At-Hook Motif Nuclear Localised Protein 18 as a Novel Modulator of Root System Architecture

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of AHL18 as a Gene Involved in Root Development and Analysis of its Expression
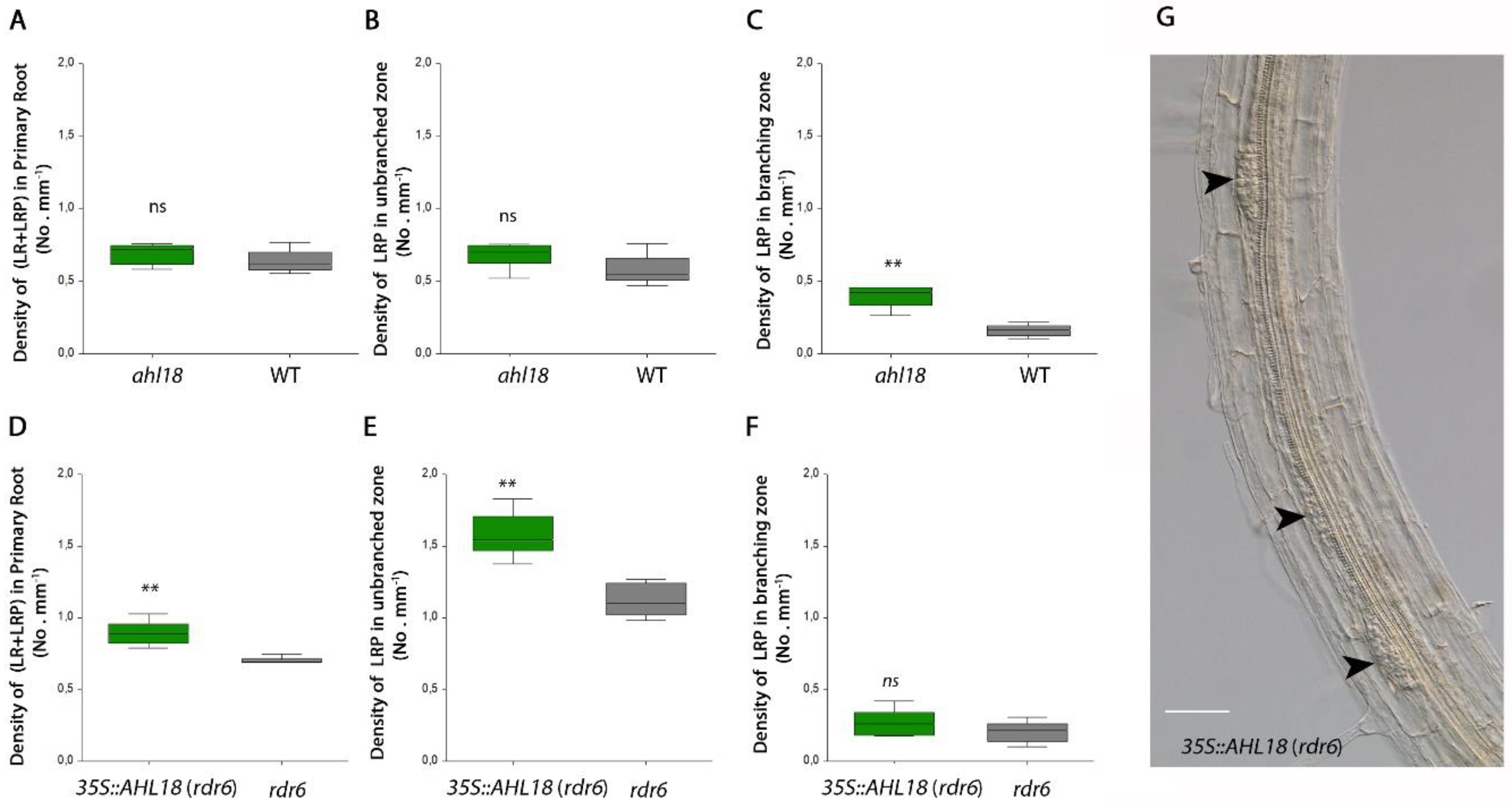
2.2. Modulation of AHL18 Expression Affects Root System Architecture
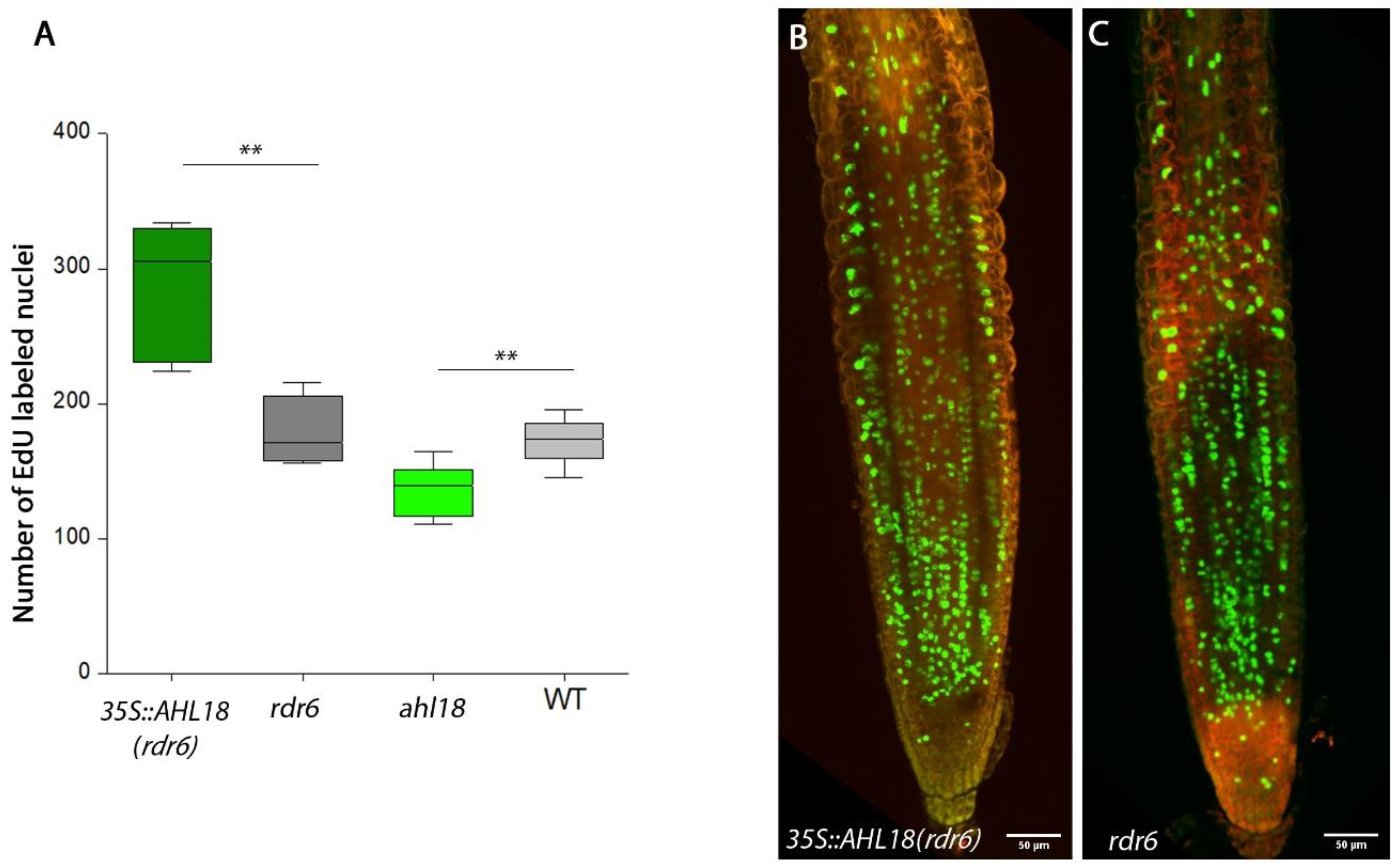
2.3. AHL18 Is Required for the Root Apical Meristem Activity
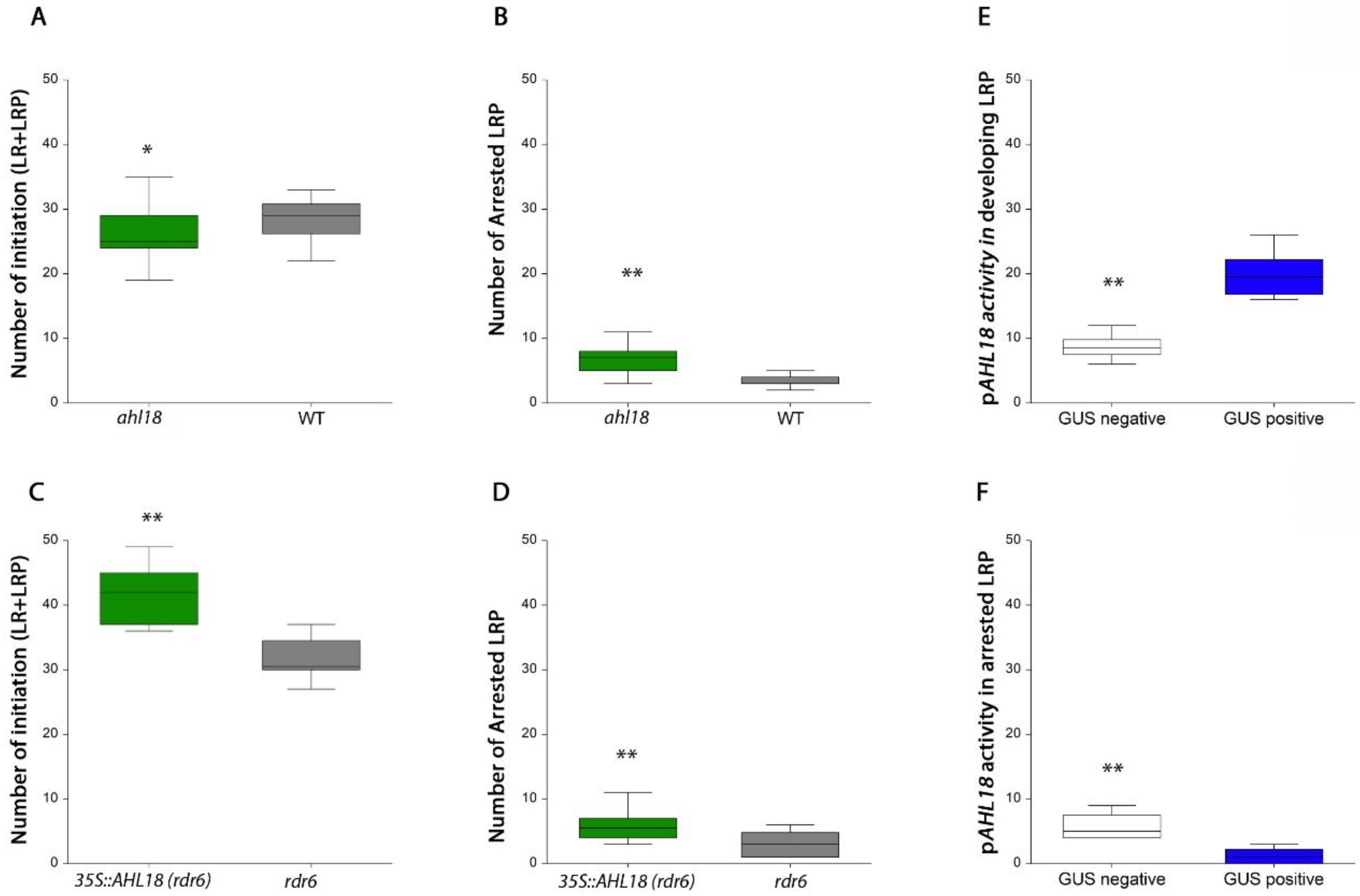
2.4. Role of AHL18 in Later Development of Lateral Root Primordia
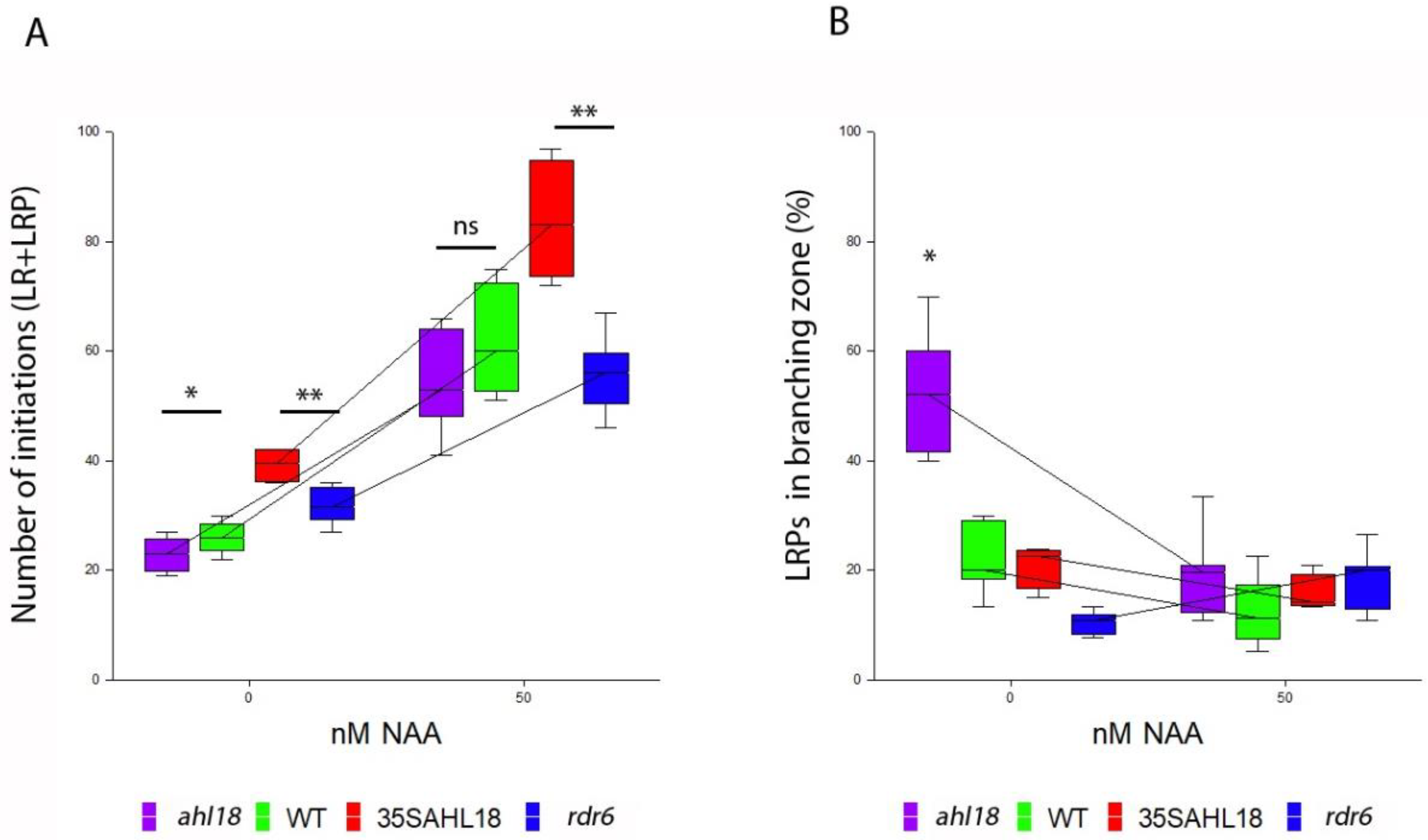
2.5. AHL18 Related Phenotypes Are not Directly Affected by Auxin
3. Discussion
3.1. Role of AHLs in Regulation of Lateral Root Development.
3.2. Root Apical Meristem and AHL18
3.3. LR Initiation and Development
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Molecular Cloning and Transgenic Lines
4.3. qPCR
4.4. Sample Processing and Microscopy
4.5. EdU Labelling
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PR | Primary root |
| LR | Lateral root |
| LRP | Lateral root primordia |
| AHL | AT-hook motif nuclear localized |
| PPC | Plants and prokaryote conserved domain |
| WT | Wild-type |
| RAM | Root apical meristem |
| EdU | 5-ethynyl-2-deoxyuridine |
References
- Malamy, J.E.; Benfey, P.N. Organization and cell differentiation in lateral roots of Arabidopsis thaliana. Development 1997, 124, 33–44. [Google Scholar]
- Torres-Martínez, H.H.; Rodríguez-Alonso, G.; Shishkova, S.; Dubrovsky, J.G. Lateral Root Primordium Morphogenesis in Angiosperms. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Banda, J.; Bellande, K.; von Wangenheim, D.; Goh, T.; Guyomarc’h, S.; Laplaze, L.; Bennett, M.J. Lateral Root Formation in Arabidopsis: A Well-Ordered LRexit. Trends Plant Sci. 2019, 24, 826–839. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Alfonso, Y.; Faulkner, C.; Pendle, A.; Miyashima, S.; Helariutta, Y.; Maule, A. Symplastic Intercellular Connectivity Regulates Lateral Root Patterning. Dev. Cell 2013, 26, 136–147. [Google Scholar] [CrossRef] [Green Version]
- Vermeer, J.E.M.; von Wangenheim, D.; Barberon, M.; Lee, Y.; Stelzer, E.H.K.; Maizel, A.; Geldner, N. A Spatial Accommodation by Neighboring Cells Is Required for Organ Initiation in Arabidopsis. Science 2014, 343, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Marhavý, P.; Montesinos, J.C.; Abuzeineh, A.; Van Damme, D.; Vermeer, J.E.M.; Duclercq, J.; Rakusová, H.; Nováková, P.; Friml, J.; Geldner, N.; et al. Targeted cell elimination reveals an auxin-guided biphasic mode of lateral root initiation. Genes Dev. 2016, 30, 471–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benkova, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertova, D.; Jurgens, G.; Friml, J. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 2003, 115, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Dubrovsky, J.G.; Sauer, M.; Napsucialy-Mendivil, S.; Ivanchenko, M.G.; Friml, J.; Shishkova, S.; Celenza, J.; Benková, E. Auxin acts as a local morphogenetic trigger to specify lateral root founder cells. Proc. Natl. Acad. Sci. USA 2008, 105, 8790–8794. [Google Scholar] [CrossRef] [Green Version]
- Péret, B.; De Rybel, B.; Casimiro, I.; Benková, E.; Swarup, R.; Laplaze, L.; Beeckman, T.; Bennett, M.J. Arabidopsis lateral root development: An emerging story. Trends Plant Sci. 2009, 14, 399–408. [Google Scholar] [CrossRef]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar] [CrossRef] [Green Version]
- Motte, H.; Vanneste, S.; Beeckman, T. Molecular and Environmental Regulation of Root Development. Annu. Rev. Plant Biol. 2019, 465–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavenus, J.; Goh, T.; Guyomarc’H, S.; Hill, K.; Lucas, M.; Voß, U.; Kenobi, K.; Wilson, M.H.; Farcot, E.; Hagen, G.; et al. Inference of the arabidopsis lateral root gene regulatory network suggests a bifurcation mechanism that defines primordia flanking and central zones. Plant Cell 2015, 27, 1368–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Favero, D.S.; Peng, H.; Neff, M.M. Arabidopsis thaliana AHL family modulates hypocotyl growth redundantly by interacting with each other via the PPC/DUF296 domain. Proc. Natl. Acad. Sci. USA 2013, 110, E4688–E4697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, S.; Matsunaga, S.; Yonemura, M.; Uchiyama, S.; Azuma, T.; Fukui, K. Identification of a novel plant MAR DNA binding protein localized on chromosomal surfaces. Plant Mol. Biol. 2004, 56, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.M.; Bhaskara, G.B.; Wen, T.N.; Lin, W.D.; Nguyen, T.T.; Chong, G.L.; Verslues, P.E. Phosphoproteomics of Arabidopsis Highly ABA-Induced1 identifies AT-Hook-Like10 phosphorylation required for stress growth regulation. Proc. Natl. Acad. Sci. USA 2019, 116, 2354–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, K.-H.; Yu, H.; Ito, T. AGAMOUS Controls GIANT KILLER, a Multifunctional Chromatin Modifier in Reproductive Organ Patterning and Differentiation. PLoS Biol. 2009, 7, e1000251. [Google Scholar] [CrossRef] [Green Version]
- Lim, P.O.; Kim, Y.; Breeze, E.; Koo, J.C.; Woo, H.R.; Ryu, J.S.; Park, D.H.; Beynon, J.; Tabrett, A.; Buchanan-Wollaston, V.; et al. Overexpression of a chromatin architecture-controlling AT-hook protein extends leaf longevity and increases the post-harvest storage life of plants. Plant J. 2007, 52, 1140–1153. [Google Scholar] [CrossRef]
- Yun, J.; Kim, Y.S.; Jung, J.H.; Seo, P.J.; Park, C.M. The AT-hook motif-containing protein AHL22 regulates flowering initiation by modifying FLOWERING LOCUS T chromatin in Arabidopsis. J. Biol. Chem. 2012, 287, 15307–15316. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Zou, Y.; Feng, N. Overexpression of AHL20 negatively regulates defenses in arabidopsis. J. Integr. Plant Biol. 2010, 52, 801–808. [Google Scholar] [CrossRef]
- Yadeta, K.A.; Hanemian, M.; Smit, P.; Hiemstra, J.A.; Pereira, A.; Marco, Y.; Thomma, B.P.H.J. DNA-Binding Protein AHL19 Mediates Verticillium Wilt Resistance. Mol. Plant Microbe Interact. 2011, 24, 1582–1591. [Google Scholar] [CrossRef] [Green Version]
- Howden, A.J.M.; Stam, R.; Martinez Heredia, V.; Motion, G.B.; ten Have, S.; Hodge, K.; Marques Monteiro Amaro, T.M.; Huitema, E. Quantitative analysis of the tomato nuclear proteome during Phytophthora capsici infection unveils regulators of immunity. New Phytol. 2017, 215, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Street, I.H.; Shah, P.K.; Smith, A.M.; Avery, N.; Neff, M.M. The AT-hook-containing proteins SOB3/AHL29 and ESC/AHL27 are negative modulators of hypocotyl growth in Arabidopsis. Plant J. 2008, 54, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, A.; Furumoto, T.; Ishida, S.; Takahashi, Y. AGF1, an AT-hook protein, is necessary for the negative feedback of AtGA3ox1 encoding GA 3-oxidase. Plant Physiol. 2007, 143, 1152–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Seo, P.J. Coordination of matrix attachment and ATP-dependent chromatin remodeling regulate auxin biosynthesis and Arabidopsis hypocotyl elongation. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Wang, X.; Lee, J.-Y.; Lee, J.-Y. Cell-to-Cell Movement of Two Interacting AT-Hook Factors in Arabidopsis Root Vascular Tissue Patterning. Plant Cell 2013, 25, 187–201. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Chen, F.; Yu, X.; Lin, C.; Fu, Y.-F. Over-expression of an AT-hook gene, AHL22, delays flowering and inhibits the elongation of the hypocotyl in Arabidopsis thaliana. Plant Mol. Biol. 2009, 71, 39–50. [Google Scholar] [CrossRef]
- Sundaresan, V.; Springer, P.; Volpe, T.; Haward, S.; Jones, J.D.G.; Dean, C.; Ma, H.; Martienssen, R. Patterns of gene action in plant development revealed by enhancer trap and gene trap transposable elements. Genes Dev. 1995, 9, 1797–1810. [Google Scholar] [CrossRef] [Green Version]
- Vielle-Calzada, J.P.; Baskar, R.; Grossniklaus, U. Delayed activation of the paternal genome during seed development. Nature 2000, 404, 91–94. [Google Scholar] [CrossRef]
- Liu, Y.-G.; Chen, Y.; Zhang, Q. Amplification of Genomic Sequences Flanking T-DNA Insertions by Thermal Asymmetric Interlaced Polymerase Chain Reaction. Methods Mol. Biol. 2005, 286, 341–348. [Google Scholar]
- Deuschle, K.; Chaudhuri, B.; Okumoto, S.; Lager, I.; Lalonde, S.; Frommer, W.B. Rapid Metabolism of Glucose Detected with FRET Glucose Nanosensors in Epidermal Cells and Intact Roots of Arabidopsis RNA-Silencing Mutants. Plant Cell Online 2006, 18, 2314–2325. [Google Scholar] [CrossRef] [Green Version]
- Dubrovsky, J.G.; Fordeb, B.G. Quantitative analysis of lateral root development: Pitfalls and how to avoid them. Plant Cell 2012, 24, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, V.B.; Dubrovsky, J.G. Estimation of the cell-cycle duration in the root apical meristem: A model of linkage between cell-cycle duration, rate of cell production, and rate of root growth. Int. J. Plant Sci. 1997, 158, 757–763. [Google Scholar] [CrossRef]
- Napsucialy-Mendivil, S.; Dubrovsky, J.G. Genetic and Phenotypic Analysis of Lateral Root Development in Arabidopsis thaliana. Methods Mol. Biol. 2018, 1761, 47–75. [Google Scholar] [PubMed]
- Jiang, H.; Moreno-Romero, J.; Santos-González, J.; De Jaeger, G.; Gevaert, K.; Van De Slijke, E.; Köhler, C. Ectopic application of the repressive histone modification H3K9me2 establishes post-zygotic reproductive isolation in Arabidopsis Thaliana. Genes Dev. 2017, 31, 1272–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.; Liu, X.; Shi, Z.; Li, D.; Zhu, L. An AT-hook protein DEPRESSED PALEA1 physically interacts with the TCP Family transcription factor RETARDED PALEA1 in rice. Biochem. Biophys. Res. Commun. 2018, 495, 487–492. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, Z.; Liu, Y.; Kong, D.; Li, T.; Yu, S.; Mei, H.; Xu, X.; Liu, H.; Chen, L.; et al. A novel gene OsAHL1 improves both drought avoidance and drought tolerance in rice. Sci. Rep. 2016, 6, 30264. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, J.; Mosquera, J.; Couceiro, J.R.; Vázquez, M.E.; Mascareñas, J.L. The AT-Hook motif as a versatile minor groove anchor for promoting DNA binding of transcription factor fragments. Chem. Sci. 2015, 6, 4767–4771. [Google Scholar] [CrossRef] [Green Version]
- Greb, T.; Lohmann, J.U. Plant Stem Cells. Curr. Biol. 2016, 26, R816–R821. [Google Scholar] [CrossRef] [Green Version]
- Gruel, J.; Landrein, B.; Tarr, P.; Schuster, C.; Refahi, Y.; Sampathkumar, A.; Hamant, O.; Meyerowitz, E.M.; Jönsson, H. An epidermis-driven mechanism positions and scales stem cell niches in plants. Sci. Adv. 2016, 2, 22–24. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Liu, Y.; Zhou, W.; Zhu, Y.; Dong, A.; Shen, W.H. Histone chaperones play crucial roles in maintenance of stem cell niche during plant root development. Plant J. 2018, 95, 86–100. [Google Scholar] [CrossRef]
- Mähönen, A.P.; Bishopp, A.; Higuchi, M.; Nieminen, K.M.; Kinoshita, K.; Törmäkangas, K.; Ikeda, Y.; Oka, A.; Kakimoto, T.; Helariutta, Y. Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 2006, 311, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Harashima, H.; Schnittger, A. The integration of cell division, growth and differentiation. Curr. Opin. Plant Biol. 2010, 13, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Mislata, A.; Schiessl, K.; Sablowski, R. Active Control of Cell Size Generates Spatial Detail during Plant Organogenesis. Curr. Biol. 2015, 25, 2991–2996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Q.; Reed, J.W. Control of auxin-regulated root development by the Arabidopsis thaliana SHY2/IAA3 gene. Development 1999, 126, 711–721. [Google Scholar]
- Moubayidin, L.; Perilli, S.; Dello Ioio, R.; Di Mambro, R.; Costantino, P.; Sabatini, S. The rate of cell differentiation controls the arabidopsis root meristem growth phase. Curr. Biol. 2010, 20, 1138–1143. [Google Scholar] [CrossRef]
- Ivanov, V.B.; Dubrovsky, J.G. Longitudinal zonation pattern in plant roots: Conflicts and solutions. Trends Plant Sci. 2013, 18, 237–243. [Google Scholar] [CrossRef]
- Di Mambro, R.; De Ruvo, M.; Pacifici, E.; Salvi, E.; Sozzani, R.; Benfey, P.N.; Busch, W.; Novak, O.; Ljung, K.; Di Paola, L.; et al. Auxin minimum triggers the developmental switch from cell division to cell differentiation in the Arabidopsis root. Proc. Natl. Acad. Sci. USA 2017, 114, E7641–E7649. [Google Scholar] [CrossRef] [Green Version]
- Di Mambro, R.; Svolacchia, N.; Dello Ioio, R.; Pierdonati, E.; Salvi, E.; Pedrazzini, E.; Vitale, A.; Perilli, S.; Sozzani, R.; Benfey, P.N.; et al. The Lateral Root Cap Acts as an Auxin Sink that Controls Meristem Size. Curr. Biol. 2019, 29, 1199–1205. [Google Scholar] [CrossRef] [Green Version]
- Napsucialy-Mendivil, S.; Alvarez-Venegas, R.; Shishkova, S.; Dubrovsky, J.G. Arabidopsis homolog of trithorax1 (ATX1) is required for cell production, patterning, and morphogenesis in root development. J. Exp. Bot. 2014, 65, 6373–6384. [Google Scholar] [CrossRef] [Green Version]
- Aida, M.; Beis, D.; Heidstra, R.; Willemsen, V.; Blilou, I.; Galinha, C.; Nussaume, L.; Noh, Y.-S.; Amasino, R.; Scheres, B. The PLETHORA Genes Mediate Patterning of the Arabidopsis Root Stem Cell Niche. Cell 2004, 119, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional Regulation of ROS Controls Transition from Proliferation to Differentiation in the Root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhang, J.; He, J.; Qin, Y.; Hua, D.; Duan, Y.; Chen, Z.; Gong, Z. ABA-Mediated ROS in Mitochondria Regulate Root Meristem Activity by Controlling PLETHORA Expression in Arabidopsis. PLoS Genet. 2014, 10, e1004791. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Tian, H.; Yue, K.; Liu, J.; Zhang, B.; Li, X.; Ding, Z. A P-Loop NTPase Regulates Quiescent Center Cell Division and Distal Stem Cell Identity through the Regulation of ROS Homeostasis in Arabidopsis Root. PLOS Genet. 2016, 12, e1006175. [Google Scholar] [CrossRef]
- Péret, B.; Larrieu, A.; Bennett, M.J. Lateral root emergence: A difficult birth. J. Exp. Bot. 2009, 60, 3637–3643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubrovsky, J.G.; Doerner, P.W.; Colón-Carmona, A.; Rost, T.L. Pericycle cell proliferation and lateral root initiation in Arabidopsis. Plant Physiol. 2000, 124, 1648–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parizot, B.; Laplaze, L.; Ricaud, L.; Boucheron-Dubuisson, E.; Bayle, V.; Bonke, M.; De Smet, I.; Poethig, S.R.; Helariutta, Y.; Haseloff, J.; et al. Diarch symmetry of the vascular bundle in arabidopsis root encompasses the pericycle and is reflected in distich lateral root initiation. Plant Physiol. 2008, 146, 140–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beeckman, T.; Burssens, S.; Inze, D. The peri-cell-cycle in Arabidopsis. J. Exp. Bot. 2001, 52, 403–411. [Google Scholar]
- Nieuwland, J.; Maughan, S.; Dewitte, W.; Scofield, S.; Sanz, L.; Murray, J.A.H. The D-type cyclin CYCD4;1 modulates lateral root density in Arabidopsis by affecting the basal meristem region. Proc. Natl. Acad. Sci. USA 2009, 106, 22528–22533. [Google Scholar] [CrossRef] [Green Version]
- Lloret, P.G.; Casero, P.J.; Pulgarín, A.; Navascués, J. The Behaviour of Two Cell Populations in the Pericycle of Allium cepa, Pisum sativum, and Daucus carota During Early Lateral Root Development. Ann. Bot. 1989, 63, 465–475. [Google Scholar] [CrossRef]
- Lavrekha, V.V.; Pasternak, T.; Ivanov, V.B.; Palme, K.; Mironova, V.V. 3D analysis of mitosis distribution highlights the longitudinal zonation and diarch symmetry in proliferation activity of the Arabidopsis thaliana root meristem. Plant J. 2017, 92, 834–845. [Google Scholar] [CrossRef] [Green Version]
- Celenza, J.L.; Grisafi, P.L.; Fink, G.R. A pathway for lateral root formation in Arabidopsis thaliana. Genes Dev. 1995, 9, 2131–2142. [Google Scholar] [CrossRef] [Green Version]
- DiDonato, R.J.; Arbuckle, E.; Buker, S.; Sheets, J.; Tobar, J.; Totong, R.; Grisafi, P.; Fink, G.R.; Celenza, J.L. Arabidopsis ALF4 encodes a nuclear-localized protein required for lateral root formation. Plant J. 2004, 37, 340–353. [Google Scholar] [CrossRef]
- Alarcón, M.V.; Salguero, J.; Lloret, P.G. Auxin modulated initiation of lateral roots is linked to pericycle cell length in Maize. Front. Plant Sci. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, T.; Toyokura, K.; Wells, D.M.; Swarup, K.; Yamamoto, M.; Mimura, T.; Weijers, D.; Fukaki, H.; Laplaze, L.; Bennett, M.J.; et al. Quiescent center initiation in the Arabidopsis lateral root primordia is dependent on the SCARECROW transcription factor. Development 2016, 143, 3363–3371. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Scheres, B. Lateral root formation and the multiple roles of auxin. J. Exp. Bot. 2018, 69, 155–167. [Google Scholar] [CrossRef]
- Laskowski, M.J.; Williams, M.E.; Nusbaum, H.C.; Sussex, I.M. Formation of lateral root meristems is a two-stage process. Development 1995, 121, 3303–3310. [Google Scholar]
- Tian, H.; De Smet, I.; Ding, Z. Shaping a root system: Regulating lateral versus primary root growth. Trends Plant Sci. 2014, 19, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Shimotohno, A.; Heidstra, R.; Blilou, I.; Scheres, B. Root stem cell niche organizer specification by molecular convergence of PLETHORA and SCARECROW transcription factor modules. Genes Dev. 2018, 32, 1085–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Smet, I.; White, P.J.; Glyn Bengough, A.; Dupuy, L.; Parizot, B.; Casimiro, I.; Heidstra, R.; Laskowski, M.; Lepetit, M.; Hochholdinger, F.; et al. Analyzing lateral root development: How to move forward. Plant Cell 2012, 24, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Swarup, K.; Benková, E.; Swarup, R.; Casimiro, I.; Péret, B.; Yang, Y.; Parry, G.; Nielsen, E.; De Smet, I.; Vanneste, S.; et al. The auxin influx carrier LAX3 promotes lateral root emergence. Nat. Cell Biol. 2008, 10, 946–954. [Google Scholar] [CrossRef]
- Dubrovsky, J.G.; Gambetta, G.A.; Hernández-Barrera, A.; Shiskova, S.; Gonzáles, I. Lateral Root Initiation in Arabidopsis: Developmental Window, Spatial Patterning, Density and Predictability. Ann. Bot. 2006, 97, 903–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobet, G.; Pagès, L.; Draye, X. A Novel Image-Analysis Toolbox Enabling Quantitative Analysis of Root System Architecture. Plant Physiol. 2011, 157, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, M.G.; Thompson, W.F. Nucleic Acids Research Rapid Isolation of High Molecular Weight Plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.L.; Moorman, A.F.M. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Soukup, A. Selected Simple Methods of Plant Cell Wall Histochemistry and Staining for Light Microscopy; Humana Press: Totowa, NJ, USA, 2014; pp. 25–40. [Google Scholar]
- Kotogány, E.; Dudits, D.; Horváth, G.V.; Ayaydin, F. A rapid and robust assay for detection of S-phase cell cycle progression in plant cells and tissues by using ethynyl deoxyuridine. Plant Methods 2010, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NCSS. NCSS 9 Statistical Software No Title 2013; NCSS LLC: Kaysville, UT, USA, 2013.









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Širl, M.; Šnajdrová, T.; Gutiérrez-Alanís, D.; Dubrovsky, J.G.; Vielle-Calzada, J.P.; Kulich, I.; Soukup, A. At-Hook Motif Nuclear Localised Protein 18 as a Novel Modulator of Root System Architecture. Int. J. Mol. Sci. 2020, 21, 1886. https://doi.org/10.3390/ijms21051886
Širl M, Šnajdrová T, Gutiérrez-Alanís D, Dubrovsky JG, Vielle-Calzada JP, Kulich I, Soukup A. At-Hook Motif Nuclear Localised Protein 18 as a Novel Modulator of Root System Architecture. International Journal of Molecular Sciences. 2020; 21(5):1886. https://doi.org/10.3390/ijms21051886
Chicago/Turabian StyleŠirl, Marek, Tereza Šnajdrová, Dolores Gutiérrez-Alanís, Joseph G. Dubrovsky, Jean Phillipe Vielle-Calzada, Ivan Kulich, and Aleš Soukup. 2020. "At-Hook Motif Nuclear Localised Protein 18 as a Novel Modulator of Root System Architecture" International Journal of Molecular Sciences 21, no. 5: 1886. https://doi.org/10.3390/ijms21051886
APA StyleŠirl, M., Šnajdrová, T., Gutiérrez-Alanís, D., Dubrovsky, J. G., Vielle-Calzada, J. P., Kulich, I., & Soukup, A. (2020). At-Hook Motif Nuclear Localised Protein 18 as a Novel Modulator of Root System Architecture. International Journal of Molecular Sciences, 21(5), 1886. https://doi.org/10.3390/ijms21051886