Role of 5-HT1A Receptor in the Anxiolytic-Relaxant Effects of Bergamot Essential Oil in Rodent

 ,
,  , , ,
, , ,  and
and 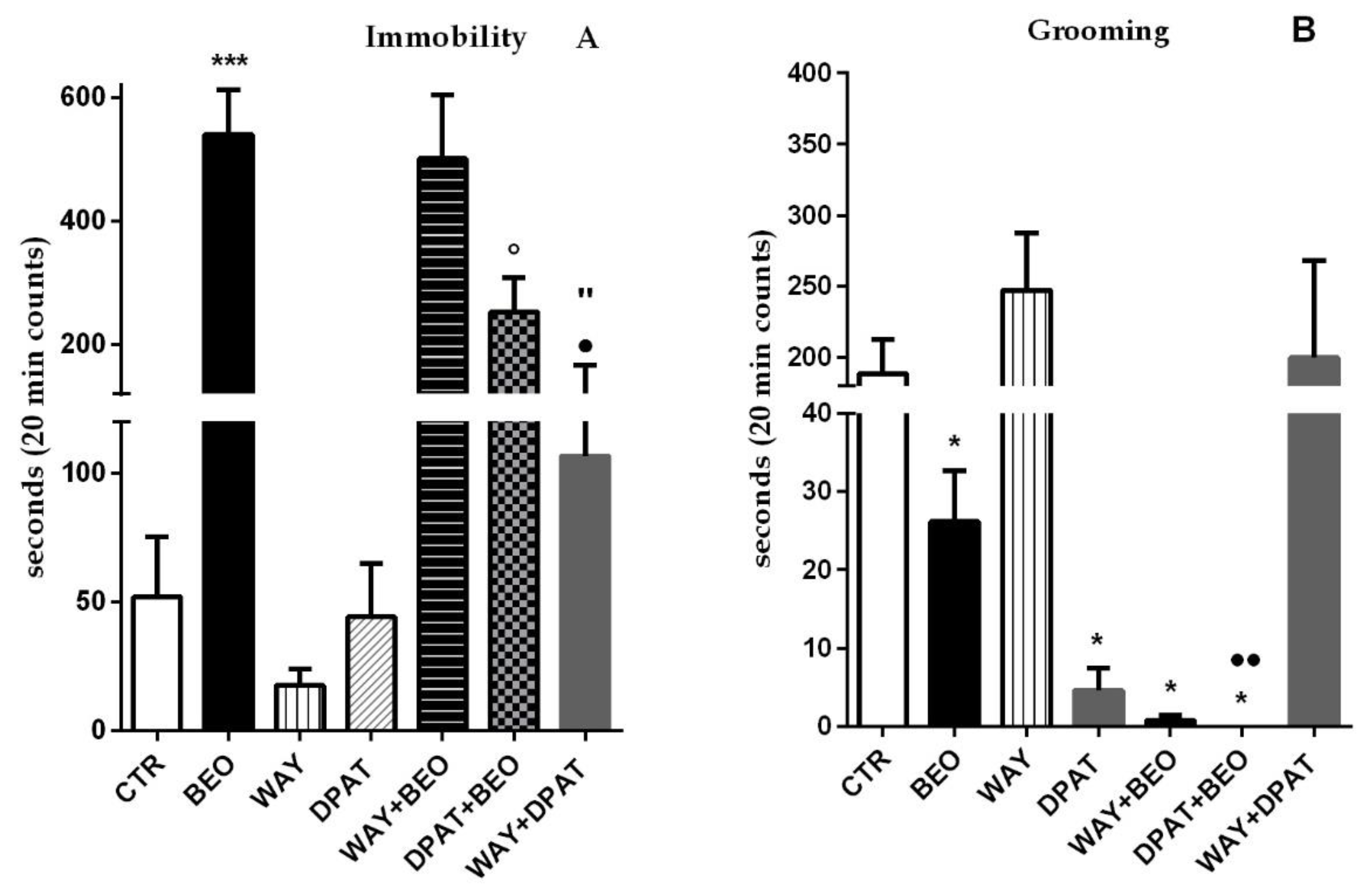{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
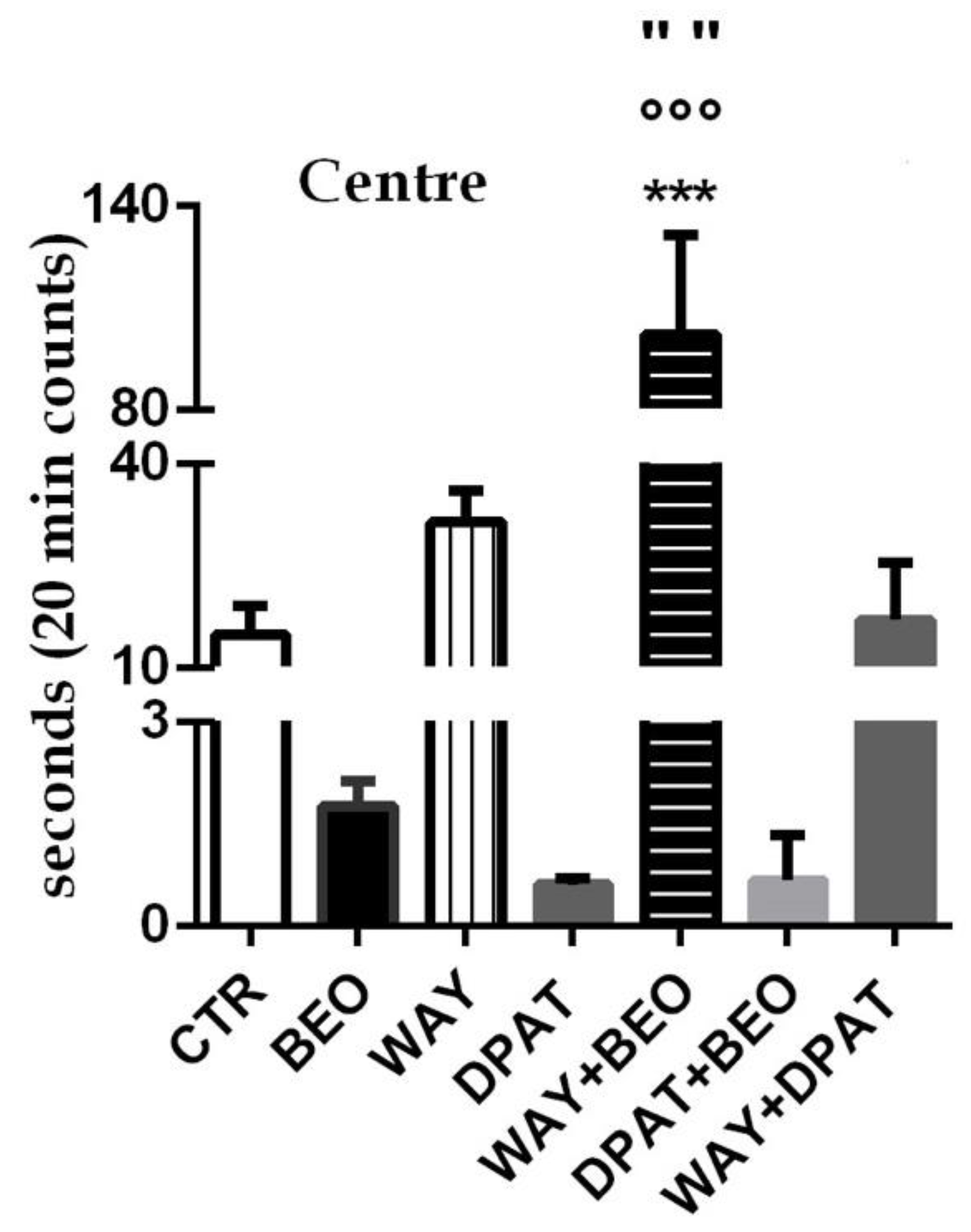
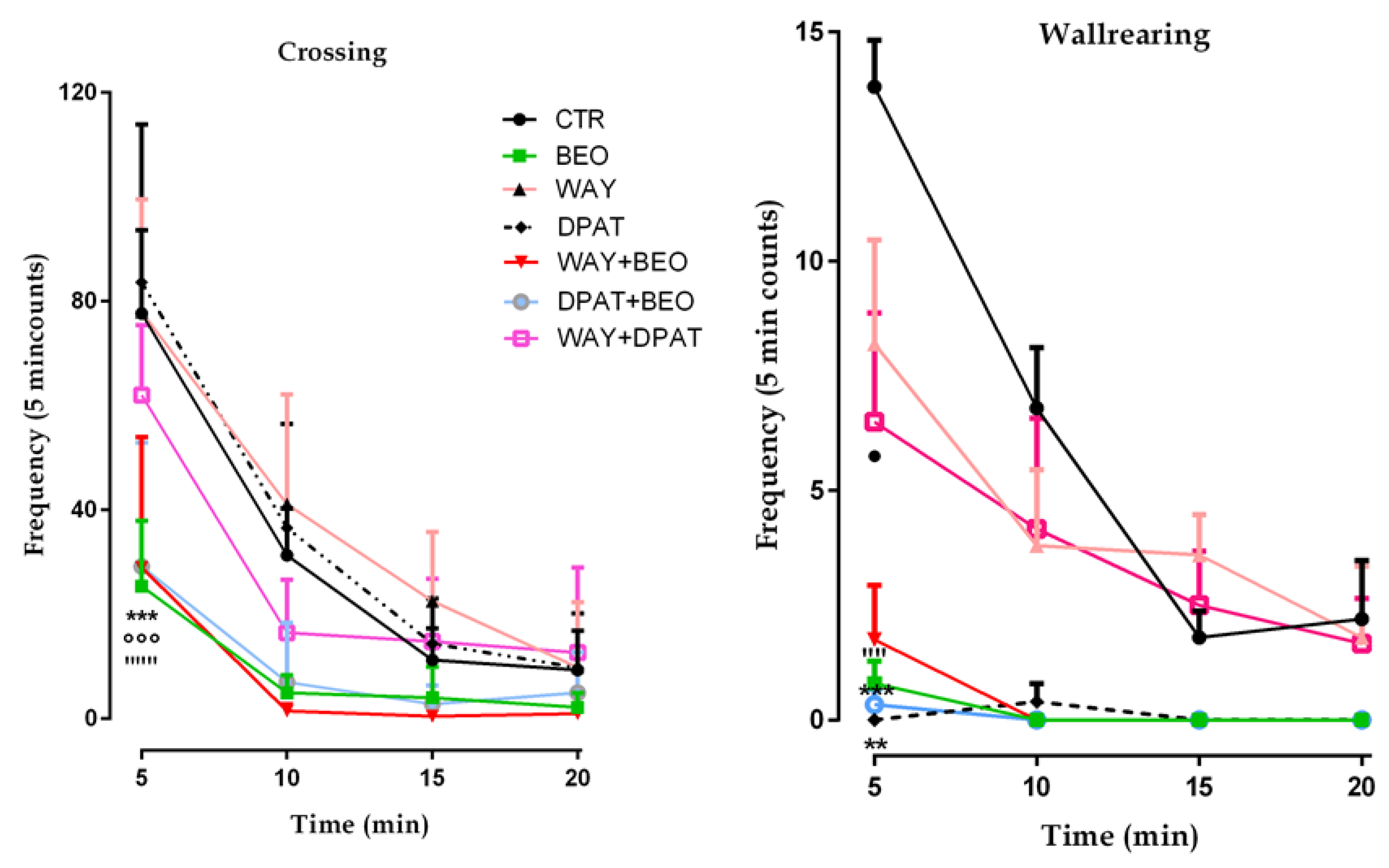
2.1. Open Field Test
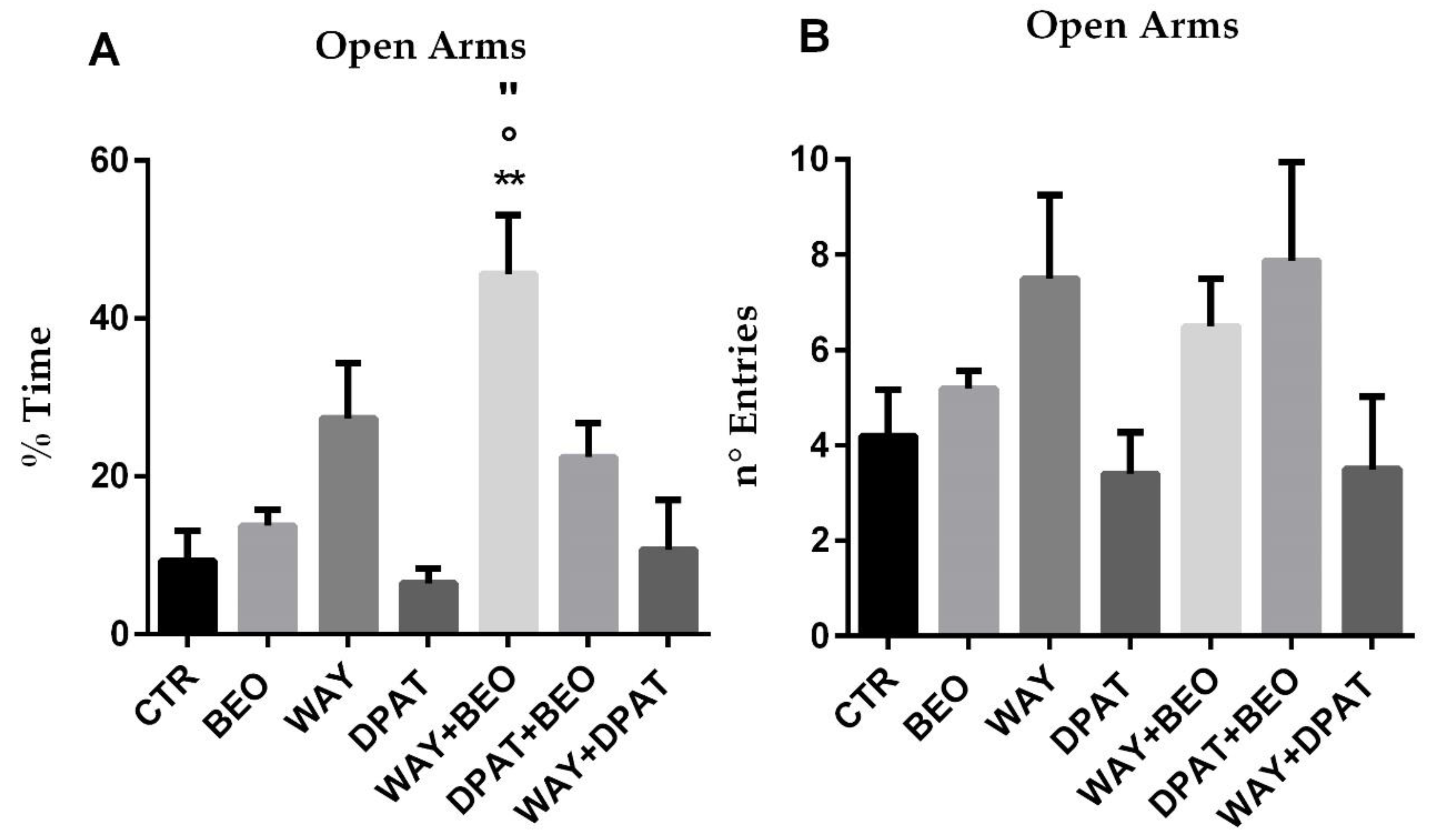
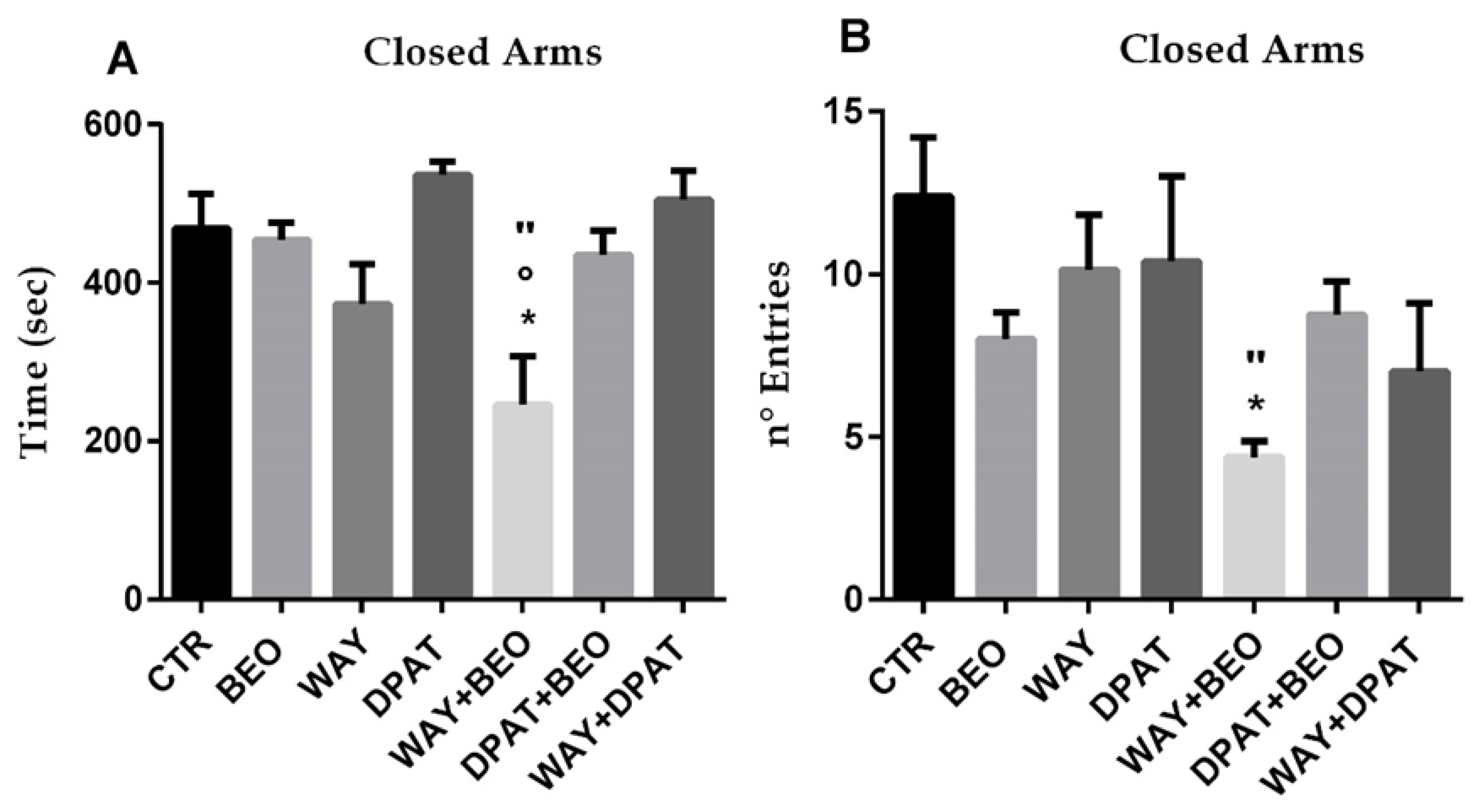
2.2. Elevated Plus Maze Test
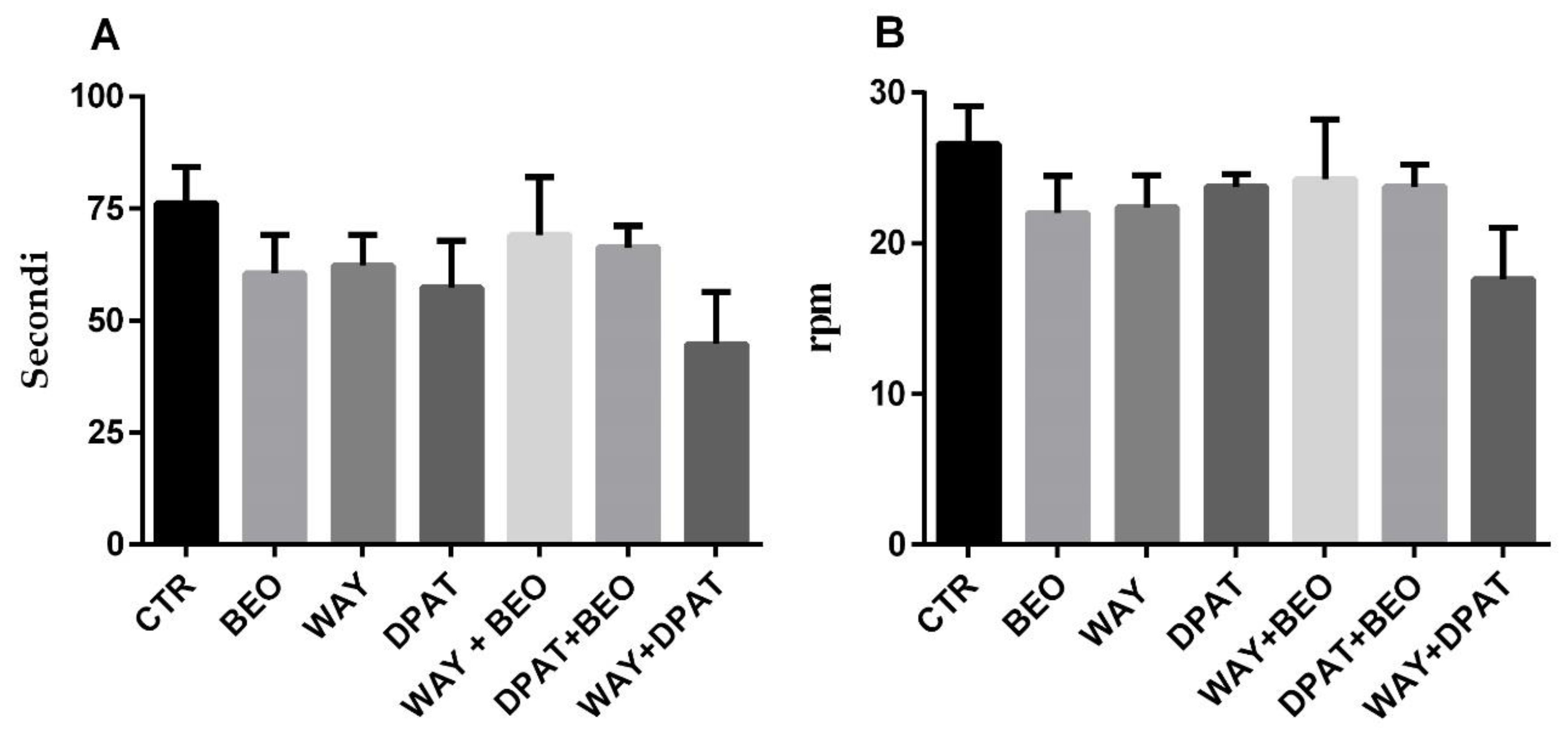
2.3. Rotarod Test
2.4. Statistics
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Analysis

4.3. Open Field Test
4.4. Elevated Plus Maze Test
4.5. Rotarod Test
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| (±)8-OH-DPAT | 8-hydroxy-2-(di-n-propylamino) tetralin |
| 5-HT | Serotonin |
| BEO | Bergamot essential oil |
| CTR | Control |
| EPM | Elevated Plus Maze |
| mGluR8 | Metabotropic glutamate receptors8 |
| OF | Open Field |
| WAY-100635 | N-{2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl}-N-(2-pyridinyl)cyclohexanecarboxamide trihydrochloride |
References
- Ni, C.H.; Hou, W.H.; Kao, C.C.; Chang, M.L.; Yu, L.F.; Wu, C.C.; Chen, C. The anxiolytic effect of aromatherapy on patients awaiting ambulatory surgery, a randomized controlled trial. Evid. Based Complement. Alternat. Med. 2013, 2013, 927419. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, E.; Kuchta, K.; Kimura, M.; Rauwald, H.W.; Kamei, T.; Imanishi, J. Effects of bergamot (Citrus bergamia (Risso) Wright & Arn.) essential oil aromatherapy on mood states, parasympathetic nervous system activity, and salivary cortisol levels in 41 healthy females. Complement. Med. Res. 2015, 22, 43–49. [Google Scholar] [CrossRef]
- Ndao, D.H.; Ladas, E.J.; Cheng, B.; Sands, S.A.; Snyder, K.T.; Garvin, J.H., Jr.; Kelly, K.M. Inhalation aromatherapy in children and adolescents undergoing stem cell infusion: Results of a placebo-controlled double-blind trial. Psychooncology 2012, 21, 247–254. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, B.; Newcomb, P.J. Trial of Essential Oils to Improve Sleep for Patients in Cardiac Rehabilitation. Altern. Compl. Med. 2019, 25, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.W.; Kim, Y.K.; Jeon, H.J. Comorbid Anxiety and Depression: Clinical and Conceptual Consideration and Transdiagnostic Treatment. Adv. Exp. Med. Biol. 2020, 1191, 219–235. [Google Scholar] [CrossRef]
- Kern, N.; Sheldrick, A.J.; Schmidt, F.M.; Minkwitz, J. Neurobiology of depression and novel antidepressant drug targets. Curr. Pharm. Des. 2012, 18, 5791–5801. [Google Scholar] [CrossRef]
- Markou, A.; Cryan, J.F. Stress, anxiety and depression: Toward new treatment strategies. Neuropharmacology 2012, 62, 1–2. [Google Scholar] [CrossRef]
- Bagetta, G.; Morrone, L.A.; Rombolà, L.; Amantea, D.; Russo, R.; Berliocchi, L.; Sakurad, S.; Sakurada, T.; Rotiroti, D.; Corasaniti, M.T. Neuropharmacology of the essential oil of bergamot. Fitoterapia 2010, 81, 453–461. [Google Scholar] [CrossRef]
- Scuteri, D.; Morrone, L.A.; Rombolà, L.; Avato, P.R.; Bilia, A.R.; Corasaniti, M.T.; Sakurada, S.; Sakurada, T.; Bagetta, G. Aromatherapy and Aromatic Plants for the Treatment of Behavioural and Psychological Symptoms of Dementia in Patients with Alzheimer’s Disease: Clinical Evidence and Possible Mechanisms. Evid. Based Complement. Alternat. Med. 2017, 2017, 9416305. [Google Scholar] [CrossRef] [Green Version]
- Scuteri, D.; Rombolà, L.; Tridico, L.; Mizoguchi, H.; Watanabe, C.; Sakurada, T.; Sakurada, S.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Neuropharmacological Properties of the Essential Oil of Bergamot for the Clinical Management of Pain-Related BPSDs. Curr. Med. Chem. 2019, 26, 3764–3774. [Google Scholar] [CrossRef]
- Donelli, D.; Antonelli, M.; Bellinazzi, C.; Gensini, G.F.; Firenzuoli, F. Effects of lavender on anxiety: A systematic review and meta-analysis. Phytomedicine 2019, 65, 153099. [Google Scholar] [CrossRef] [PubMed]
- Sakurada, T.; Mizoguchi, H.; Kuwahata, H.; Katsuyama, S.; Komatsu, T.; Morrone, L.A.; Corasaniti, M.T.; Bagetta, G.; Sakurada, S. Intraplantar injection of bergamot essential oil induces peripheral antinociception mediated by opioid mechanism. Pharmacol. Biochem. Behav. 2011, 97, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Kuwahata, H.; Komatsu, T.; Katsuyama, S.; Corasaniti, M.T.; Bagetta, G.; Sakurada, S.; Sakurada, T.; Takahama, K. Peripherally injected linalool and bergamot essential oil attenuate mechanical allodynia via inhibiting spinal ERK phosphorylation. Pharmacol. Biochem. Behav. 2013, 103, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Katsuyama, S.; Otowa, A.; Kamio, S.; Sato, K.; Yagi, T.; Kishikawa, Y.; Komatsu, T.; Bagetta, G.; Sakurada, T.; Nakamura, H. Effect of plantar subcutaneous administration of bergamot essential oil and linalool on formalin-induced nociceptive behavior in mice. Biomed. Res. 2015, 36, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rombolà, L.; Amantea, D.; Russo, R.; Adornetto, A.; Berliocchi, L.; Tridico, L.; Corasaniti, M.T.; Sakurada, S.; Sakurada, T.; Bagetta, G.; et al. Rational Basis for the Use of Bergamot Essential Oil in Complementary Medicine to Treat Chronic Pain. Mini Rev. Med. Chem. 2016, 16, 721–728. [Google Scholar] [CrossRef]
- Scuteri, D.; Crudo, M.; Rombolà, L.; Watanabe, C.; Mizoguchi, H.; Sakurada, S.; Sakurada, T.; Greco, R.; Corasaniti, M.T.; Morrone, L.A.; et al. Antinociceptive effect of inhalation of the essential oil of bergamot in mice. Fitoterapia 2018, 129, 20–24. [Google Scholar] [CrossRef]
- Rombolà, L.; Tridico, L.; Scuteri, D.; Sakurada, T.; Sakurada, S.; Mizoguchi, H.; Avato, P.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Bergamot Essential Oil Attenuates Anxiety-Like Behaviour in Rats. Molecules 2017, 22, 614. [Google Scholar] [CrossRef]
- Rombolà, L.; Scuteri, D.; Adornetto, A.; Straface, M.; Sakurada, T.; Sakurada, S.; Mizoguchi, H.; Corasaniti, M.T.; Bagetta, G.; Tonin, P.; et al. Anxiolytic-Like Effects of Bergamot Essential Oil Are Insensitive to Flumazenil in Rats. Evid. Bas. Complement. Alternat. Med. 2019, 14, 2156873. [Google Scholar] [CrossRef]
- Rombolà, L.; Corasaniti, M.T.; Rotiroti, D.; Tassorelli, C.; Sakurada, S.; Bagetta, G.; Morrone, L.A. Effects of systemic administration of the essential oil of bergamot (BEO) on gross behaviour and EEG power spectra recorded from the rat hippocampus and cerebral cortex. Funct. Neurol. 2009, 24, 107–112. [Google Scholar]
- Morrone, L.A.; Rombola, L.; Pelle, C.; Corasaniti, M.T.; Zappettini, S.; Paudice, P.; Bonanno, G.; Bagetta, G. The essential oil of bergamot enhances the levels of amino acid neurotransmitters in the hippocampus of rat: Implication of monoterpene hydrocarbons. Pharmacol. Res. 2007, 55, 255–262. [Google Scholar] [CrossRef]
- Lesch, K.P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Müller, C.R.; Hamer, D.H.; Murphy, D.L. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 1996, 274, 1527–1531. [Google Scholar] [CrossRef] [PubMed]
- Ressler, K.J.; Nemeroff, C.B. Role of serotonergic and noradrenergic systems in the pathophysiology of depression and anxiety disorders. Depress. Anxiety 2000, 12, 2–19. [Google Scholar] [CrossRef]
- Holmes, A.; Murphy, D.L.; Crawley, J.N. Abnormal behavioral phenotypes of serotonin transporter knockout mice: Parallels with human anxiety and depression. Biol. Psychiatry 2003, 54, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Komiya, M.; Takeuchi, T.; Harada, E. Lemon oil vapor causes an anti-stress effect via modulating the 5-HT and DA activities in mice. Behav. Brain Res. 2006, 172, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Chioca, L.R.; Ferro, M.M.; Baretta, I.P.; Oliveira, S.M.; Silva, C.R.; Ferreira, J.; Losso, E.M.; Andreatini, R. Anxiolytic-like effect of lavender essential oil inhalation in mice: Participation of serotonergic but not GABAA/benzodiazepine neurotransmission. J. Ethnopharmacol. 2013, 147, 412–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, C.; Cury, T.C.; Cassettari, B.O.; Takahira, R.K.; Flório, J.C.; Costa, M. Citrus aurantium L. essential oil exhibits anxiolytic-like activity mediated by 5-HT(1A)-receptors and reduces cholesterol after repeated oral treatment. BMC Complement. Altern. Med. 2013, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Tabari, A.; Moghaddam, H.A.; Maggi, F.; Benelli, G. Anxiolytic and antidepressant activities of Pelargonium roseum essential oil on Swiss albino mice: Possible involvement of serotonergic transmission. Phytother. Res. 2018, 32, 1014–1022. [Google Scholar] [CrossRef]
- López, V.; Nielsen, B.; Solas, M.; Ramírez, M.J.; Jäger, A.K. Exploring Pharmacological Mechanisms of Lavender (Lavandula angustifolia) Essential Oil on Central Nervous System Targets. Front. Pharmacol. 2017, 8, 280. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, L.; Feng, L.; Yao, L. Cananga odorata essential oil reverses the anxiety induced by 1-(3-chlorophenyl) piperazine through regulating the MAPK pathway and serotonin system in mice. J. Ethnopharmacol. 2018, 219, 23–30. [Google Scholar] [CrossRef] [PubMed]
- File, S.E.; Gonzalez, L.E.; Andrews, N. Comparative study of pre- and postsynaptic 5-HT1A receptor modulation of anxiety in two ethological animal tests. J. Neurosci. 1996, 16, 4810–4815. [Google Scholar] [CrossRef] [PubMed]
- Spiacci, A., Jr.; Pobbe, R.L.H.; Matthiesen, M.; Zangrossi, H., Jr. 5-HT1A receptors of the rat dorsal raphe lateral wings and dorsomedial subnuclei differentially control anxiety- and panic-related defensive responses. Neuropharmacology 2016, 107, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Forster, E.A.; Cliffe, I.A.; Bill, D.J.; Dover, G.M.; Jones, D.; Reilly, Y.; Fletcher, A. A pharmacological profile of the selective silent 5-HT1A receptor antagonist, WAY-100635. Eur. J. Pharmacol. 1995, 281, 81–88. [Google Scholar] [CrossRef]
- Middlemiss, D.N.; Fozard, J.R. 8-OH-DPAT discriminates between subtypes of the 5HT1 recognition site. Eur. J. Pharmacol. 1983, 90, 151–153. [Google Scholar] [CrossRef]
- Chopin, P.; Briley, M. Animal models of anxiety: The effect of compounds that modify 5-HT neurotransmission. Trends Pharmacol. Sci. 1987, 8, 383–388. [Google Scholar] [CrossRef]
- Akimova, E.; Lanzenberger, R.; Kasper, S. The serotonin-1A receptor in anxiety disorders. Biol. Psychiatry 2009, 66, 627–635. [Google Scholar] [CrossRef]
- Olivier, J.D.A.; Olivier, B. Translational Studies in the Complex Role of Neurotransmitter Systems in Anxiety and Anxiety Disorders. Adv. Exp. Med. Biol. 2020, 1191, 121–140. [Google Scholar]
- Depoortère, R.; Bardin, L.; Varney, M.A.; Newman-Tancredi, A. Serotonin 5-HT1A Receptor Biased Agonists Display Differential Anxiolytic Activity in a Rat Social Interaction Model. ACS Chem. Neurosci. 2019, 10, 3101–3107. [Google Scholar] [CrossRef]
- Andrade, T.G.; Zangrossi, H., Jr.; Graeff, F.G. The median raphe nucleus in anxiety revisited. J. Psychopharmacol. 2013, 27, 1107–1115. [Google Scholar] [CrossRef]
- Ciranna, L. Serotonin as a modulator of glutamate- and GABA-mediated neurotransmission: Implications in physiological functions and in pathology. Curr. Neuropharmacol. 2006, 4, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Bijlsma, E.Y.; Ter Heegde, F.; Verdouw, M.P.; Garssen, J.; Newman-Tancredi, A.; Groenink, L. Activation of somatodendritic 5-HT1A autoreceptors reduces the acquisition and expression of cued fear in the rat fear-potentiated startle test. Psychopharmacology 2019, 236, 1171–1185. [Google Scholar] [CrossRef] [Green Version]
- Celada, P.; Bortolozzi, A.; Artigas, F. Serotonin 5-HT1A receptors as targets for agents to treat psychiatric disorders: Rationale and current status of research. CNS Drugs 2013, 27, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, A.; Forster, E.A.; Bill, D.J.; Brown, G.; Cliffe, I.A.; Hartley, J.E.; Jones, D.E.; McLenachan, A.; Stanhope, K.J.; Critchley, D.J.; et al. Electrophysiological, biochemical, neurohormonal and behavioural studies with WAY-100635, a potent, selective and silent 5-HT1A receptor antagonist. Behav. Brain Res. 1996, 73, 337–353. [Google Scholar] [CrossRef]
- Hjorth, S.; Westlin, D.; Bengtsson, H.J. WAY100635-induced augmentation of the 5-HT-elevating action of citalopram: Relative importance of the dose of the 5-HT1A (auto) receptor blocker versus that of the 5-HT reuptake inhibitor. Neuropharmacology 1997, 36, 461–465. [Google Scholar] [CrossRef]
- Muller, C.P.; Carey, R.J.; De Souza Silva, M.A.; Jocham, G.; Huston, J.P. Cocaine increases serotonergic activity in the hippocampus and nucleus accumbens in vivo: 5-HT1A-receptor antagonism blocks behavioural but potentiates serotonergic activation. Synapse 2002, 45, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Fornal, C.A.; Metzler, C.W.; Gallegos, R.A.; Veasez, S.C.; McCreary, A.C.; Jacobs, B.L. WAY-100635, a potent and selective 5-hydroxytryptamine1A antagonist, increases serotonergic neuronal activity in behaving cats: Comparison with (S)-WAY-100135. J. Pharmacol. Exp. Ther. 1996, 278, 752–762. [Google Scholar] [PubMed]
- Mundey, M.K.; Fletcher, A.; Marsden, C.A. Effects of 8-OH-DPAT and 5-HT1A antagonists WAY 100135 and WAY 100635, on guinea-pig behaviour and dorsal raphe 5-HT neuron firing. Br. J. Pharmacol. 1996, 117, 750–756. [Google Scholar] [CrossRef] [Green Version]
- Carey, R.J.; DePalma, G.; Damianopoulos, E.; Hopkins, A.; Shanahan, A.; Müller, C.P.; Huston, J.P. Dopaminergic and serotonergic autoreceptor stimulation effects are equivalent and additive in the suppression of spontaneous and cocaine induced locomotor activity. Brain Res. 2004, 1019, 134–143. [Google Scholar] [CrossRef]
- Carey, R.J.; DePalma, G.; Damianopoulos, E.; Müller, C.P.; Huston, J.P. The 5-HT1A receptor and behavioral stimulation in the rat: Effects of 8-OHDPAT on spontaneous and cocaine-induced behavior. Psychopharmacology 2004, 177, 46–54. [Google Scholar] [CrossRef]
- Carey, R.J.; Shanahan, A.; Damianopoulos, E.N.; Müller, C.P.; Huston, J.P. Behavior selectively elicited by novel stimuli: Modulation by the 5-HT1A agonist 8-OHDPAT and antagonist WAY-100635. Behav. Pharmacol. 2008, 19, 361–364. [Google Scholar] [CrossRef]
- Cao, B.J.; Rodgers, R.J. Influence of 5-HT1A receptor antagonism on plus-maze behaviour in mice. II. WAY 100635, SDZ 216-525 and NAN-190. Pharmacol. Biochem. Behav. 1997, 58, 593–603. [Google Scholar] [CrossRef]
- Griebel, G.; Rodgers, R.J.; Perrault, G.; Sanger, D.J. Behavioural profiles in the mouse defence test battery suggest anxiolytic potential of 5-HT (1A) receptor antagonists. Psychopharmacology 1999, 144, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Griebel, G.; Rodgers, R.J.; Perrault, G.; Sanger, D.J. The effects of compounds varying in selectivity as 5-HT (1A) receptor antagonists in three rat models of anxiety. Neuropharmacology 2000, 39, 1848–1857. [Google Scholar] [CrossRef]
- Barros, M.; Mello, E.L.; Maior, R.S.; Müller, C.P.; de Souza Silva, M.A.; Carey, R.J.; Huston, J.P.; Tomaz, C. Anxiolytic-like effects of the selective 5-HT1A receptor antagonist WAY 100635 in non-human primates. Eur. J. Pharmacol. 2003, 482, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, R.J.; Griebel, G.; Guardiola-Lemaître, B.; Brush, M.M.; Lee, N.; Blanchard, D.C. An ethopharmacological analysis of selective activation of 5-HT1A receptors: The mouse 5-HT1A syndrome. Pharmacol. Biochem. Behav. 1997, 57, 897–908. [Google Scholar] [CrossRef]
- Critchley, M.A.E.; Handley, S.L. Effects in the X-maze anxiety model of agents acting at 5-HT1 and 5-HT2 receptors. Psychopharmacology 1987, 89, 502–506. [Google Scholar] [CrossRef]
- Quines, C.B.; Rosa, S.G.; Da Rocha, J.T.; Gai, B.M.; Bortolatto, C.F.; Duarte, M.M.; Nogueira, C.W. Monosodium glutamate, a food additive, induces depressive-like and anxiogenic-like behaviors in young rats. Life Sci. 2014, 107, 27–31. [Google Scholar] [CrossRef]
- Onaolapo, O.J.; Aremu, O.S.; Onaolapo, A.Y. Monosodium glutamate-associated alterations in open field, anxiety-related and conditioned place preference behaviours in mice. Naunyn. Schmied. Arch. Pharmacol. 2017, 390, 677–689. [Google Scholar] [CrossRef]
- Taylor, N.E.; Pei, J.; Zhang, J.; Vlasov, K.Y.; Davis, T.; Taylor, E.; Weng, F.J.; Van Dort, C.J.; Solt, K.; Brown, E.N. The Role of Glutamatergic and Dopaminergic Neurons in the Periaqueductal Gray/Dorsal Raphe: Separating Analgesia and Anxiety. eNeuro 2019, 6, ENEURO.0018-18.2019. [Google Scholar] [CrossRef]
- Schmitz, D.; Empson, R.M.; Heinemann, U. Serotonin and 8-OH-DPAT reduce excitatory transmission in rat hippocampal area CA1 via reduction in presumed presynaptic Ca2 + entry. Brain Res. 1995, 701, 249–254. [Google Scholar] [CrossRef]
- Robbins, M.J.; Starr, K.R.; Honey, A.; Soffin, E.M.; Rourke, C.; Jones, G.A.; Kelly, F.M.; Strum, J.; Melarange, R.A.; Harris, A.J.; et al. Evaluation of the mGlu8 receptor as a putative therapeutic target in schizophrenia. Brain. Res. 2007, 1152, 215–227. [Google Scholar] [CrossRef]
- Bahi, A. Decreased anxiety, voluntary ethanol intake and ethanol-induced CPP acquisition following activation of the metabotropic glutamate receptor 8 “mGluR8”. Pharmacol. Biochem. Behav. 2017, 155, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, E.; Marabese, I.; de Novellis, V.; Rossi, F.; Maione, S. Supraspinal metabotropic glutamate receptors: A target for pain relief and beyond. Eur. J. Neurosci. 2014, 39, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Jastrzbska-Więsek, M.; Partyka, A.; Rychtyk, J.; Śniecikowska, J.; Kołaczkowski, M.; Wesoowska, A.; Varney, M.A.; Newman-Tancredi, A. Activity of Serotonin 5-HT1A Receptor Biased Agonists in Rat: Anxiolytic and Antidepressant-like properties. ACS Chem. Neurosci. 2018, 9, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Tabari, M.A.; Bagherpour, M.A.T. Evidence for the involvement of the GABAergic, but not serotonergic transmission in the anxiolytic-like effect of bisabolol in the mouse elevated plus maze. Naunyn-Schmiedeberg’s. Arch. Pharmacol. 2017, 390, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Walsh, R.N.; Cummins, R.A. The Open-Field Test: A critical review. Psychol. Bull. 1976, 83, 482–504. [Google Scholar] [CrossRef]
- Pellow, S.; Chopin, P.; File, S.E.; Briley, M. Validation of open closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Meth. 1985, 14, 149–167. [Google Scholar] [CrossRef]
- Jones, B.J.; Roberts, D.J. The quantitative measurement of motor incoordination in naive mice using an accelerating rotarod. J. Pharm. Pharmacol. 1968, 20, 302–304. [Google Scholar] [CrossRef]
- Scuteri, D.; Rombolà, L.; Morrone, L.A.; Bagetta, G.; Sakurada, S.; Sakurada, T.; Tonin, P.; Corasaniti, M.T. Neuropharmacology of the Neuropsychiatric Symptoms of Dementia and Role of Pain: Essential Oil of Bergamot as a Novel Therapeutic Approach. Int. J. Mol. Sci. 2019, 20, 3327. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rombolà, L.; Scuteri, D.; Watanabe, C.; Sakurada, S.; Hamamura, K.; Sakurada, T.; Tonin, P.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Role of 5-HT1A Receptor in the Anxiolytic-Relaxant Effects of Bergamot Essential Oil in Rodent. Int. J. Mol. Sci. 2020, 21, 2597. https://doi.org/10.3390/ijms21072597
Rombolà L, Scuteri D, Watanabe C, Sakurada S, Hamamura K, Sakurada T, Tonin P, Corasaniti MT, Bagetta G, Morrone LA. Role of 5-HT1A Receptor in the Anxiolytic-Relaxant Effects of Bergamot Essential Oil in Rodent. International Journal of Molecular Sciences. 2020; 21(7):2597. https://doi.org/10.3390/ijms21072597
Chicago/Turabian StyleRombolà, Laura, Damiana Scuteri, Chizuko Watanabe, Shinobu Sakurada, Kengo Hamamura, Tsukasa Sakurada, Paolo Tonin, Maria Tiziana Corasaniti, Giacinto Bagetta, and Luigi Antonio Morrone. 2020. "Role of 5-HT1A Receptor in the Anxiolytic-Relaxant Effects of Bergamot Essential Oil in Rodent" International Journal of Molecular Sciences 21, no. 7: 2597. https://doi.org/10.3390/ijms21072597
APA StyleRombolà, L., Scuteri, D., Watanabe, C., Sakurada, S., Hamamura, K., Sakurada, T., Tonin, P., Corasaniti, M. T., Bagetta, G., & Morrone, L. A. (2020). Role of 5-HT1A Receptor in the Anxiolytic-Relaxant Effects of Bergamot Essential Oil in Rodent. International Journal of Molecular Sciences, 21(7), 2597. https://doi.org/10.3390/ijms21072597