Structural Characterization of an ACP from Thermotoga maritima: Insights into Hyperthermal Adaptation


Abstract
:
1. Introduction
2. Results
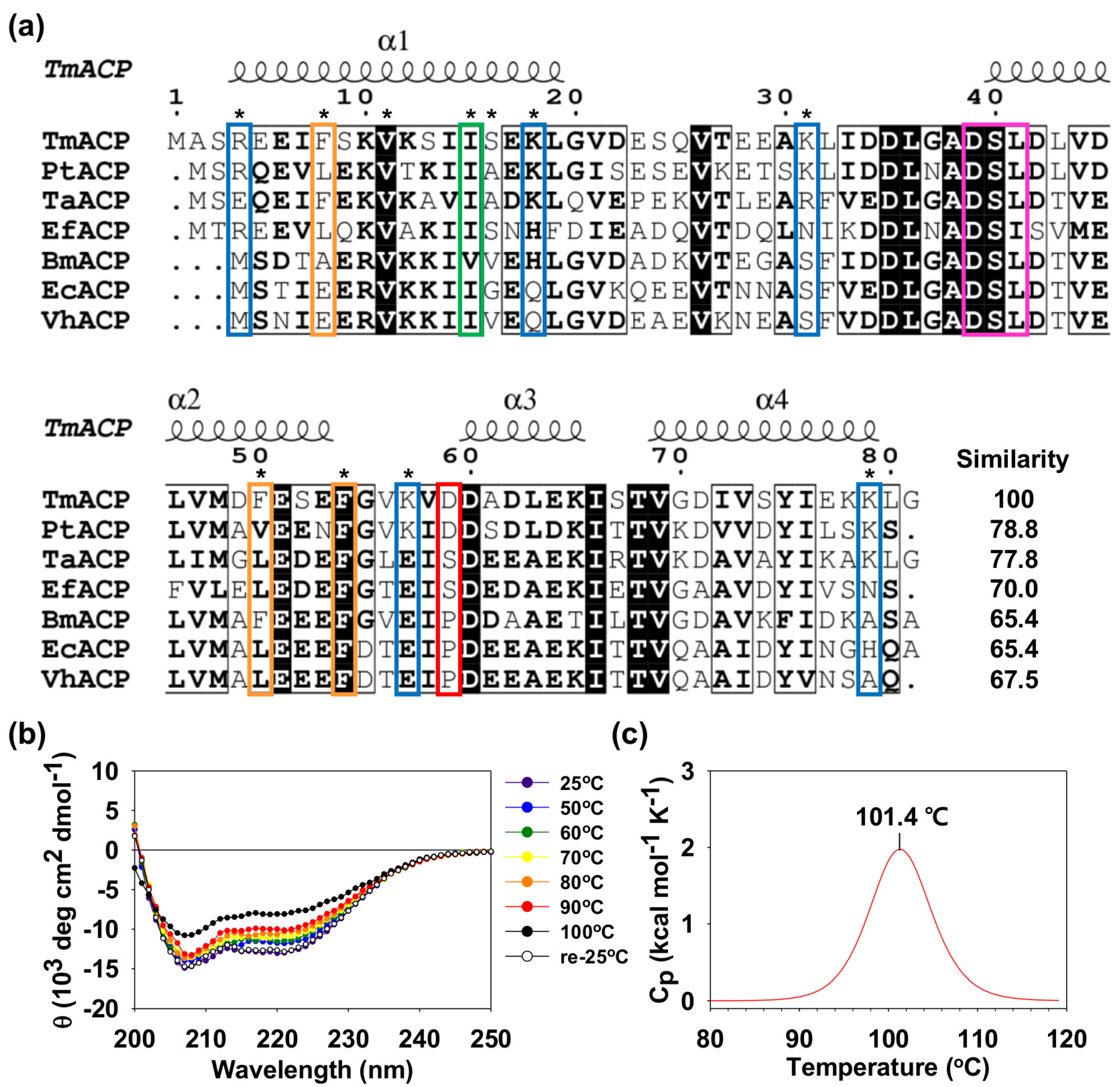
2.1. Thermostability of Tm-ACP
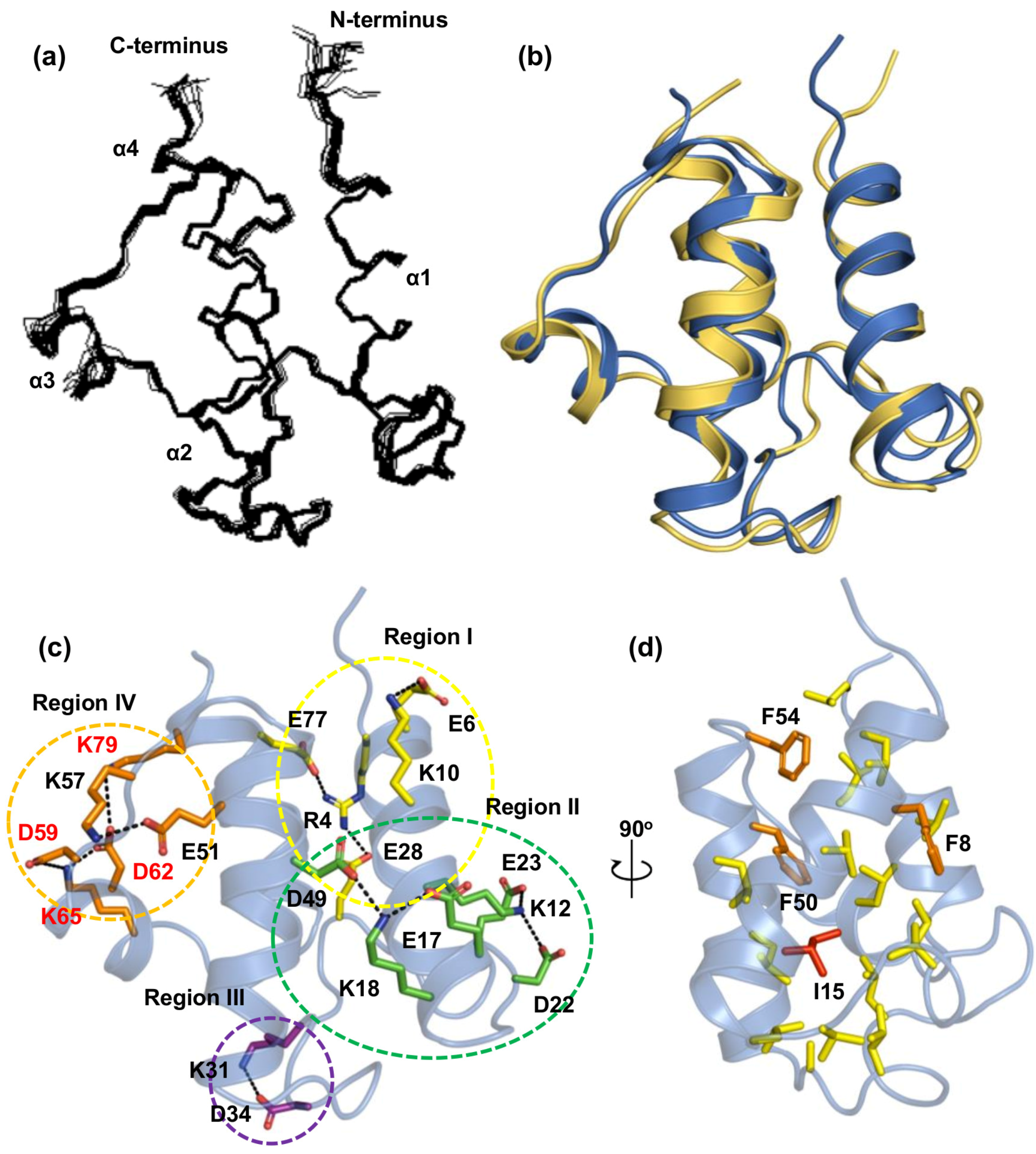
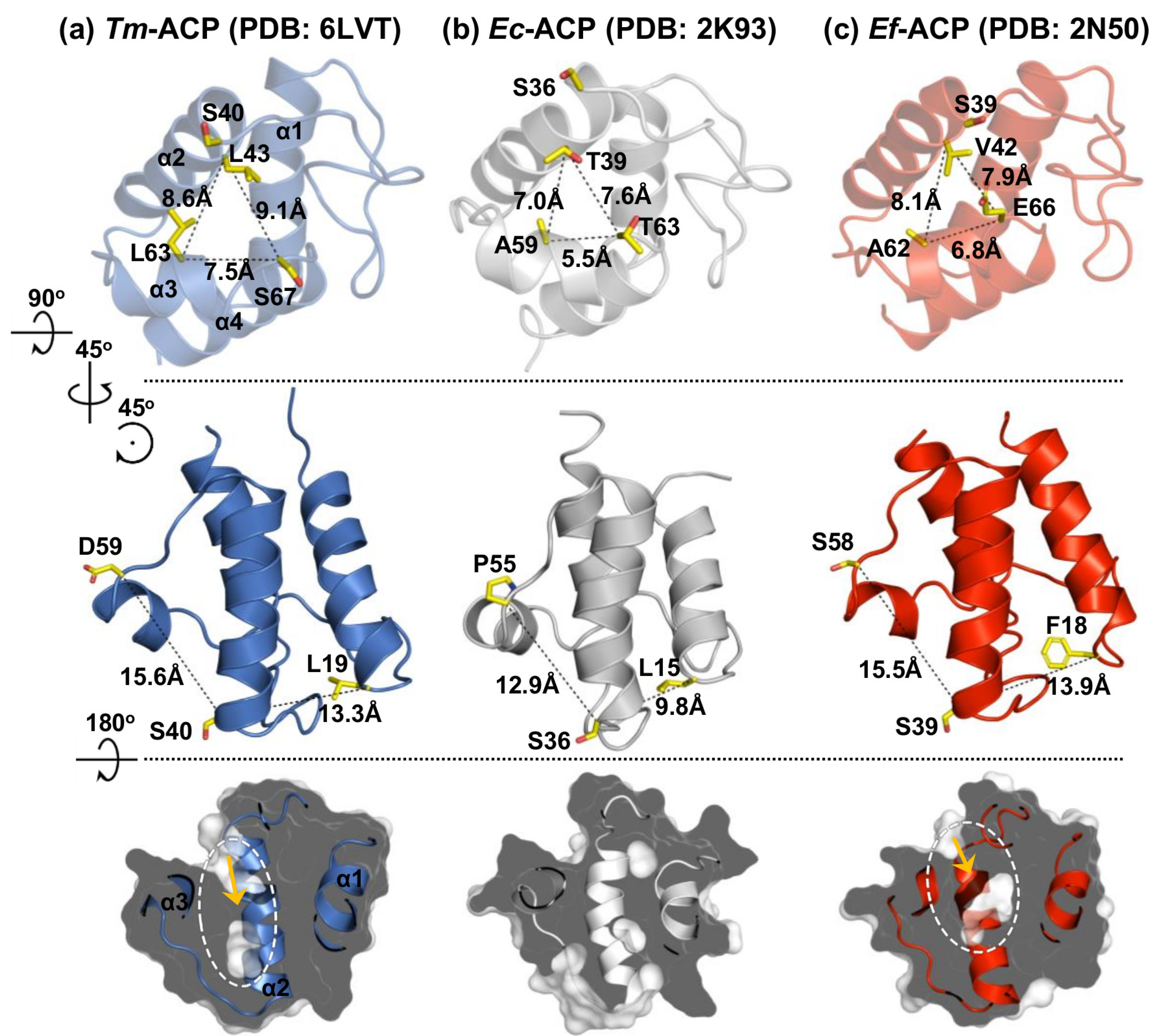
2.2. Tertiary Structure of Tm-ACP
2.3. Key Residues Contributing to the Thermostability of Tm-ACP
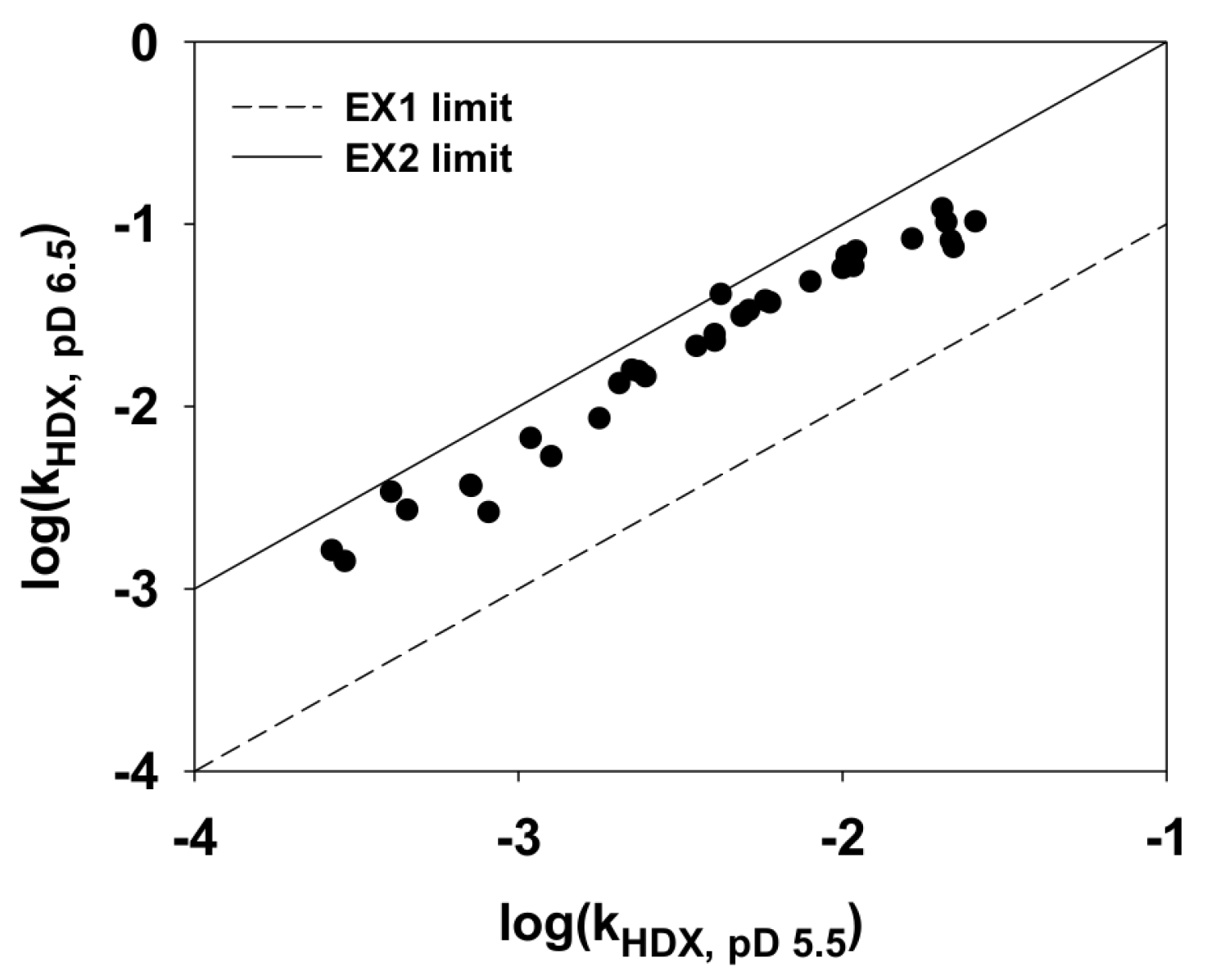
2.4. Hydrogen/Deuterium (H/D) Exchange Experiments
2.5. Chemical Denaturation of Tm-ACP
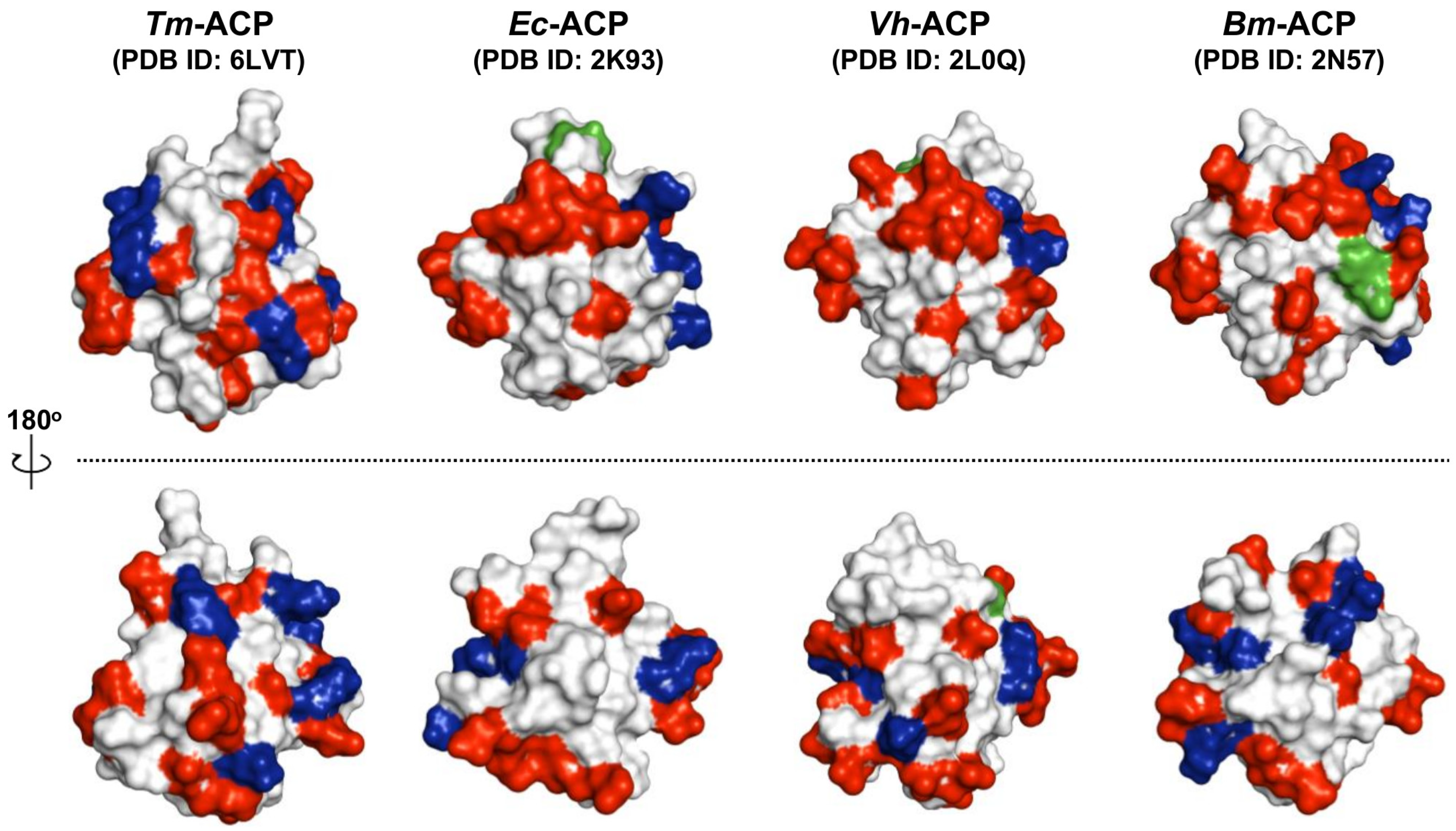
3. Discussion
4. Materials and Methods
4.1. Cloning, Expression, Isotopic Enrichment, and Purification
4.2. Site-Directed Mutagenesis
4.3. NMR Experiments and Assignments
4.4. Solution Structure Calculation
4.5. Hydrogen/Deuterium Exchange Experiments
4.6. Circular Dichroism Experiment
4.7. Chemical Denaturation Experiment
4.8. Differential Scanning Calorimetry
4.9. X-ray Crystallography
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACP | Acyl carrier protein |
| ACPS | Holo-ACP synthase |
| Bm | Brucella melitenis |
| CD | Circular dichroism |
| CoA | Coenzyme A |
| DSC | Differential scanning calorimetry |
| DSL | Asp-Ser-Leu |
| DSS | 2,2-Dimethyl-2-silapentane-5-sulfonate |
| DTT | Dithiothreitol |
| Ec | Escherichia coli |
| Ef | Enterococcus faecalis |
| FAS | Fatty acid synthesis |
| H/D | Hydrogen/deuterium |
| HSQC | Heteronuclear single quantum coherence spectroscopy |
| IPTG | Isopropyl β-D-1-thiogalactopyranoside |
| NOE | Nuclear Overhauser effect |
| OD | Optical density |
| PCR | polymerase chain reaction |
| PDB | Protein data bank |
| PEG | Polyethylene glycol |
| Pt | Pseudothermotoga thermarum |
| RDC | Residual dipolar coupling |
| RMSD | Root mean square deviation |
| RMSZ | Root mean square of Z-scores |
| Ta | Thermus aquaticus |
| Tm | Thermotoga maritima |
| Vh | Vibrio harveyi |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutant | Direction | Primer Sequence |
|---|---|---|
| R4E | forward | 5′-gaa gga gat ata cat atg gcc agt gaa gaa gaa att ttt tct aaa gtg aaa tc-3′ |
| reverse | 5′-gat ttc act tta gaa aaa att tct tct tca ctg gcc ata tgt ata tct cct tc-3′ | |
| K10E | forward | 5′-cgg gaa gaa att ttt tct gaa gtg aaa tcc atc atc tc-3′ |
| reverse | 5′-gag atg atg gat ttc act tca gaa aaa att tct tcc cg-3′ | |
| K12E | forward | 5′-gaa att ttt tct aaa gtg gaa tcc atc atc tct gaa-3′ |
| reverse | 5′-ttc aga gat gat gga ttc cac ttt aga aaa aat ttc-3′ | |
| K18E | forward | 5′-cca tca tct ctg aag aat tgg ggg tcg atg-3′ |
| reverse | 5′-cat cga ccc cca att ctt cag aga tga tgg-3′ | |
| K31E | forward | 5′-gtg acg gaa gag gcc gaa ttg att gat gat ctg gga g-3′ |
| reverse | 5′-ctc cca gat cat caa tca att cgg cct ctt ccg tca c-3′ | |
| K57E | forward | 5′-gtg agt tcg gcg ttg aag tcg atg acg ccg-3′ |
| reverse | 5′-cgg cgt cat cga ctt caa cgc cga act cac-3′ | |
| K79E | forward | 5′-gtc agc tac att gaa aaa gaa tta ggc tag gga tcc ggc tgc-3′ |
| reverse | 5′-gca gcc gga tcc cta gcc taa ttc ttt ttc aat gta gct gac-3′ | |
| S16G | forward | 5′-cta aag tga aat cca tca tcg gtg aaa aat tgg ggg tcg atg aat c -3′ |
| reverse | 5′-gat tca tcg acc ccc aat ttt tca ccg atg atg gat ttc act tta g-3′ | |
| F8A | forward | 5′-gcc agt cgg gaa gaa att gcg tct aaa gtg aaa tcc tag-3′ |
| reverse | 5′-cta gga ttt cac ttt aga cgc aat ttc ttc ccg act ggc-3′ | |
| F50A | forward | 5′-gac ctg gta atg gac gcg gag agt gag ttc gg-3′ |
| reverse | 5′-ccg aac tca ctc tcc gcg tcc att acc agg tc-3′ | |
| F54A | forward | 5′-gga ctt tga gag tga ggc ggg cgt taa agt cg′a tg-3′ |
| reverse | 5′-cat cga ctt taa cgc ccg cct cac tct caa agt cc-3′ | |
| V11A | forward | 5′-cgg gaa gaa att ttt tct aaa gcg aaa tcc atc atc tct gaa aaa ttg g-3′ |
| reverse | 5′-cca att ttt cag aga tga tgg att tcg ctt tag aaa aaa ttt ctt ccc g-3′ | |
| I15A | forward | 5′-gaa att ttt tct aaa gtg aaa tcc atc gcc tct gaa aaa ttg ggg gtc g-3′ |
| reverse | 5′-cga ccc cca att ttt cag agg cga tgg att tca ctt tag aaa aaa ttt c-3′ | |
| I72A | forward | 5′-gaa aat ctc tac tgt ggg cga cgc cgt cag cta cat tga aaa aaa att ag-3′ |
| reverse | 5′-cta att ttt ttt caa tgt agc tga cgg cgt cgc cca cag tag aga ttt tc-3′ | |
| V73A | forward | 5′-cta ctg tgg gcg aca ttg cca gct aca ttg aaa aaa aat tag-3′ |
| reverse | 5′-cta att ttt ttt caa tgt agc tgg caa tgt cgc cca cag tag-3′ |
References
- Chan, D.I.; Vogel, H.J. Current understanding of fatty acid biosynthesis and the acyl carrier protein. Biochem. J. 2010, 430, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.; Kong, R.; Murugan, E.; Ho, C.L.; Liang, Z.X.; Yang, D. Solution structures of the acyl carrier protein domain from the highly reducing type I iterative polyketide synthase CalE8. PLoS ONE 2011, 6, e20549. [Google Scholar] [CrossRef] [PubMed]
- Masoudi, A.; Raetz, C.R.H.; Zhou, P.; Pemble, C.W. Chasing acyl carrier protein through a catalytic cycle of lipid A production. Nature 2014, 505, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heaton, M.P.; Neuhaus, F.C. Role of the D-alanyl carrier protein in the biosynthesis of D-alanyl-lipoteichoic acid. J. Bacteriol. 1994, 176, 681–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, O.; Spaink, H.P.; Kennedy, E.P. Isolation of the Rhizobium leguminosarum NodF nodulation protein: NodF carries a 4'-phosphopantetheine prosthetic group. J. Bacteriol. 1991, 173, 2872–2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Issartel, J.P.; Koronakis, V.; Hughes, C. Activation of Escherichia coli prohaemolysin to the mature toxin by acyl carrier protein-dependent fatty acylation. Nature 1991, 351, 759–761. [Google Scholar] [CrossRef]
- Wakil, S.J.; Stoops, J.K.; Joshi, V.C. Fatty acid synthesis and its regulation. Annu. Rev. Biochem. 1983, 52, 537–579. [Google Scholar] [CrossRef]
- Crosby, J.; Crump, M.P. The structural role of the carrier protein--active controller or passive carrier. Nat. Prod. Rep. 2012, 29, 1111–1137. [Google Scholar] [CrossRef]
- Chan, D.I.; Stockner, T.; Tieleman, D.P.; Vogel, H.J. Molecular Dynamics Simulations of the Apo-, Holo-, and Acyl-forms of Escherichia coli Acyl Carrier Protein. J. Biol. Chem. 2008, 283, 33620–33629. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.; Haushalter, R.W.; Lee, D.J.; Markwick, P.R.L.; Bruegger, J.; Caldara-Festin, G.; Finzel, K.; Jackson, D.R.; Ishikawa, F.; O’Dowd, B.; et al. Trapping the dynamic acyl carrier protein in fatty acid biosynthesis. Nature 2014, 505, 427–431. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.N.; Zhang, Y.M.; Jie, Z.; Rock, C.O. Key residues responsible for acyl carrier protein (ACP) and beta-ketoacyl-acyl carrier protein reductase (FabG) interaction. Biophys. J. 2004, 86, 495. [Google Scholar]
- Flugel, R.S.; Hwangbo, Y.; Lambalot, R.H.; Cronan, J.E.; Walsh, C.T. Holo-(acyl carrier protein) synthase and phosphopantetheinyl transfer in Escherichia coli. J. Biol. Chem. 2000, 275, 959–968. [Google Scholar] [CrossRef] [Green Version]
- Sztain, T.; Patel, A.; Lee, D.J.; Davis, T.D.; McCammon, J.A.; Burkart, M.D. Modifying the Thioester Linkage Affects the Structure of the Acyl Carrier Protein. Angew. Chem. Int. Ed. Engl. 2019, 58, 10888–10892. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.N.; Londergan, C.H.; Charkoudian, L.K. Probing the phosphopantetheine arm conformations of acyl carrier proteins using vibrational spectroscopy. J. Am. Chem. Soc. 2014, 136, 11240–11243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, D.I.; Chu, B.C.; Lau, C.K.; Hunter, H.N.; Byers, D.M.; Vogel, H.J. NMR solution structure and biophysical characterization of Vibrio harveyi acyl carrier protein A75H: Effects of divalent metal ions. J. Biol. Chem. 2010, 285, 30558–30566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crump, M.P.; Crosby, J.; Dempsey, C.E.; Parkinson, J.A.; Murray, M.; Hopwood, D.A.; Simpson, T.J. Solution structure of the actinorhodin polyketide synthase acyl carrier protein from Streptomyces coelicolor A3(2). Biochemistry 1997, 36, 6000–6008. [Google Scholar] [CrossRef]
- Holak, T.A.; Kearsley, S.K.; Kim, Y.; Prestegard, J.H. Three-dimensional structure of acyl carrier protein determined by NMR pseudoenergy and distance geometry calculations. Biochemistry 1988, 27, 6135–6142. [Google Scholar] [CrossRef]
- Park, Y.G.; Jung, M.C.; Song, H.; Jeong, K.W.; Bang, E.; Hwang, G.S.; Kim, Y. Novel Structural Components Contribute to the High Thermal Stability of Acyl Carrier Protein from Enterococcus faecalis. J. Biol. Chem. 2016, 291, 1692–1702. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.K.; Sharma, S.K.; Surolia, A.; Surolia, N.; Sarma, S.P. Solution structures of conformationally equilibrium forms of holo-acyl carrier protein (PfACP) from Plasmodium falciparum provides insight into the mechanism of activation of ACPs. Biochemistry 2006, 45, 6904–6916. [Google Scholar] [CrossRef] [Green Version]
- Wong, H.C.; Liu, G.H.; Zhang, Y.M.; Rock, C.O.; Zheng, J. The solution structure of acyl carrier protein from Mycobacterium tuberculosis. J. Biol. Chem. 2002, 277, 15874–15880. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.Y.; Tam, A.; Lin, L.; Hixon, J.; Fritz, C.C.; Powers, R. Solution structure of B. subtilis acyl carrier protein. Structure 2001, 9, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Barnwal, R.P.; Van Voorhis, W.C.; Varani, G. NMR structure of an acyl-carrier protein from Borrelia burgdorferi. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 1137–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnwal, R.P.; Kaur, M.; Heckert, A.; Gartia, J.; Varani, G. Comparative structure, dynamics and evolution of acyl-carrier proteins from Borrelia burgdorferi, Brucella melitensis and Rickettsia prowazekii. Biochem. J. 2020, 477, 491–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Kovrigin, E.L.; Eletr, Z. NMR studies of Escherichia coli acyl carrier protein: Dynamic and structural differences of the apo- and holo-forms. Biochem. Biophys. Res. Commun. 2006, 341, 776–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.; Xiao, T.; Fan, J.; Yang, D. An off-pathway folding intermediate of an acyl carrier protein domain coexists with the folded and unfolded states under native conditions. Angew. Chem. Int. Ed. Engl. 2014, 53, 2358–2361. [Google Scholar] [CrossRef] [PubMed]
- Colizzi, F.; Masetti, M.; Recanatini, M.; Cavalli, A. Atomic-Level Characterization of the Chain-Flipping Mechanism in Fatty-Acids Biosynthesis. J. Phys. Chem. Lett. 2016, 7, 2899–2904. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, D. Equilibrium folding dynamics of meACP in water, heavy water, and low concentration of urea. Sci. Rep. 2017, 7, 16156. [Google Scholar] [CrossRef]
- Arya, R.; Sharma, B.; Dhembla, C.; Pal, R.K.; Patel, A.K.; Sundd, M.; Ghosh, B.; Makde, R.D.; Kundu, S. A conformational switch from a closed apo- to an open holo-form equips the acyl carrier protein for acyl chain accommodation. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 163–174. [Google Scholar] [CrossRef]
- Brininger, C.; Spradlin, S.; Cobani, L.; Evilia, C. The more adaptive to change, the more likely you are to survive: Protein adaptation in extremophiles. Semin. Cell Dev. Biol. 2018, 84, 158–169. [Google Scholar] [CrossRef]
- Hait, S.; Mallik, S.; Basu, S.; Kundu, S. Finding the generalized molecular principles of protein thermal stability. Proteins 2019. [Google Scholar] [CrossRef]
- Huber, R.; Langworthy, T.A.; König, H.; Thomm, M.; Woese, C.R.; Sleytr, U.B.; Stetter, K.O. Thermotoga maritima sp. nov. represents a new genus of unique extremely thermophilic eubacteria growing up to 90°C. Arch. Microbiol. 1986, 144, 324–333. [Google Scholar] [CrossRef]
- Carballeira, N.M.; Reyes, M.; Sostre, A.; Huang, H.S.; Verhagen, M.F.J.M.; Adams, M.W.W. , Unusual fatty acid compositions of the hyperthermophilic archaeon Pyrococcus furiosus and the bacterium Thermotoga maritima. J. Bacteriol. 1997, 179, 2766–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, Y. Thermal adaptation of the archaeal and bacterial lipid membranes. Archaea 2012, 2012, 789652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, L.A.; Sturtevant, J.M.; Prestegard, J.H. Kinetics and thermodynamics of thermal denaturation in acyl carrier protein. Protein Sci. 1994, 3, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [PubMed]
- Cornilescu, G.; Marquardt, J.L.; Ottiger, M.; Bax, A. Validation of protein structure from anisotropic carbonyl chemical shifts in a dilute liquid crystalline phase. J. Am. Chem. Soc. 1998, 120, 6836–6837. [Google Scholar] [CrossRef]
- Ramelot, T.A.; Rossi, P.; Forouhar, F.; Lee, H.W.; Yang, Y.; Ni, S.; Unser, S.; Lew, S.; Seetharaman, J.; Xiao, R.; et al. Structure of a specialized acyl carrier protein essential for lipid A biosynthesis with very long-chain fatty acids in open and closed conformations. Biochemistry 2012, 51, 7239–7249. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.N.; Zhang, Y.M.; Rock, C.O.; Zheng, J.J. Structural modification of acyl carrier protein by butyryl group. Protein Sci. 2009, 18, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Stark, J.L.; Markley, J.L. PONDEROSA-C/S: Client-server based software package for automated protein 3D structure determination. J. Biomol. NMR 2014, 60, 73–75. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Cornilescu, G.; Dashti, H.; Eghbalnia, H.R.; Tonelli, M.; Westler, W.M.; Butcher, S.E.; Henzler-Wildman, K.A.; Markley, J.L. Integrative NMR for biomolecular research. J. Biomol. NMR 2016, 64, 307–332. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, A.; Tejero, R.; Montelione, G.T. Evaluating protein structures determined by structural genomics consortia. Proteins 2007, 66, 778–795. [Google Scholar] [CrossRef] [PubMed]
- Frederick, A.F.; Kay, L.E.; Prestegard, J.H. Location of divalent ion sites in acyl carrier protein using relaxation perturbed 2D NMR. FEBS Lett. 1988, 238, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Kleywegt, G.J. Validation of protein models from Calpha coordinates alone. J. Mol. Biol. 1997, 273, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Milne, J.S.; Mayne, L.; Englander, S.W. Primary structure effects on peptide group hydrogen exchange. Proteins 1993, 17, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Englander, S.W. Protein folding intermediates and pathways studied by hydrogen exchange. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 213–238. [Google Scholar] [CrossRef] [Green Version]
- Laity, J.H.; Montelione, G.T.; Scheraga, H.A. Comparison of local and global stability of an analogue of a disulfide-folding intermediate with those of the wild-type protein in bovine pancreatic ribonuclease A: Identification of specific regions of stable structure along the oxidative folding pathway. Biochemistry 1999, 38, 16432–16442. [Google Scholar] [CrossRef]
- Santoro, M.M.; Bolen, D.W. A test of the linear extrapolation of unfolding free energy changes over an extended denaturant concentration range. Biochemistry 1992, 31, 4901–4907. [Google Scholar] [CrossRef]
- Pace, C.N. Determination and analysis of urea and guanidine hydrochloride denaturation curves. Methods Enzymol. 1986, 131, 266–280. [Google Scholar]
- Schellman, J.A. The thermodynamic stability of proteins. Annu. Rev. Biophys. Biophys. Chem. 1987, 16, 115–137. [Google Scholar] [CrossRef]
- Padmanabhan, S.; Laurents, D.V.; Fernandez, A.M.; Elias-Arnanz, M.; Ruiz-Sanz, J.; Mateo, P.L.; Rico, M.; Filimonov, V.V. Thermodynamic analysis of the structural stability of phage 434 Cro protein. Biochemistry 1999, 38, 15536–15547. [Google Scholar] [CrossRef] [Green Version]
- Zavodszky, P.; Kardos, J.; Svingor, A.; Petsko, G.A. Adjustment of conformational flexibility is a key event in the thermal adaptation of proteins. Proc. Natl. Acad. Sci. USA 1998, 95, 7406–7411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaenicke, R.; Bohm, G. The stability of proteins in extreme environments. Curr. Opin. Struct. Biol. 1998, 8, 738–748. [Google Scholar] [CrossRef]
- Gershenson, A.; Schauerte, J.A.; Giver, L.; Arnold, F.H. Tryptophan phosphorescence study of enzyme flexibility and unfolding in laboratory-evolved thermostable esterases. Biochemistry 2000, 39, 4658–4665. [Google Scholar] [CrossRef] [PubMed]
- Vieille, C.; Zeikus, G.J. Hyperthermophilic enzymes: Sources, uses, and molecular mechanisms for thermostability. Microbiol. Mol. Biol. Rev. 2001, 65, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Cui, Q.; Tong, Y.; Feng, Y.; Shan, L.; Huang, L.; Wang, J. A stabilizing alpha/beta-hydrophobic core greatly contributes to hyperthermostability of archaeal [P62A]Ssh10b. Biochemistry 2008, 47, 11212–11221. [Google Scholar] [CrossRef]
- Tych, K.M.; Batchelor, M.; Hoffmann, T.; Wilson, M.C.; Hughes, M.L.; Paci, E.; Brockwell, D.J.; Dougan, L. Differential Effects of Hydrophobic Core Packing Residues for Thermodynamic and Mechanical Stability of a Hyperthermophilic Protein. Langmuir 2016, 32, 7392–7402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roujeinikova, A.; Simon, W.J.; Gilroy, J.; Rice, D.W.; Rafferty, J.B.; Slabas, A.R. Structural studies of fatty acyl-(acyl carrier protein) thioesters reveal a hydrophobic binding cavity that can expand to fit longer substrates. J. Mol. Biol. 2007, 365, 135–145. [Google Scholar] [CrossRef]
- Zhu, L.; Zou, Q.; Cao, X.; Cronan, J.E. Enterococcus faecalis Encodes an Atypical Auxiliary Acyl Carrier Protein Required for Efficient Regulation of Fatty Acid Synthesis by Exogenous Fatty Acids. mBio 2019, 10, e00577-19. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Jeong, K.W.; Jin, B.; Ryu, K.S.; Kim, E.H.; Ahn, J.H.; Kim, Y. Structural and dynamic features of cold-shock proteins of Listeria monocytogenes, a psychrophilic bacterium. Biochemistry 2013, 52, 2492–2504. [Google Scholar] [CrossRef]
- Jeong, K.W.; Kang, D.I.; Lee, E.; Shin, A.; Jin, B.; Park, Y.G.; Lee, C.K.; Kim, E.H.; Jeon, Y.H.; Kim, E.E.; et al. Structure and backbone dynamics of vanadate-bound PRL-3: Comparison of 15N nuclear magnetic resonance relaxation profiles of free and vanadate-bound PRL-3. Biochemistry 2014, 53, 4814–4825. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.; Jeong, K.W.; Kim, Y. Structure and flexibility of the thermophilic cold-shock protein of Thermus aquaticus. Biochem. Biophys. Res. Commun. 2014, 451, 402–407. [Google Scholar] [CrossRef]
- Lee, Y.; Kwak, C.; Jeong, K.W.; Durai, P.; Ryu, K.S.; Kim, E.H.; Cheong, C.; Ahn, H.C.; Kim, H.J.; Kim, Y. Tyr51: Key Determinant of the Low Thermostability of the Colwellia psychrerythraea Cold-Shock Protein. Biochemistry 2018, 57, 3625–3640. [Google Scholar] [CrossRef]
- Delaglio, F.; Grzesiek, S.; Vuister, G.W.; Zhu, G.; Pfeifer, J.; Bax, A. NMRPipe: A multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR 1995, 6, 277–293. [Google Scholar] [CrossRef]
- Lee, W.; Tonelli, M.; Markley, J.L. NMRFAM-SPARKY: Enhanced software for biomolecular NMR spectroscopy. Bioinformatics 2015, 31, 1325–1327. [Google Scholar] [CrossRef] [Green Version]
- Chou, J.J.; Gaemers, S.; Howder, B.; Louis, J.M.; Bax, A. A simple apparatus for generating stretched polyacrylamide gels, yielding uniform alignment of proteins and detergent micelles. J. Biomol. NMR 2001, 21, 377–382. [Google Scholar] [CrossRef]
- Sass, H.J.; Musco, G.; Stahl, S.J.; Wingfield, P.T.; Grzesiek, S. Solution NMR of proteins within polyacrylamide gels: Diffusional properties and residual alignment by mechanical stress or embedding of oriented purple membranes. J. Biomol. NMR 2000, 18, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Cordier, F.; Dingley, A.J.; Grzesiek, S. A doublet-separated sensitivity-enhanced HSQC for the determination of scalar and dipolar one-bond J-couplings. J. Biomol. NMR 1999, 13, 175–180. [Google Scholar] [CrossRef]
- Gangadhara, B.N.; Laine, J.M.; Kathuria, S.V.; Massi, F.; Matthews, C.R. Clusters of Branched Aliphatic Side Chains Serve As Cores of Stability in the Native State of the HisF TIM Barrel Protein. J. Mol. Biol. 2013, 425, 1065–1081. [Google Scholar] [CrossRef] [Green Version]
- Molday, R.S.; Englander, S.W.; Kallen, R.G. Primary structure effects on peptide group hydrogen exchange. Biochemistry 1972, 11, 150–158. [Google Scholar] [CrossRef]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta 2005, 1751, 119–139. [Google Scholar] [CrossRef]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]






| Tm-ACP 1 | Pt-ACP 2 | Ta-ACP 3 | Ef-ACP 4 | Bm-ACP 5 | Ec-ACP 6 | Vh-ACP 7 | |
|---|---|---|---|---|---|---|---|
| pI 8 | 4.13 | 4.28 | 4.29 | 3.87 | 3.97 | 3.98 | 3.79 |
| Acidic (Glu/Asp) | 21 (10/11) | 19 (8/11) | 21 (14/7) | 21 (11/10) | 20 (9/11) | 20 (14/6) | 22 (14/8) |
| Basic (Arg/Lys) | 9 (1/8) | 10 (1/9) | 10 (2/8) | 5 (1/4) | 6 (1/5) | 5 (1/4) | 6 (1/5) |
| Restraints 1 | |
|---|---|
| Total | 1108 |
| Conformationally restricting distance constraints | |
| Short Range ((i– j) <= 1) | 301 |
| Medium Range (1 < (i – j) ≤ 5) | 266 |
| Long Range ((i – j) > 5) | 196 |
| Dihedral angle constraints | |
| Phi | 77 |
| Psi | 77 |
| Hydrogen-bond constraints | 114 |
| Residual dipolar coupling (RDC) constraints | 77 |
| Xplor-NIH pseudo-potential energy (kJ/mol) 2 | 3251 |
| Average Rmsd to the Mean Coordinates (Å) 3 | |
| Backbone atoms (all / ordered residues 4) | 0.3/0.2 |
| Heavy atoms (all / ordered residues 4) | 0.7/0.6 |
| Ramachandran Plot Summary from PROCHECK (%) 3 | |
| Most favored regions | 96.4 |
| Allowed regions | 3.6 |
| Disallowed regions | 0.0 |
| Average Number of Violations Per Conformer 2 | |
| Distance constraint violations (> 0.2 Å) | 0 |
| Angle constraint violations (> 10°) | 0 |
| Data collection | |
| Space group | P212121 |
| Unit-cell | |
| a, b, c (Å) | 23.83, 61.95, 95.72 |
| α, β, γ (o) | 90, 90, 90 |
| Resolution (Å) | 28.37–2.29 |
| Unique reflections | 49,345 |
| Redundancy | 7.3 |
| Completeness for range (%) | 98.2 |
| Mean I/σ(I) | 26.65 (at 2.29 Å) |
| Rmerge (%) 1 | 8.1 |
| Refinement 2 | |
| No. of reflections (overall) | 6797 |
| No. of reflections (test set) | 671 |
| R-factor 3 | 0.2217(0.2104 at 2.29 Å) |
| R-free 4 | 0.2841(0.2762 at 2.29 Å) |
| RMSZ 5 / RMSD | |
| Bond lengths (Å) | 0.42/0.008 |
| Bond angles (o) | 0.63/1.086 |
| Wilson B-factor (Å2) 6 | 19.5 |
| Ramachandran Plot Summary from PROCHECK (%) 2 | |
| Most favored regions | 97.2 |
| Allowed regions | 2.8 |
| Disallowed regions | 0.0 |
| Average B-factors (Å2) 7 | |
| Protein | 27.3 |
| Water | 27.9 |
| Ligand | 40.0 |
| Related Interaction | Phenotype | Tm (°C) |
|---|---|---|
| Wild-type | 100.4 | |
| Electrostatic | R4E | 86.1 |
| K10E | 80.4 | |
| K12E | 76.9 | |
| K18E | 82.1 | |
| K31E | 90.0 | |
| K57E | 95.8 | |
| K79E | 89.8 | |
| Hydrogen bond | S16G | 94.0 |
| Hydrophobic | F8A | 94.1 |
| F50A | 90.7 | |
| F54A | 81.8 | |
| V11A | 91.6 | |
| I15A | 84.6 | |
| I72A | 90.1 | |
| V73A | 89.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.; Jang, A.; Jeong, M.-C.; Park, N.; Park, J.; Lee, W.C.; Cheong, C.; Kim, Y. Structural Characterization of an ACP from Thermotoga maritima: Insights into Hyperthermal Adaptation. Int. J. Mol. Sci. 2020, 21, 2600. https://doi.org/10.3390/ijms21072600
Lee Y, Jang A, Jeong M-C, Park N, Park J, Lee WC, Cheong C, Kim Y. Structural Characterization of an ACP from Thermotoga maritima: Insights into Hyperthermal Adaptation. International Journal of Molecular Sciences. 2020; 21(7):2600. https://doi.org/10.3390/ijms21072600
Chicago/Turabian StyleLee, Yeongjoon, Ahjin Jang, Min-Cheol Jeong, Nuri Park, Jungwoo Park, Woo Cheol Lee, Chaejoon Cheong, and Yangmee Kim. 2020. "Structural Characterization of an ACP from Thermotoga maritima: Insights into Hyperthermal Adaptation" International Journal of Molecular Sciences 21, no. 7: 2600. https://doi.org/10.3390/ijms21072600
APA StyleLee, Y., Jang, A., Jeong, M. -C., Park, N., Park, J., Lee, W. C., Cheong, C., & Kim, Y. (2020). Structural Characterization of an ACP from Thermotoga maritima: Insights into Hyperthermal Adaptation. International Journal of Molecular Sciences, 21(7), 2600. https://doi.org/10.3390/ijms21072600