The Interactions between the Antimicrobial Peptide P-113 and Living Candida albicans Cells Shed Light on Mechanisms of Antifungal Activity and Resistance

 ,
, 
Abstract
:1. Introduction
2. Results
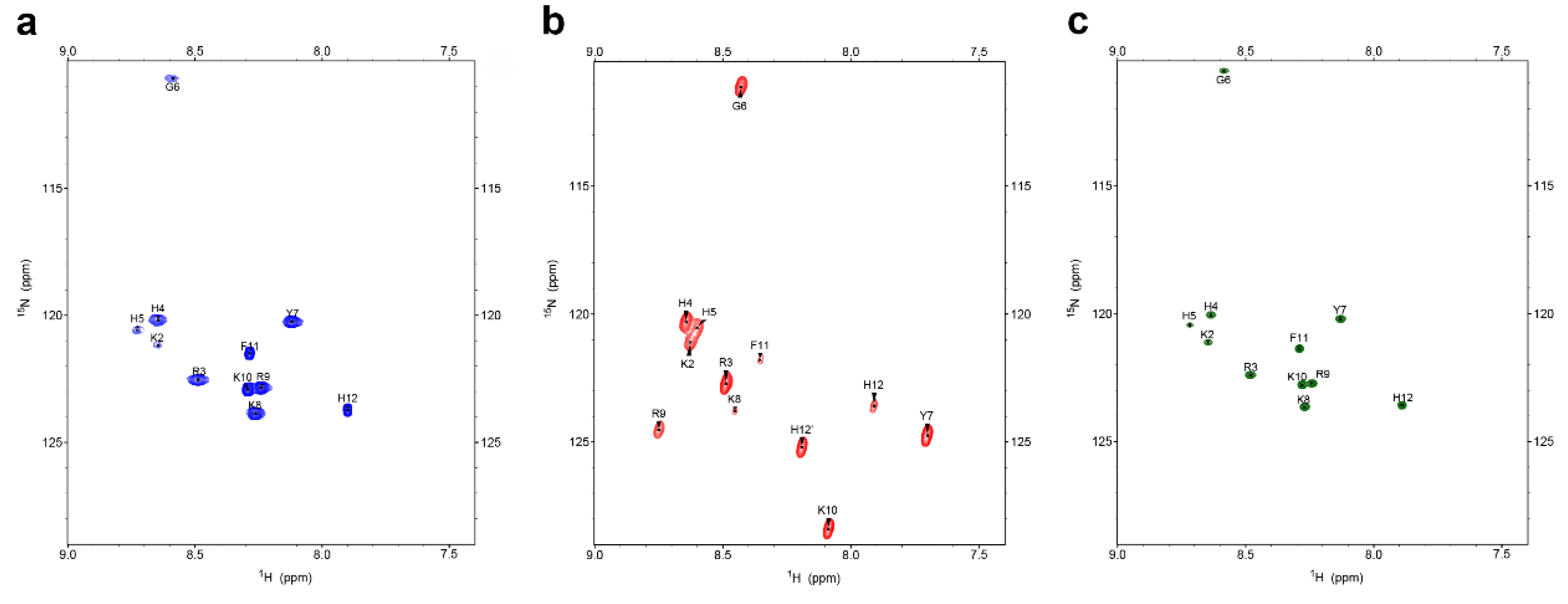
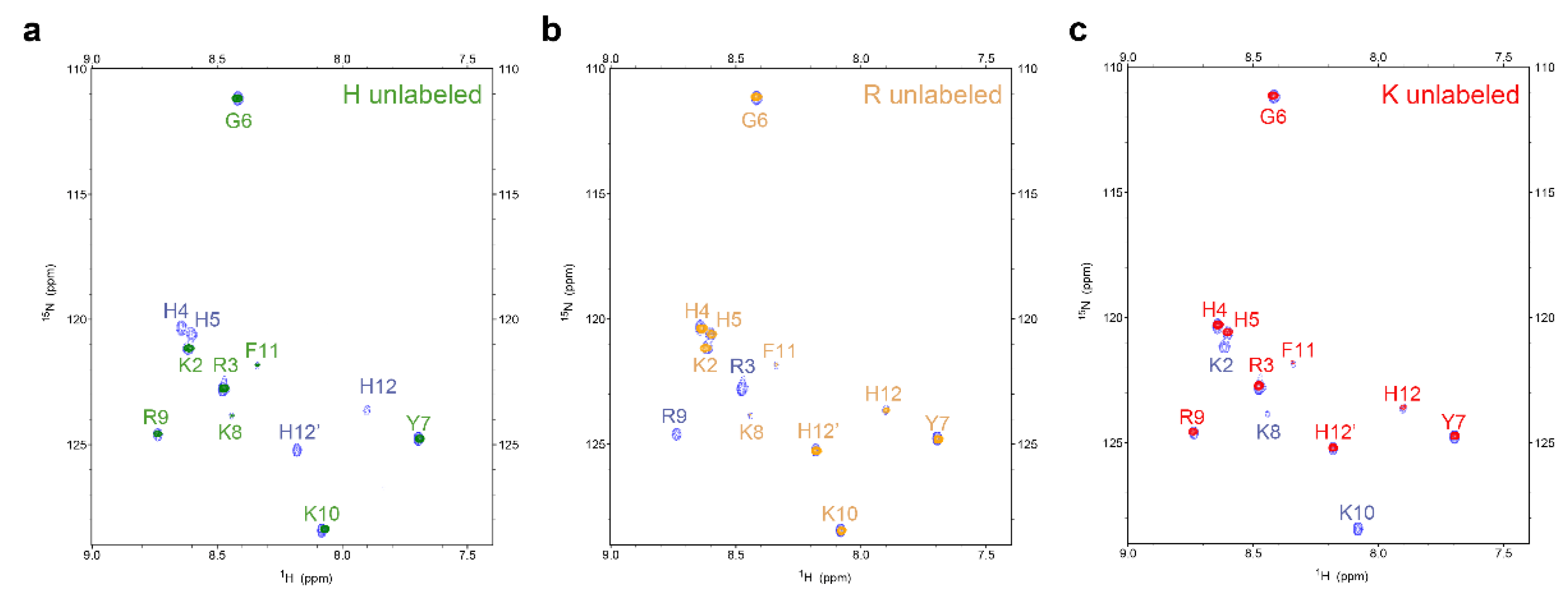
2.1. Interactions with C. albicans Causes Chemical Shift Changes in P-113 over the Course of 24 Hours
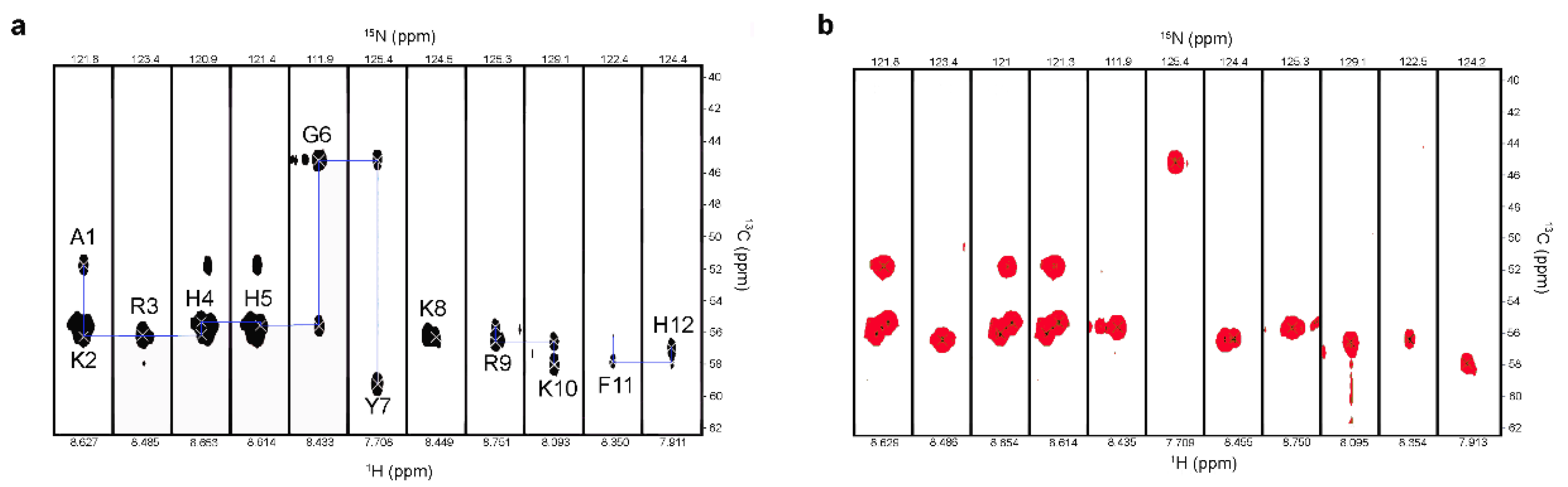
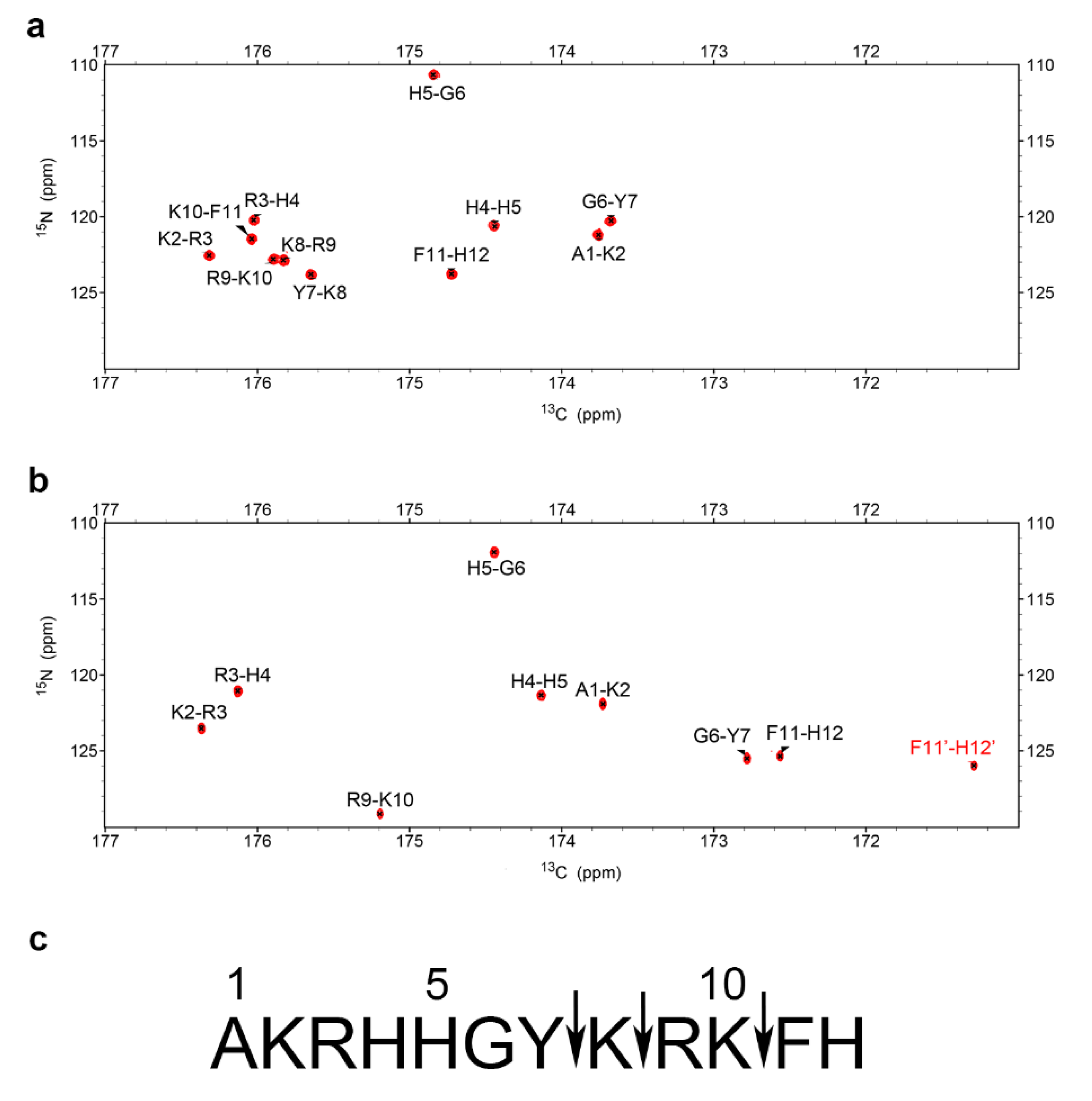
2.2. Characterization of P-113 Degradation Fragments by NMR
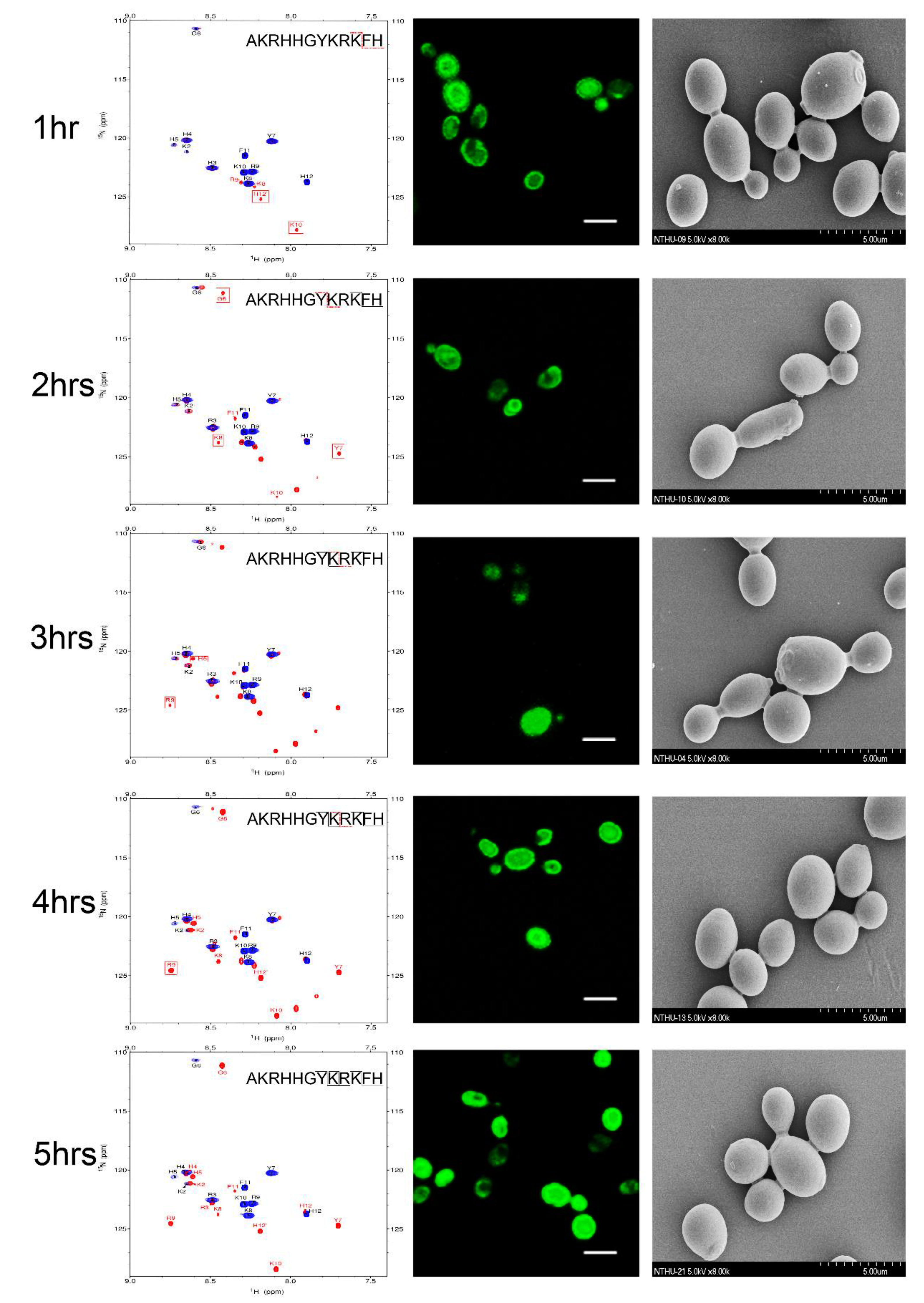
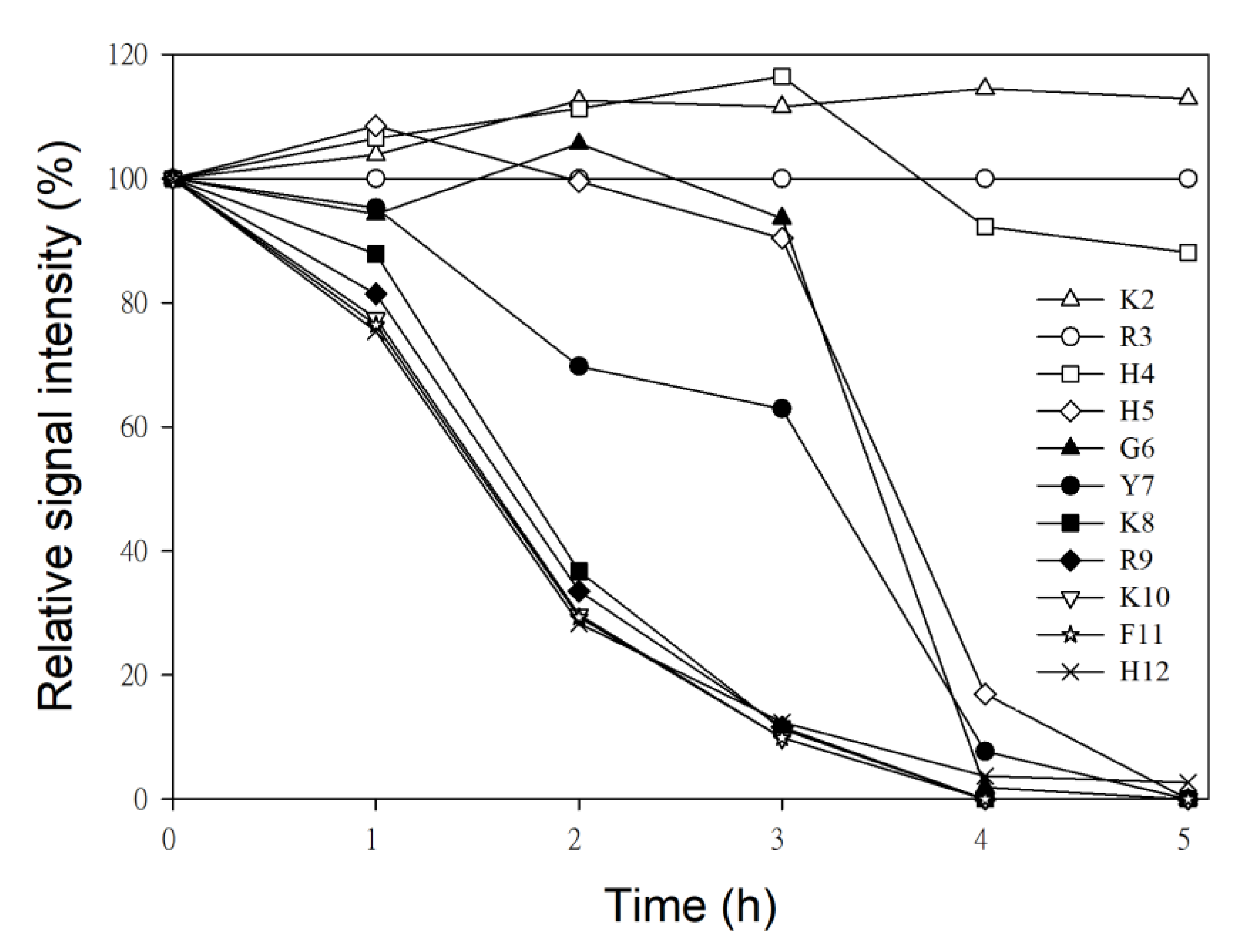
2.3. NMR Time Course Study of P-113 Degradation by C. albicans
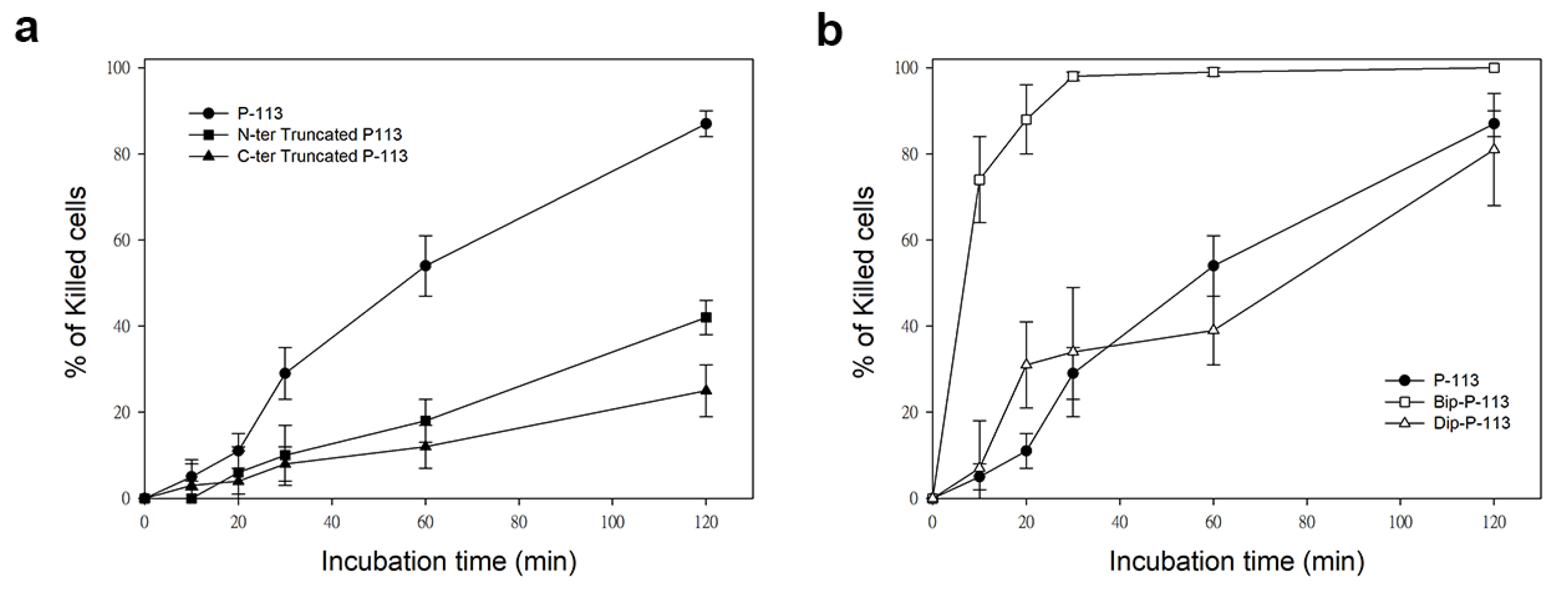
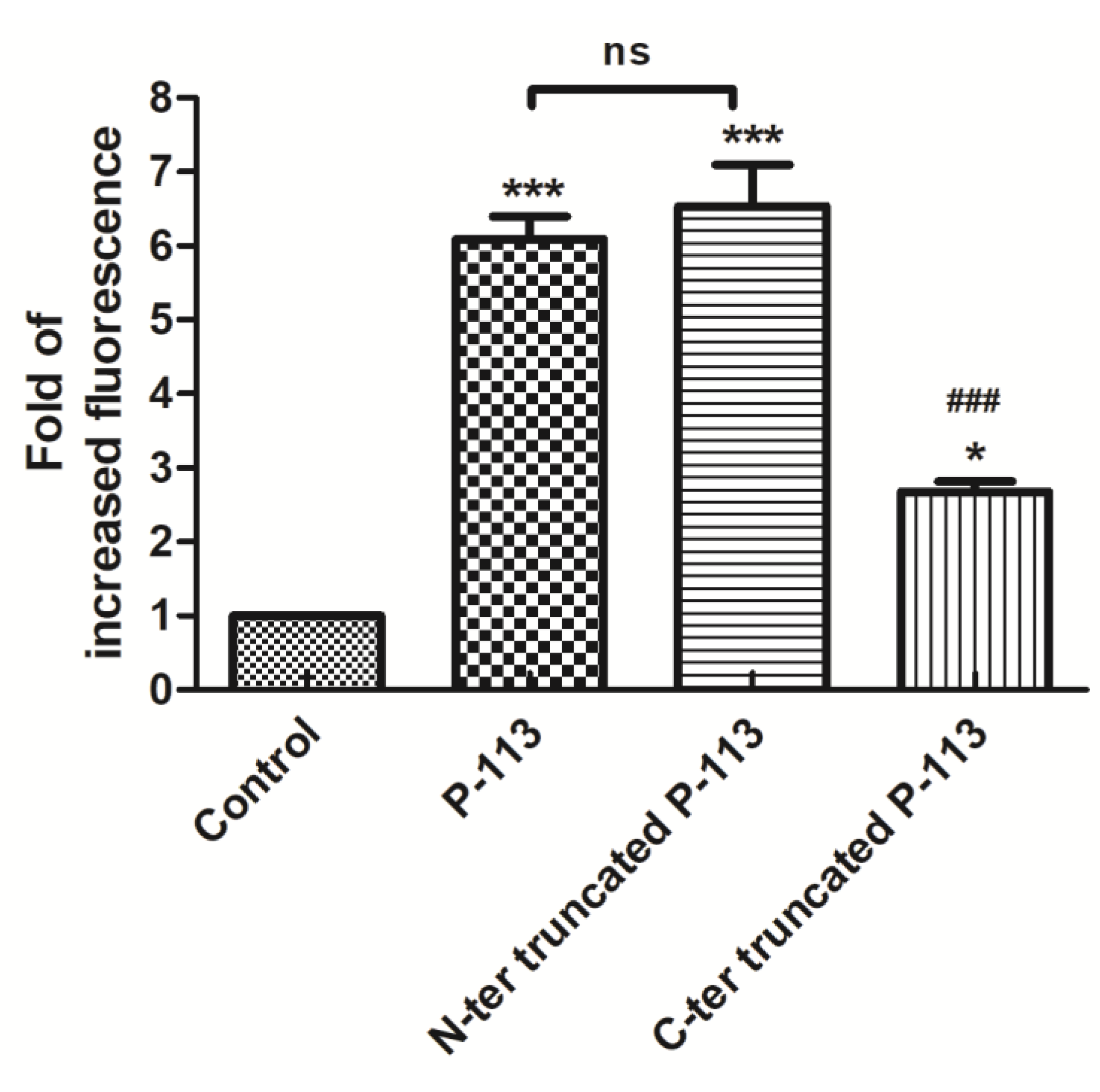
2.4. Functional Role of N- and C-Terminal Amino Acids in the Antifungal Activity of P-113
2.5. Substituting His 4, 5, and 12 of P-113 for Non-Natural Bulky Amino Acids Altered the Anti-Candida Mechanism
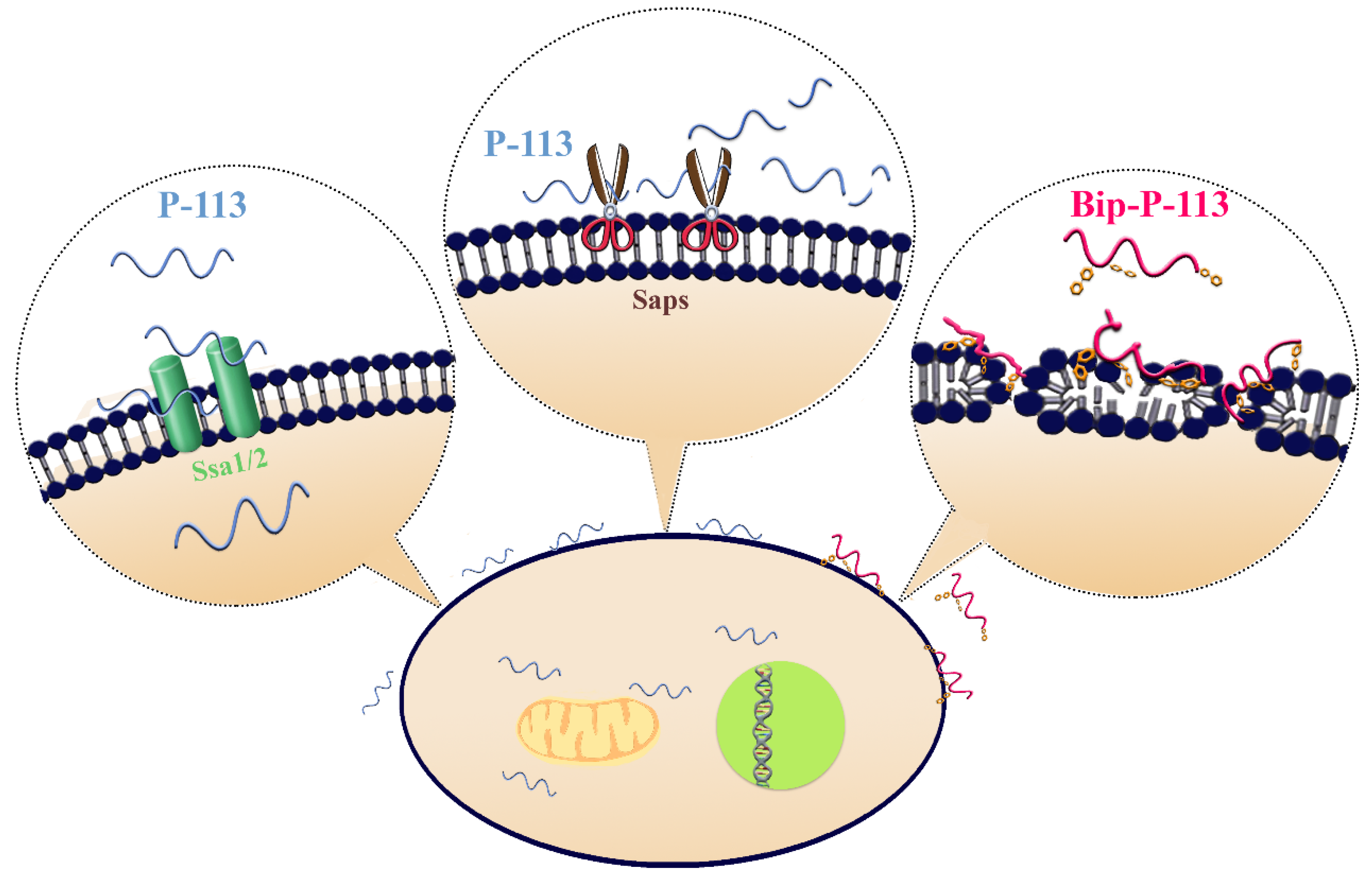
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Peptides
4.3. Yeast Strain
4.4. Nuclear Magnetic Resonance Spectroscopy
4.5. Inhibition of Degradation by Aspartyl Protease Inhibitors
4.6. Expression and Purification of Amino-Acid-Selective 15N-Unlabeled P-113
4.7. Scanning Electron Microscopy Analysis
4.8. Confocal Laser Microscopy
4.9. Anticandidal Activity Assay
4.10. Time Killing Assay
4.11. Fluorescence Microscopy
4.12. Flow Cytometry
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beck-Sague, C.; Jarvis, W.R. Secular trends in the epidemiology of nosocomial fungal infections in the United States, 1980–1990. National Nosocomial Infections Surveillance System. J. Infect. Dis. 1993, 167, 1247–1251. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Sudbery, P. Candida albicans, a major human fungal pathogen. J. Microbiol. 2011, 49, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C.; Jacobsen, M.D. Multilocus sequence typing of pathogenic Candida species. Eukaryot. Cell 2008, 7, 1075–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothstein, D.M.; Spacciapoli, P.; Tran, L.T.; Xu, T.; Roberts, F.D.; Dalla Serra, M.; Buxton, D.K.; Oppenheim, F.G.; Friden, P. Anticandida activity is retained in P-113, a 12-amino-acid fragment of histatin 5. Antimicrob. Agents Chemother. 2001, 45, 1367–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajjan, U.S.; Tran, L.T.; Sole, N.; Rovaldi, C.; Akiyama, A.; Friden, P.M.; Forstner, J.F.; Rothstein, D.M. P-113D, an antimicrobial peptide active against Pseudomonas aeruginosa, retains activity in the presence of sputum from cystic fibrosis patients. Antimicrob. Agents Chemother. 2001, 45, 3437–3444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmerhorst, E.J.; Oppenheim, F.G.; Choi, L.; Cheng, J.W.; Reiner, N.E. Evaluation of a new host-derived synthetic antifungal peptide (PAC-113) in the treatment of oral candidiasis. In Proceedings of the International Meeting on Antimicrobial Chemotherapy in Clinical Practice (ACCP), Portofino, Italy, 15–17 November 2007. [Google Scholar]
- Mickels, N.; McManus, C.; Massaro, J.; Friden, P.; Braman, V.; D’Agostino, R.; Oppenheim, F.; Warbington, M.; Dibart, S.; Van Dyke, T. Clinical and microbial evaluation of a histatin-containing mouthrinse in humans with experimental gingivitis. J. Clin. Periodontol. 2001, 28, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.S.; Li, X.S.; Sun, J.N.; Edgerton, M. The P-113 fragment of histatin 5 requires a specific peptide sequence for intracellular translocation in Candida albicans, which is independent of cell wall binding. Antimicrob. Agents Chemother. 2008, 52, 497–504. [Google Scholar] [CrossRef] [Green Version]
- Georgopoulos, C.; Welch, W.J. Role of the major heat shock proteins as molecular chaperones. Annu. Rev. Cell Biol. 1993, 9, 601–634. [Google Scholar] [CrossRef]
- Bromuro, C.; La Valle, R.; Sandini, S.; Urbani, F.; Ausiello, C.M.; Morelli, L.; Fe d’Ostiani, C.; Romani, L.; Cassone, A. A 70-kilodalton recombinant heat shock protein of Candida albicans is highly immunogenic and enhances systemic murine candidiasis. Ifect. Immun. 1998, 66, 2154–2162. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.Y.; Tu, C.H.; Yip, B.S.; Chen, H.L.; Cheng, H.T.; Huang, K.C.; Lo, H.J.; Cheng, J.W. Easy strategy to increase salt resistance of antimicrobial peptides. Antimicrob. Agents Chemother. 2011, 55, 4918–4921. [Google Scholar] [CrossRef] [Green Version]
- Cutler, J.E. Putative virulence factors of Candida albicans. Annu. Rev. Microbiol. 1991, 45, 187–218. [Google Scholar] [CrossRef] [PubMed]
- Naglik, J.R.; Challacombe, S.J.; Hube, B. Candida albicans secreted aspartyl proteinases in virulence and pathogenesis. Microbiol. Mol. Biol. Rev. 2003, 67, 400–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albrecht, A.; Felk, A.; Pichova, I.; Naglik, J.R.; Schaller, M.; de Groot, P.; MacCallum, D.; Odds, F.C.; Schafer, W.; Klis, F.; et al. Glycosylphosphatidylinositol-anchored proteases of Candida albicans target proteins necessary for both cellular processes and host-pathogen interactions. J. Biol. Chem. 2006, 281, 688–694. [Google Scholar] [CrossRef] [Green Version]
- Hube, B. Candida albicans secreted aspartyl proteinases. Curr. Top. Med. Mycol. 1996, 7, 55–69. [Google Scholar] [PubMed]
- Rapala-Kozik, M.; Bochenska, O.; Zawrotniak, M.; Wolak, N.; Trebacz, G.; Gogol, M.; Ostrowska, D.; Aoki, W.; Ueda, M.; Kozik, A. Inactivation of the antifungal and immunomodulatory properties of human cathelicidin LL-37 by aspartic proteases produced by the pathogenic yeast Candida albicans. Infect. Immun. 2015, 83, 2518–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochenska, O.; Rapala-Kozik, M.; Wolak, N.; Kamysz, W.; Grzywacz, D.; Aoki, W.; Ueda, M.; Kozik, A. Inactivation of human kininogen-derived antimicrobial peptides by secreted aspartic proteases produced by the pathogenic yeast Candida albicans. Biol. Chem. 2015, 396, 1369–1375. [Google Scholar] [CrossRef]
- Meiller, T.F.; Hube, B.; Schild, L.; Shirtliff, M.E.; Scheper, M.A.; Winkler, R.; Ton, A.; Jabra-Rizk, M.A. A novel immune evasion strategy of candida albicans: Proteolytic cleavage of a salivary antimicrobial peptide. PLoS ONE 2009, 4, e5039. [Google Scholar] [CrossRef] [Green Version]
- Bochenska, O.; Rapala-Kozik, M.; Wolak, N.; Aoki, W.; Ueda, M.; Kozik, A. The action of ten secreted aspartic proteases of pathogenic yeast Candida albicans on major human salivary antimicrobial peptide, histatin 5. Acta Biochim. Pol. 2016, 63, 403–410. [Google Scholar] [CrossRef] [Green Version]
- Di Grazia, A.; Cappiello, F.; Cohen, H.; Casciaro, B.; Luca, V.; Pini, A.; Di, Y.P.; Shai, Y.; Mangoni, M.L. D-Amino acids incorporation in the frog skin-derived peptide esculentin-1a(1-21)NH2 is beneficial for its multiple functions. Amino Acids 2015, 47, 2505–2519. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Samperio, P. Peptidomimetics as a new generation of antimicrobial agents: Current progress. Infect. Drug Resist. 2014, 7, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Ikonomova, S.P.; Moghaddam-Taaheri, P.; Jabra-Rizk, M.A.; Wang, Y.; Karlsson, A.J. Engineering improved variants of the antifungal peptide histatin 5 with reduced susceptibility to Candida albicans secreted aspartic proteases and enhanced antimicrobial potency. FEBS J. 2018, 285, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Fidel PL, Jr. Candida-host interactions in HIV disease: Relationships in oropharyngeal candidiasis. Adv. Dent. Res. 2006, 19, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, N.C.; Nery, J.M.; Dias, A.L. Aspartic proteinases of Candida spp.: Role in pathogenicity and antifungal resistance. Mycoses 2014, 57, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gropp, K.; Schild, L.; Schindler, S.; Hube, B.; Zipfel, P.F.; Skerka, C. The yeast Candida albicans evades human complement attack by secretion of aspartic proteases. Mol. Immunol. 2009, 47, 465–475. [Google Scholar] [CrossRef] [PubMed]
- McKendrick, A.J.; Wilson, M.I.; Main, D.M. Oral Candida and long-term tetracycline therapy. Arch. Oral Biol. 1967, 12, 281–290. [Google Scholar] [CrossRef]
- Samaranayake, L.P.; MacFarlane, T.W.; Lamey, P.J.; Ferguson, M.M. A comparison of oral rinse and imprint sampling techniques for the detection of yeast, coliform and Staphylococcus aureus carriage in the oral cavity. J. Oral Pathol. 1986, 15, 386–388. [Google Scholar] [CrossRef]
- Van Dyke, T.; Paquette, D.; Grossi, S.; Braman, V.; Massaro, J.; D’Agostino, R.; Dibart, S.; Friden, P. Clinical and microbial evaluation of a histatin-containing mouthrinse in humans with experimental gingivitis: A phase-2 multi-center study. J. Clin. Periodontol. 2002, 29, 168–176. [Google Scholar] [CrossRef]
- Han, J.; Jyoti, M.A.; Song, H.Y.; Jang, W.S. Antifungal activity and action mechanism of histatin 5-halocidin hybrid peptides against Candida ssp. PLoS ONE 2016, 11, e0150196. [Google Scholar] [CrossRef] [Green Version]
- Chih, Y.H.; Wang, S.Y.; Yip, B.S.; Cheng, K.T.; Hsu, S.Y.; Wu, C.L.; Yu, H.Y.; Cheng, J.W. Dependence on size and shape of non-nature amino acids in the enhancement of lipopolysaccharide (LPS) neutralizing activities of antimicrobial peptides. J. Colloid Interface Sci. 2019, 533, 492–502. [Google Scholar] [CrossRef]
- Cheng, K.T.; Wu, C.L.; Yip, B.S.; Yu, H.Y.; Cheng, H.T.; Chih, Y.H.; Cheng, J.W. High level expression and purification of the clinically active antimicrobial peptide P-113 in Escherichia coli. Molecules 2018, 23, 800. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | Molecular Weight (Da) | MH Broth | LYM | ||
|---|---|---|---|---|---|---|
| Control | 50 mM NaCl | 100 mM NaCl | ||||
| P-113 * | FITC-AKRHHGYKRKFH-NH2 | 2066.36 | >50 | 3.13 | 25 | 50 |
| N-ter truncated P-113 | FITC-RHHGYKRKFH-NH2 | 1867.14 | >50 | 25 | >50 | >50 |
| C-ter truncated P-113 | FITC-AKRHHGYKRK-NH2 | 1781.49 | >50 | 50 | >50 | >50 |
| Bip-P-113 | FITC-AKR(Bip)(Bip)GYKRKF(Bip)-NH2 | 2324.62 | 50 | 6.25 | 6.25 | 12.5 |
| Dip-P-113 | FITC-AKR(Dip)(Dip)GYKRKF(Dip)-NH2 | 2324.62 | >50 | 12.5 | 12.5 | 25 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, K.-T.; Wu, C.-L.; Yip, B.-S.; Chih, Y.-H.; Peng, K.-L.; Hsu, S.-Y.; Yu, H.-Y.; Cheng, J.-W. The Interactions between the Antimicrobial Peptide P-113 and Living Candida albicans Cells Shed Light on Mechanisms of Antifungal Activity and Resistance. Int. J. Mol. Sci. 2020, 21, 2654. https://doi.org/10.3390/ijms21072654
Cheng K-T, Wu C-L, Yip B-S, Chih Y-H, Peng K-L, Hsu S-Y, Yu H-Y, Cheng J-W. The Interactions between the Antimicrobial Peptide P-113 and Living Candida albicans Cells Shed Light on Mechanisms of Antifungal Activity and Resistance. International Journal of Molecular Sciences. 2020; 21(7):2654. https://doi.org/10.3390/ijms21072654
Chicago/Turabian StyleCheng, Kuang-Ting, Chih-Lung Wu, Bak-Sau Yip, Ya-Han Chih, Kuang-Li Peng, Su-Ya Hsu, Hui-Yuan Yu, and Jya-Wei Cheng. 2020. "The Interactions between the Antimicrobial Peptide P-113 and Living Candida albicans Cells Shed Light on Mechanisms of Antifungal Activity and Resistance" International Journal of Molecular Sciences 21, no. 7: 2654. https://doi.org/10.3390/ijms21072654
APA StyleCheng, K. -T., Wu, C. -L., Yip, B. -S., Chih, Y. -H., Peng, K. -L., Hsu, S. -Y., Yu, H. -Y., & Cheng, J. -W. (2020). The Interactions between the Antimicrobial Peptide P-113 and Living Candida albicans Cells Shed Light on Mechanisms of Antifungal Activity and Resistance. International Journal of Molecular Sciences, 21(7), 2654. https://doi.org/10.3390/ijms21072654