Dynamics Studies of DNA with Non-canonical Structure Using NMR Spectroscopy


{kind=link}
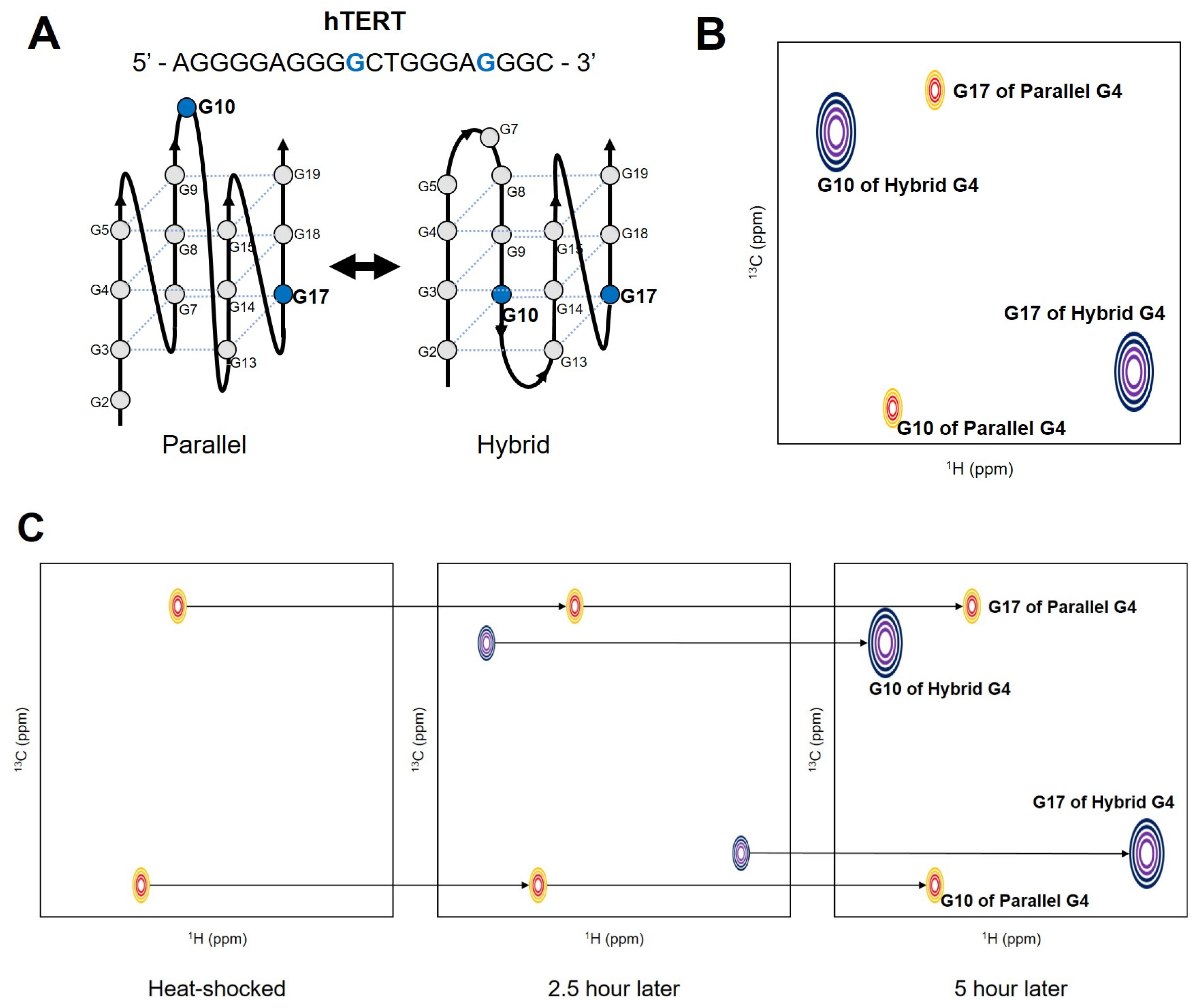{kind=link}
{kind=link}
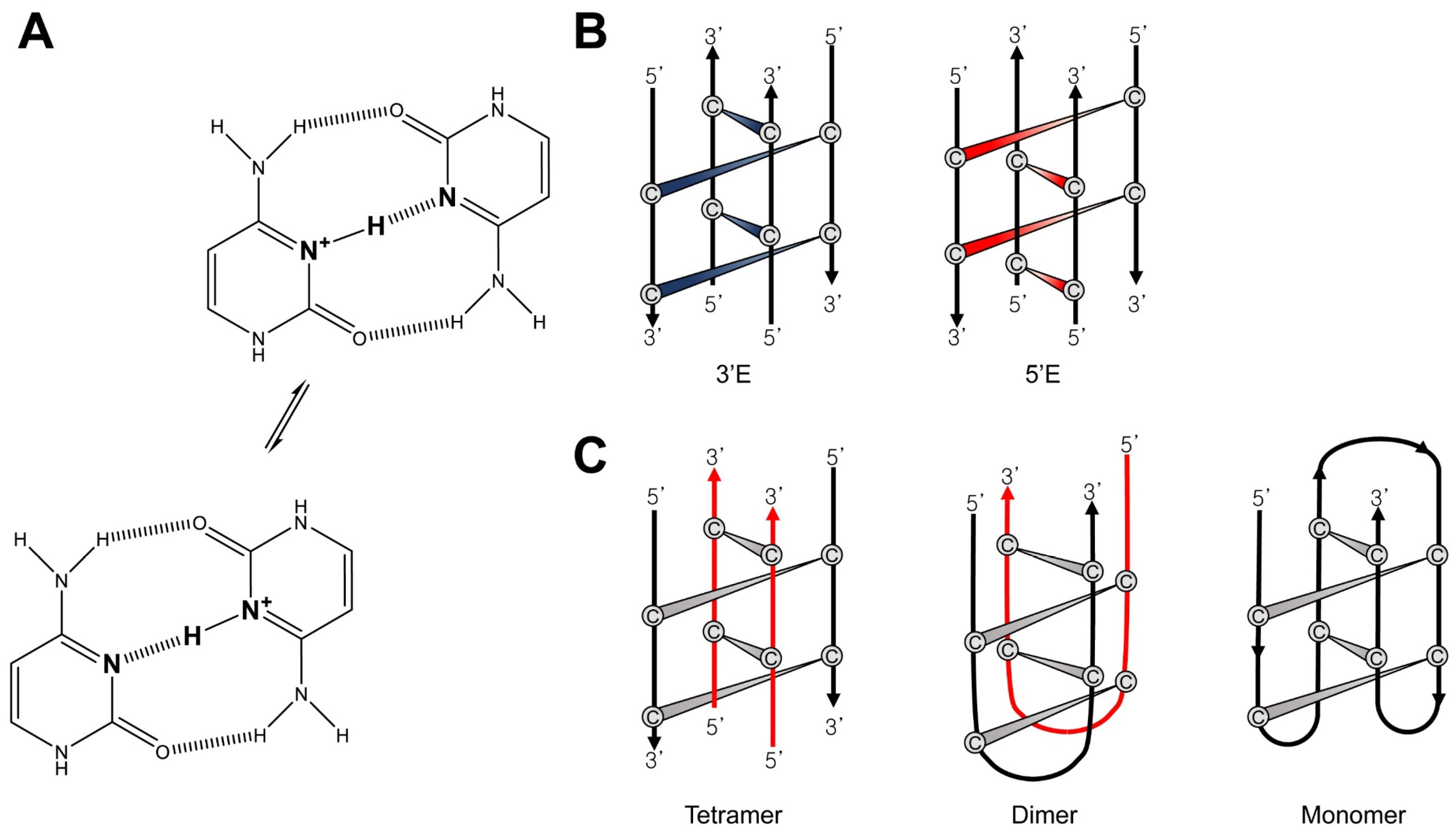{kind=link}
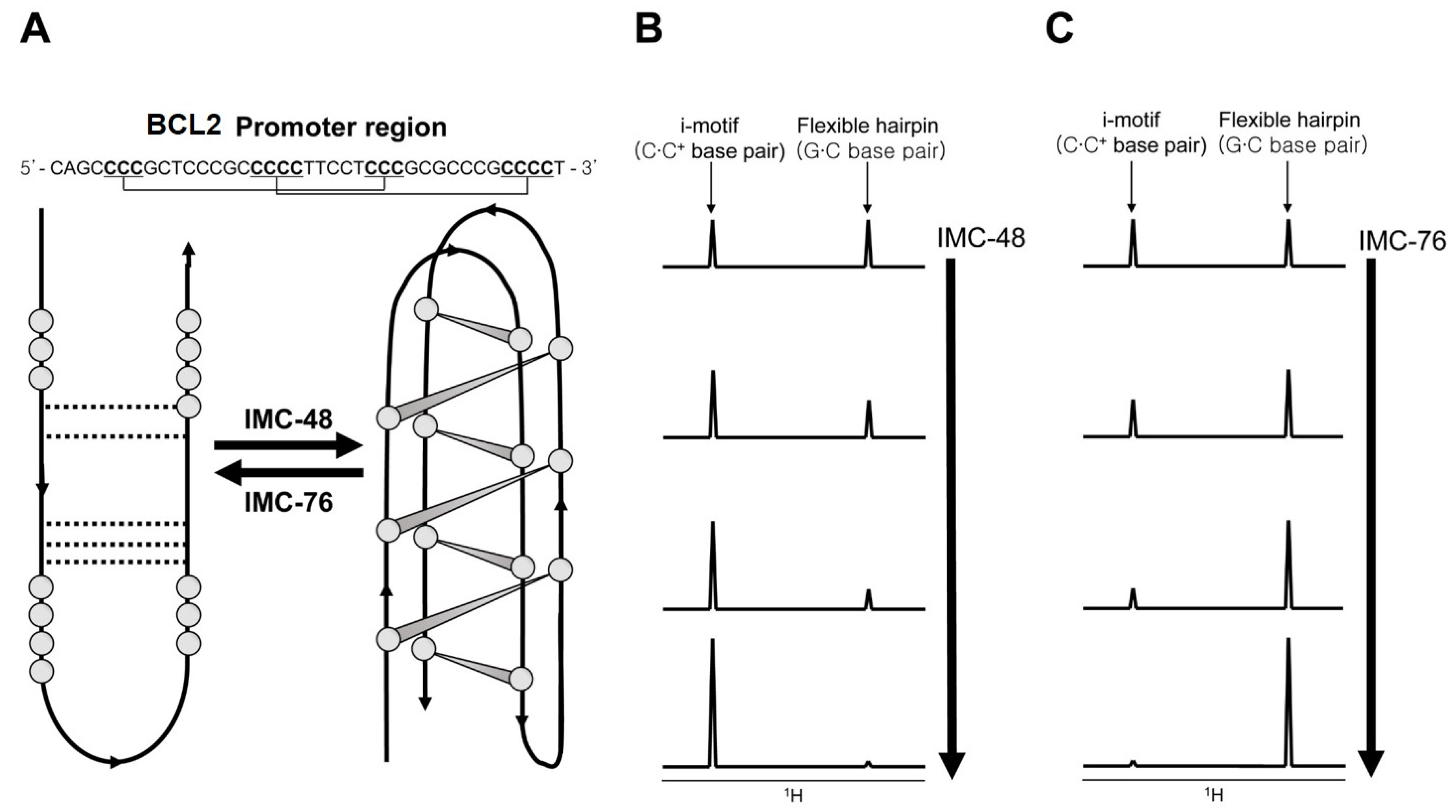{kind=link}
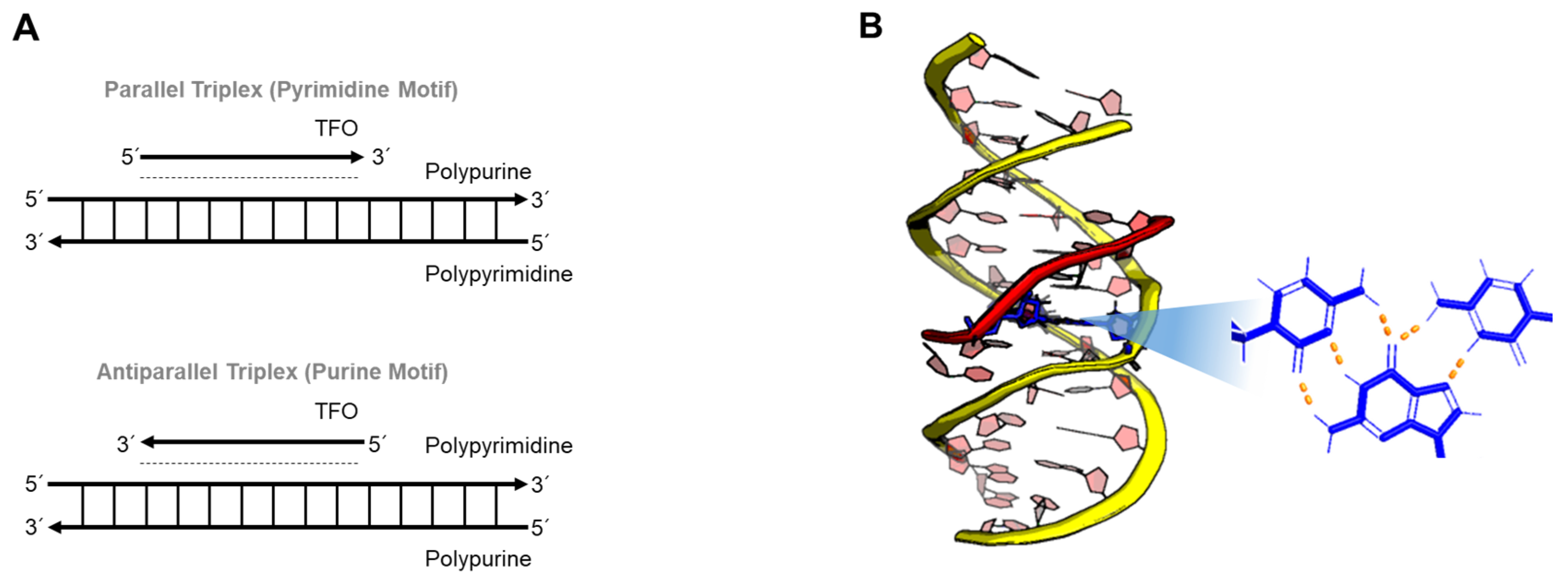{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. G-Quadruplex (G4)
2.1. G4 DNA
2.2. G4-Protein Interaction
2.3. G4-Ligand Interaction
3. I-Motif
3.1. I-Motif DNA
3.2. I-Motif Binding Ligands and Proteins
4. DNA Triplex
5. Z-DNA
5.1. Z-DNA
5.2. Z-DNA Complexes with ZBPs
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| G4 | G-quadruplex |
| BEST | Band-selective Excitation Short-Transient |
| HMGB1 | High Mobility Group B1 |
| RGG3 | The third RGG motif |
| TLS | Translocated in liposarcoma |
| FUS | Fused in sarcoma |
| TERRA | Telomeric repeat-containing RNA |
| CSP | The chemical shift perturbation |
| CD | Circular dichroism |
| WRN | Werner syndrome protein |
| BLM | Bloom syndrome protein |
| RQC | RecQ C-terminal |
| CPMG | Car-Purcell-Meiboom-Gill |
| BMVC-8C3O | 3,6-bis(1-methyl-4-vinylpyridiumiodide)-9-(1-(1-methyl-piperidiniumiodide)-3,6,9-trioxaundecane |
| BTC | Bis-triazolylcarbazole |
| smFRET | single-molecule Förster resonance energy transfer |
| C∙C+ | Cytidine and protonated cytidine base-pair |
| BCL2 | Human B-cell lymphoma gene-2 |
| RRM12 | RRM1 and RRM2 domains |
| TFO | Triplex-forming oligonucleotide |
| ZBPs | Z-DNA binding proteins |
| DAI | DNA-dependent activator of IFN-regulatory factor |
| PKZ | ZBP-containing protein kinase |
| hZαADAR1 | Zα domain of human ADAR1 |
| yabZαE3L | Zα domain of yatapoxvirus E3L |
| caZαPKZ | Zα domain of PKZ from Carassius auratus |
References
- Gallie, D.R. A tale of two termini: A functional interaction between the termini of an mRNA is a prerequisite for efficient translation initiation. Gene 1998, 216, 1–11. [Google Scholar] [CrossRef]
- Yagil, G. Paranemic structures of DNA and their role in DNA unwinding. Crit. Rev. Biochem. Mol. Biol. 1991, 26, 475–559. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H. Switching and signaling at the telomere. Cell 2001, 106, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Aubert, G.; Lansdorp, P.M. Telomeres and aging. Physiol. Rev. 2008, 88, 557–579. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, L.; Mergny, J.-L.; Leroy, J.-L.; Hélène, C. Inability of RNA to form the i-motif: implications for triplex formation. Biochemistry 1996, 35, 8715–8722. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Majima, T. Conformational changes of non-B DNA. Chem. Soc. Rev. 2011, 40, 5893–5909. [Google Scholar] [CrossRef]
- Wang, G.; Vasquez, K.M. Impact of alternative DNA structures on DNA damage, DNA repair, and genetic instability. DNA Repair 2014, 19, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Halder, S.; Bhattacharyya, D. RNA structure and dynamics: A base pairing perspective. Prog. Biophys. Mol. Biol. 2013, 113, 264–283. [Google Scholar] [CrossRef]
- Heus, H.A.; Hilbers, C.W. Structures of non-canonical tandem base pairs in RNA helices: Review. Nucleosides Nucleotides Nucleic Acids 2003, 22, 559–571. [Google Scholar] [CrossRef]
- Uhlenbeck, O.C.; Pardi, A.; Feigon, J. RNA structure comes of age. Cell 1997, 90, 833–840. [Google Scholar] [CrossRef] [Green Version]
- Wöhnert, J.; Dingley, A.J.; Stoldt, M.; Görlach, M.; Grzesiek, S.; Brown, L.R. Direct identification of NH...N hydrogen bonds in non-canonical base pairs of RNA by NMR spectroscopy. Nucleic Acids Res. 1999, 27, 3104–3110. [Google Scholar]
- Rachwal, P.A.; Brown, T.; Fox, K.R. Effect of G-tract length on the topology and stability of intramolecular DNA quadruplexes. Biochemistry 2007, 46, 3036–3044. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.T. G-quartets 40 years later: From 5’-GMP to molecular biology and supramolecular chemistry. Angew. Chem. Int. Ed. 2004, 43, 668–698. [Google Scholar] [CrossRef]
- Wright, E.P.; Huppert, J.L.; Waller, Z.A.E. Identification of multiple genomic DNA sequences which form i-motif structures at neutral pH. Nucleic Acids Res. 2017, 45, 2951–2959. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.M.; Ding, Y.; Rogers, R.A.; Zhu, J.; Zhu, J.; Burton, A.D.; Carlisle, C.B.; Burrows, C.J. 4n–1 is a “Sweet Spot” in DNA i-motif folding of 2′-deoxycytidine homopolymers. J. Am. Chem. Soc. 2017, 139, 4682–4689. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Parkinson, G.N.; Neidle, S. Crystal structure of the potassium form of an Oxytricha nova G-quadruplex. J. Mol. Biol. 2002, 320, 189–200. [Google Scholar] [CrossRef]
- Smirnov, I.; Shafer, R.H. Effect of loop sequence and size on DNA aptamer stability. Biochemistry 2000, 39, 1462–1468. [Google Scholar] [CrossRef]
- Asamitsu, S.; Obata, S.; Yu, Z.; Bando, T.; Sugiyama, H. Recent progress of targeted G-quadruplex-preferred ligands toward cancer therapy. Molecules 2019, 24, 429. [Google Scholar] [CrossRef] [Green Version]
- De Magis, A.; Manzo, S.G.; Russo, M.; Marinello, J.; Morigi, R.; Sordet, O.; Capranico, G. DNA damage and genome instability by G-quadruplex ligands are mediated by R loops in human cancer cells. Proc. Natl. Acad. Sci. USA 2019, 116, 816. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, J.; Adhikari, S.; Balasubramanian, S. The structure and function of DNA G-quadruplexes. Trends Chem. 2020, 2, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Bessi, I.; Jonker, H.R.A.; Richter, C.; Schwalbe, H. Involvement of long-lived intermediate states in the complex folding pathway of the human telomeric G-quadruplex. Angew. Chem. Int. Ed. 2015, 54, 8444–8448. [Google Scholar] [CrossRef] [PubMed]
- Harkness, R.W.V.; Mittermaier, A.K. G-register exchange dynamics in guanine quadruplexes. Nucleic Acids Res. 2016, 44, 3481–3494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, N.Q.; Phan, A.T. Monomer–dimer equilibrium for the 5′–5′ stacking of propeller-type parallel-stranded G-quadruplexes: NMR structural study. Chem. Eur. J. 2012, 18, 14752–14759. [Google Scholar] [CrossRef]
- Seenisamy, J.; Rezler, E.M.; Powell, T.J.; Tye, D.; Gokhale, V.; Joshi, C.S.; Siddiqui-Jain, A.; Hurley, L.H. The dynamic character of the G-quadruplex element in the c-MYC promoter and modification by TMPyP4. J. Am. Chem. Soc. 2004, 126, 8702–8709. [Google Scholar] [CrossRef] [PubMed]
- Adrian, M.; Heddi, B.; Phan, A.T. NMR spectroscopy of G-quadruplexes. Methods 2012, 57, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Harkness, R.W.; Mittermaier, A.K. G-quadruplex dynamics. Biochim. Biophys. Acta - Proteins Proteom 2017, 1865, 1544–1554. [Google Scholar] [CrossRef]
- Nußbaumer, F.; Juen, M.A.; Gasser, C.; Kremser, J.; Müller, T.; Tollinger, M.; Kreutz, C. Synthesis and incorporation of 13C-labeled DNA building blocks to probe structural dynamics of DNA by NMR. Nucleic Acids Res. 2017, 45, 9178–9192. [Google Scholar] [CrossRef] [Green Version]
- Guéron, M.; Kochoyan, M.; Leroy, J.-L. A single mode of DNA base-pair opening drives imino proton exchange. Nature 1987, 328, 89–92. [Google Scholar] [CrossRef]
- Guéron, M.; Leroy, J.L. Studies of base pair kinetics by NMR measurement of proton exchange. Methods Enzymol. 1995, 261, 383–413. [Google Scholar]
- Russu, I.M. Probing site-specific energetics in proteins and nucleic acids by hydrogen exchange and nuclear magnetic resonance spectroscopy. Methods Enzymol. 2004, 379, 152–175. [Google Scholar]
- Krueger, A.; Protozanova, E.; Frank-Kamenetskii, M.D. Sequence-dependent base pair opening in DNA double helix. Biophys. J. 2006, 90, 3091–3099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolova, E.N.; Zhou, H.; Gottardo, F.L.; Alvey, H.S.; Kimsey, I.J.; Al-Hashimi, H.M. A historical account of Hoogsteen base-pairs in duplex DNA. Biopolymers 2013, 99, 955–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.-R.; Kim, N.-H.; Jin, H.-S.; Seo, Y.-J.; Lee, J.; Lee, J.-H. Base-pair opening dynamics of nucleic acids in relation to their biological function. Comput. Struct. Biotechnol. J. 2019, 17, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Grün, J.T.; Hennecker, C.; Klötzner, D.-P.; Harkness, R.W.; Bessi, I.; Heckel, A.; Mittermaier, A.K.; Schwalbe, H. Conformational dynamics of strand register shifts in DNA G-quadruplexes. J. Am. Chem. Soc. 2020, 142, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Lescop, E.; Kern, T.; Brutscher, B. Guidelines for the use of band-selective radiofrequency pulses in hetero-nuclear NMR: Example of longitudinal-relaxation-enhanced BEST-type 1H–15N correlation experiments. J. Magn. Reson. 2010, 203, 190–198. [Google Scholar] [CrossRef]
- Brcic, J.; Plavec, J. NMR structure of a G-quadruplex formed by four d(G4C2) repeats: Insights into structural polymorphism. Nucleic Acids Res. 2018, 46, 11605–11617. [Google Scholar]
- DeJesus-Hernandez, M.; Mackenzie, I.R.; Boeve, B.F.; Boxer, A.L.; Baker, M.; Rutherford, N.J.; Nicholson, A.M.; Finch, N.A.; Flynn, H.; Adamson, J.; et al. Expanded GGGGCC hexanucleotide repeat in noncoding region of C9ORF72 causes chromosome 9p-linked FTD and ALS. Neuron 2011, 72, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Renton, A.E.; Majounie, E.; Waite, A.; Simón-Sánchez, J.; Rollinson, S.; Gibbs, J.R.; Schymick, J.C.; Laaksovirta, H.; Van Swieten, J.C.; Myllykangas, L.; et al. A hexanucleotide repeat expansion in C9ORF72 is the cause of chromosome 9p21-linked ALS-FTD. Neuron 2011, 72, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Varizhuk, A.M.; Protopopova, A.D.; Tsvetkov, V.B.; Barinov, N.A.; Podgorsky, V.V.; Tankevich, M.V.; Vlasenok, M.A.; Severov, V.V.; Smirnov, I.P.; Dubrovin, E.V.; et al. Polymorphism of G4 associates: From stacks to wires via interlocks. Nucleic Acids Res. 2018, 46, 8978–8992. [Google Scholar] [CrossRef]
- Mendoza, O.; Bourdoncle, A.; Boulé, J.-B.; Brosh, R.M., Jr.; Mergny, J.-L. G-quadruplexes and helicases. Nucleic Acids Res. 2016, 44, 1989–2006. [Google Scholar] [CrossRef] [Green Version]
- Sauer, M.; Paeschke, K. G-quadruplex unwinding helicases and their function in vivo. Biochem. Soc. Trans. 2017, 45, 1173–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brázda, V.; Hároníková, L.; Liao, J.C.C.; Fojta, M. DNA and RNA quadruplex-binding proteins. Int. J. Mol. Sci 2014, 15, 17493–17517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüger, A.C.; Raarup, M.K.; Nielsen, M.M.; Kristensen, M.; Besenbacher, F.; Kjems, J.; Birkedal, V. Interaction of hnRNP A1 with telomere DNA G-quadruplex structures studied at the single molecule level. Eur. Biophys. J. 2010, 39, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.; Buncher, N.; Opresko, P.L.; Myong, S. POT1-TPP1 regulates telomeric overhang structural dynamics. Structure 2012, 20, 1872–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, J.; Madanayake, T.W.; Iaccarino, N.; Novellino, E.; Randazzo, A.; Hurley, L.H.; Pagano, B. HMGB1 binds to the KRAS promoter G-quadruplex: A new player in oncogene transcriptional regulation? Chem. Commun. 2018, 54, 9442–9445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, J.; Cerofolini, L.; Brancaccio, D.; Giuntini, S.; Iaccarino, N.; Zizza, P.; Iachettini, S.; Biroccio, A.; Novellino, E.; Rosato, A.; et al. Insights into telomeric G-quadruplex DNA recognition by HMGB1 protein. Nucleic Acids Res. 2019, 47, 9950–9966. [Google Scholar] [CrossRef] [Green Version]
- Kondo, K.; Mashima, T.; Oyoshi, T.; Yagi, R.; Kurokawa, R.; Kobayashi, N.; Nagata, T.; Katahira, M. Plastic roles of phenylalanine and tyrosine residues of TLS/FUS in complex formation with the G-quadruplexes of telomeric DNA and TERRA. Sci. Rep. 2018, 8, 2864. [Google Scholar] [CrossRef]
- Ghosh, M.; Singh, M. RGG-box in hnRNPA1 specifically recognizes the telomere G-quadruplex DNA and enhances the G-quadruplex unfolding ability of UP1 domain. Nucleic Acids Res. 2018, 46, 10246–10261. [Google Scholar] [CrossRef]
- Chatterjee, S.; Zagelbaum, J.; Savitsky, P.; Sturzenegger, A.; Huttner, D.; Janscak, P.; Hickson, I.D.; Gileadi, O.; Rothenberg, E. Mechanistic insight into the interaction of BLM helicase with intra-strand G-quadruplex structures. Nat. Commun. 2014, 5, 5556. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.-Q.; Hou, X.-M.; Li, M.; Dou, S.-X.; Xi, X.-G. BLM unfolds G-quadruplexes in different structural environments through different mechanisms. Nucleic Acids Res. 2015, 43, 4614–4626. [Google Scholar] [CrossRef] [Green Version]
- Ketkar, A.; Voehler, M.; Mukiza, T.; Eoff, R.L. Residues in the RecQ C-terminal domain of the human Werner syndrome helicase are involved in unwinding G-quadruplex DNA. J. Biol. Chem. 2017, 292, 3154–3163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Lee, A.-R.; Ryu, K.-S.; Lee, J.-H.; Park, C.-J. NMR investigation of the interaction between the RecQ C-terminal domain of human Bloom syndrome protein and G-quadruplex DNA from the human c-Myc promoter. J. Mol. Biol. 2019, 431, 794–806. [Google Scholar] [CrossRef] [PubMed]
- Kitano, K.; Kim, S.-Y.; Hakoshima, T. Structural basis for DNA strand separation by the unconventional winged-helix domain of RecQ helicase WRN. Structure 2010, 18, 177–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, J.A.; Savitsky, P.; Allerston, C.K.; Bizard, A.H.; Özer, Ö.; Sarlós, K.; Liu, Y.; Pardon, E.; Steyaert, J.; Hickson, I.D.; et al. Crystal structure of the Bloom’s syndrome helicase indicates a role for the HRDC domain in conformational changes. Nucleic Acids Res. 2015, 43, 5221–5235. [Google Scholar] [CrossRef]
- Palmer, A.G.; Kroenke, C.D.; Loria, J.P. Nuclear magnetic resonance methods for quantifying microsecond-to-millisecond motions in biological macromolecules. Methods Enzymol. 2001, 339, 204–238. [Google Scholar]
- Neidle, S. Human telomeric G-quadruplex: The current status of telomeric G-quadruplexes as therapeutic targets in human cancer. FEBS J. 2010, 277, 1118–1125. [Google Scholar] [CrossRef]
- Sun, Z.-Y.; Wang, X.-N.; Cheng, S.-Q.; Su, X.-X.; Ou, T.-M. Developing novel G-quadruplex ligands: From interaction with nucleic acids to interfering with nucleic acid–protein interaction. Molecules 2019, 24, 396. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui-Jain, A.; Grand, C.L.; Bearss, D.J.; Hurley, L.H. Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription. Proc. Natl. Acad. Sci. USA 2002, 99, 11593–11598. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.-J.; Wu, Y.-L.; Tanaka, Y.; Zhang, W. Small molecules targeting c-Myc oncogene: Promising anti-cancer therapeutics. Int. J. Biol. Sci. 2014, 10, 1084–1096. [Google Scholar] [CrossRef]
- Duarte, A.R.; Cadoni, E.; Ressurreição, A.S.; Moreira, R.; Paulo, A. Design of modular G-quadruplex ligands. Chem. Med. Chem. 2018, 13, 869–893. [Google Scholar] [CrossRef]
- Wang, Z.-F.; Li, M.-H.; Chen, W.-W.; Hsu, S.-T.D.; Chang, T.-C. A novel transition pathway of ligand-induced topological conversion from hybrid forms to parallel forms of human telomeric G-quadruplexes. Nucleic Acids Res. 2016, 44, 3958–3968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, M.; Ghosh, S.; Panda, D.; Bessi, I.; Schwalbe, H.; Bhattacharyya, K.; Dash, J. Small molecule regulated dynamic structural changes of human G-quadruplexes. Chem. Sci. 2016, 7, 3279–3285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musso, L.; Mazzini, S.; Rossini, A.; Castagnoli, L.; Scaglioni, L.; Artali, R.; Di Nicola, M.; Zunino, F.; Dallavalle, S. C-MYC G-quadruplex binding by the RNA polymerase I inhibitor BMH-21 and analogues revealed by a combined NMR and biochemical Approach. BBA-GEN SUBJECTS. 2018, 1862, 615–629. [Google Scholar] [CrossRef]
- Dutta, D.; Debnath, M.; Müller, D.; Paul, R.; Das, T.; Bessi, I.; Schwalbe, H.; Dash, J. Cell penetrating thiazole peptides inhibit c-MYC expression via site-specific targeting of c-MYC G-quadruplex. Nucleic Acids Res. 2018, 46, 5355–5365. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Banerjee, N.; Roychowdhury, T.; Dutta, A.; Chattopadhyay, S.; Chatterjee, S. Site-specific amino acid substitution in dodecameric peptides determines the stability and unfolding of c-MYC quadruplex promoting apoptosis in cancer cells. Nucleic Acids Res. 2018, 46, 9932–9950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehring, K.; Leroy, J.-L.; Guéron, M. A tetrameric DNA structure with protonated cytosine-cytosine base pairs. Nature. 1993, 363, 561–565. [Google Scholar] [CrossRef]
- Choi, J.; Kim, S.; Tachikawa, T.; Fujitsuka, M.; Majima, T. pH-induced intramolecular folding dynamics of i-motif DNA. J. Am. Chem. Soc. 2011, 133, 16146–16153. [Google Scholar] [CrossRef]
- Mergny, J.-L.; Lacroix, L.; Han, X.; Leroy, J.-L.; Helene, C. Intramolecular folding of pyrimidine oligodeoxynucleotides into an i-DNA motif. J. Am. Chem. Soc. 1995, 117, 8887–8898. [Google Scholar] [CrossRef]
- Chambers, V.S.; Marsico, G.; Boutell, J.M.; Di Antonio, M.; Smith, G.P.; Balasubramanian, S. High-throughput sequencing of DNA G-quadruplex structures in the human genome. Nat. Biotechnol. 2015, 33, 877–881. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Hurley, L.H. The Importance of Negative Superhelicity in inducing the formation of G-quadruplex and i-motif structures in the c-Myc promoter: Implications for drug targeting and control of gene expression. J. Med. Chem. 2009, 52, 2863–2874. [Google Scholar] [CrossRef] [Green Version]
- Brooks, T.A.; Kendrick, S.; Hurley, L. Making sense of G-quadruplex and i-motif functions in oncogene promoters. FEBS J. 2010, 277, 3459–3469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzatko, S.; Krafcikova, M.; Hänsel-Hertsch, R.; Fessl, T.; Fiala, R.; Loja, T.; Krafcik, D.; Mergny, J.-L.; Foldynova-Trantirkova, S.; Trantirek, L. Evaluation of the stability of DNA i-motifs in the nuclei of living mammalian cells. Angew Chem. Int. Ed. 2018, 57, 2165–2169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, A.T.; Leroy, J.L. Intramolecular i-motif structures of telomeric DNA. J. Biomol. Struct. Dyn. 2000, 17, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Yang, Z.; Liu, D. DNA nanotechnology based on i-motif structures. Acc. Chem. Res. 2014, 47, 1853–1860. [Google Scholar] [CrossRef]
- Abou Assi, H.; Garavís, M.; González, C.; Damha, M.J. i-Motif DNA: Structural features and significance to cell biology. Nucleic Acids Res. 2018, 46, 8038–8056. [Google Scholar] [CrossRef] [Green Version]
- Day, H.A.; Pavlou, P.; Waller, Z.A.E. i-Motif DNA: Structure, stability and targeting with ligands. Bioorg. Med. Chem. 2014, 22, 4407–4418. [Google Scholar] [CrossRef]
- Schah-Mohammedi, P.; Shenderovich, I.G.; Detering, C.; Limbach, H.-H.; Tolstoy, P.M.; Smirnov, S.N.; Denisov, G.S.; Golubev, N.S. Hydrogen/deuterium-isotope effects on NMR chemical shifts and symmetry of homoconjugated hydrogen-bonded ions in polar solution. J. Am. Chem. Soc. 2000, 122, 12878–12879. [Google Scholar] [CrossRef]
- Lieblein, A.L.; Krämer, M.; Dreuw, A.; Fürtig, B.; Schwalbe, H. The nature of hydrogen bonds in cytidine···H+···cytidine DNA base pairs. Angew. Chem. Int. Ed. 2012, 51, 4067–4070. [Google Scholar] [CrossRef]
- Canalia, M.; Leroy, J.L. Structure, internal motions and association–dissociation kinetics of the i-motif dimer of d(5mCCTCACTCC). Nucleic Acids Res. 2005, 33, 5471–5481. [Google Scholar] [CrossRef] [Green Version]
- Leroy, J.L.; Gehring, K.; Kettani, A.; Gueron, M. Acid multimers of oligodeoxycytidine strands: Stoichiometry, base-pair characterization, and proton exchange properties. Biochemistry 1993, 32, 6019–6031. [Google Scholar] [CrossRef]
- Esmaili, N.; Leroy, J.L. i-motif solution structure and dynamics of the d(AACCCC) and d(CCCCAA) tetrahymena telomeric repeats. Nucleic Acids Res. 2005, 33, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieblein, A.L.; Buck, J.; Schlepckow, K.; Fürtig, B.; Schwalbe, H. Time-resolved NMR spectroscopic studies of DNA i-motif folding reveal kinetic partitioning. Angew. Chem. Int. Ed. 2012, 51, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Fedoroff, O.Y.; Rangan, A.; Chemeris, V.V.; Hurley, L.H. Cationic porphyrins promote the formation of i-motif DNA and bind peripherally by a nonintercalative mechanism. Biochemistry 2000, 39, 15083–15090. [Google Scholar] [CrossRef] [PubMed]
- Alberti, P.; Ren, J.; Teulade-Fichou, M.P.; Guittat, L.; Riou, J.-F.; Chaires, J.B.; Hélène, C.; Vigneron, J.-P.; Lehn, J.-M.; Mergny, J.-L. Interaction of an acridine dimer with DNA quadruplex structures. J. Biomol. Struct. Dyn. 2001, 19, 505–513. [Google Scholar] [CrossRef]
- Sedghi Masoud, S.; Nagasawa, K. i-motif-binding ligands and their effects on the structure and biological functions of i-motif. Chem. Pharm. Bull. (Tokyo) 2018, 66, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Kendrick, S.; Kang, H.-J.; Alam, M.P.; Madathil, M.M.; Agrawal, P.; Gokhale, V.; Yang, D.; Hecht, S.M.; Hurley, L.H. The Dynamic Character of the BCL2 Promoter i-Motif Provides a Mechanism for Modulation of Gene Expression by Compounds That Bind Selectively to the Alternative DNA Hairpin Structure. J. Am. Chem. Soc. 2014, 136, 4161–4171. [Google Scholar] [CrossRef]
- Lacroix, L.; Liénard, H.; Labourier, E.; Djavaheri-Mergny, M.; Lacoste, J.; Leffers, H.; Tazi, J.; Hélène, C.; Mergny, J.L. Identification of two human nuclear proteins that recognise the cytosine-rich strand of human telomeres in vitro. Nucleic Acids Res. 2000, 28, 1564–1575. [Google Scholar] [CrossRef] [Green Version]
- Lannes, L.; Young, P.; Richter, C.; Morgner, N.; Schwalbe, H. Interaction of the N-terminal tandem domains of hnRNP LL with the BCL2 promoter i-motif DNA sequence. ChemBioChem 2017, 18, 2033–2044. [Google Scholar] [CrossRef]
- Felsenfeld, G.; Davies, D.R.; Rich, A. Formation of a three-stranded polynucleotide molecule. J. Am. Chem. Soc. 1957, 79, 2023–2024. [Google Scholar] [CrossRef]
- Moser, H.E.; Dervan, P.B. Sequence-specific cleavage of double helical DNA by triple helix formation. Science 1987, 238, 645–650. [Google Scholar] [CrossRef] [Green Version]
- Beal, P.A.; Dervan, P.B. Second structural motif for recognition of DNA by oligonucleotide-directed triple-helix formation. Science 1991, 251, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Mirkin, S.M.; Lyamichev, V.I.; Drushlyak, K.N.; Dobrynin, V.N.; Filippov, S.A.; Frank-Kamenetskii, M.D. DNA H form requires a homopurine-homopyrimidine mirror repeat. Nature 1987, 330, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Vasquez, K.M. Triplex technology in studies of DNA damage, DNA repair, and mutagenesis. Biochimie 2011, 93, 1197–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooney, S.M.; Moore, P.D. Antiparallel, intramolecular triplex DNA stimulates homologous recombination in human cells. Proc. Natl. Acad. Sci. USA 1995, 92, 2141–2144. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Seidman, M.M.; Glazer, P.M. Mutagenesis in mammalian cells induced by triple helix formation and transcription-coupled repair. Science 1996, 271, 802–805. [Google Scholar] [CrossRef]
- Portes-Sentis, S.; Sergeant, A.; Gruffat, H. A particular DNA structure is required for the function of a cis-acting component of the Epstein-Barr virus OriLyt origin of replication. Nucleic Acids Res. 1997, 25, 1347–1354. [Google Scholar] [CrossRef] [Green Version]
- Plum, G.E.; Park, Y.W.; Singleton, S.F.; Dervan, P.B.; Breslauer, K.J. Thermodynamic characterization of the stability and the melting behavior of a DNA triplex: A spectroscopic and calorimetric study. Proc. Natl. Acad. Sci. USA 1990, 87, 9436–9440. [Google Scholar] [CrossRef] [Green Version]
- Plum, G.E.; Pilch, D.S.; Singleton, S.F.; Breslauer, K.J. Nucleic acid hybridization: Triplex stability and energetics. Annu. Rev. Biophys. Biomol. Struct. 1995, 24, 319–350. [Google Scholar] [CrossRef]
- Reither, S.; Jeltsch, A. Specificity of DNA triple helix formation analyzed by a FRET assay. BMC Biochem. 2002, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.E.; Kim, T.; Kim, S.Y.; Kim, S.W. Detection of triple helix DNA formation of guanine-rich oligonucleotide in sodium ion abundant buffer by cross-checking FRET scheme. Chem. Phys. Lett. 2010, 490, 230–233. [Google Scholar] [CrossRef]
- Idili, A.; Vallée-Bélisle, A.; Ricci, F. Programmable pH-triggered DNA nanoswitches. J. Am. Chem. Soc. 2014, 136, 5836–5839. [Google Scholar] [CrossRef] [PubMed]
- Shields, G.C.; Laughton, C.A.; Orozco, M. Molecular dynamics simulations of the d(T·A·T) triple helix. J. Am. Chem. Soc. 1997, 119, 7463–7469. [Google Scholar] [CrossRef]
- Shields, G.C.; Laughton, C.A.; Orozco, M. Molecular dynamics simulation of a PNA·DNA·PNA triple helix in aqueous solution. J. Am. Chem. Soc. 1998, 120, 5895–5904. [Google Scholar] [CrossRef]
- Esguerra, M.; Nilsson, L.; Villa, A. Triple helical DNA in a duplex context and base pair opening. Nucleic Acids Res. 2014, 42, 11329–11338. [Google Scholar] [CrossRef] [Green Version]
- Ramreddy, T.; Kombrabail, M.; Krishnamoorthy, G.; Rao, B.J. Site-specific dynamics in TAT triplex DNA as revealed by time-domain fluorescence of 2-aminopurine. J. Phys. Chem. B 2009, 113, 6840–6846. [Google Scholar] [CrossRef]
- Powell, S.W.; Jiang, L.; Russu, I.M. Proton exchange and base pair opening in a DNA triple helix. Biochemistry 2001, 40, 11065–11072. [Google Scholar] [CrossRef]
- Jiang, L. Proton exchange and local stability in a DNA triple helix containing a G.TA triad. Nucleic Acids Res. 2001, 29, 4231–4237. [Google Scholar] [CrossRef] [Green Version]
- Coman, D. Site-resolved stabilization of a DNA triple helix by magnesium ions. Nucleic Acids Res. 2004, 32, 878–883. [Google Scholar] [CrossRef] [Green Version]
- Eick, A.; Riechert-Krause, F.; Weisz, K. Binding and NMR structural studies on indoloquinoline-oligonucleotide conjugates targeting duplex DNA. Bioconjugate Chem. 2012, 23, 1127–1137. [Google Scholar] [CrossRef]
- Dickerhoff, J.; Riechert-Krause, F.; Seifert, J.; Weisz, K. Exploring multiple binding sites of an indoloquinoline in triple-helical DNA: A paradigm for DNA triplex-selective intercalators. Biochimie 2014, 107, 327–337. [Google Scholar] [CrossRef]
- Ha, S.C.; Lowenhaupt, K.; Rich, A.; Kim, Y.-G.; Kim, K.K. Crystal structure of a junction between B-DNA and Z-DNA reveals two extruded bases. Nature 2005, 437, 1183–1186. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.H.; Quigley, G.J.; Kolpak, F.J.; Crawford, J.L.; Van Boom, J.H.; Van der Marel, G.; Rich, A. Molecular structure of a left-handed double helical DNA fragment at atomic resolution. Nature 1979, 282, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Pohl, F.M.; Jovin, T.M. Salt-induced co-operative conformational change of a synthetic DNA: Equilibrium and kinetic studies with poly (dG-dC). J. Mol. Biol. 1972, 67, 375–396. [Google Scholar] [CrossRef]
- Wittig, B.; Wölfl, S.; Dorbic, T.; Vahrson, W.; Rich, A. Transcription of human c-myc in permeabilized nuclei is associated with formation of Z-DNA in three discrete regions of the gene. EMBO J. 1992, 11, 4653–4663. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, H.; Chen, X.; Kirby, M.; Brown, P.O.; Zhao, K. Regulation of CSF1 promoter by the SWI/SNF-like BAF complex. Cell 2001, 106, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, A.; Mimura, J.; Harada, N.; Itoh, K. Nrf2 activation is associated with Z-DNA formation in the human HO-1 promoter. Nucleic Acids Res. 2013, 41, 5223–5234. [Google Scholar] [CrossRef] [Green Version]
- Takaoka, A.; Taniguchi, T. Cytosolic DNA recognition for triggering innate immune responses. Adv. Drug Delivery Rev. 2008, 60, 847–857. [Google Scholar] [CrossRef]
- Kim, Y.-G.; Lowenhaupt, K.; Oh, D.-B.; Kim, K.K.; Rich, A. Evidence that vaccinia virulence factor E3L binds to Z-DNA in vivo: Implications for development of a therapy for poxvirus infection. Proc. Natl. Acad. Sci. USA 2004, 101, 1514–1518. [Google Scholar] [CrossRef] [Green Version]
- Bothe, J.R.; Lowenhaupt, K.; Al-Hashimi, H.M. Sequence-specific B-DNA flexibility modulates Z-DNA formation. J. Am. Chem. Soc. 2011, 133, 2016–2018. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.; Kim, D.; Kim, K.K.; Kim, Y.-G.; Hohng, S. Intrinsic Z-DNA is stabilized by the conformational selection mechanism of Z-DNA-binding proteins. J. Am. Chem. Soc. 2011, 133, 668–671. [Google Scholar] [CrossRef]
- Bae, S.; Kim, Y.; Kim, D.; Kim, K.K.; Kim, Y.-G.; Hohng, S. Energetics of Z-DNA binding protein-mediated helicity reversals in DNA, RNA, and DNA–RNA Duplexes. J. Phys. Chem. B 2013, 117, 13866–13871. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kim, S.H.; Hong, S.C. Minute negative superhelicity is sufficient to induce the B-Z transition in the presence of low tension. Proc. Natl. Acad. Sci. USA 2010, 107, 4985–4990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Lim, S.-H.; Lee, A.-R.; Kwon, D.H.; Song, H.K.; Lee, J.-H.; Cho, M.; Johner, A.; Lee, N.-K.; Hong, S.-C. Unveiling the pathway to Z-DNA in the protein-induced B–Z transition. Nucleic Acids Res. 2018, 46, 4129–4137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bothe, J.R.; Lowenhaupt, K.; Al-Hashimi, H.M. Incorporation of CC steps into Z-DNA: Interplay between B–Z junction and Z-DNA helical formation. Biochemistry 2012, 51, 6871–6879. [Google Scholar] [CrossRef]
- Dumat, B.; Larsen, A.F.; Wilhelmsson, L.M. Studying Z-DNA and B- to Z-DNA transitions using a cytosine analogue FRET-pair. Nucleic Acids Res. 2016, 44, e101Ce101. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhang, Y.X.; Sugiyama, H.; Umano, T.; Osuga, H.; Tanaka, K. (P)-helicene displays chiral selection in binding to Z-DNA. J. Am. Chem. Soc. 2004, 126, 6566–6567. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, Y.; An, R.; Liang, X.; Li, Q.; Wang, H.; Wang, H.; Fan, Y.; Dong, P.; Li, J.; et al. Topologically constrained formation of stable Z-DNA from normal sequence under physiological conditions. J. Am. Chem. Soc. 2019, 141, 7758–7764. [Google Scholar] [CrossRef]
- Kladde, M.P.; Kohwi, Y.; Kohwi-Shigematsu, T.; Gorski, J. The non-B-DNA structure of d(CA/TG)n differs from that of Z-DNA. Proc. Natl. Acad. Sci. USA 1994, 91, 1898–1902. [Google Scholar] [CrossRef] [Green Version]
- Peck, L.J.; Wang, J.C. Energetics of B-to-Z transition in DNA. Proc. Natl. Acad. Sci. USA 1983, 80, 6206–6210. [Google Scholar] [CrossRef] [Green Version]
- Ellison, M.J.; Feigon, J.; Kelleher, R.J.; Wang, A.H.; Habener, J.F.; Rich, A. An assessment of the Z-DNA forming potential of alternating dA-dT stretches in supercoiled plasmids. Biochemistry 1986, 25, 3648–3655. [Google Scholar] [CrossRef]
- McLean, M.J.; Blaho, J.A.; Kilpatrick, M.W.; Wells, R.D. Consecutive A X T pairs can adopt a left-handed DNA structure. Proc. Natl. Acad. Sci. USA 1986, 83, 5884–5888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, M.; Fita, I.; Lloveras, J.; Subirana, J.A.; Bardella, F.; Huynh-Dinh, T.; Igolen, J. Structure of d(CACGTG), a Z-DNA hexamer containing AT base pairs. Nucleic Acids Res. 1988, 16, 8695–8705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichman, B.F.; Schroth, G.P.; Basham, B.E.; Ho, P.S. The intrinsic structure and stability of out-of-alternation base pairs in Z-DNA. Nucleic Acids Res. 1999, 27, 543–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abascal, J.L.F.; Domercq, M.; Gil Montoro, J.C. Computer simulation of the ionic atmosphere around Z-DNA. J. Phys. Chem. B 2006, 110, 25080–25090. [Google Scholar] [CrossRef]
- Herbert, A.; Rich, A. The biology of left-handed Z-DNA. J. Biol. Chem. 1996, 271, 11595–11598. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.-A.; Rich, A. Biological function of the vaccinia virus Z-DNA-binding protein E3L: Gene transactivation and antiapoptotic activity in HeLa cells. Proc. Natl. Acas. Sci. USA 2005, 102, 12759–12764. [Google Scholar] [CrossRef] [Green Version]
- Rich, A.; Zhang, S. Timeline: Z-DNA: The long road to biological function. Nat. Rev. Genet. 2003, 4, 566–572. [Google Scholar] [CrossRef]
- Tashiro, R.; Sugiyama, H. A Nanothermometer based on the different π stackings of B- and Z-DNA. Angew Chem. Int. Ed. 2003, 42, 6018–6020. [Google Scholar] [CrossRef]
- Tashiro, R.; Sugiyama, H. Biomolecule-based switching devices that respond inversely to thermal stimuli. J. Am. Chem. Soc. 2005, 127, 2094–2097. [Google Scholar] [CrossRef]
- Behe, M.; Felsenfeld, G. Effects of methylation on a synthetic polynucleotide: The B--Z transition in poly(dG-m5dC) poly(dG-m5dC). Proc. Natl. Acad. Sci. USA 1981, 78, 1619–1623. [Google Scholar] [CrossRef] [Green Version]
- Temiz, N.A.; Donohue, D.E.; Bacolla, A.; Luke, B.T.; Collins, J.R. The role of methylation in the intrinsic dynamics of B- and Z-DNA. PLoS ONE 2012, 7, e35558. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ikeda, R.; Sugiyama, H. 8-Methylguanosine: A powerful Z-DNA stabilizer. J. Am. Chem. Soc. 2003, 125, 13519–13524. [Google Scholar] [CrossRef] [PubMed]
- Kochoyan, M.; Leroy, J.L.; Guéron, M. Proton exchange and base-pair lifetimes in a deoxy-duplex containing a purine-pyrimidine step and in the duplex of inverse sequence. J. Mol. Biol. 1987, 196, 599–609. [Google Scholar] [CrossRef]
- Kochoyan, M.; Leroy, J.L.; Guéron, M. Processes of base-pair opening and proton exchange in Z-DNA. Biochemistry 1990, 29, 4799–4805. [Google Scholar] [CrossRef]
- Kang, Y.-M.; Bang, J.; Lee, E.-H.; Ahn, H.-C.; Seo, Y.-J.; Kim, K.K.; Kim, Y.-G.; Choi, B.-S.; Lee, J.-H. NMR Spectroscopic elucidation of the B−Z transition of a DNA double helix induced by the Zα domain of human ADAR1. J. Am. Chem. Soc. 2009, 131, 11485–11491. [Google Scholar] [CrossRef]
- Herbert, A.; Lowenhaupt, K.; Spitzner, J.; Rich, A. Chicken double-stranded RNA adenosine deaminase has apparent specificity for Z-DNA. Proc. Natl. Acad. Sci. USA 1995, 92, 7550–7554. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, T.; Rould, M.A.; Lowenhaupt, K.; Herbert, A.; Rich, A. Crystal structure of the Zα domain of the human editing enzyme ADAR1 bound to left-handed Z-DNA. Science 1999, 284, 1841–1845. [Google Scholar] [CrossRef]
- Wang, Z.; Choi, M.K.; Ban, T.; Yanai, H.; Negishi, H.; Lu, Y.; Tamura, T.; Takaoka, A.; Nishikura, K.; Taniguchi, T. Regulation of innate immune responses by DAI (DLM-1/ZBP1) and other DNA-sensing molecules. Proc. Natl. Acad. Sci. USA. 2008, 105, 5477–5482. [Google Scholar] [CrossRef] [Green Version]
- Yanai, H.; Savitsky, D.; Tamura, T.; Taniguchi, T. Regulation of the cytosolic DNA-sensing system in innate immunity: A current view. Curr. Opin. Immunol. 2009, 21, 17–22. [Google Scholar] [CrossRef]
- Kim, Y.-G.; Muralinath, M.; Brandt, T.; Pearcy, M.; Hauns, K.; Lowenhaupt, K.; Jacobs, B.L.; Rich, A. A role for Z-DNA binding in vaccinia virus pathogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 6974–6979. [Google Scholar] [CrossRef] [Green Version]
- Rothenburg, S.; Deigendesch, N.; Dittmar, K.; Koch-Nolte, F.; Haag, F.; Lowenhaupt, K.; Rich, A. A PKR-like eukaryotic initiation factor 2α kinase from zebrafish contains Z-DNA binding domains instead of dsRNA binding domains. Proc. Natl. Acad. Sci. USA 2005, 102, 1602–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.-R.; Kim, N.-H.; Seo, Y.-J.; Choi, S.-R.; Lee, J.-H. Thermodynamic model for B-Z transition of DNA induced by Z-DNA binding proteins. Molecules 2018, 23, 2748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.-H.; Seo, Y.-J.; Ahn, H.-C.; Kang, Y.-M.; Kim, H.-E.; Lee, Y.-M.; Choi, B.-S.; Lee, J.-H. NMR study of hydrogen exchange during the B-Z transition of a DNA duplex induced by the Zα domains of yatapoxvirus E3L. FEBS Letters 2010, 584, 4453–4457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.-R.; Park, C.-J.; Cheong, H.-K.; Ryu, K.-S.; Park, J.-W.; Kwon, M.-Y.; Lee, J.; Kim, K.K.; Choi, B.-S.; Lee, J.-H. Solution structure of the Z-DNA binding domain of PKR-like protein kinase from Carassius auratus and quantitative analyses of the intermediate complex during B–Z transition. Nucleic Acids Res. 2016, 44, 2936–2948. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.-R.; Hwang, J.; Hur, J.H.; Ryu, K.-S.; Kim, K.K.; Choi, B.-S.; Kim, N.-K.; Lee, J.-H. NMR dynamics study reveals the Zα domain of human ADAR1 associates with and dissociates from Z-RNA more slowly than Z-DNA. ACS Chem. Biol. 2019, 14, 245–255. [Google Scholar] [CrossRef]
- Ha, S.C.; Choi, J.; Hwang, H.-Y.; Rich, A.; Kim, Y.-G.; Kim, K.K. The structures of non-CG-repeat Z-DNAs co-crystallized with the Z-DNA-binding domain, hZαADAR1. Nucleic Acids Res. 2008, 37, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-E.; Ahn, H.-C.; Lee, Y.-M.; Lee, E.-H.; Seo, Y.-J.; Kim, Y.-G.; Kim, K.K.; Choi, B.-S.; Lee, J.-H. The Zβ domain of human DAI binds to Z-DNA via a novel B-Z transition pathway. FEBS Lett. 2011, 585, 772–778. [Google Scholar] [CrossRef] [Green Version]
- Sheardy, R.D.; Suh, D.; Kurzinsky, R.; Doktycz, M.J.; Benight, A.S.; Chaires, J.B. Sequence dependence of the free energy of B-Z junction formation in deoxyoligonucleotides. J. Mol. Biol. 1993, 231, 475–488. [Google Scholar] [CrossRef]
- Sheardy, R.D.; Levine, N.; Marotta, S.; Suh, D.; Chaires, J.B. A thermodynamic investigation of the melting of B-Z junction forming DNA oligomers. Biochemistry 1994, 33, 1385–1391. [Google Scholar] [CrossRef]
- González, V.M.; Fuertes, M.A.; Pérez, J.M. Kinetics of the salt-induced B-to Z-DNA transition. Biophys. J. 1998, 27, 417–423. [Google Scholar] [CrossRef]
- Kim, D.; Hur, J.; Park, K.; Bae, S.; Shin, D.; Ha, S.C.; Hwang, H.-Y.; Hohng, S.; Lee, J.-H.; Lee, S.; et al. Distinct Z-DNA binding mode of a PKR-like protein kinase containing a Z-DNA binding domain (PKZ). Nucleic Acids Res. 2014, 42, 5937–5948. [Google Scholar] [CrossRef] [PubMed] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, K.-I.; Kim, J.; Park, C.-J.; Lee, J.-H. Dynamics Studies of DNA with Non-canonical Structure Using NMR Spectroscopy. Int. J. Mol. Sci. 2020, 21, 2673. https://doi.org/10.3390/ijms21082673
Oh K-I, Kim J, Park C-J, Lee J-H. Dynamics Studies of DNA with Non-canonical Structure Using NMR Spectroscopy. International Journal of Molecular Sciences. 2020; 21(8):2673. https://doi.org/10.3390/ijms21082673
Chicago/Turabian StyleOh, Kwang-Im, Jinwoo Kim, Chin-Ju Park, and Joon-Hwa Lee. 2020. "Dynamics Studies of DNA with Non-canonical Structure Using NMR Spectroscopy" International Journal of Molecular Sciences 21, no. 8: 2673. https://doi.org/10.3390/ijms21082673
APA StyleOh, K. -I., Kim, J., Park, C. -J., & Lee, J. -H. (2020). Dynamics Studies of DNA with Non-canonical Structure Using NMR Spectroscopy. International Journal of Molecular Sciences, 21(8), 2673. https://doi.org/10.3390/ijms21082673