Recovery of Tendon Characteristics by Inhibition of Aberrant Differentiation of Tendon-Derived Stem Cells from Degenerative Tendinopathy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
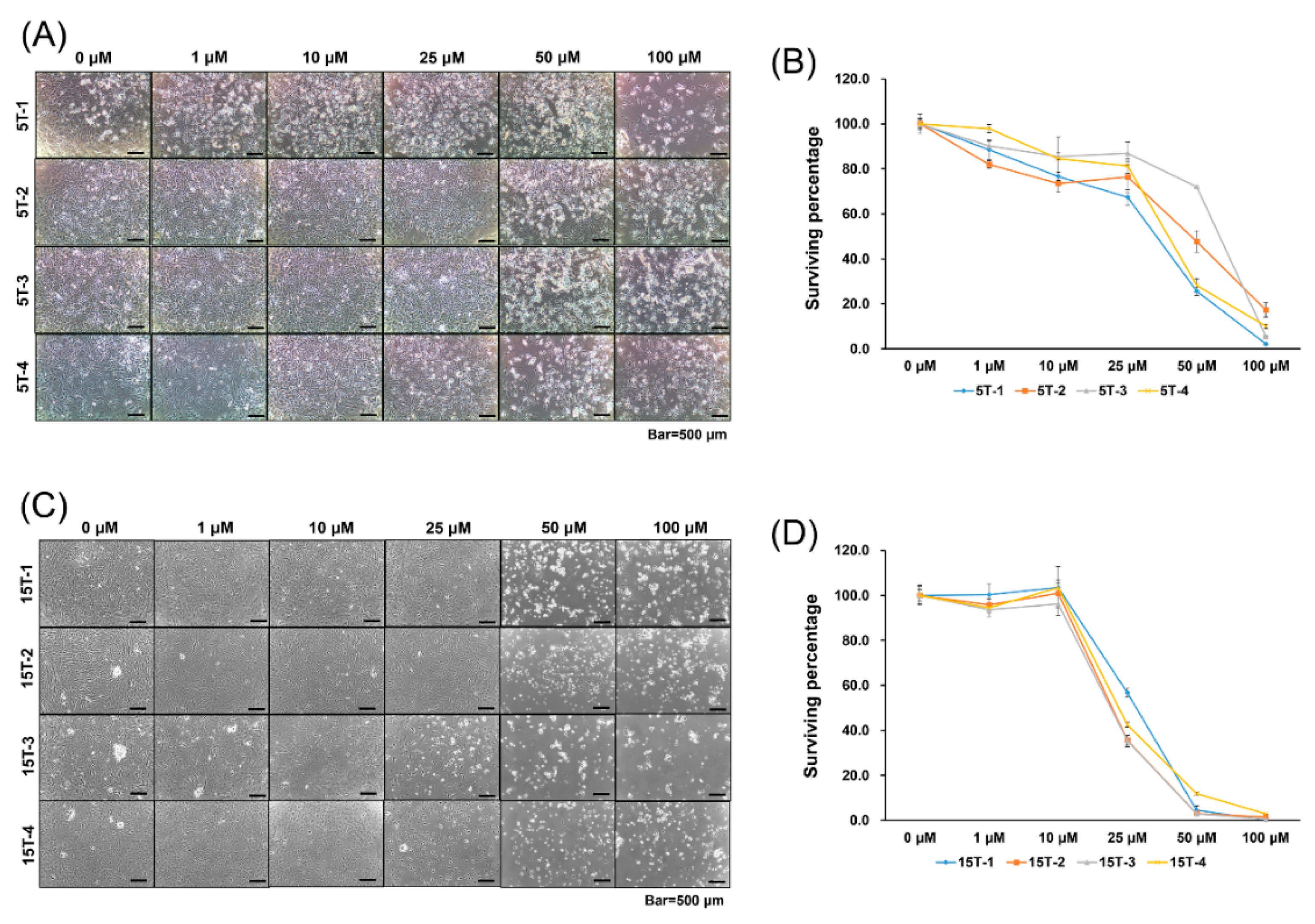
2.1. Effects of AGA and T0070907 on Cytotoxicity Of TDSCs
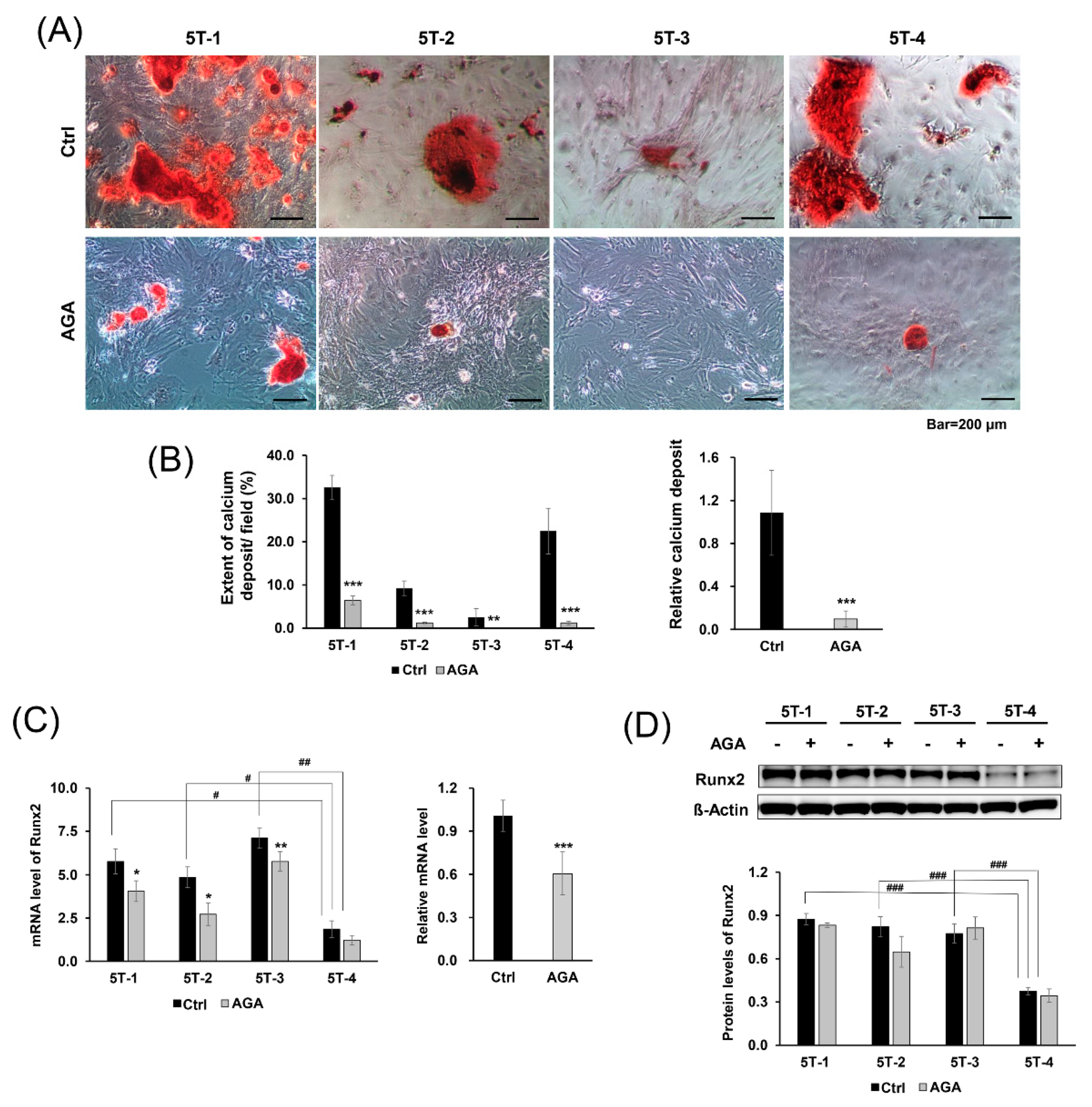
2.2. AGA Blocked Osteogenic Differentiation and Inhibited Expression of Runx2 at the mRNA Level
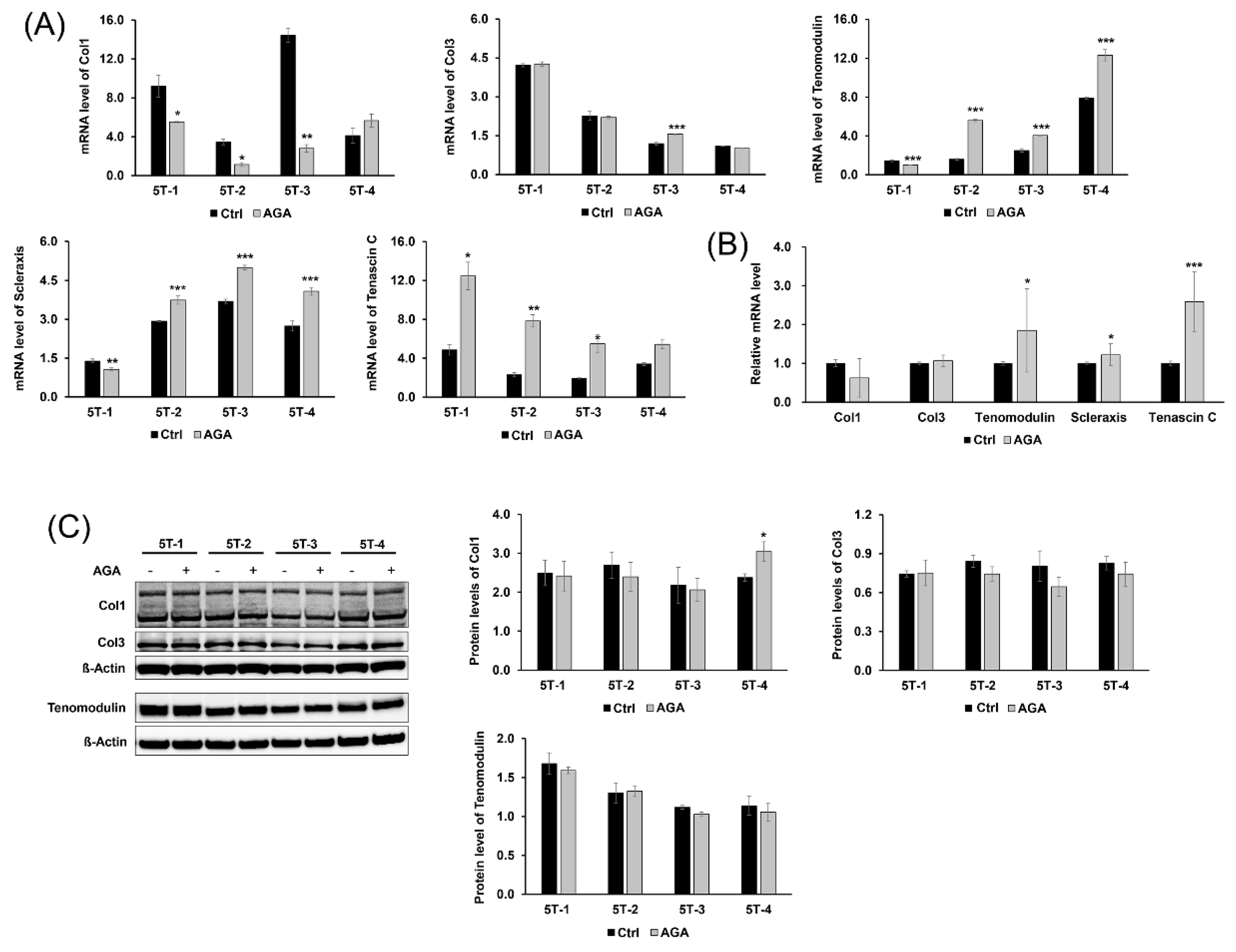
2.3. AGA Induced Tenogenic Differentiation and Regenerative Capacity of TDSCS in Part
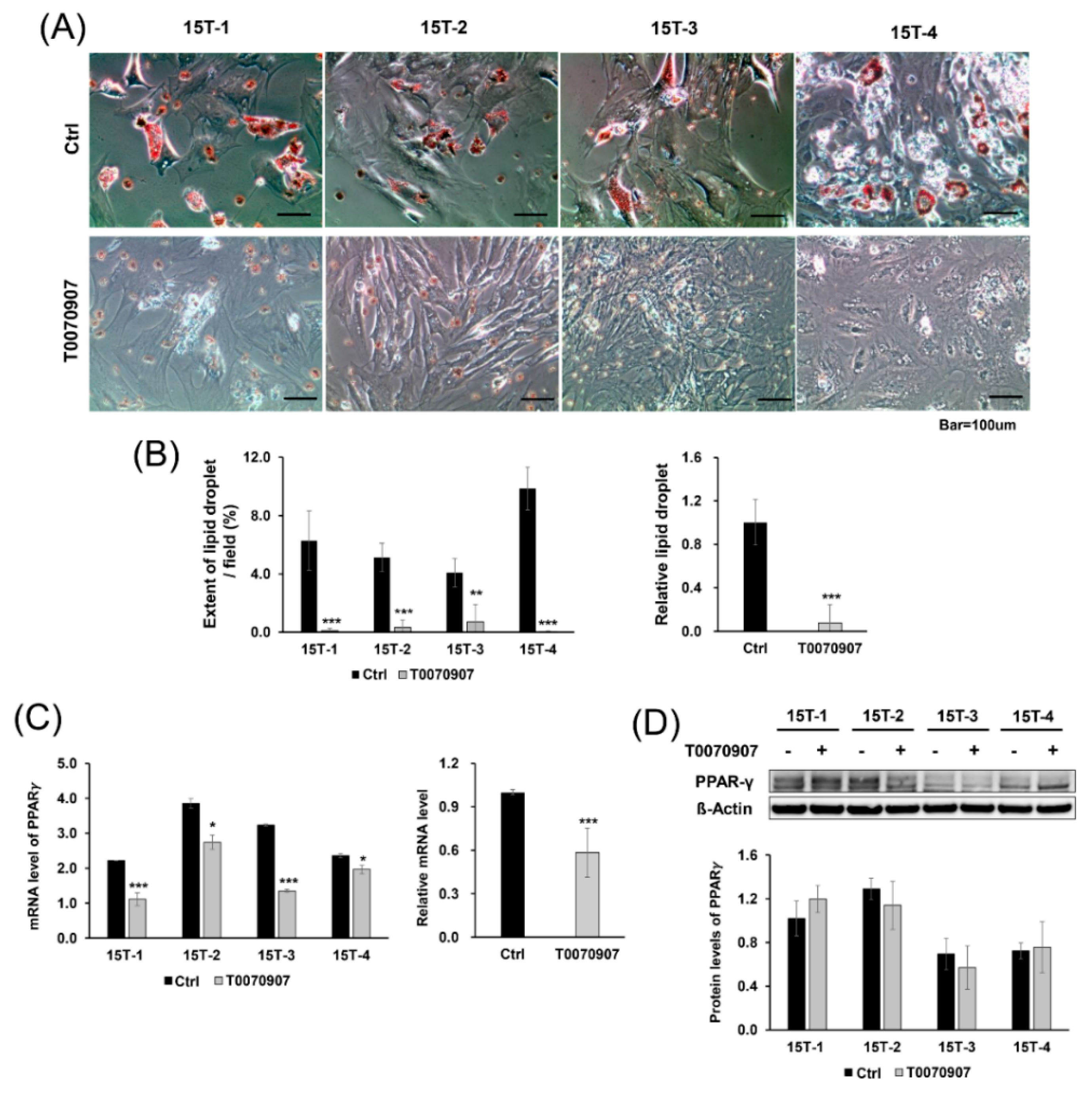
2.4. T0070907 Blocked Adipogenic Differentiation and Inhibited Expression of PPARγ at the mRNA Level
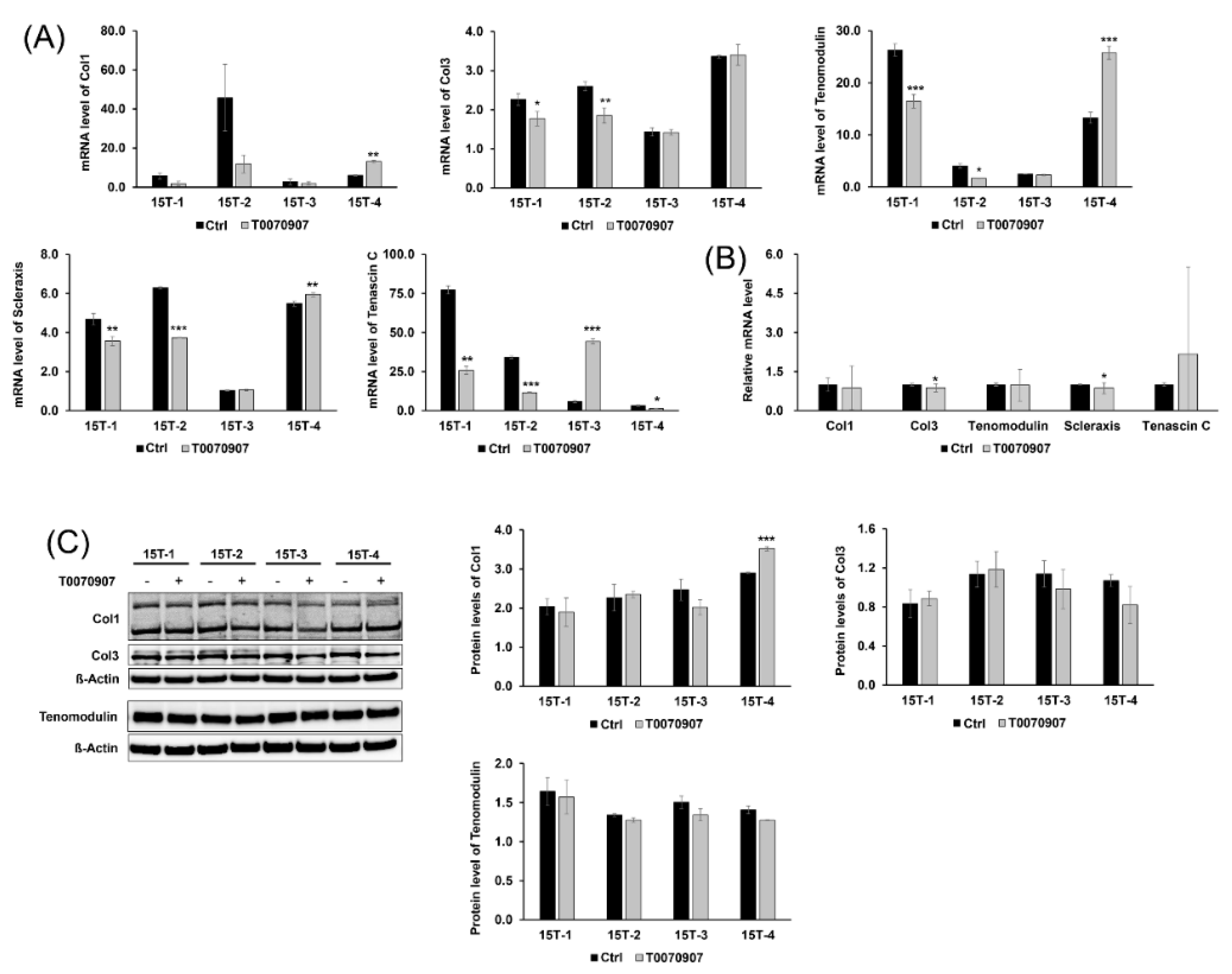
2.5. T0070907 Had No Effect on the Tenogenic Differentiation and Regenerative Capacity of TDSCs
3. Discussion
4. Materials and Methods
4.1. Extraction of TDSCs from Tendinopathy Models
4.2. Selection of Optimal Doses for Blocking Agents
4.3. Inhibition of Aberrant Differentiation by Blocking Agents
4.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.5. Western Blot
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TDSCs | tendon-derived stem cells |
| AGA | 18α-glycyrrhetinic acid |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| Runx2 | Runt-related transcription factor 2 |
| Col1 | collagen type I |
| Col3 | collagen type III |
| OCT4 | octamer-binding transcription factor 4 |
| SSEA4 | stage-specific embryonic antigen-4 |
References
- Wang, J.H.; Iosifidis, M.I.; Fu, F.H. Biomechanical basis for tendinopathy. Clin. Orthop. Relat. Res. 2006, 443, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Abate, M.; Silbernagel, K.G.; Siljeholm, C.; Di Iorio, A.; De Amicis, D.; Salini, V.; Werner, S.; Paganelli, R. Pathogenesis of tendinopathies: Inflammation or degeneration? Arthritis Res. Ther. 2009, 11, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, R.K.; Smith, M.M.; Martin, J.H.; Clarke, J.L.; Dart, A.J.; Little, C.B.; Clarke, E.C. Chondroitin sulphate glycosaminoglycans contribute to widespread inferior biomechanics in tendon after focal injury. J. Biomech. 2016, 49, 2694–2701. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Zhang, J.; Zhao, G.; Zhou, Y.; Zhang, C.Q.; Wang, J.H. Creating an Animal Model of Tendinopathy by Inducing Chondrogenic Differentiation with Kartogenin. PLoS ONE 2016, 11, e0148557. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Song, D.H.; Kim, S.J. Characteristics of tendon derived stem cells according to different factors to induce the tendinopathy. J. Cell Physiol. 2018, 233, 6196–6206. [Google Scholar] [CrossRef]
- Almalki, S.G.; Agrawal, D.K. Key transcription factors in the differentiation of mesenchymal stem cells. Differentiation 2016, 92, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Regulation of bone development and extracellular matrix protein genes by RUNX2. Cell Tissue Res. 2010, 339, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Li, Z.; Hou, Y.; Fang, W. Potential mechanisms underlying the Runx2 induced osteogenesis of bone marrow mesenchymal stem cells. Am. J. Transl. Res. 2015, 7, 2527–2535. [Google Scholar]
- Ling, M.; Huang, P.; Islam, S.; Heruth, D.P.; Li, X.; Zhang, L.Q.; Li, D.Y.; Hu, Z.; Ye, S.Q. Epigenetic regulation of Runx2 transcription and osteoblast differentiation by nicotinamide phosphoribosyltransferase. Cell Biosci. 2017, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.; Wang, Y.; Liu, H.; Nan, X.; Wong, S.; Peng, S.; Gu, Y.; Zhao, H.; Feng, H. Mutant Runx2 regulates amelogenesis and osteogenesis through a miR-185-5p-Dlx2 axis. Cell Death Dis. 2017, 8, 3221. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Regulation of Proliferation, Differentiation and Functions of Osteoblasts by Runx2. Int. J. Mol. Sci. 2019, 20, 1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, A.; Barak, Y.; Nagy, L.; Liao, D.; Tontonoz, P.; Evans, R.M. PPAR-gamma dependent and independent effects on macrophage-gene expression in lipid metabolism and inflammation. Nat. Med. 2001, 7, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Siersbaek, R.; Nielsen, R.; Mandrup, S. PPARgamma in adipocyte differentiation and metabolism--novel insights from genome-wide studies. FEBS Lett. 2010, 584, 3242–3249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefterova, M.I.; Haakonsson, A.K.; Lazar, M.A.; Mandrup, S. PPARγ and the global map of adipogenesis and beyond. Trends Endocrinol. Metab. 2014, 25, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Jeong, M.; Lee, S.S.; Song, J. Camptothecin and topotecan inhibit adipocyte differentiation by inducing degradation of PPARgamma. Biochem. Biophys. Res. Commun. 2015, 463, 1122–1128. [Google Scholar] [CrossRef]
- Chen, L.; Chen, Y.; Zhang, S.; Ye, L.; Cui, J.; Sun, Q.; Li, K.; Wu, H.; Liu, L. MiR-540 as a novel adipogenic inhibitor impairs adipogenesis via suppression of PPARgamma. J. Cell Biochem. 2015, 116, 969–976. [Google Scholar] [CrossRef]
- Tamori, Y.; Masugi, J.; Nishino, N.; Kasuga, M. Role of peroxisome proliferator-activated receptor-gamma in maintenance of the characteristics of mature 3T3-L1 adipocytes. Diabetes 2002, 51, 2045–2055. [Google Scholar] [CrossRef] [Green Version]
- D’Alimonte, I.; Lannutti, A.; Pipino, C.; Di Tomo, P.; Pierdomenico, L.; Cianci, E.; Antonucci, I.; Marchisio, M.; Romano, M.; Stuppia, L.; et al. Wnt signaling behaves as a "master regulator" in the osteogenic and adipogenic commitment of human amniotic fluid mesenchymal stem cells. Stem. Cell Rev. Rep. 2013, 9, 642–654. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.P.; Deng, M.T.; Chen, L.Y.; Fang, N.; Du, C.; Chen, L.P.; Zou, Y.Q.; Dai, J.H.; Zhu, M.L.; Wang, W.; et al. Shp2 regulates chlorogenic acid-induced proliferation and adipogenic differentiation of bone marrow-derived mesenchymal stem cells in adipogenesis. Mol. Med. Rep. 2015, 11, 4489–4495. [Google Scholar] [CrossRef]
- Yuan, Z.; Li, Q.; Luo, S.; Liu, Z.; Luo, D.; Zhang, B.; Zhang, D.; Rao, P.; Xiao, J. PPARγ and Wnt Signaling in Adipogenic and Osteogenic Differentiation of Mesenchymal Stem Cells. Curr. Stem. Cell Res. Ther. 2016, 11, 216–225. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, W.; Lim, C.; Chung, S.G. Treatment of Lateral Epicondylosis by Using Allogeneic Adipose-Derived Mesenchymal Stem Cells: A Pilot Study. Stem. Cells 2015, 33, 2995–3005. [Google Scholar] [CrossRef]
- Usuelli, F.G.; Grassi, M.; Maccario, C.; Vigano, M.; Lanfranchi, L.; Alfieri Montrasio, U.; de Girolamo, L. Intratendinous adipose-derived stromal vascular fraction (SVF) injection provides a safe, efficacious treatment for Achilles tendinopathy: Results of a randomized controlled clinical trial at a 6-month follow-up. Knee Surg. Sports Traumatol. Arthrosc. 2018, 26, 2000–2010. [Google Scholar] [CrossRef]
- Kim, S.J.; Yeo, S.M.; Noh, S.J.; Ha, C.W.; Lee, B.C.; Lee, H.S.; Kim, S.J. Effect of platelet-rich plasma on the degenerative rotator cuff tendinopathy according to the compositions. J. Orthop.Surg. Res. 2019, 14, 408. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Martinez-Williams, C.; Gilbert, K.A.; Rannels, D.E. Inhibition of gap junction communication in alveolar epithelial cells by 18alpha-glycyrrhetinic acid. Am. J. Physiol. 1999, 276, L1018–L1026. [Google Scholar]
- Talbot, J.; Brion, R.; Lamora, A.; Mullard, M.; Morice, S.; Heymann, D.; Verrecchia, F. Connexin43 intercellular communication drives the early differentiation of human bone marrow stromal cells into osteoblasts. J. Cell Physiol. 2018, 233, 946–957. [Google Scholar] [CrossRef]
- Jeong, H.M.; Cho, D.H.; Jin, Y.H.; Chung, J.O.; Chung, M.Y.; Chung, D.J.; Lee, K.Y. Inhibition of osteoblastic differentiation by warfarin and 18-alpha-glycyrrhetinic acid. Arch. Pharm. Res. 2011, 34, 1381–1387. [Google Scholar] [CrossRef]
- Lee, G.; Elwood, F.; McNally, J.; Weiszmann, J.; Lindstrom, M.; Amaral, K.; Nakamura, M.; Miao, S.; Cao, P.; Learned, R.M.; et al. T0070907, a selective ligand for peroxisome proliferator-activated receptor gamma, functions as an antagonist of biochemical and cellular activities. J. Biol. Chem. 2002, 277, 19649–19657. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, A.; Tomimoto, A.; Fujita, K.; Sugiyama, M.; Takahashi, H.; Ikeda, I.; Hosono, K.; Endo, H.; Yoneda, K.; Iida, H.; et al. Inhibition of peroxisome proliferator-activated receptor gamma activity suppresses pancreatic cancer cell motility. Cancer Sci. 2008, 99, 1892–1900. [Google Scholar] [CrossRef]
- Schaefer, K.L.; Wada, K.; Takahashi, H.; Matsuhashi, N.; Ohnishi, S.; Wolfe, M.M.; Turner, J.R.; Nakajima, A.; Borkan, S.C.; Saubermann, L.J. Peroxisome proliferator-activated receptor gamma inhibition prevents adhesion to the extracellular matrix and induces anoikis in hepatocellular carcinoma cells. Cancer Res. 2005, 65, 2251–2259. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, A.; Haraguchi, N.; Tsuchiya, H.; Ikeda, Y.; Hama, S.; Kogure, K. Peroxisome proliferator-activated receptor gamma (PPARgamma)-independent specific cytotoxicity against immature adipocytes induced by PPARgamma antagonist T0070907. Biol. Pharm. Bull. 2013, 36, 1428–1434. [Google Scholar] [CrossRef] [Green Version]
- Zaytseva, Y.Y.; Wallis, N.K.; Southard, R.C.; Kilgore, M.W. The PPARgamma antagonist T0070907 suppresses breast cancer cell proliferation and motility via both PPARgamma-dependent and -independent mechanisms. Anticancer Res. 2011, 31, 813–823. [Google Scholar] [PubMed]
- Takahashi, H.; Fujita, K.; Fujisawa, T.; Yonemitsu, K.; Tomimoto, A.; Ikeda, I.; Yoneda, M.; Masuda, T.; Schaefer, K.; Saubermann, L.J.; et al. Inhibition of peroxisome proliferator-activated receptor gamma activity in esophageal carcinoma cells results in a drastic decrease of invasive properties. Cancer Sci. 2006, 97, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.D.; Goldenberg, D.M.; Blumenthal, R.D. Potential of peroxisome proliferator-activated receptor gamma antagonist compounds as therapeutic agents for a wide range of cancer types. PPAR Res. 2008, 2008, 494161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosco, D.B.; Roycik, M.D.; Jin, Y.; Schwartz, M.A.; Lively, T.J.; Zorio, D.A.; Sang, Q.A. A new synthetic matrix metalloproteinase inhibitor reduces human mesenchymal stem cell adipogenesis. PLoS ONE 2017, 12, e0172925. [Google Scholar] [CrossRef]
- Liang, X.; Shang, Z.; Wei, T. Role of Cx43-Based Gap Junction in Murine Osteoblast-Like Mc3t3-E1Cells Exposed to 17-Βestradiol. Mol. Biol. 2017, 6. [Google Scholar] [CrossRef]
- Lui, P.P. Markers for the identification of tendon-derived stem cells in vitro and tendon stem cells in situ—Update and future development. Stem. Cell Res. Ther. 2015, 6, 106. [Google Scholar] [CrossRef] [Green Version]
- Wagenhauser, M.U.; Pietschmann, M.F.; Sievers, B.; Docheva, D.; Schieker, M.; Jansson, V.; Muller, P.E. Collagen type I and decorin expression in tenocytes depend on the cell isolation method. BMC Musculoskelet Disord. 2012, 13, 140. [Google Scholar] [CrossRef] [Green Version]
- Riley, G. Tendinopathy—From basic science to treatment. Nat. Clin. Pract. Rheumatol. 2008, 4, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [Green Version]
- Simonin, M.A.; Gegout-Pottie, P.; Minn, A.; Gillet, P.; Netter, P.; Terlain, B. Pefloxacin-induced achilles tendon toxicity in rodents: Biochemical changes in proteoglycan synthesis and oxidative damage to collagen. Antimicrob. Agents Chemother. 2000, 44, 867–872. [Google Scholar] [CrossRef] [Green Version]
- Glazebrook, M.A.; Wright, J.R., Jr.; Langman, M.; Stanish, W.D.; Lee, J.M. Histological analysis of achilles tendons in an overuse rat model. J. Orthop. Res. 2008, 26, 840–846. [Google Scholar] [CrossRef]
- Chen, L.; Liu, J.P.; Tang, K.L.; Wang, Q.; Wang, G.D.; Cai, X.H.; Liu, X.M. Tendon derived stem cells promote platelet-rich plasma healing in collagenase-induced rat achilles tendinopathy. Cell Physiol. Biochem. 2014, 34, 2153–2168. [Google Scholar] [CrossRef]
- De Mos, M.; Koevoet, W.; van Schie, H.T.; Kops, N.; Jahr, H.; Verhaar, J.A.; van Osch, G.J. In vitro model to study chondrogenic differentiation in tendinopathy. Am. J. Sports Med. 2009, 37, 1214–1222. [Google Scholar] [CrossRef]
- Rui, Y.F.; Lui, P.P.; Wong, Y.M.; Tan, Q.; Chan, K.M. Altered fate of tendon-derived stem cells isolated from a failed tendon-healing animal model of tendinopathy. Stem. Cells Dev. 2013, 22, 1076–1085. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, J.H. The effects of mechanical loading on tendons--an in vivo and in vitro model study. PLoS ONE 2013, 8, e71740. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.J.; Oh, H.W.; Chang, J.W.; Kim, S.J. Recovery of Tendon Characteristics by Inhibition of Aberrant Differentiation of Tendon-Derived Stem Cells from Degenerative Tendinopathy. Int. J. Mol. Sci. 2020, 21, 2687. https://doi.org/10.3390/ijms21082687
Kim SJ, Oh HW, Chang JW, Kim SJ. Recovery of Tendon Characteristics by Inhibition of Aberrant Differentiation of Tendon-Derived Stem Cells from Degenerative Tendinopathy. International Journal of Molecular Sciences. 2020; 21(8):2687. https://doi.org/10.3390/ijms21082687
Chicago/Turabian StyleKim, Sun Jeong, Hae Won Oh, Jong Wook Chang, and Sang Jun Kim. 2020. "Recovery of Tendon Characteristics by Inhibition of Aberrant Differentiation of Tendon-Derived Stem Cells from Degenerative Tendinopathy" International Journal of Molecular Sciences 21, no. 8: 2687. https://doi.org/10.3390/ijms21082687
APA StyleKim, S. J., Oh, H. W., Chang, J. W., & Kim, S. J. (2020). Recovery of Tendon Characteristics by Inhibition of Aberrant Differentiation of Tendon-Derived Stem Cells from Degenerative Tendinopathy. International Journal of Molecular Sciences, 21(8), 2687. https://doi.org/10.3390/ijms21082687