Differential Expression and Function of Bicellular Tight Junctions in Skin and Oral Wound Healing


Abstract
:1. Introduction
2. Results
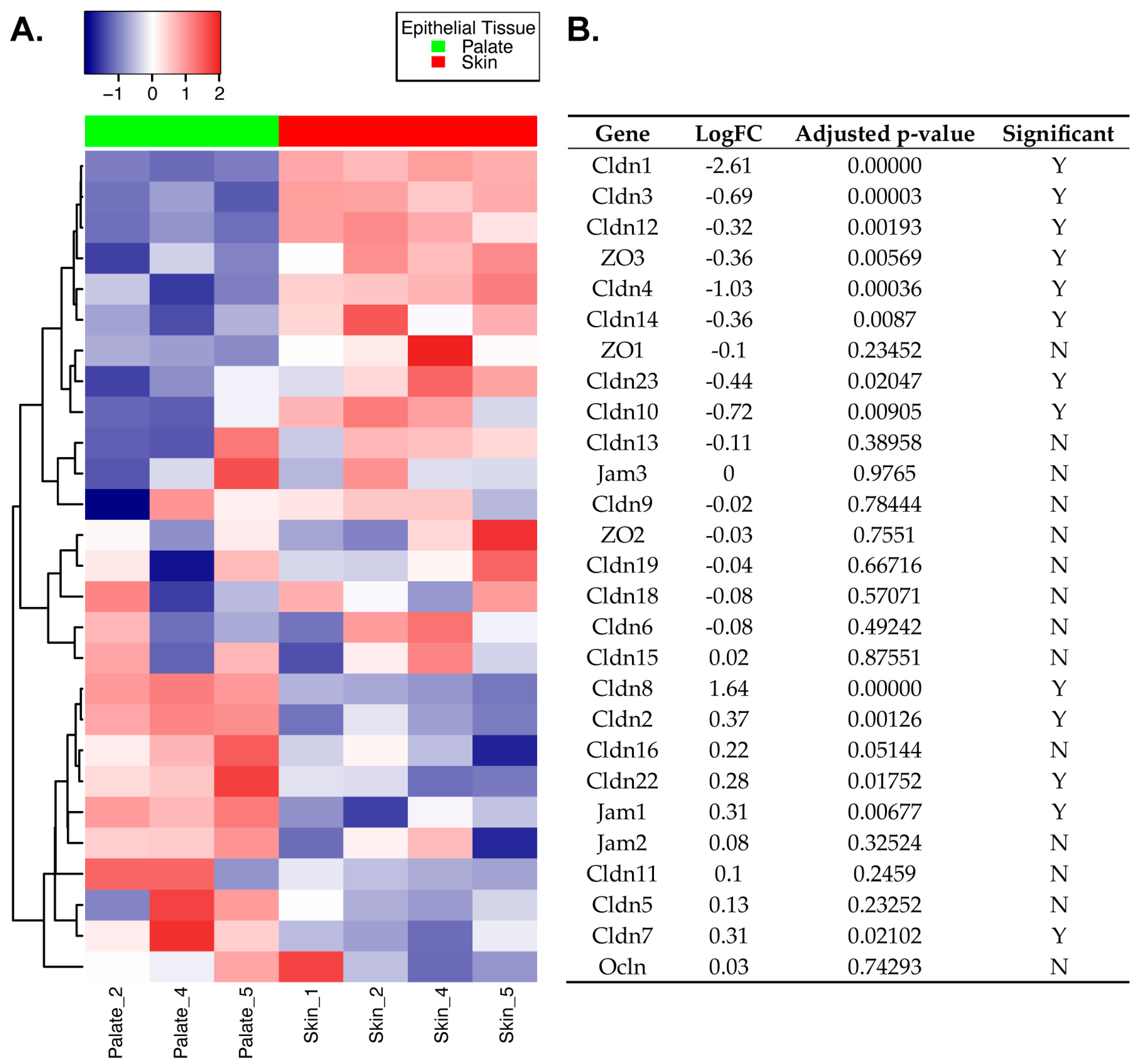
2.1. Differential Expression of TJ Molecules in Primary Mouse Skin and Oral Epithelial Cells
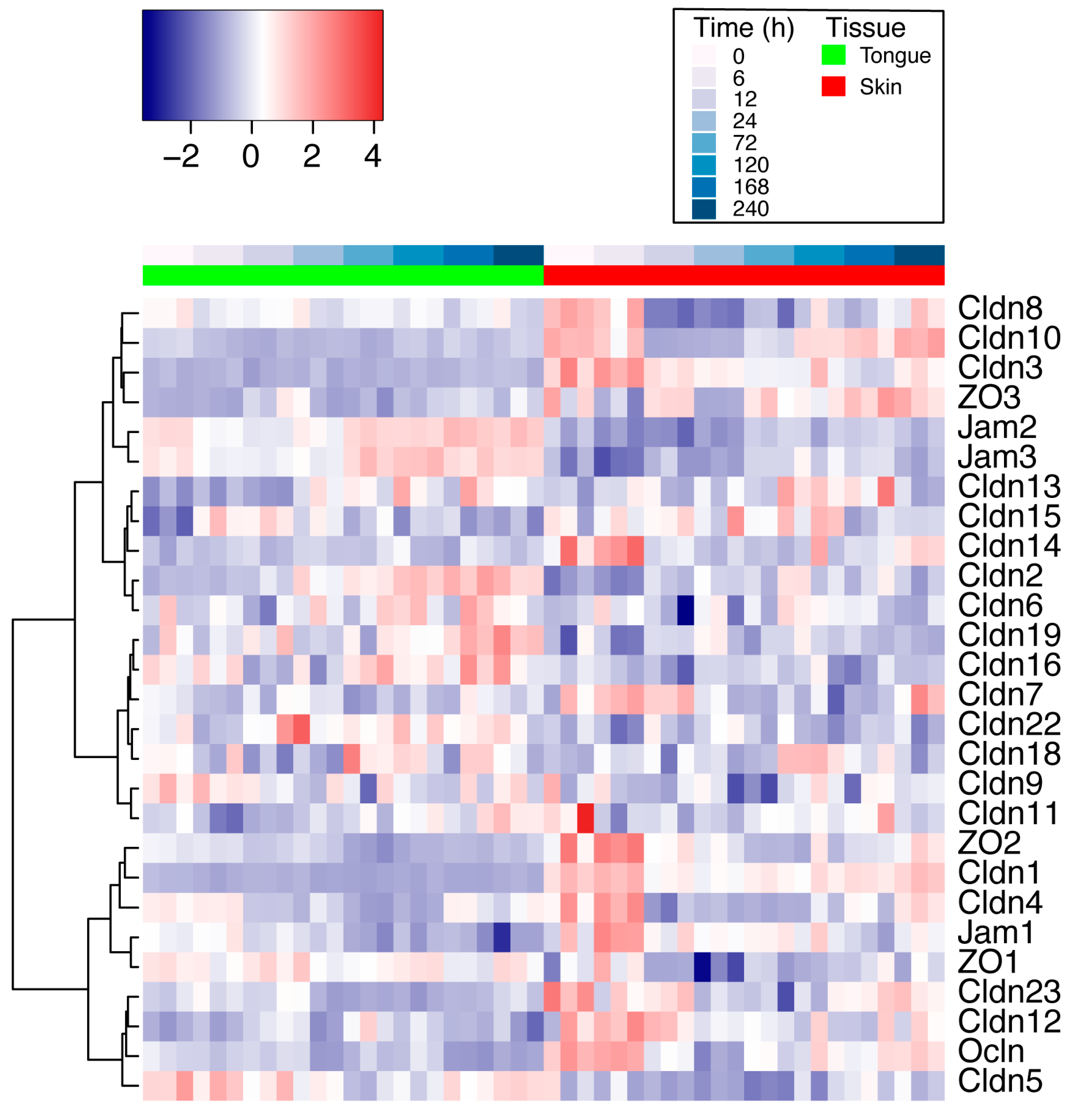
2.2. Differential Expression of TJ Molecules in Mouse Skin and Oral Wounds In Vivo
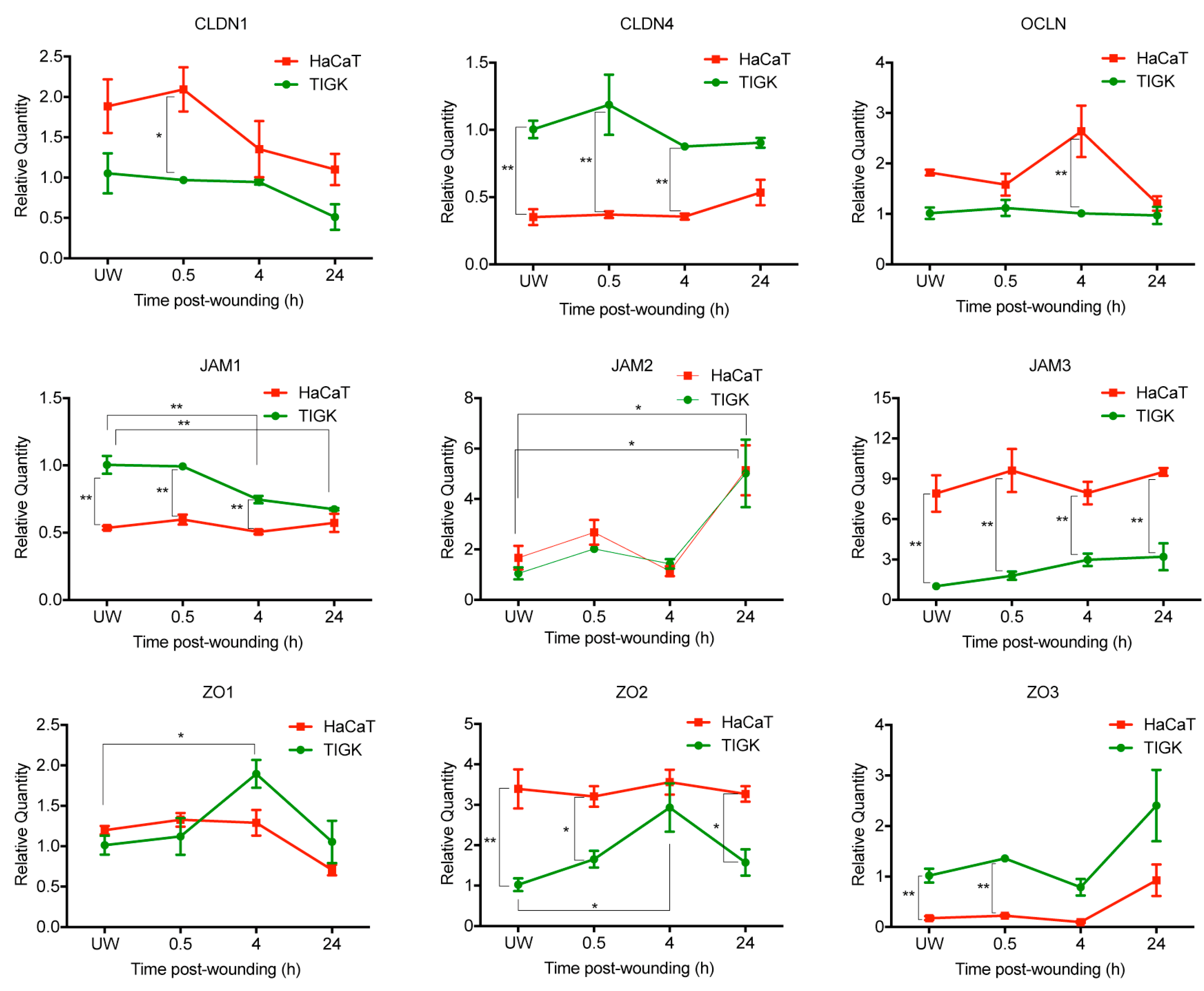
2.3. Differential Expression of TJ Molecules in Human Skin and Oral Keratinocytes after Injury
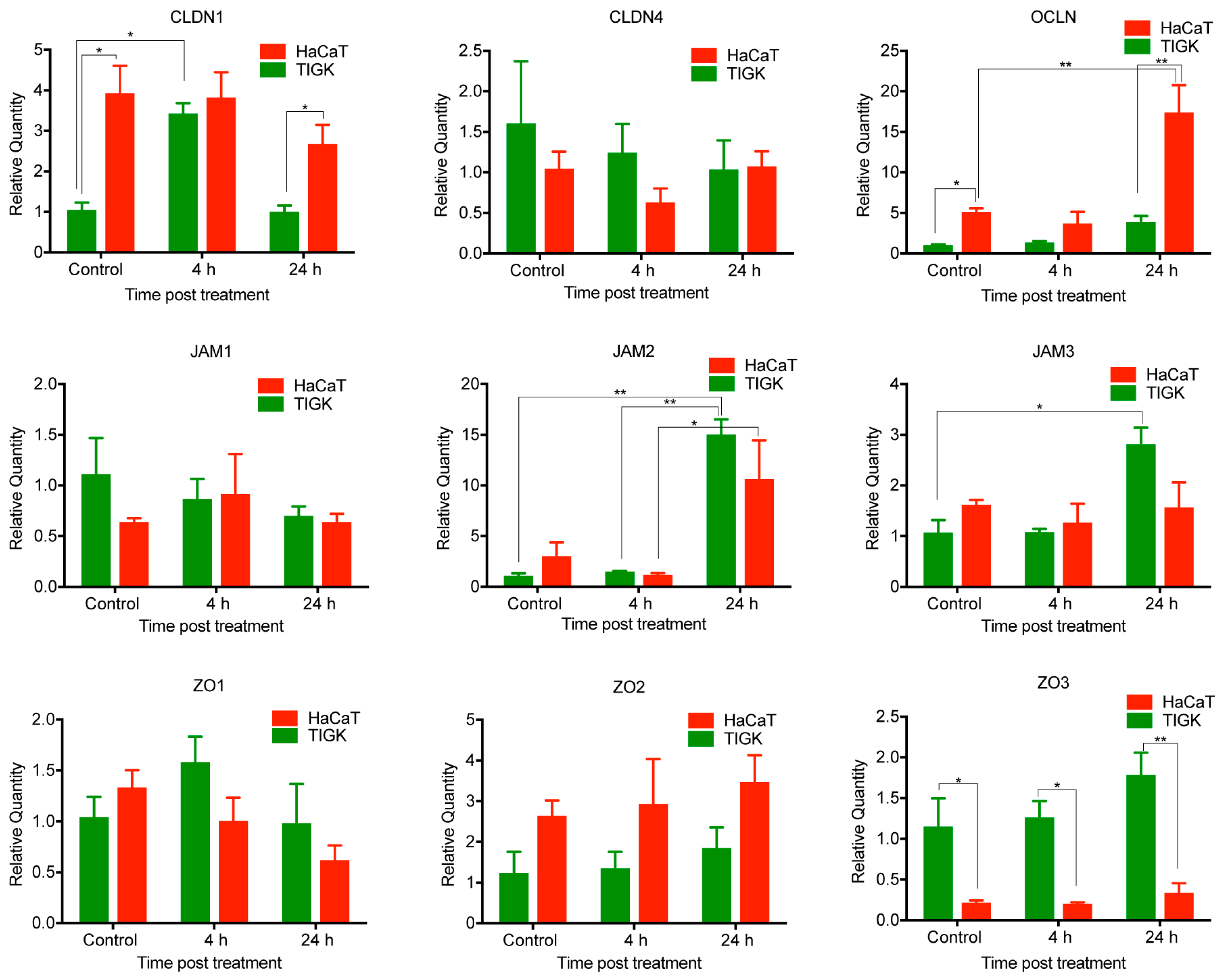
2.4. Differential Expression of TJ Molecules in Human Skin and Oral Keratinocytes after LPS Treatment
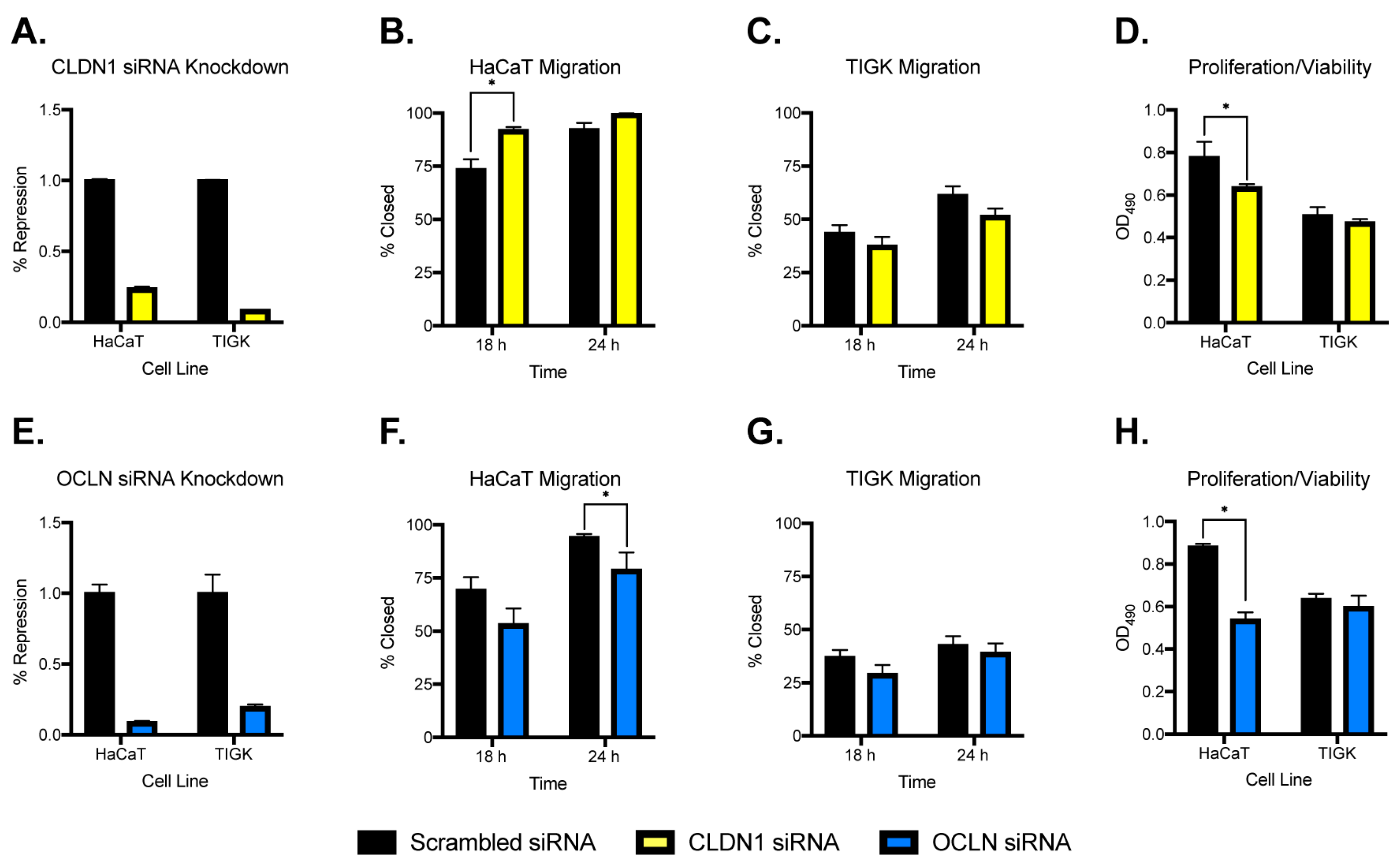
2.5. Inhibition of Claudin 1 or Occludin Alters Migration and Proliferation/Viability of Skin Keratinocytes but Not Oral Keratinocytes
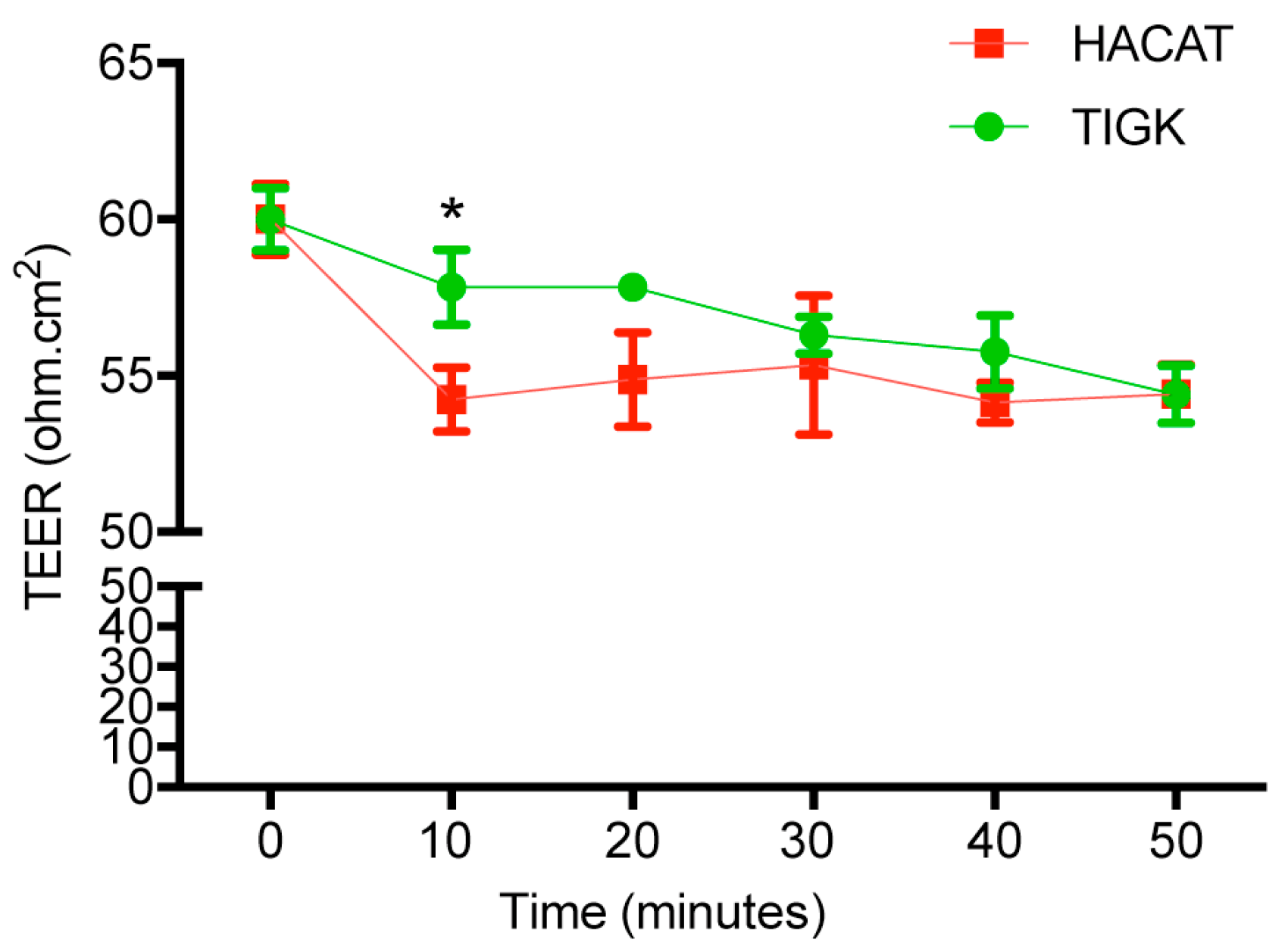
2.6. Human Skin and Gingival Keratinocytes Had Distinct Cell Permeability after Histamine Treatment
3. Discussion
4. Materials and Methods
4.1. Animals and Wound Models for Microarray
4.2. Microarray Data Retrieval and Normalization
4.3. Microarray Differential Expression Analysis
4.4. Cell Culture
4.5. Keratinocyte Migration and Proliferation/Viability after Occludin or Claudin 1 siRNA Knockdown
4.6. Real-Time PCR (RT-PCR)
4.7. TEER Measurement
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CLDN | claudin |
| DE | differentially expressed |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| HaCaT | immortalized human skin keratinocytes |
| JAM | junctional adhesion molecules |
| LPS | lipopolysaccharide |
| OCLN | occludin |
| TEER | transepithelial electrical resistance |
| TIGK | human telomerase immortalized gingival keratinocytes |
| TJ | tight junction |
| ZO | zonula occludens |
References
- Guo, S.; Dipietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.W.; Berg, J.H.; Mackenzie, I.C. Quantitative evaluation of regional differences between epithelia in the adult mouse. Arch. Oral Biol. 1981, 26, 1063–1067. [Google Scholar] [CrossRef]
- Chen, L.; Arbieva, Z.H.; Guo, S.; Marucha, P.T.; Mustoe, T.A.; DiPietro, L.A. Positional differences in the wound transcriptome of skin and oral mucosa. BMC Genom. 2010, 11, 471. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Gajendrareddy, P.K.; DiPietro, L.A. Differential expression of HIF-1alpha in skin and mucosal wounds. J. Dent. Res. 2012, 91, 871–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrementi, M.E.; Ferreira, A.M.; Zender, C.; DiPietro, L.A. Site-specific production of TGF-beta in oral mucosal and cutaneous wounds. Wound Repair Regen. 2008, 16, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Szpaderska, A.M.; Walsh, C.G.; Steinberg, M.J.; DiPietro, L.A. Distinct patterns of angiogenesis in oral and skin wounds. J. Dent. Res. 2005, 84, 309–314. [Google Scholar] [CrossRef]
- Szpaderska, A.M.; Zuckerman, J.D.; DiPietro, L.A. Differential injury responses in oral mucosal and cutaneous wounds. J. Dent. Res. 2003, 82, 621–626. [Google Scholar] [CrossRef]
- Turabelidze, A.; Guo, S.; Chung, A.Y.; Chen, L.; Dai, Y.; Marucha, P.T.; DiPietro, L.A. Intrinsic differences between oral and skin keratinocytes. PLoS ONE 2014, 9, e101480. [Google Scholar] [CrossRef]
- Iglesias-Bartolome, R.; Uchiyama, A.; Molinolo, A.A.; Abusleme, L.; Brooks, S.R.; Callejas-Valera, J.L.; Edwards, D.; Doci, C.; Asselin-Labat, M.L.; Onaitis, M.W.; et al. Transcriptional signature primes human oral mucosa for rapid wound healing. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Mackenzie, I.C.; Hill, M.W. Connective tissue influences on patterns of epithelial architecture and keratinization in skin and oral mucosa of the adult mouse. Cell Tissue Res. 1984, 235, 551–559. [Google Scholar] [CrossRef]
- Brandner, J.M. Importance of Tight Junctions in Relation to Skin Barrier Function. Curr. Probl. Dermatol. 2016, 49, 27–37. [Google Scholar] [PubMed]
- Kirschner, N.; Brandner, J.M. Barriers and more: Functions of tight junction proteins in the skin. Ann. N. Y. Acad. Sci. 2012, 1257, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Kyuno, D.; Yamaguchi, H.; Ito, T.; Kono, T.; Kimura, Y.; Imamura, M.; Konno, T.; Hirata, K.; Sawada, N.; Kojima, T. Targeting tight junctions during epithelial to mesenchymal transition in human pancreatic cancer. World J. Gastroenterol. 2014, 20, 10813–10824. [Google Scholar] [CrossRef] [PubMed]
- Raleigh, D.R.; Marchiando, A.M.; Zhang, Y.; Shen, L.; Sasaki, H.; Wang, Y.; Long, M.; Turner, J.R. Tight junction-associated MARVEL proteins marveld3, tricellulin, and occludin have distinct but overlapping functions. Mol. Biol. Cell 2010, 21, 1200–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkas, A.E.; Capaldo, C.T.; Nusrat, A. Regulation of epithelial proliferation by tight junction proteins. Ann. N. Y. Acad. Sci. 2012, 1258, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Volksdorf, T.; Heilmann, J.; Eming, S.A.; Schawjinski, K.; Zorn-Kruppa, M.; Ueck, C.; Vidal, Y.S.S.; Windhorst, S.; Jucker, M.; Moll, I.; et al. Tight Junction Proteins Claudin-1 and Occludin Are Important for Cutaneous Wound Healing. Am. J. Pathol. 2017, 187, 1301–1312. [Google Scholar] [CrossRef] [Green Version]
- Mandel, I.; Paperna, T.; Glass-Marmor, L.; Volkowich, A.; Badarny, S.; Schwartz, I.; Vardi, P.; Koren, I.; Miller, A. Tight junction proteins expression and modulation in immune cells and multiple sclerosis. J. Cell. Mol. Med. 2012, 16, 765–775. [Google Scholar] [CrossRef]
- Chen, L.; Schrementi, M.E.; Ranzer, M.J.; Wilgus, T.A.; DiPietro, L.A. Blockade of mast cell activation reduces cutaneous scar formation. PLoS ONE 2014, 9, e85226. [Google Scholar] [CrossRef] [Green Version]
- Egozi, E.I.; Ferreira, A.M.; Burns, A.L.; Gamelli, R.L.; Dipietro, L.A. Mast cells modulate the inflammatory but not the proliferative response in healing wounds. Wound Repair Regen. 2003, 11, 46–54. [Google Scholar] [CrossRef]
- Weller, K.; Foitzik, K.; Paus, R.; Syska, W.; Maurer, M. Mast cells are required for normal healing of skin wounds in mice. FASEB J. 2006, 20, 2366–2368. [Google Scholar] [CrossRef]
- Numata, Y.; Terui, T.; Okuyama, R.; Hirasawa, N.; Sugiura, Y.; Miyoshi, I.; Watanabe, T.; Kuramasu, A.; Tagami, H.; Ohtsu, H. The accelerating effect of histamine on the cutaneous wound-healing process through the action of basic fibroblast growth factor. J. Investig. Dermatol. 2006, 126, 1403–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedetto, A.; Rafaels, N.M.; McGirt, L.Y.; Ivanov, A.I.; Georas, S.N.; Cheadle, C.; Berger, A.E.; Zhang, K.; Vidyasagar, S.; Yoshida, T.; et al. Tight junction defects in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2011, 127, 773–786.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedetto, A.; Yoshida, T.; Fridy, S.; Park, J.E.; Kuo, I.H.; Beck, L.A. Histamine and Skin Barrier: Are Histamine Antagonists Useful for the Prevention or Treatment of Atopic Dermatitis? J. Clin. Med. 2015, 4, 741–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malminen, M.; Koivukangas, V.; Peltonen, J.; Karvonen, S.L.; Oikarinen, A.; Peltonen, S. Immunohistological distribution of the tight junction components ZO-1 and occludin in regenerating human epidermis. Br. J. Dermatol. 2003, 149, 255–260. [Google Scholar] [CrossRef]
- Andersen, L. Cell junctions in squamous epithelium during wound healing in palatal mucosa of guinea pigs. Scand. J. Dent. Res. 1980, 88, 328–339. [Google Scholar] [CrossRef]
- Shi, J.; Barakat, M.; Chen, D.; Chen, L. Bicellular Tight Junctions and Wound Healing. Int. J. Mol. Sci. 2018, 19, 3862. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Simoes, A.; Chen, Z.; Zhao, Y.; Wu, X.; Dai, Y.; DiPietro, L.A.; Zhou, X. Overexpression of the Oral Mucosa-Specific microRNA-31 Promotes Skin Wound Closure. Int. J. Mol. Sci. 2019, 20, 3679. [Google Scholar] [CrossRef] [Green Version]
- Simoes, A.; Chen, L.; Chen, Z.; Zhao, Y.; Gao, S.; Marucha, P.T.; Dai, Y.; DiPietro, L.A.; Zhou, X. Differential microRNA profile underlies the divergent healing responses in skin and oral mucosal wounds. Sci. Rep. 2019, 9, 7160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokumasu, R.; Tamura, A.; Tsukita, S. Time- and dose-dependent claudin contribution to biological functions: Lessons from claudin-1 in skin. Tissue Barriers 2017, 5, e1336194. [Google Scholar] [CrossRef]
- Cheng, B.; Rong, A.; Zhou, Q.; Li, W. CLDN8 promotes colorectal cancer cell proliferation, migration, and invasion by activating MAPK/ERK signaling. Cancer Manag. Res. 2019, 11, 3741–3751. [Google Scholar] [CrossRef] [Green Version]
- Ashikari, D.; Takayama, K.I.; Obinata, D.; Takahashi, S.; Inoue, S. CLDN8, an androgen-regulated gene, promotes prostate cancer cell proliferation and migration. Cancer Sci. 2017, 108, 1386–1393. [Google Scholar] [CrossRef] [Green Version]
- Stamatovic, S.M.; Johnson, A.M.; Sladojevic, N.; Keep, R.F.; Andjelkovic, A.V. Endocytosis of tight junction proteins and the regulation of degradation and recycling. Ann. N. Y. Acad. Sci. 2017, 1397, 54–65. [Google Scholar] [CrossRef]
- Glatzer, F.; Gschwandtner, M.; Ehling, S.; Rossbach, K.; Janik, K.; Klos, A.; Baumer, W.; Kietzmann, M.; Werfel, T.; Gutzmer, R. Histamine induces proliferation in keratinocytes from patients with atopic dermatitis through the histamine 4 receptor. J. Allergy Clin. Immunol. 2013, 132, 1358–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gschwandtner, M.; Mildner, M.; Mlitz, V.; Gruber, F.; Eckhart, L.; Werfel, T.; Gutzmer, R.; Elias, P.M.; Tschachler, E. Histamine suppresses epidermal keratinocyte differentiation and impairs skin barrier function in a human skin model. Allergy 2013, 68, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Kishioka, C.; Ishinaga, H.; Sakakura, Y.; Majima, Y. Histamine alters gene expression in cultured human nasal epithelial cells. J. Allergy Clin. Immunol. 2001, 107, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Guo, S.; Ranzer, M.J.; DiPietro, L.A. Toll-like receptor 4 has an essential role in early skin wound healing. J. Investig. Dermatol. 2013, 133, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Furuse, M.; Hata, M.; Furuse, K.; Yoshida, Y.; Haratake, A.; Sugitani, Y.; Noda, T.; Kubo, A.; Tsukita, S. Claudin-based tight junctions are crucial for the mammalian epidermal barrier: A lesson from claudin-1-deficient mice. J. Cell Biol. 2002, 156, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Eijssen, L.M.; Jaillard, M.; Adriaens, M.E.; Gaj, S.; de Groot, P.J.; Muller, M.; Evelo, C.T. User-friendly solutions for microarray quality control and pre-processing on ArrayAnalysis.org. Nucleic Acids Res. 2013, 41, W71–W76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, M.; Wang, P.; Boyd, A.D.; Kostov, G.; Athey, B.; Jones, E.G.; Bunney, W.E.; Myers, R.M.; Speed, T.P.; Akil, H.; et al. Evolving gene/transcript definitions significantly alter the interpretation of GeneChip data. Nucleic Acids Res. 2005, 33, e175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. Affy—Analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Phipson, B.; Lee, S.; Majewski, I.J.; Alexander, W.S.; Smyth, G.K. Robust hyperparameter estimation protects against hypervariable genes and improves power to detect differential expression. Ann. Appl. Stat. 2016, 10, 946–963. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Zhao, S.; Guo, Y.; Sheng, Q.; Shyr, Y. Heatmap3: An improved heatmap package with more powerful and convenient features. BMC Bioinform. 2014, 15 (Suppl. 10), P16. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Moffatt-Jauregui, C.E.; Robinson, B.; de Moya, A.V.; Brockman, R.D.; Roman, A.V.; Cash, M.N.; Culp, D.J.; Lamont, R.J. Establishment and characterization of a telomerase immortalized human gingival epithelial cell line. J. Periodontal Res. 2013, 48, 713–721. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Chen, L.; DiPietro, L.A. Production and function of pigment epithelium-derived factor in isolated skin keratinocytes. Exp. Dermatol. 2014, 23, 436–438. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | 6 h | 12 h | 24 h | 72 h | 120 h | 168 h | 240 h |
|---|---|---|---|---|---|---|---|
| Cldn1 | 0.41 | −0.77 | −1.01 | −0.61 | −0.58 | -- | -- |
| Cldn2 | -- | -- | 0.35 | 0.37 | 0.42 | 0.35 | -- |
| Cldn3 | -- | -- | −0.73 | −1.07 | −0.7 | −1.41 | -- |
| Cldn4 | 1 | −2.14 | −1.79 | −1.94 | −1.53 | −1.03 | -- |
| Cldn5 | -- | -- | −0.61 | −0.88 | -- | -- | -- |
| Cldn6 | -- | -- | -- | -- | -- | -- | -- |
| Cldn7 | -- | -- | -- | -- | -- | -- | -- |
| Cldn8 | -- | −2.8 | −2.71 | −2.23 | −1.44 | −1.73 | -- |
| Cldn9 | -- | -- | -- | -- | -- | -- | -- |
| Cldn10 | −0.74 | −2.86 | −2.74 | −2.03 | −0.77 | −0.62 | -- |
| Cldn11 | −0.56 | −0.49 | −0.51 | −0.36 | −0.38 | -- | −0.47 |
| Cldn12 | -- | -- | -- | -- | -- | -- | -- |
| Cldn13 | -- | -- | -- | -- | -- | 0.48 | -- |
| Cldn14 | -- | −0.64 | −0.84 | −0.91 | -- | −0.65 | -- |
| Cldn15 | -- | -- | -- | -- | -- | -- | -- |
| Cldn16 | -- | -- | -- | -- | -- | -- | -- |
| Cldn18 | -- | -- | -- | -- | 0.31 | -- | -- |
| Cldn19 | -- | -- | -- | -- | -- | -- | -- |
| Cldn22 | -- | -- | -- | -- | -- | -- | -- |
| Cldn23 | −1.02 | −0.66 | −1.54 | −1.91 | −1.37 | −0.82 | -- |
| Jam1 | 0.57 | -- | -- | -- | -- | -- | -- |
| Jam2 | −0.47 | −0.61 | −0.36 | -- | -- | -- | -- |
| Jam3 | −0.56 | -- | -- | 0.45 | 0.68 | 0.5 | -- |
| Ocln | -- | −0.75 | −1.41 | −0.77 | −0.58 | −0.61 | -- |
| ZO1 | 0.33 | -- | −0.46 | -- | -- | -- | -- |
| ZO2 | 0.73 | -- | −0.53 | −0.96 | −0.74 | −0.65 | -- |
| ZO3 | −0.75 | -- | −0.78 | -- | -- | -- | -- |
| Gene Symbol | 6 h | 12 h | 24 h | 72 h | 120 h | 168 h | 240 h |
|---|---|---|---|---|---|---|---|
| Cldn1 | -- | -- | -- | -- | -- | -- | -- |
| Cldn2 | -- | -- | 0.39 | 0.46 | 0.66 | 0.76 | 0.62 |
| Cldn3 | -- | -- | -- | -- | -- | -- | -- |
| Cldn4 | -- | −1.1 | −0.97 | −1.63 | −1.51 | -- | -- |
| Cldn5 | -- | -- | -- | −1.11 | −0.84 | -- | -- |
| Cldn6 | -- | -- | -- | -- | -- | -- | -- |
| Cldn7 | -- | -- | -- | -- | -- | -- | -- |
| Cldn8 | -- | -- | -- | -- | -- | -- | -- |
| Cldn9 | -- | -- | -- | -- | -- | -- | -- |
| Cldn10 | -- | −0.62 | -- | −0.61 | -- | -- | -- |
| Cldn11 | -- | -- | -- | -- | -- | -- | -- |
| Cldn12 | -- | -- | -- | -- | -- | -- | -- |
| Cldn13 | -- | -- | -- | -- | 0.47 | 0.47 | -- |
| Cldn14 | -- | -- | -- | -- | -- | -- | -- |
| Cldn15 | 0.72 | 0.71 | 0.51 | -- | -- | -- | -- |
| Cldn16 | -- | -- | -- | -- | -- | -- | -- |
| Cldn18 | -- | -- | -- | -- | -- | -- | -- |
| Cldn19 | -- | -- | -- | -- | -- | -- | -- |
| Cldn22 | -- | -- | -- | -- | -- | -- | -- |
| Cldn23 | -- | -- | -- | -- | -- | -- | -- |
| Jam1 | -- | -- | -- | −0.41 | -- | -- | −0.56 |
| Jam2 | −0.44 | −0.64 | −0.35 | -- | -- | -- | -- |
| Jam3 | -- | −0.42 | -- | -- | 0.43 | -- | -- |
| Ocln | -- | -- | -- | -- | -- | −0.52 | -- |
| ZO1 | -- | -- | -- | -- | -- | -- | -- |
| ZO2 | -- | -- | -- | −0.64 | -- | -- | -- |
| ZO3 | -- | -- | -- | -- | -- | -- | -- |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leonardo, T.R.; Shi, J.; Chen, D.; Trivedi, H.M.; Chen, L. Differential Expression and Function of Bicellular Tight Junctions in Skin and Oral Wound Healing. Int. J. Mol. Sci. 2020, 21, 2966. https://doi.org/10.3390/ijms21082966
Leonardo TR, Shi J, Chen D, Trivedi HM, Chen L. Differential Expression and Function of Bicellular Tight Junctions in Skin and Oral Wound Healing. International Journal of Molecular Sciences. 2020; 21(8):2966. https://doi.org/10.3390/ijms21082966
Chicago/Turabian StyleLeonardo, Trevor R., Junhe Shi, Dandan Chen, Harsh M. Trivedi, and Lin Chen. 2020. "Differential Expression and Function of Bicellular Tight Junctions in Skin and Oral Wound Healing" International Journal of Molecular Sciences 21, no. 8: 2966. https://doi.org/10.3390/ijms21082966
APA StyleLeonardo, T. R., Shi, J., Chen, D., Trivedi, H. M., & Chen, L. (2020). Differential Expression and Function of Bicellular Tight Junctions in Skin and Oral Wound Healing. International Journal of Molecular Sciences, 21(8), 2966. https://doi.org/10.3390/ijms21082966