Wood Architecture and Composition Are Deeply Remodeled in Frost Sensitive Eucalyptus Overexpressing CBF/DREB1 Transcription Factors

 , and
, and
Abstract
:1. Introduction
2. Results
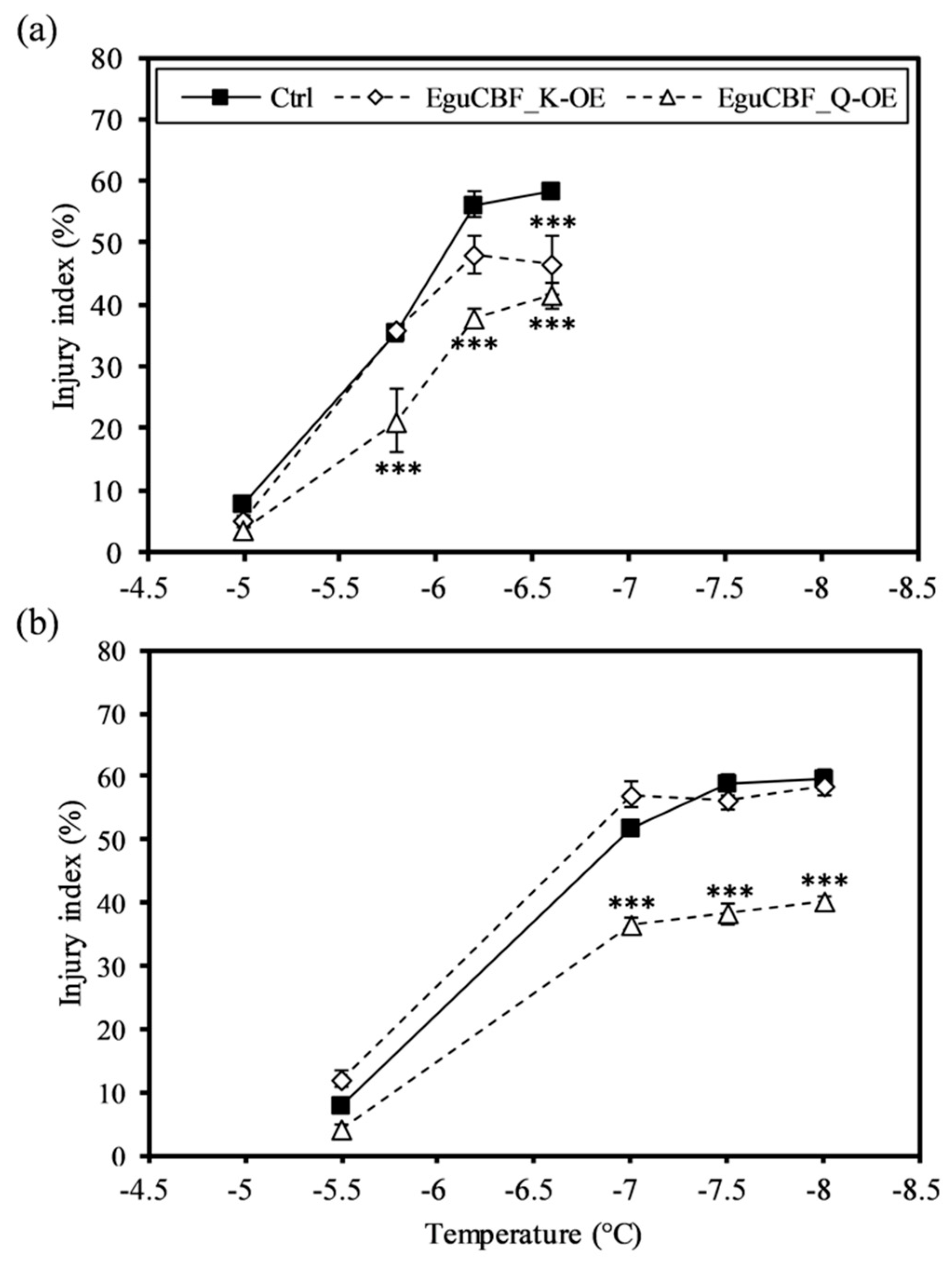
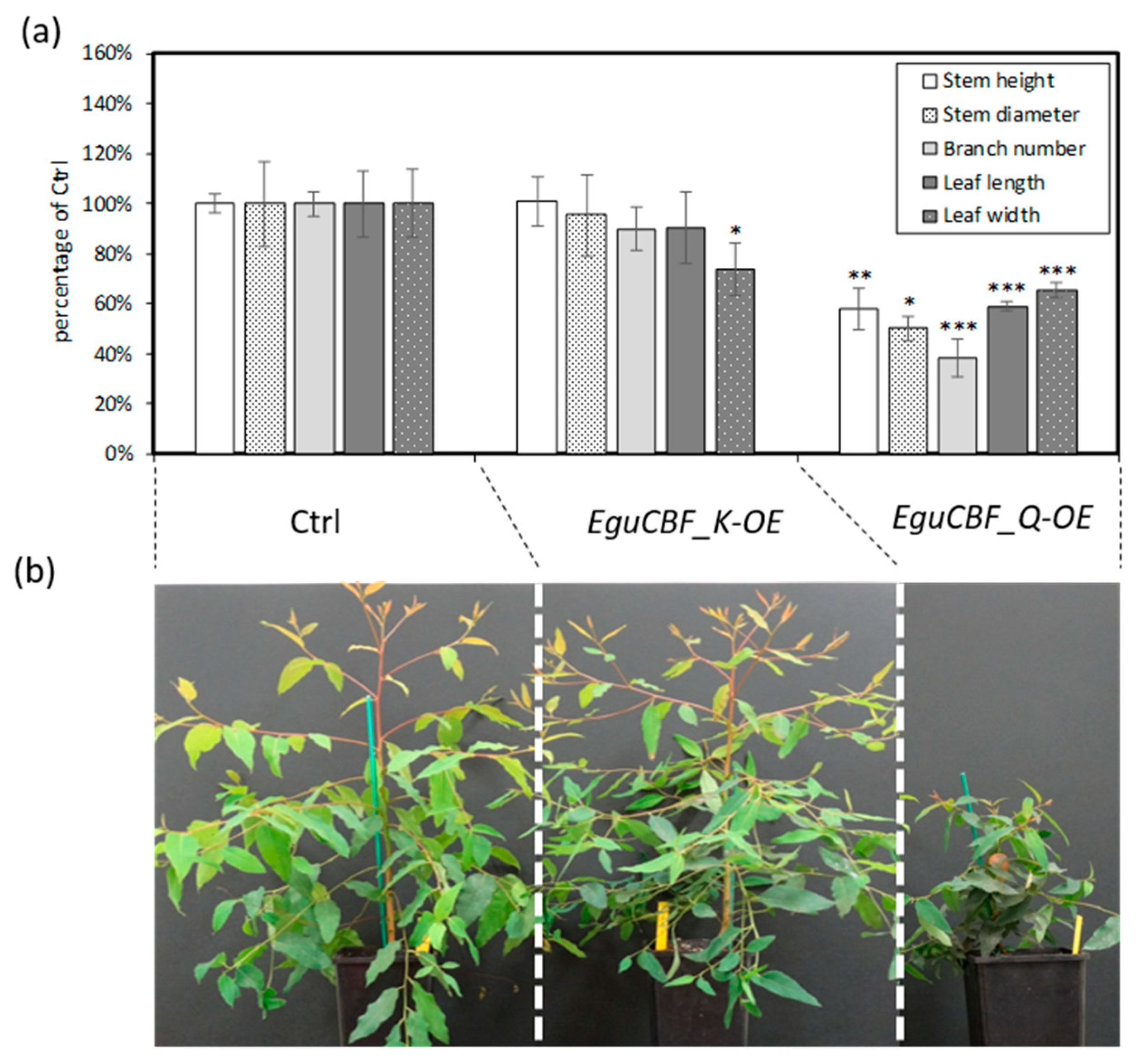
2.1. CBF Overexpression Confers Frost Tolerance to Eucalyptus Transgenic Lines While Negatively Impacting Plant Growth and Development
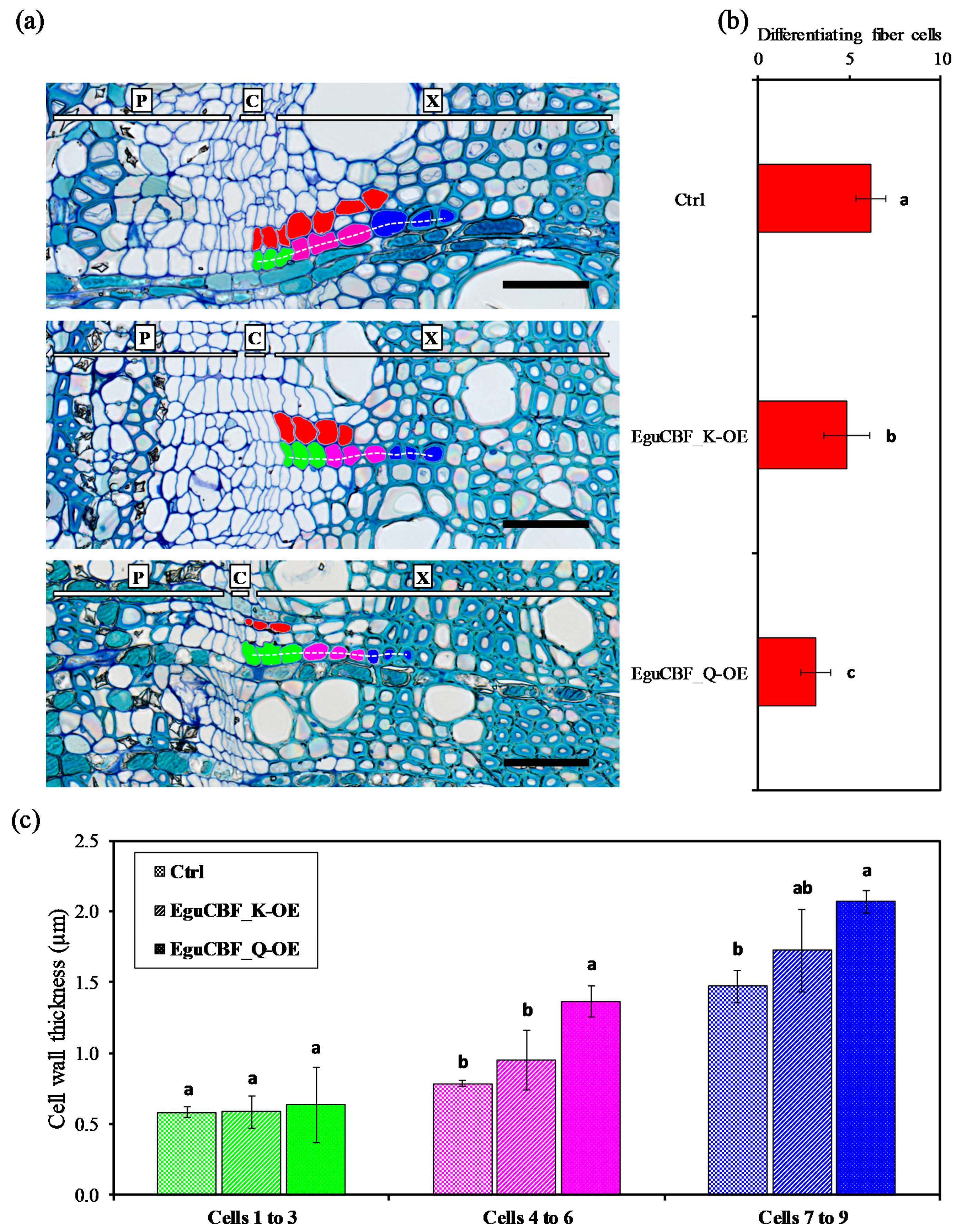
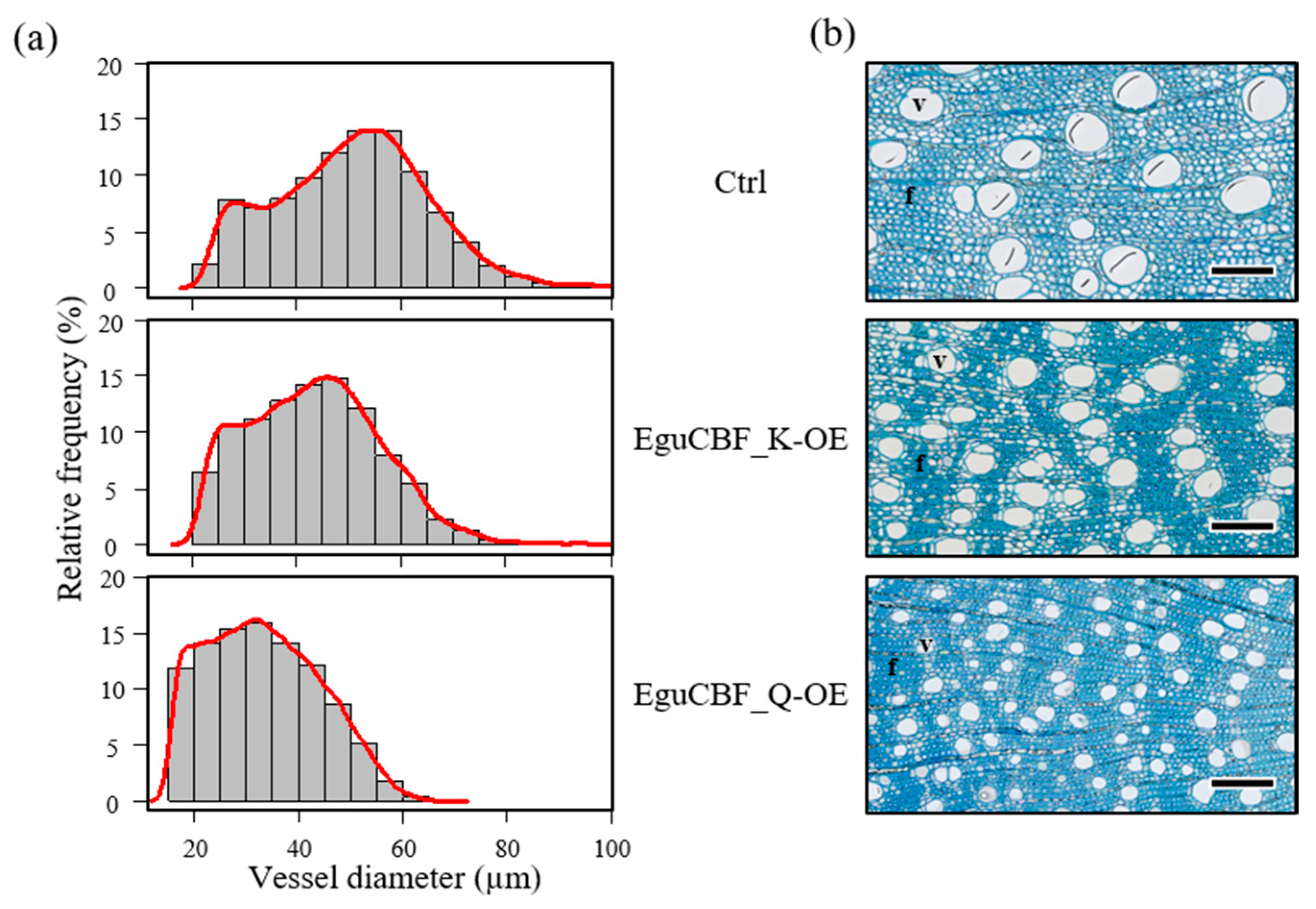
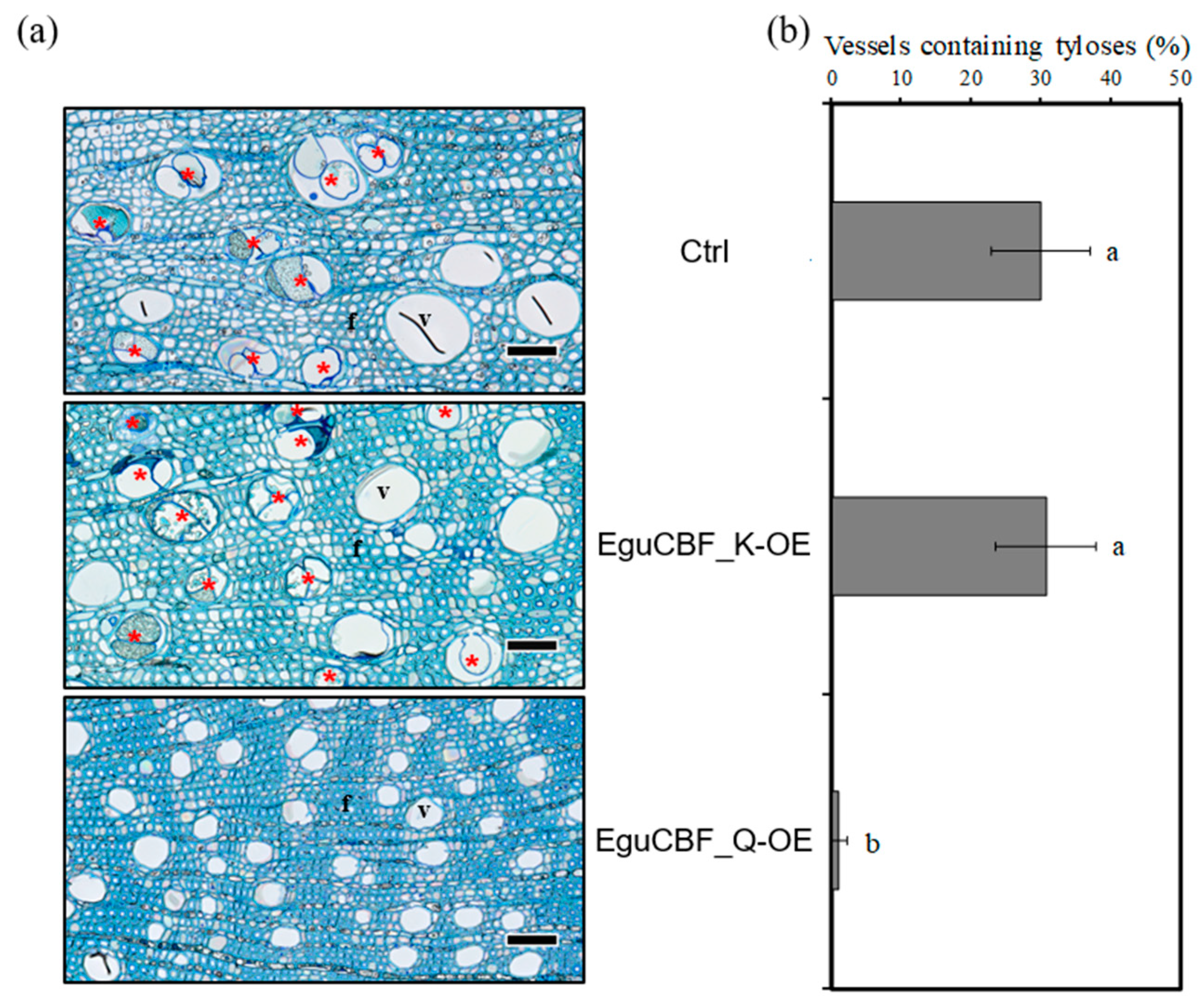
2.2. CBF Overexpression Impacts Xylem Differentiation and Architecture in Eucalyptus Transgenic Lines
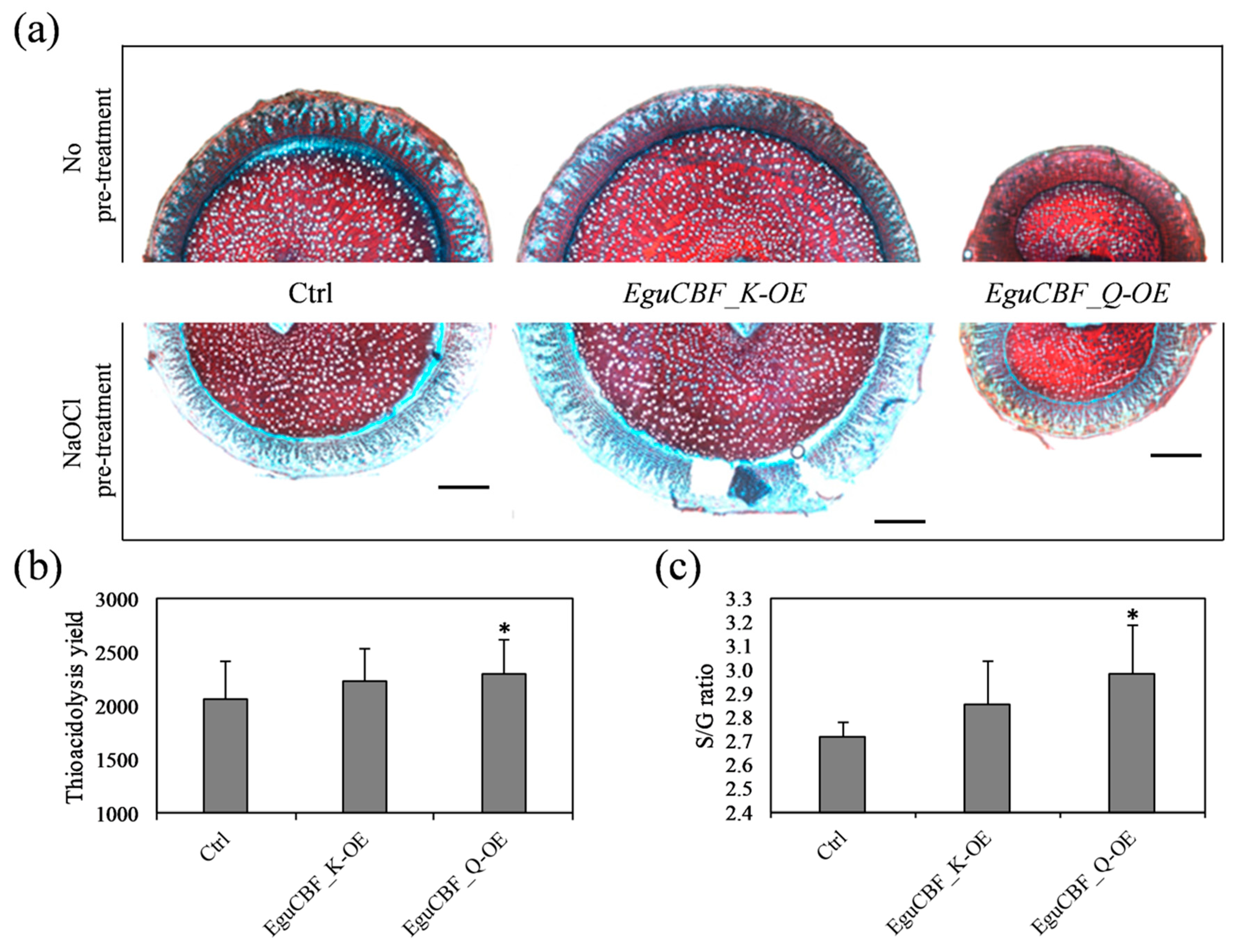
2.3. Both Lignin Content and Composition Are Modified in CBF-Overexpressors
2.4. Most Lignin Biosynthetic Genes Are Up-Regulated in CBF-Overexpressors
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Freezing Tolerance Assessment Using the Ion Leakage Method
4.3. Microscopy and Histochemistry
4.4. Lignin Content and Monomer Composition
4.5. RNA Isolation and RT-qPCR Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 4CL | 4-hydroxycinnamate CoA ligase |
| ANOVA | Analysis of variance |
| AP2/ERF | APETALA2/ethylene-responsive element binding factor |
| BAP | Benzylaminopurine |
| C3H | p-coumarate 3-hydroxylase |
| C4H | Cinnamate-4-hydroxylase |
| CAD | Cinnamyl alcohol dehydrogenase |
| CaMV | Cauliflower mosaic virus |
| CBF | CRT-repeat binding factors |
| DREB | Dehydration responsive element binding protein |
| CCoAOMT | Caffeoyl-CoA 3-O-methyltransferase |
| CCR | Cinnamoyl-CoA reductase |
| cDNA | Complementary DNA |
| COMT | Caffeic acid O-methyltransferase |
| CTAB | Cetyltrimethylammonium bromide |
| Ec | Electrolyte conductivity |
| F5H | Ferulate 5-hydroxylase |
| G | Guaiacyl |
| HCT | Hydroxycinnamoyl transferase |
| HSD | Honestly significant difference |
| IBA | Indol-butyric acid |
| OE | Overexpression |
| PAL | Phenylalanine ammonialyase |
| PCR | Polymerase chain reaction |
| RTqPCR | Real time quantitative polymerase chain reaction |
| S | Syringyl |
| SCW | Secondary cell wall |
| TBO | Toluidine blue O |
| TF | Transcription factor |
| VI | Vulnerability index |
References
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Myburg, A.A.; Grattapaglia, D.; Tuskan, G.A.; Hellsten, U.; Hayes, R.D.; Grimwood, J.; Jenkins, J.; Lindquist, E.; Tice, H.; Bauer, D.; et al. The genome of Eucalyptus grandis. Nature 2014, 510, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Teulieres, C.; Bossinger, G.; Moran, G.; Marque, C. Stress studies in Eucalyptus. Plant Stress. 2007, 1, 197–215. [Google Scholar]
- Ployet, R.; Soler, M.; Carocha, V.; Ladouce, N.; Alves, A.; Rodrigues, J.-C.; Harvengt, L.; Marque, C.; Teulières, C.; Grima-Pettenati, J.; et al. Long cold exposure induces transcriptional and biochemical remodelling of xylem secondary cell wall in Eucalyptus. Tree Physiol. 2018, 38, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Butnor, J.R.; Johnsen, K.H.; Anderson, P.H.; Hall, K.B.; Halman, J.M.; Hawley, G.J.; Maier, C.A.; Schaberg, P.G. Growth, photosynthesis, and cold tolerance of eucalyptus benthamii planted in the piedmont of North Carolina. For. Sci. 2019, 65, 59–67. [Google Scholar]
- Choat, B.; Medek, D.E.; Stuart, S.A.; Pasquet-Kok, J.; Egerton, J.J.G.; Salari, H.; Sack, L.; Ball, M.C. Xylem traits mediate a trade-off between resistance to freeze-thaw-induced embolism and photosynthetic capacity in overwintering evergreens. New Phytol. 2011, 191, 996–1005. [Google Scholar] [CrossRef]
- Mauriat, M.; Le Provost, G.; Rozenberg, P.; Delzon, S.; Breda, N.; Clair, B.; Coutand, C.; Domec, J.-C.; Fourcaud, T.; Grima-Pettenati, J.; et al. Wood formation in trees. In Tree Biotechnology; Ramawat, K.G., Merillon, J.M., Ahuja, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2014; p. 656. ISBN 9781466597143. [Google Scholar]
- Zinkgraf, M.; Liu, L.; Groover, A.; Filkov, V. Identifying gene coexpression networks underlying the dynamic regulation of wood-forming tissues in Populus under diverse environmental conditions. New Phytol. 2017, 214, 1464–1478. [Google Scholar] [CrossRef] [Green Version]
- Pfautsch, S.; Harbusch, M.; Wesolowski, A.; Smith, R.; Macfarlane, C.; Tjoelker, M.G.; Reich, P.B.; Adams, M.A. Climate determines vascular traits in the ecologically diverse genus Eucalyptus. Ecol. Lett. 2016, 19, 240–248. [Google Scholar] [CrossRef]
- Camargo, E.L.O.; Nascimento, L.C.; Soler, M.; Salazar, M.M.; Lepikson-Neto, J.; Marques, W.L.; Alves, A.; Teixeira, P.J.P.L.; Mieczkowski, P.; Carazzolle, M.F.; et al. Contrasting nitrogen fertilization treatments impact xylem gene expression and secondary cell wall lignification in Eucalyptus. BMC Plant Biol. 2014, 14, 256. [Google Scholar] [CrossRef] [Green Version]
- Paux, E.; Carocha, V.; Marques, C.; De Sousa, A.M.; Borralho, N.; Sivadon, P.; Grima-Pettenati, J. Transcript profiling of Eucalyptus xylem genes during tension wood formation. New Phytol. 2005, 167, 89–100. [Google Scholar] [CrossRef]
- Ye, Z.-H.; Zhong, R. Molecular control of wood formation in trees. J. Exp. Bot. 2015, 66, 4119–4131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camargo, E.L.O.; Ployet, R.; Cassan-Wang, H.; Mounet, F.; Grima-Pettenati, J. Digging in wood: New insights in the regulation of wood formation in tree species. Adv. Bot. Res. 2018, 89, 201–233. [Google Scholar]
- Taylor-Teeples, M.; Lin, L.; de Lucas, M.; Turco, G.; Toal, T.W.; Gaudinier, A.; Young, N.F.; Trabucco, G.M.; Veling, M.T.; Lamothe, R.; et al. An Arabidopsis gene regulatory network for secondary cell wall synthesis. Nature 2015, 517, 571–575. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Wang, Y.; Wang, L.; Hu, P.; Wang, Y.; Jia, Y.; Zhang, C.; Zhang, Y.; Zhang, Y.; Wang, C.; et al. Expression of the MYB transcription factor gene BplMYB46 affects abiotic stress tolerance and secondary cell wall deposition in Betula platyphylla. Plant Biotechnol. J. 2017, 15, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Fu, X.; Liu, R.; Guo, L.; Ran, L.; Li, C.; Tian, Q.; Jiao, B.; Wang, B.; Luo, K.; et al. PtoMYB170 positively regulates lignin deposition during wood formation in poplar and confers drought tolerance in transgenic Arabidopsis. Tree Physiol. 2017, 37, 1713–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Lee, C.-M.; Doherty, C.J.; Gilmour, S.J.; Kim, Y.; Thomashow, M.F. Regulation of the Arabidopsis CBF regulon by a complex low-temperature regulatory network. Plant J. 2015, 82, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Lata, C.; Prasad, M. Role of DREBs in regulation of abiotic stress responses in plants. J. Exp. Bot. 2011, 62, 4731–4748. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, M.; Jaiswal, A.; Taj, G.; Jaiswal, J.P.; Qureshi, M.I.; Singh, N.K. DREB1/CBF transcription factors: Their structure, function and role in abiotic stress tolerance in plants. J. Genet. 2012, 91, 385–395. [Google Scholar] [CrossRef]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta-Gene Regul. Mech. 2012, 1819, 86–96. [Google Scholar] [CrossRef]
- Navarro, M.; Ayax, C.; Martinez, Y.; Laur, J.; El Kayal, W.; Marque, C.; Teulières, C. Two EguCBF1 genes overexpressed in Eucalyptus display a different impact on stress tolerance and plant development. Plant Biotechnol. J. 2011, 9, 50–63. [Google Scholar] [CrossRef]
- Cao, P.B.; Azar, S.; SanClemente, H.; Mounet, F.; Dunand, C.; Marque, G.; Marque, C.; Teulières, C. Genome-wide analysis of the AP2/ERF family in eucalyptus grandis: An intriguing over-representation of stress-responsive DREB1/CBF genes. PLoS ONE 2015, 10, e0121041. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.C.; Cao, P.B.; San Clemente, H.; Ployet, R.; Mounet, F.; Ladouce, N.; Harvengt, L.; Marque, C.; Teulieres, C. Special trends in CBF and DREB2 groups in Eucalyptus gunnii vs. Eucalyptus grandis suggest that CBF are master players in the trade-off between growth and stress resistance. Physiol. Plant. 2017, 159, 445–467. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.; Marque, G.; Ayax, C.; Keller, G.; Borges, J.P.; Marque, C.; Teulières, C. Complementary regulation of four Eucalyptus CBF genes under various cold conditions. J. Exp. Bot. 2009, 60, 2713–2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffen, K.L.; Arora, R.; Palta, J.P. Relative sensitivity of photosynthesis and respiration to freeze-thaw stress in herbaceous species. Plant Physiol. 1989, 89, 1372–1379. [Google Scholar] [CrossRef] [Green Version]
- Kolb, K.J.; Sperry, J.S. Differences in drought adaptation between subspecies of sagebrush (Artemisia tridentata). Ecology 1999, 80, 2373–2384. [Google Scholar] [CrossRef] [Green Version]
- Hacke, U.G.; Sperry, J.S.; Wheeler, J.K.; Castro, L. Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol. 2006, 26, 689–701. [Google Scholar] [CrossRef]
- Tixier, A.; Cochard, H.; Badel, E.; Dusotoit-Coucaud, A.; Jansen, S.; Herbette, S. Arabidopsis thaliana as a model species for xylem hydraulics: Does size matter? J. Exp. Bot. 2013, 64, 2295–2305. [Google Scholar] [CrossRef] [Green Version]
- Arbellay, E.; Fonti, P.; Stoffel, M. Duration and extension of anatomical changes in wood structure after cambial injury. J. Exp. Bot. 2012, 63, 3271–3277. [Google Scholar] [CrossRef] [Green Version]
- Méchin, V.; Laluc, A.; Legée, F.; Cézard, L.; Denoue, D.; Barrière, Y.; Lapierre, C. Impact of the brown-midrib bm 5 mutation on maize lignins. J. Agric. Food Chem. 2014, 62, 5102–5107. [Google Scholar] [CrossRef]
- Carocha, V.; Soler, M.; Hefer, C.; Cassan-Wang, H.; Fevereiro, P.; Myburg, A.A.; Paiva, J.A.P.; Grima-Pettenati, J. Genome-wide analysis of the lignin toolbox of Eucalyptus grandis. New Phytol. 2015, 206, 1297–1313. [Google Scholar] [CrossRef] [Green Version]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Kayal, W.; Navarro, M.; Marque, G.; Keller, G.; Marque, C.; Teulieres, C. Expression profile of CBF-like transcriptional factor genes from Eucalyptus in response to cold. J. Exp. Bot. 2006, 57, 2455–2469. [Google Scholar] [CrossRef] [PubMed]
- Bhalerao, R.P.; Fischer, U. Environmental and hormonal control of cambial stem cell dynamics. J. Exp. Bot. 2017, 68, 79–87. [Google Scholar] [CrossRef]
- Bourne, A.E.; Creek, D.; Peters, J.M.R.; Ellsworth, D.S.; Choat, B. Species climate range influences hydraulic and stomatal traits in Eucalyptus species. Ann. Bot. 2017, 120, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Xu, H.; Li, H.; Wei, D.; Lin, J.; Li, X. Seasonal development of cambial activity in relation to xylem formation in Chinese fir. J. Plant Physiol. 2016, 195, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Schrader, J.; Nilsson, J.; Mellerowicz, E.; Berglund, A.; Nilsson, P.; Hertzberg, M.; Sandberg, G. A high-resolution transcript profile across the wood-forming meristem of poplar identifies potential regulators of cambial stem cell identity. Plant Cell 2004, 16, 2278–2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendall, S.L.; Hellwege, A.; Marriot, P.; Whalley, C.; Graham, I.A.; Penfield, S. Induction of dormancy in Arabidopsis summer annuals requires parallel regulation of DOG1 and hormone metabolism by low temperature and CBF transcription factors. Plant Cell 2011, 23, 2568–2580. [Google Scholar] [CrossRef] [Green Version]
- Wisniewski, M.; Norelli, J.; Bassett, C.; Artlip, T.; Macarisin, D. Ectopic expression of a novel peach (Prunus persica) CBF transcription factor in apple (Malus × domestica) results in short-day induced dormancy and increased cold hardiness. Planta 2011, 233, 971–983. [Google Scholar] [CrossRef]
- Wisniewski, M.; Norelli, J.; Artlip, T. Overexpression of a peach CBF gene in apple: A model for understanding the integration of growth, dormancy, and cold hardiness in woody plants. Front. Plant Sci. 2015, 6, 85. [Google Scholar] [CrossRef] [Green Version]
- Artlip, T.S.; Wisniewski, M.E.; Bassett, C.L.; Norelli, J.L. CBF gene expression in peach leaf and bark tissues is gated by a circadian clock. Tree Physiol. 2013, 33, 866–877. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zhang, Z.; Xie, S.; Si, T.; Li, Y.; Zhu, J.K. Mutational evidence for the critical role of CBF transcription factors in cold acclimation in Arabidopsis. Plant Physiol. 2016, 171, 2744–2759. [Google Scholar] [PubMed] [Green Version]
- Vander Willigen, C.; Pammenter, N.W. Relationship between growth and xylem hydraulic characteristics of clones of Eucalyptus spp. at contrasting sites. Tree Physiol. 1997, 18, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, A.L.; Ewers, F.W.; Pratt, R.B.; Paddock, W.A.; Davis, S.D. Do xylem fibers affect vessel cavitation resistance? Plant Physiol. 2005, 139, 546–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barotto, A.J.; Monteoliva, S.; Gyenge, J.; Martinez-Meier, A.; Fernandez, M.E. Functional relationships between wood structure and vulnerability to xylem cavitation in races of Eucalyptus globulus differing in wood density. Tree Physiol. 2018, 38, 243–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gall, H.; Philippe, F.; Domon, J.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar]
- Bajji, M.; Kinet, J.M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Carlquist, S. Ecological factors in wood evolution: A floristic approach. Am. J. Bot. 1977, 64, 887–896. [Google Scholar] [CrossRef]
- James, S.A.; Meinzer, F.C.; Goldstein, G.; Woodruff, D.; Jones, T.; Restom, T.; Mejia, M.; Clearwater, M.; Campanello, P. Axial and radial water transport and internal water storage in tropical forest canopy trees. Oecologia 2003, 134, 37–45. [Google Scholar] [CrossRef]
- Hajek, P.; Kurjak, D.; von Wühlisch, G.; Delzon, S.; Schuldt, B. Intraspecific variation in wood anatomical, hydraulic, and foliar traits in ten european beech provenances differing in growth yield. Front. Plant Sci. 2016, 7, 791. [Google Scholar] [CrossRef] [Green Version]
- Cassan-Wang, H.; Soler, M.; Yu, H.; Camargo, E.L.O.; Carocha, V.; Ladouce, N.; Savelli, B.; Paiva, J.A.P.; Leplé, J.-C.; Grima-Pettenati, J. Reference genes for high-throughput quantitative reverse transcription-PCR analysis of gene expression in organs and tissues of Eucalyptus grown in various environmental conditions. Plant Cell Physiol. 2012, 53, 2101–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D.; Kong, Y.; Kong, D.; Gao, Q.; Zhou, G.; Sandberg, G.; Tuominen, H.; Moritz, T.; Bhalerao, R.; et al. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | EguCBF_K-OE | EguCBF_Q-OE | |
|---|---|---|---|
| Vessel diameter (µm) | 52.7 ± 3.4 a | 44.9 ± 4.3 b | 34.6 ± 5.4 c |
| Vessel density (N/mm−2) | 67.4 ± 10.8 c | 103.6 ± 18.2 b | 169.9 ± 39.5 a |
| Porosity (%) | 14.5 ± 1.4 b | 16.1 ± 1.5 a | 15.4 ± 2.6 ab |
| Average vessel hydraulic mean diameter (µm) | 62.8 ± 4.7 a | 54.2 ± 6.6 b | 41.4 ± 6.4 c |
| Theoretical area-specific hydraulic conductivity (kg/m−1/MPa−1/s−1) | 15.8 ± 2.9 a | 13.1 ± 3.6 b | 7.7 ± 3.2 c |
| Potential vulnerability index | 0.81 ± 0.16 a | 0.45 ± 0.11 b | 0.22 ± 0.08 c |
| Control | EguCBF_K-OE | EguCBF_Q-OE | |
|---|---|---|---|
| Fiber lumen diameter (µm) | 5.51 ± 0.62 a | 5.05 ± 0.74 b | 4.17 ± 0.41 c |
| Fiber density (N/mm−2) | 9793.1 ± 1639.2 c | 10594.6 ± 1684.9 b | 13378.7 ± 2214.1 a |
| Fiber cell wall thickness (µm) | 3.00 ± 0.49 a | 3.01 ± 0.70 a | 2.85 ± 0.47 a |
| Cell wall area fraction (%) | 58.19 ± 2.99 b | 56.66 ± 5.37 b | 63.19 ± 5.21 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, P.B.; Ployet, R.; Nguyen, C.; Dupas, A.; Ladouce, N.; Martinez, Y.; Grima-Pettenati, J.; Marque, C.; Mounet, F.; Teulières, C. Wood Architecture and Composition Are Deeply Remodeled in Frost Sensitive Eucalyptus Overexpressing CBF/DREB1 Transcription Factors. Int. J. Mol. Sci. 2020, 21, 3019. https://doi.org/10.3390/ijms21083019
Cao PB, Ployet R, Nguyen C, Dupas A, Ladouce N, Martinez Y, Grima-Pettenati J, Marque C, Mounet F, Teulières C. Wood Architecture and Composition Are Deeply Remodeled in Frost Sensitive Eucalyptus Overexpressing CBF/DREB1 Transcription Factors. International Journal of Molecular Sciences. 2020; 21(8):3019. https://doi.org/10.3390/ijms21083019
Chicago/Turabian StyleCao, Phi Bang, Raphaël Ployet, Chien Nguyen, Annabelle Dupas, Nathalie Ladouce, Yves Martinez, Jacqueline Grima-Pettenati, Christiane Marque, Fabien Mounet, and Chantal Teulières. 2020. "Wood Architecture and Composition Are Deeply Remodeled in Frost Sensitive Eucalyptus Overexpressing CBF/DREB1 Transcription Factors" International Journal of Molecular Sciences 21, no. 8: 3019. https://doi.org/10.3390/ijms21083019
APA StyleCao, P. B., Ployet, R., Nguyen, C., Dupas, A., Ladouce, N., Martinez, Y., Grima-Pettenati, J., Marque, C., Mounet, F., & Teulières, C. (2020). Wood Architecture and Composition Are Deeply Remodeled in Frost Sensitive Eucalyptus Overexpressing CBF/DREB1 Transcription Factors. International Journal of Molecular Sciences, 21(8), 3019. https://doi.org/10.3390/ijms21083019