Human Ovarian Cortex biobanking: A Fascinating Resource for Fertility Preservation in Cancer

 , ,
, ,  , , ,
, , , 
Abstract
:1. Introduction
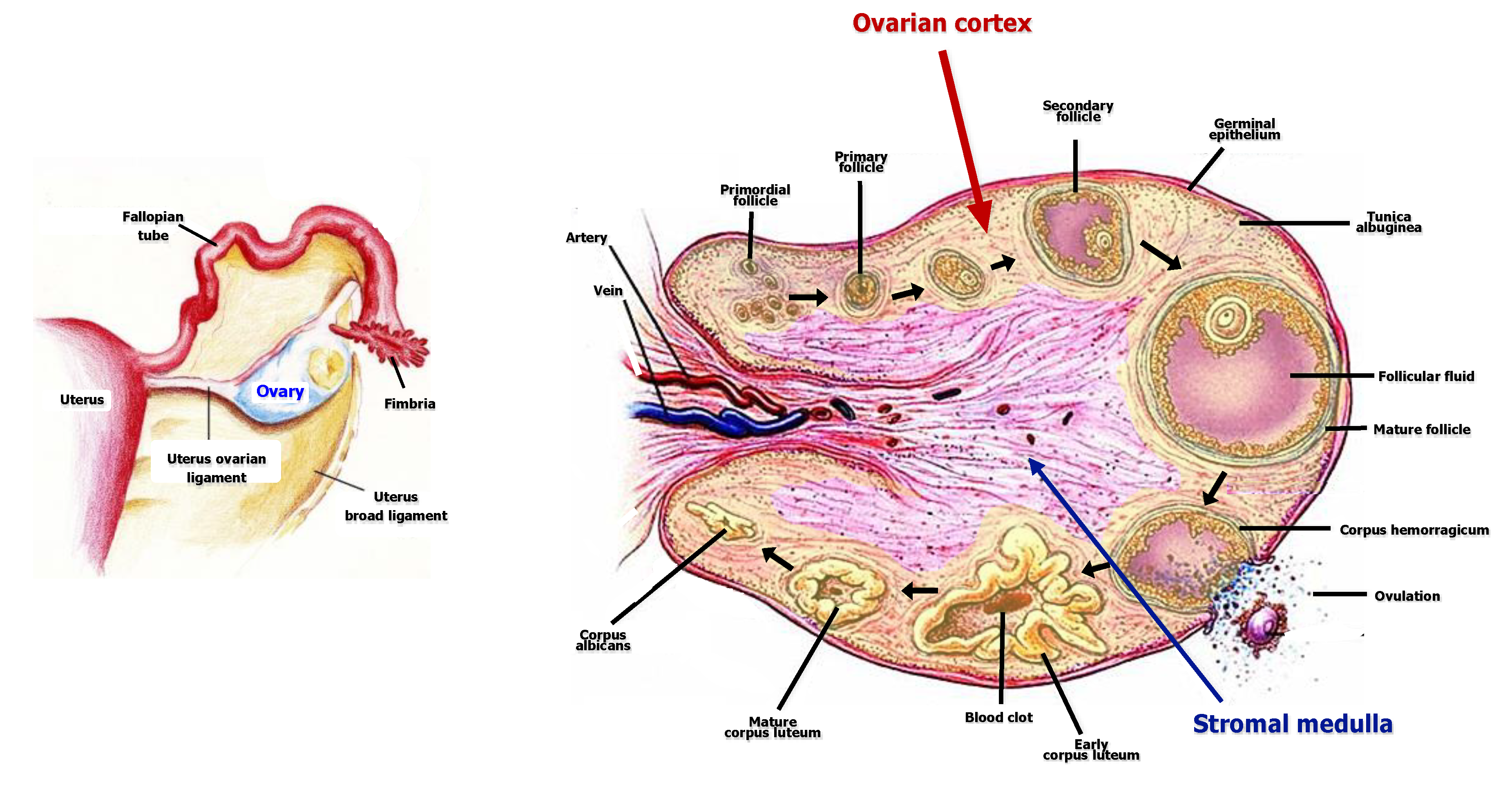
2. Ovarian Structure and Components
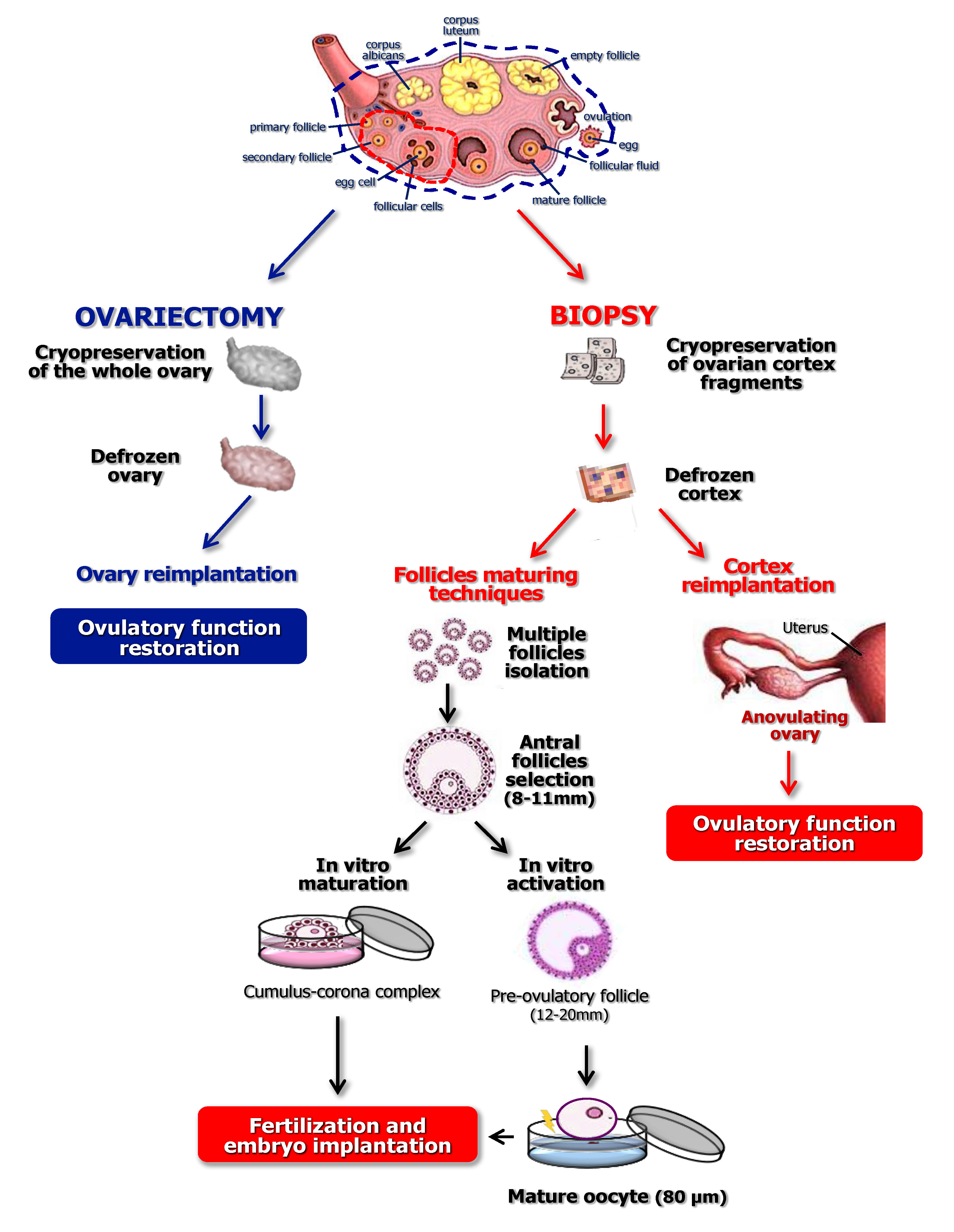
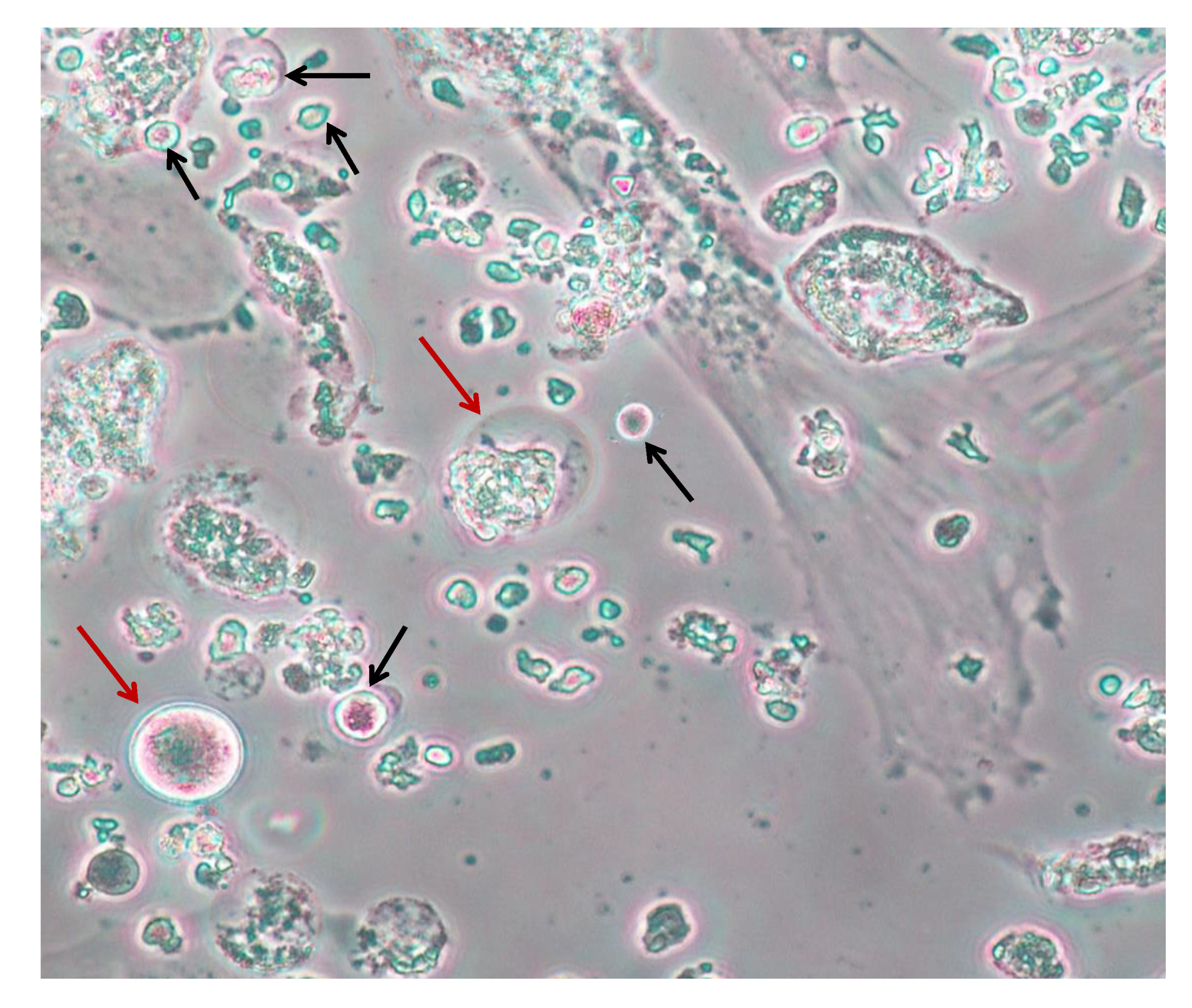
3. Current Techniques Using Cryopreserved Ovarian Cortex
4. The Ovarian Cortex Transplantation to Prevent the Cancer Treatment Induced Infertility
5. Ovarian Stem Cells (OSCs): A Novel Resource of the Ovarian Cortex
6. Potential Utilization of OSCs for Female Fertility
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| FP | fertility preservation |
| OR | ovarian reserve |
| GnRHa | gonadotropin releasing hormone agonist |
| COS | controlled ovarian stimulation |
| MFG | multiple follicular growth |
| OSCs | ovarian stem cells |
| OLCs | oocyte-like cells |
| POI | premature ovarian failure |
| PGCs | primordial germ cells |
| Bmp2 | bone morphogenetic protein 2 |
| GCs | granulosa cells |
| UGCs | undifferentiated granulosa cells |
| Wnt4 | Wingless-type MMTV integration site family member 4 |
| Foxl2 | Forkhead box L2 |
| FSH-R | Follicle stimulating hormone receptor |
| OTC | Ovarian tissue cryopreservation |
| IVM | in vitro maturation |
| IVA | in vitro activation |
| Ddx4 | DEAD box polypeptide 4 |
| ddPCR | droplet digital PCR |
| PSC | pluripotent stem cells |
References
- Inhorn, M.; Patrizio, P. Infertility around the globe: New thinking on gender, reproductive technologies and global movements in the 21st century. Hum. Reprod. Update 2015, 21, 411–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Cancer Society. Cancer Facts & Figures 2013. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2013.html (accessed on 17 January 2013).
- Molina, J.; Barton, D.; Loprinzi, C. Chemotherapy-induced ovarian failure - Manifestations and management. Drug Saf. 2005, 28, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Akahori, T.; Woods, D.C.; Tilly, J.L. Female Fertility Preservation through Stem Cell-based Ovarian Tissue Reconstitution in Vitro and Ovarian Regeneration in Vivo. Clin. Med. Insights Reprod. Health 2019, 13, 1179558119848007. [Google Scholar] [CrossRef] [PubMed]
- Bedoschi, G.; Navarro, P.; Oktay, K. Chemotherapy-induced damage to ovary: Mechanisms and clinical impact. Future Oncol. 2016, 12, 2333–2344. [Google Scholar] [CrossRef] [Green Version]
- Blumenfeld, Z. How to preserve fertility in young women exposed to chemotherapy? The role of GnRH agonist cotreatment in addition to cryopreservation of embrya, oocytes, or ovaries. Oncologist 2007, 12, 1044–1054. [Google Scholar] [CrossRef]
- Loren, A.W.; Mangu, P.B.; Beck, L.N.; Brennan, L.; Magdalinski, A.J.; Partridge, A.H.; Quinn, G.; Wallace, W.H.; Oktay, K. American Society of Clinical Oncology. Fertility preservation for patients with cancer: American society of clinical oncology clinical practice guideline update. J. Clin. Oncol. 2013, 31, 2500–2510. [Google Scholar] [CrossRef]
- Peccatori, F.A.; Azim, H.A., Jr.; Orecchia, R.; Pavlidis, N. ESMO Guidelines Working Group. Cancer, pregnancy and fertility: ESMO clinical practice guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2013, 24 (Suppl. 6), vi160–vi170. [Google Scholar] [CrossRef]
- Parmegiani, L.; Bertocci, F.; Garello, C.; Salvarani, M.; Tambuscio, G.; Fabbri, R. Efficiency of human oocyte slow freezing: Results from five assisted reprodction centres. Reprod. Biomed. Online 2009, 18, 352–359. [Google Scholar] [CrossRef]
- Annan, J.J.; Gudi, A.; Bhide, P.; Shah, A.; Homburg, R. Biochemical pregnancy during assisted conception: A little bit pregnant. J. Clin. Med. Res. 2013, 5, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, V.; Lappi, M.; Bonu, M.; Borini, A. Oocyte slow freezing using a 0.2-0.3 M sucrose concentration protocol: Is it really the time to trash the cryopreservation machine? Fertil. Steril. 2012, 97, 1101–1107. [Google Scholar] [CrossRef]
- Lim, J.-H.; Chian, R.-C. In vitro maturation of human immature oocytes. J. Reprod. Stem Cell Biotechnol. 2010, 1, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Silber, S.; Kagawa, N.; Kuwayama, M.; Gosden, R. Duration of fertility after fresh and frozen ovary transplantation. Fertil. Steril. 2010, 94, 2191–2196. [Google Scholar] [CrossRef] [PubMed]
- Von Wolff, M.; Donnez, J.; Hovatta, O.; Keros, V.; Maltaris, T.; Montag, M.; Salle, B.; Sonmezer, M.; Andersen, C. Cryopreservation and autotransplantation of human ovarian tissue prior to cytotoxic therapy - A technique in its infancy but already successful in fertility preservation. Eur. J. Cancer 2009, 45, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Canning, J.; Kaneko, T.; Pru, J.; Tilly, J. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature 2004, 428, 145–150. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, L.; Kang, J.; Xie, W.; Li, X.; Wu, C.; Xu, B.; Wu, J. Production of fat-1 transgenic rats using a post-natal female germline stem cell line. Mol. Hum. Reprod. 2014, 20, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Wylie, C. Germ cells. Cell 1999, 96, 165–174. [Google Scholar] [CrossRef]
- Saitou, M.; Payer, B.; O’Carroll, D.; Ohinata, Y.; Surani, M. Blimp1 and the emergence of the germ line during development in the mouse. Cell Cycle 2005, 4, 1736–1740. [Google Scholar] [CrossRef] [Green Version]
- Lawson, K.; Dunn, N.; Roelen, B.; Zeinstra, L.; Davis, A.; Wright, C.; Korving, J.; Hogan, B. Bmp4 is required for the generation of primordial germ cells in the mouse embryo. Genes Dev. 1999, 13, 424–436. [Google Scholar] [CrossRef]
- Ying, Y.; Qi, X.; Zhao, G. Induction of primordial germ cells from murine epiblasts by synergistic action of BMP4 and BMP8B signaling pathways. Proc. Natl. Acad. Sci. USA 2001, 98, 7858–7862. [Google Scholar] [CrossRef] [Green Version]
- Ying, Y.; Zhao, G. Cooperation of endoderm-derived BMP2 and extraembryonic ectoderm-derived BMP4 in primordial germ cell generation in the mouse. Dev. Biol. 2001, 232, 484–492. [Google Scholar] [CrossRef] [Green Version]
- Marlow, F. Primordial Germ Cell Specification and Migration. F1000Res 2015, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, F.W.; Wilson, J.D. Conversion of androgen to estrogen by the human fetal ovary. J. Clin. Endocrinol. Metab. 1978, 47, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Saito, D. Epithelial-to-mesenchymal transition-based morphogenesis of dorsal mesentery and gonad. Semin. Cell Dev. Biol. 2019, 92, 105–112. [Google Scholar] [CrossRef] [PubMed]
- McLaren, A. Germ and somatic cell lineages in the developing gonad. Mol. Cell. Endocrinol. 2000, 163, 3–9. [Google Scholar] [CrossRef]
- Hansen, K.; Knowlton, N.; Thyer, A.; Charleston, J.; Soules, M.; Klein, N. A new model of reproductive aging: The decline in ovarian non-growing follicle number from birth to menopause. Hum. Reprod. 2008, 23, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Findlay, J.; Hutt, K.; Hickey, M.; Anderson, R. How Is the Number of Primordial Follicles in the Ovarian Reserve Established? Biol. Reprod. 2015, 93. [Google Scholar] [CrossRef]
- Faddy, M.; Gosden, R.; Gougeon, A.; Richardson, S.; Nelson, J. Accelerated disappearance of ovarian follicles in midlife - implications for forecasting menopause. Hum. Reprod. 1992, 7, 1342–1346. [Google Scholar] [CrossRef]
- Choi, J.; Yoo, H. Control of puberty: Genetics, endocrinology, and environment. Curr. Opin. Endocrinol. Diabetes Obes. 2013, 20, 62–68. [Google Scholar] [CrossRef]
- Horan, C.; Williams, S. Oocyte stem cells: Fact or fantasy? Reproduction 2017, 154, R23–R35. [Google Scholar] [CrossRef] [Green Version]
- Woods, D.C.; Tilly, J.L. Germline stem cells in adult mammalian ovaries. Ten Crit. Top. Reprod. Med. 2013, 10–12. [Google Scholar]
- Donnez, J.; Dolmans, M. Ovarian cortex transplantation: 60 reported live births brings the success and worldwide expansion of the technique towards routine clinical practice. J. Assist. Reprod. Genet. 2015, 32, 1167–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, K.; Chian, R. Maturation in vitro of immature human oocytes for clinical use. Hum. Reprod. Update 1998, 4, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Jeruss, J.; Woodruff, T. Current Concepts: Preservation of Fertility in Patients with Cancer. N. Engl. J. Med. 2009, 360, 902–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.; Mamsen, L.; Kristensen, S. Freezing of ovarian tissue and clinical opportunities. Reproduction 2019, 158, F27–F34. [Google Scholar] [CrossRef]
- Carrillo, L.; Seidman, D.; Cittadini, E.; Meirow, D. The role of fertility preservation in patients with endometriosis. J. Assist. Reprod. Genet. 2016, 33, 317–323. [Google Scholar] [CrossRef]
- Jeve, Y.B.; Gelbaya, T.; Fatum, M. Time to consider ovarian tissue cryopreservation for girls with Turner’s syndrome: An opinion paper. Hum. Reprod. Open 2019, 2019, hoz016. [Google Scholar] [CrossRef] [Green Version]
- Jensen, A.; Rechnitzer, C.; Macklon, K.; Ifversen, M.; Birkebaek, N.; Clausen, N.; Sorensen, K.; Fedder, J.; Ernst, E.; Andersen, C. Cryopreservation of ovarian tissue for fertility preservation in a large cohort of young girls: Focus on pubertal development. Hum. Reprod. 2017, 32, 154–164. [Google Scholar] [CrossRef]
- Oktay, K.; Karlikaya, G. Ovarian function after transplantation of frozen, banked autologous ovarian tissue. N. Engl. J. Med. 2000, 342, 1919. [Google Scholar] [CrossRef]
- Donnez, J.; Dolmans, M.; Demylle, D.; Jadoul, P.; Pirard, C.; Squifflet, J.; Martinez-Madrid, B.; van Langendonckt, A. Livebirth after orthotopic transplantation of cryopreserved ovarian tissue. Lancet 2004, 364, 1405–1410. [Google Scholar] [CrossRef]
- Revelli, A.; Marchino, G.; Dolfin, E.; Molinari, E.; Delle Piane, L.; Salvagno, F.; Benedetto, C. Live birth after orthotopic grafting of autologous cryopreserved ovarian tissue and spontaneous conception in Italy. Fertil. Steril. 2013, 99, 227–230. [Google Scholar] [CrossRef]
- Donnez, J.; Dolmans, M. Fertility Preservation in Women. N. Engl. J. Med. 2017, 377, 1657–1665. [Google Scholar] [CrossRef] [PubMed]
- Gellert, S.; Pors, S.; Kristensen, S.; Bay-Bjorn, A.; Ernst, E.; Andersen, C. Transplantation of frozen-thawed ovarian tissue: An update on worldwide activity published in peer-reviewed papers and on the Danish cohort. J. Assist. Reprod. Genet. 2018, 35, 561–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, F. International Society for Fertility Preservation–ESHRE–ASRM Expert Working Group2. Update on fertility preservation from the Barcelona International Society for Fertility Preservation-ESHRE-ASRM 2015 expert meeting: Indications, results and future perspectives. Fertil. Steril. 2017, 108, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Donnez, J.; Silber, S.; Andersen, C.; Demeestere, I.; Piver, P.; Meirow, D.; Pellicer, A.; Dolmans, M. Children born after autotransplantation of cryopreserved ovarian tissue. A review of 13 live births. Ann. Med. 2011, 43, 437–450. [Google Scholar] [CrossRef]
- Meirow, D.; Levron, J.; Eldar-Geva, T.; Hardan, I.; Fridman, E.; Zalel, Y.; Schiff, E.; Dor, J. Pregnancy after transplantation of cryopreserved ovarian tissue in a patient with ovarian failure after chemotherapy. N. Engl. J. Med. 2005, 353, 318–321. [Google Scholar] [CrossRef] [Green Version]
- Dolmans, M.; Martinez-Madrid, B.; Gadisseux, E.; Guiot, Y.; Yuan, W.; Torre, A.; Camboni, A.; van Langendonckt, A.; Donnez, J. Short-term transplantation of isolated human ovarian follicles and cortical tissue into nude mice. Reproduction 2007, 134, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Amorim, C.; Van Langendonckt, A.; David, A.; Dolmans, M.; Donnez, J. Survival of human pre-antral follicles after cryopreservation of ovarian tissue, follicular isolation and in vitro culture in a calcium alginate matrix. Hum. Reprod. 2009, 24, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Silber, S. Fresh Ovarian Tissue and Whole Ovary Transplantation. Semin. Reprod. Med. 2009, 27, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Bromer, J.; Patrizio, P. Fertility Preservation: The Rationale for Cryopreservation of the Whole Ovary. Semin. Reprod. Med. 2009, 27, 465–471. [Google Scholar] [CrossRef]
- Gavish, Z.; Peer, G.; Hadassa, R.; Yoram, C.; Meirow, D. Follicle activation and burn-out contribute to post-transplantation follicle loss in ovarian tissue grafts: The effect of graft thickness. Hum. Reprod. 2014, 29, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Gavish, Z.; Spector, I.; Peer, G.; Schlatt, S.; Wistuba, J.; Roness, H.; Meirow, D. Follicle activation is a significant and immediate cause of follicle loss after ovarian tissue transplantation. J. Assist. Reprod. Genet. 2018, 35, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, R.; Pasquinelli, G.; Keane, D.; Magnani, V.; Paradisi, R.; Venturoli, S. Optimization of protocols for human ovarian tissue cryopreservation with sucrose, 1,2-propanediol and human serum. Reprod. Biomed. Online 2010, 21, 819–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Costa, P.; Correia, S.; Gouveia-Oliveira, A.; Negreiro, F.; Jorge, S.; Cidadao, A.; Carvalho, M.; Plancha, C. Effects of mouse ovarian tissue cryopreservation on granulosa cell-oocyte interaction. Hum. Reprod. 2005, 20, 1607–1614. [Google Scholar] [CrossRef] [Green Version]
- Roness, H.; Meirow, D. Follicle reserve loss in ovarian tissue transplantation. Reproduction 2019, 158, F35–F44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Chen, H.; Yin, H.; Kim, S.; Tan, S.; Gosden, R. Fertility after intact ovary transplantation. Nature 2002, 415, 385. [Google Scholar] [CrossRef] [PubMed]
- Bedaiwy, M.; Jeremias, E.; Gurunluoglu, R.; Hussein, M.; Siemianow, M.; Biscotti, C.; Falcone, T. Restoration of ovarian function after autotransplantation of intact frozen-thawed sheep ovaries with microvascular anastomosis. Fertil. Steril. 2003, 79, 594–602. [Google Scholar] [CrossRef]
- Martinez-Madrid, B.; Dolmans, M.; van Langendonckt, A.; Defrere, S.; Donnez, J. Freeze-thawing intact human ovary with its vascular pedicle with device a passive cooling. Fertil. Steril. 2004, 82, 1390–1394. [Google Scholar] [CrossRef]
- Salehnia, M.; Moghadam, E.; Velojerdi, M. Ultrastructure of follicles after vitrification of mouse ovarian tissue. Fertil. Steril. 2002, 78, 644–645. [Google Scholar] [CrossRef]
- Migishima, F.; Suzuki-Migishima, R.; Song, S.; Kuramochi, T.; Azuma, S.; Nishijima, M.; Yokoyama, M. Successful cryopreservation of mouse ovaries by vitrification. Biol. Reprod. 2003, 68, 881–887. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, R.; Vicenti, R.; Macciocca, M.; Pasquinelli, G.; Paradisi, R.; Battaglia, C.; Martino, N.; Venturoli, S. Good Preservation of Stromal Cells and No Apoptosis in Human Ovarian Tissue after Vitrification. Biomed. Res. Int. 2014. [Google Scholar] [CrossRef]
- Diaz-Garcia, C.; Domingo, J.; Garcia-Velasco, J.; Herraiz, S.; Mirabet, V.; Iniesta, I.; Cobo, A.; Remohi, J.; Pellicer, A. Oocyte vitrification versus ovarian cortex transplantation in fertility preservation for adult women undergoing gonadotoxic treatments: A prospective cohort study. Fertil. Steril. 2018, 109, 478–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talevi, R.; Barbato, V.; Fiorentino, I.; Braun, S.; De Stefano, C.; Ferraro, R.; Sudhakaran, S.; Gualtieri, R. Successful slush nitrogen vitrification of human ovarian tissue. Fertil. Steril. 2016, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbato, V.; Gualtieri, R.; Capriglione, T.; Pallotta, M.; Braun, S.; Di Nardo, M.; Costanzo, V.; Ferraro, R.; Catapano, G.; Talevi, R. Slush nitrogen vitrification of human ovarian tissue does not alter gene expression and improves follicle health and progression in long-term in vitro culture. Fertil. Steril. 2018, 110, 1356–1366. [Google Scholar] [CrossRef]
- Hourvitz, A.; Yerushalmi, G.; Maman, E.; Raanani, H.; Elizur, S.; Brengauz, M.; Orvieto, R.; Dor, J.; Meirow, D. Combination of ovarian tissue harvesting and immature oocyte collection for fertility preservation increases preservation yield. Reprod. Biomed. Online 2015, 31, 497–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Jiang, H.; Kristensen, S.; Andersen, C. Vitrification of in vitro matured oocytes collected from surplus ovarian medulla tissue resulting from fertility preservation of ovarian cortex tissue. J. Assist. Reprod. Genet. 2016, 33, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Kedem, A.; Yerushalmi, G.; Brengauz, M.; Raanani, H.; Orvieto, R.; Hourvitz, A.; Meirow, D. Outcome of immature oocytes collection of 119 cancer patients during ovarian tissue harvesting for fertility preservation. J. Assist. Reprod. Genet. 2018, 35, 851–856. [Google Scholar] [CrossRef]
- Dolmans, M.; Marinescu, C.; Saussoy, P.; Van Langendonckt, A.; Amorim, C.; Donnez, J. Reimplantation of cryopreserved ovarian tissue from patients with acute lymphoblastic leukemia is potentially unsafe. Blood 2010, 116, 2908–2914. [Google Scholar] [CrossRef]
- Meirow, D.; Roness, H.; Kristensen, S.; Andersen, C. Optimizing outcomes from ovarian tissue cryopreservation and transplantation; activation versus preservation. Hum. Reprod. 2015, 30, 2453–2456. [Google Scholar] [CrossRef]
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, Y.; Takae, S.; Ho, C.; Kawamura, N.; Tamura, M.; Hashimoto, S.; et al. Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 17474–17479. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Kawamura, K.; Cheng, Y.; Liu, S.; Klein, C.; Duan, E.; Hsueh, A. Activation of dormant ovarian follicles to generate mature eggs. Proc. Natl. Acad. Sci. USA 2010, 107, 10280–10284. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Yoshioka, N.; Takae, S.; Sugishita, Y.; Tamura, M.; Hashimoto, S.; Morimoto, Y.; Kawamura, K. Successful fertility preservation following ovarian tissue vitrification in patients with primary ovarian insufficiency. Hum. Reprod. 2015, 30, 608–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoop, D.; Cobo, A.; Silber, S. Fertility preservation for age-related fertility decline. Lancet 2014, 384, 1311–1319. [Google Scholar] [CrossRef]
- Donnez, J.; Jadoul, P.; Squifflet, J.; van Langendonckt, A.; Donnez, O.; Van Eyck, A.; Marinescu, C.; Dolmans, M. Ovarian tissue cryopreservation and transplantation in cancer patients. Best Pr. Res. Clin. Obs. Gynaecol. 2010, 24, 87–100. [Google Scholar] [CrossRef]
- Demeestere, I.; Simon, P.; Emiliani, S.; Delbaere, A.; Englert, Y. Orthotopic and heterotopic ovarian tissue transplantation. Hum. Reprod. Update 2009, 15, 649–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leporrier, M.; von Theobald, P.; Roffe, J.; Muller, G. A new technique to protect ovarian-function before pelvic irradiation - heterotopic ovarian autotransplantation. Cancer 1987, 60, 2201–2204. [Google Scholar] [CrossRef]
- Marconi, G.; Quintana, R.; RuedaLeverone, N.; Vighi, S. Accidental ovarian autograft after a laparoscopic surgery: Case report. Fertil. Steril. 1997, 68, 364–366. [Google Scholar] [CrossRef]
- Demeestere, I.; Simon, P.; Emiliani, S.; Delbaere, A.; Englert, Y. Fertility preservation: Successful transplantation of cryopreserved ovarian tissue in a young patient previously treated for Hodgkin’s disease. Oncologist 2007, 12, 1437–1442. [Google Scholar] [CrossRef] [Green Version]
- Imbert, R.; Moffa, F.; Tsepelidis, S.; Simon, P.; Delbaere, A.; Devreker, F.; Dechene, J.; Ferster, A.; Veys, I.; Fastrez, M.; et al. Safety and usefulness of cryopreservation of ovarian tissue to preserve fertility: A 12-year retrospective analysis. Hum. Reprod. 2014, 29, 1931–1940. [Google Scholar] [CrossRef] [Green Version]
- Stern, C.; Gook, D.; Hale, L.; Agresta, F.; Oldham, J.; Rozen, G.; Jobling, T. First reported clinical pregnancy following heterotopic grafting of cryopreserved ovarian tissue in a woman after a bilateral oophorectomy. Hum. Reprod. 2013, 28, 2996–2999. [Google Scholar] [CrossRef] [Green Version]
- Stern, C.; Toledo, M.; Hale, L.; Gook, D.; Edgar, D. The first Australian experience of heterotopic grafting of cryopreserved ovarian tissue: Evidence of establishment of normal ovarian function. Aust. N. Z. J. Obs. Gynaecol. 2011, 51. [Google Scholar] [CrossRef]
- Tammiste, T.; Kask, K.; Padrik, P.; Idla, K.; Rosenstein, K.; Jatsenko, T.; Veerus, P.; Salumets, A. A case report and follow-up of the first live birth after heterotopic transplantation of cryopreserved ovarian tissue in Eastern Europe. Bmc Womens Health 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Xie, Y.; Wang, Y.; Li, S. Vitrification versus slow freezing for human ovarian tissue cryopreservation: A systematic review and meta-anlaysis. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Dolmans, M.; Diaz, C.; Pellicer, A. Ovarian cortex transplantation: Time to move on from experimental studies to open clinical application. Fertil. Steril. 2015, 104, 1097–1098. [Google Scholar] [CrossRef] [PubMed]
- Van der Ven, H.; Liebenthron, J.; Beckmann, M.; Toth, B.; Korell, M.; Krussel, J.; Frambach, T.; Kupka, M.; Hohl, M.; Winkler-Crepaz, K.; et al. Ninety-five orthotopic transplantations in 74 women of ovarian tissue after cytotoxic treatment in a fertility preservation network: Tissue activity, pregnancy and delivery rates. Hum. Reprod. 2016, 31, 2031–2041. [Google Scholar] [CrossRef]
- Meirow, D.; Ra’anani, H.; Shapira, M.; Brenghausen, M.; Chaim, S.; Aviel-Ronen, S.; Amariglio, N.; Schiff, E.; Orvieto, R.; Dor, J. Transplantations of frozen-thawed ovarian tissue demonstrate high reproductive performance and the need to revise restrictive criteria. Fertil. Steril. 2016, 106, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Demeestere, I.; Simon, P.; Dedeken, L.; Moffa, F.; Tsepelidis, S.; Brachet, C.; Delbaere, A.; Devreker, F.; Ferster, A. Live birth after autograft of ovarian tissue cryopreserved during childhood. Hum. Reprod. 2015, 30, 2107–2109. [Google Scholar] [CrossRef] [Green Version]
- Matthews, S.J.; Picton, H.; Ernst, E.; Andersen, C.Y. Successful pregnancy in a woman previously suffering from β-thalassemia following transplantation of ovarian tissue cryopreserved before puberty. Minerva Ginecol. 2018, 70, 432–435. [Google Scholar] [CrossRef]
- Von Wolff, M.; Sanger, N.; Liebenthron, J. Is Ovarian Tissue Cryopreservation and Transplantation Still Experimental? It Is a Matter of Female Age and Type of Cancer. J. Clin. Oncol. 2018, 36. [Google Scholar] [CrossRef] [Green Version]
- Dunlop, C.; Brady, B.; McLaughlin, M.; Telfer, E.; White, J.; Cowie, F.; Zahra, S.; Wallace, W.; Anderson, R. Re-implantation of cryopreserved ovarian cortex resulting in restoration of ovarian function, natural conception and successful pregnancy after haematopoietic stem cell transplantation for Wilms tumour. J. Assist. Reprod. Genet. 2016, 33, 1615–1620. [Google Scholar] [CrossRef] [Green Version]
- Oktay, K. Ovarian tissue cryopreservation and transplantation: Preliminary findings and implications for cancer patients. Hum. Reprod. Update 2001, 7, 526–534. [Google Scholar] [CrossRef] [Green Version]
- Dolmans, M.; Luyckx, V.; Donnez, J.; Andersen, C.; Greve, T. Risk of transferring malignant cells with transplanted frozen-thawed ovarian tissue. Fertil. Steril. 2013, 99, 1514–1522. [Google Scholar] [CrossRef] [PubMed]
- Greve, T.; Clasen-Linde, E.; Andersen, M.; Andersen, M.; Sorensen, S.; Rosendahl, M.; Ralfkiaer, E.; Andersen, C. Cryopreserved ovarian cortex from patients with leukemia in complete remission contains no apparent viable malignant cells. Blood 2012, 120, 4311–4316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapira, M.; Raanani, H.; Barshack, I.; Amariglio, N.; Derech-Haim, S.; Marciano, M.; Schiff, E.; Orvieto, R.; Meirow, D. First delivery in a leukemia survivor after transplantation of cryopreserved ovarian tissue, evaluated for leukemia cells contamination. Fertil. Steril. 2018, 109, 48–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, C.; Timmer-Bosscha, H.; Wijchman, J.; de Leij, L.; Hollema, H.; Heineman, M.; de Vries, E. An in vitro model for purging of tumour cells from ovarian tissue. Hum. Reprod. 2004, 19, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Soares, M.; Saussoy, P.; Maskens, M.; Reul, H.; Amorim, C.; Donnez, J.; Dolmans, M. Eliminating malignant cells from cryopreserved ovarian tissue is possible in leukaemia patients. Br. J. Haematol. 2017, 178, 231–239. [Google Scholar] [CrossRef]
- Chiti, M.; Dolmans, M.; Mortiaux, L.; Zhuge, F.; Ouni, E.; Shahri, P.; Van Ruymbeke, E.; Champagne, S.; Donnez, J.; Amorim, C. A novel fibrin-based artificial ovary prototype resembling human ovarian tissue in terms of architecture and rigidity. J. Assist. Reprod. Genet. 2018, 35, 41–48. [Google Scholar] [CrossRef]
- Telfer, E.; Fauser, B. Important steps towards materializing the dream of developing an artificial ovary. Reprod. Biomed. Online 2016, 33, 333–334. [Google Scholar] [CrossRef]
- McLaughlin, M.; Albertini, D.; Wallace, W.; Anderson, R.; Telfer, E. Metaphase II oocytes from human unilaminar follicles grown in a multi-step culture system. Mol. Hum. Reprod. 2018, 24, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Jensen, A.; Kristensen, S.; Macklon, K.; Jeppesen, J.; Fedder, J.; Ernst, E.; Andersen, C. Outcomes of transplantations of cryopreserved ovarian tissue to 41 women in Denmark. Hum. Reprod. 2015, 30, 2838–2845. [Google Scholar] [CrossRef] [Green Version]
- Oktay, K.; Bedoschi, G.; Pacheco, F.; Turan, V.; Emirdar, V. First pregnancies, live birth, and in vitro fertilization outcomes after transplantation of frozen-banked ovarian tissue with a human extracellular matrix scaffold using robot-assisted minimally invasive surgery. Am. J. Obs. Gynecol. 2016, 214. [Google Scholar] [CrossRef] [Green Version]
- Zhai, J.; Yao, G.; Dong, F.; Bu, Z.; Cheng, Y.; Sato, Y.; Hu, L.; Zhang, Y.; Wang, J.; Dai, S.; et al. In Vitro Activation of Follicles and Fresh Tissue Auto-transplantation in Primary Ovarian Insufficiency Patients. J. Clin. Endocrinol. Metab. 2016, 101, 4405–4412. [Google Scholar] [CrossRef] [PubMed]
- Fabregues, F.; Ferreri, J.; Calafell, J.; Moreno, V.; Borras, A.; Manau, D.; Carmona, F. Pregnancy after drug-free in vitro activation of follicles and fresh tissue autotransplantation in primary ovarian insufficiency patient: A case report and literature review. J. Ovarian Res. 2018, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, T.; Johnson, J.; White, Y.; St John, J. The molecular characterization of porcine egg precursor cells. Oncotarget 2017, 8, 63484–63505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, D.C.; Tilly, J.L. Woods and Tilly reply. Nat. Med. 2015, 21, 1118–1121. [Google Scholar] [CrossRef] [Green Version]
- Guo, K.; Li, C.; Wang, X.; He, D.; Zheng, P. Germ stem cells are active in postnatal mouse ovary under physiological conditions. Mol. Hum. Reprod. 2016, 22, 316–328. [Google Scholar] [CrossRef] [Green Version]
- Park, E.; Tilly, J. Use of DEAD-box polypeptide-4 (Ddx4) gene promoter-driven fluorescent reporter mice to identify mitotically active germ cells in post-natal mouse ovaries. Mol. Hum. Reprod. 2015, 21, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Parte, S.; Bhartiya, D.; Patel, H.; Daithankar, V.; Chauhan, A.; Zaveri, K.; Hinduja, I. Dynamics associated with spontaneous differentiation of ovarian stem cells in vitro. J. Ovarian Res. 2014, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Silvestris, E.; Cafforio, P.; D’Oronzo, S.; Felici, C.; Silvestris, F.; Loverro, G. In vitro differentiation of human oocyte-like cells from oogonial stem cells: Single-cell isolation and molecular characterization. Hum. Reprod. 2018, 33, 464–473. [Google Scholar] [CrossRef] [Green Version]
- White, Y.; Woods, D.; Takai, Y.; Ishihara, O.; Seki, H.; Tilly, J. Oocyte formation by mitotically active germ cells purified from ovaries of reproductive-age women. Nat. Med. 2012, 18, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Telfer, E.; Albertini, D. The quest for human ovarian stem cells. Nat. Med. 2012, 18, 353–354. [Google Scholar] [CrossRef]
- Truman, A.; Tilly, J.; Woods, D. Ovarian regeneration: The potential for: Stem cell contribution in the postnatal ovary to sustained endocrine function. Mol. Cell. Endocrinol. 2017, 445, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anchan, R.; Gerami-Naini, B.; Lindsey, J.; Ho, J.; Kiezun, A.; Lipskind, S.; Ng, N.; LiCausi, J.; Kim, C.; Brezina, P.; et al. Efficient Differentiation of Steroidogenic and Germ-Like Cells from Epigenetically-Related iPSCs Derived from Ovarian Granulosa Cells. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yager, J.; Davidson, N. Mechanisms of disease: Estrogen carcinogenesis in breast cancer. N. Engl. J. Med. 2006, 354, 270–282. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FP Procedures | Age | Pros | Cons | References |
|---|---|---|---|---|
| Oocyte cryopreservation | -Postpuberal women | -Well established technique -High percentage of success | -Need for COS and cycle dependence -Need to delay oncological treatment -Oncogenic risk for hormonal-cancers -Not applicable in females with poor OR | [7]. Loren AW et al, 2013. [9] Parmegiani L et al, 2009. [10]. Annan JJ et al, 2013 |
| Embryo cryopreservation | -Postpuberal women | -Well established technique -Good percentage of success | -Need for COS and cycle dependence -Need to delay oncological treatment -Oncogenic risk for hormonal-cancers -Not applicable in females with poor OR -Limited to few countries | [8]. Peccatori FA et al, 2013. [10]. Annan JJ et al, 2013. [11]. Bianchii V et al, 2012 |
| Ovarian cortex cryopreservation | -Prepuberal women -Postpuberal women | -Immediate application -No need for COS or cycle dependence -No oncogenic risk for hormone sensitive cancers | -Experimental technique-Pelvic surgery -Oncogenic risk after replantation -Variable risk of unsuccess due to the oocyte depletion in implanted fragments -No possibility to select most viable oocytes -Limited to expert infertility centers | [34]. Jeruss J et al, 2009. [35]. Andersen C et al, 2019. [44]. Martinez F et al, 2015 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silvestris, E.; De Palma, G.; Canosa, S.; Palini, S.; Dellino, M.; Revelli, A.; Paradiso, A.V. Human Ovarian Cortex biobanking: A Fascinating Resource for Fertility Preservation in Cancer. Int. J. Mol. Sci. 2020, 21, 3245. https://doi.org/10.3390/ijms21093245
Silvestris E, De Palma G, Canosa S, Palini S, Dellino M, Revelli A, Paradiso AV. Human Ovarian Cortex biobanking: A Fascinating Resource for Fertility Preservation in Cancer. International Journal of Molecular Sciences. 2020; 21(9):3245. https://doi.org/10.3390/ijms21093245
Chicago/Turabian StyleSilvestris, Erica, Giuseppe De Palma, Stefano Canosa, Simone Palini, Miriam Dellino, Alberto Revelli, and Angelo Virgilio Paradiso. 2020. "Human Ovarian Cortex biobanking: A Fascinating Resource for Fertility Preservation in Cancer" International Journal of Molecular Sciences 21, no. 9: 3245. https://doi.org/10.3390/ijms21093245
APA StyleSilvestris, E., De Palma, G., Canosa, S., Palini, S., Dellino, M., Revelli, A., & Paradiso, A. V. (2020). Human Ovarian Cortex biobanking: A Fascinating Resource for Fertility Preservation in Cancer. International Journal of Molecular Sciences, 21(9), 3245. https://doi.org/10.3390/ijms21093245