Tumor-Targeting Peptides Search Strategy for the Delivery of Therapeutic and Diagnostic Molecules to Tumor Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Biopanning of Linear Phage Libraries Ph.D.-12 and Ph.D.-7 on Cells and Tumors U-87 MG
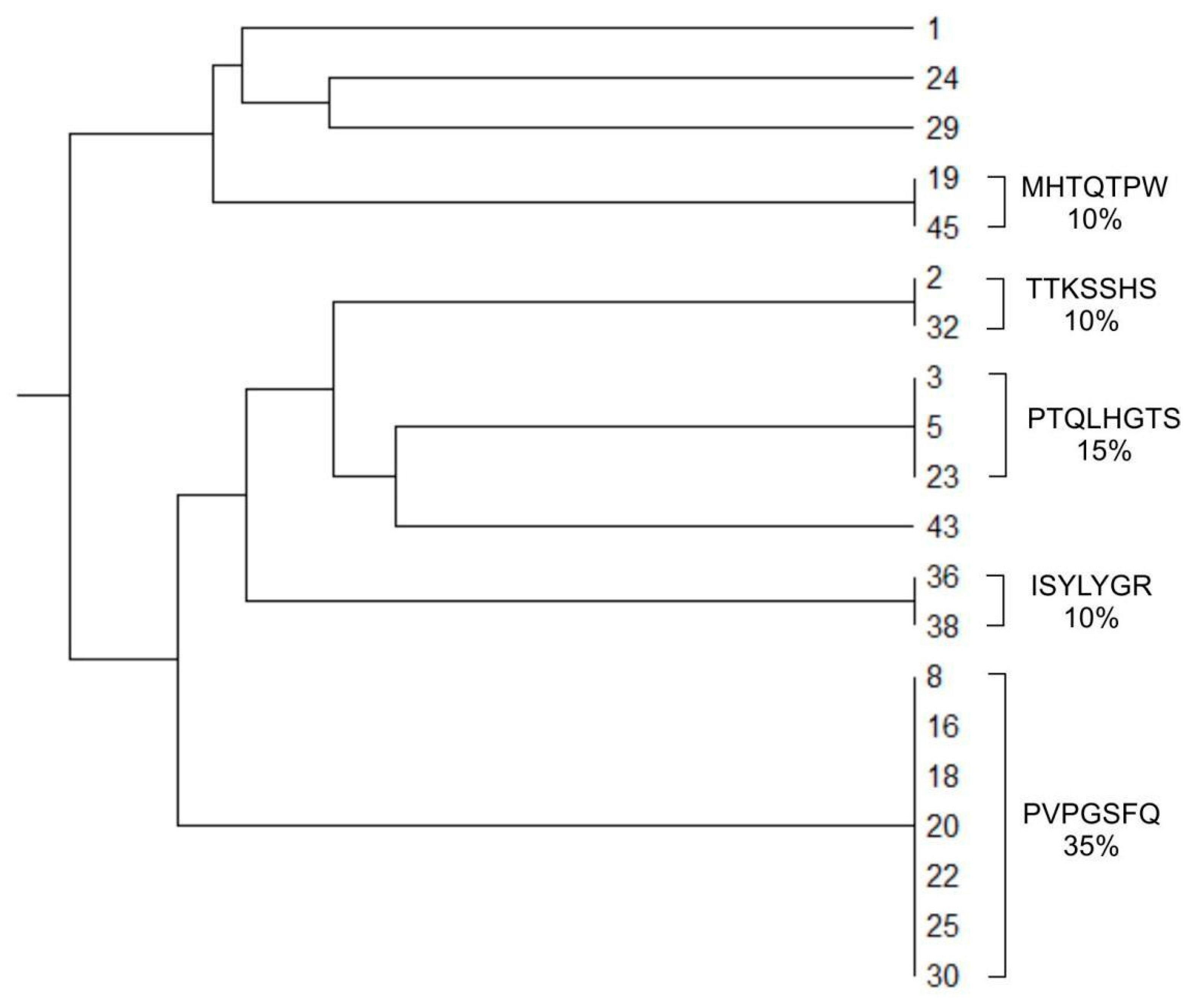
2.2. Biopanning of the Circular Phage Peptide Library Ph.D.-C7C In Vivo and In Vitro
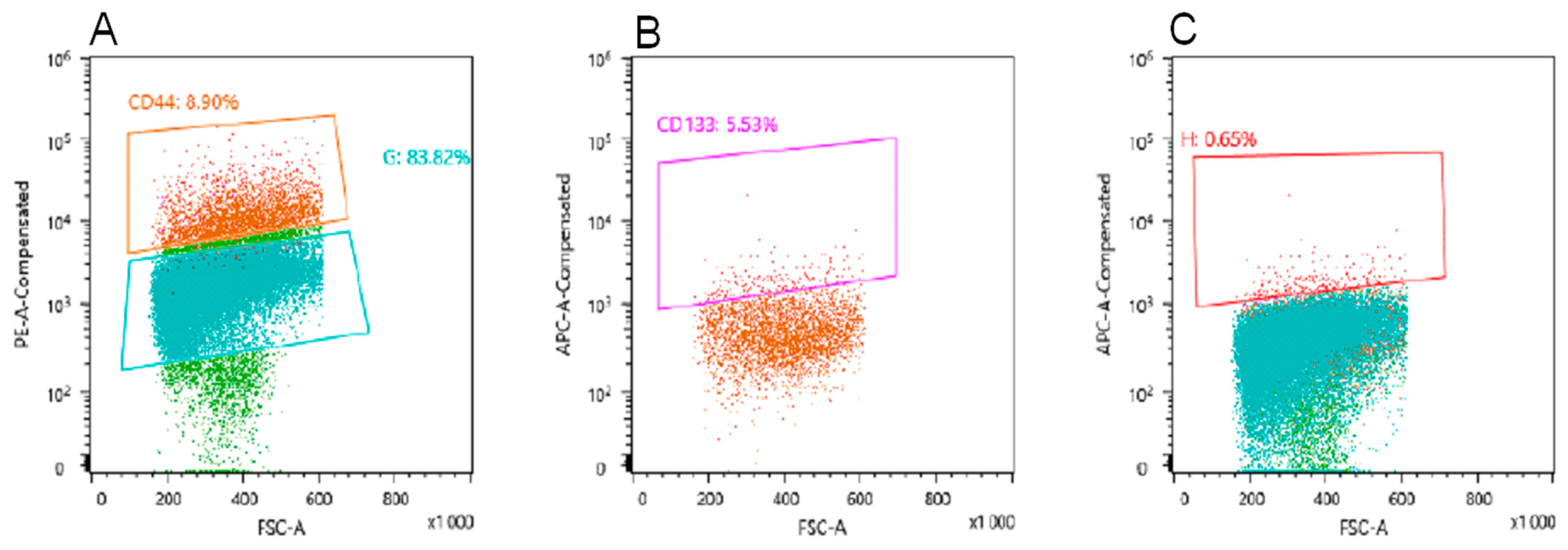
2.3. Obtaining a Population of CD44+/CD133+ U-87 MG Cells for Selection of Bacteriophages Displaying Peptides Specific to CSCs
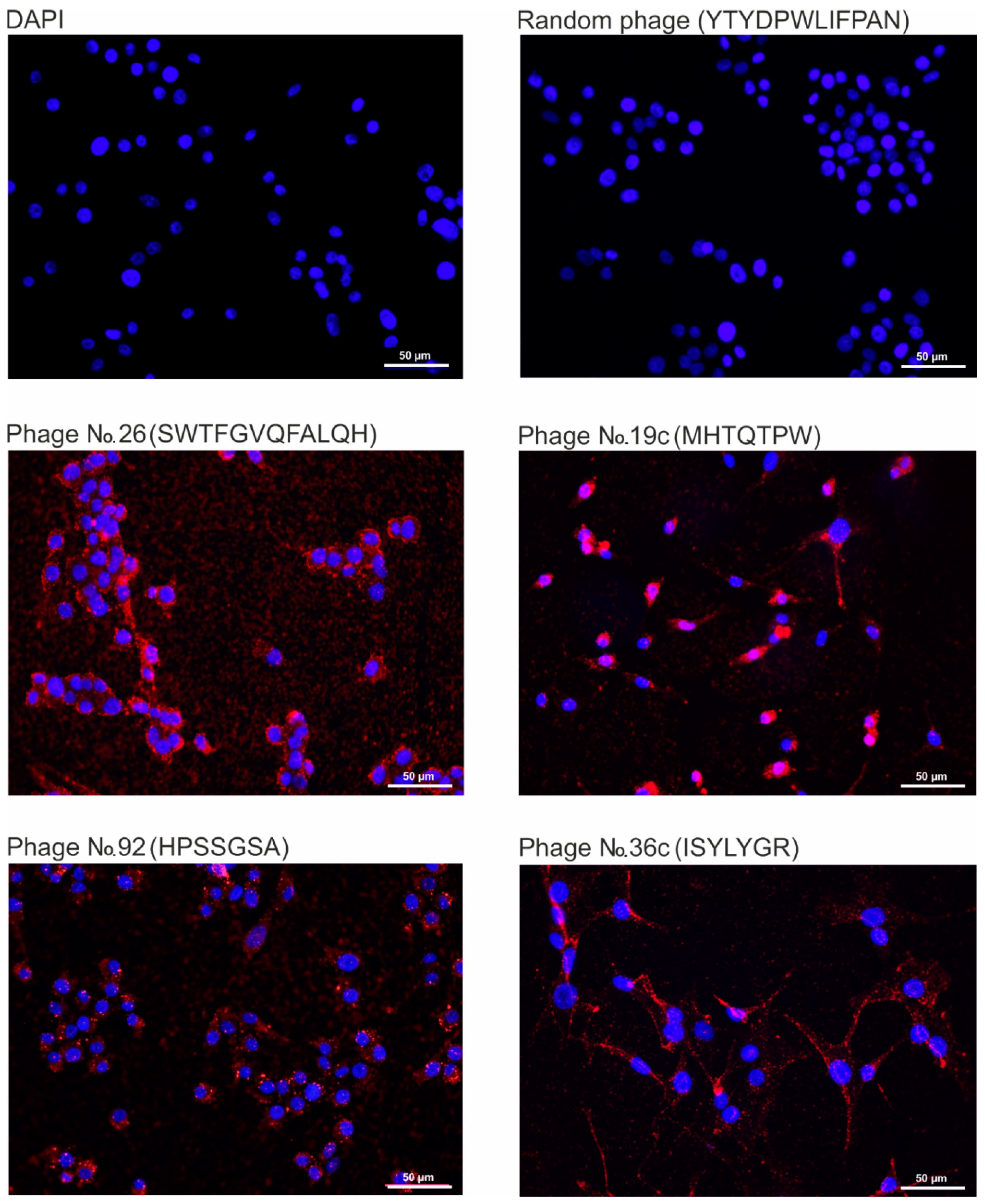
2.4. Analysis of the Binding Specificity of Bacteriophages, Displaying Selected Peptides, to Human Glioblastoma Cells U-87 MG
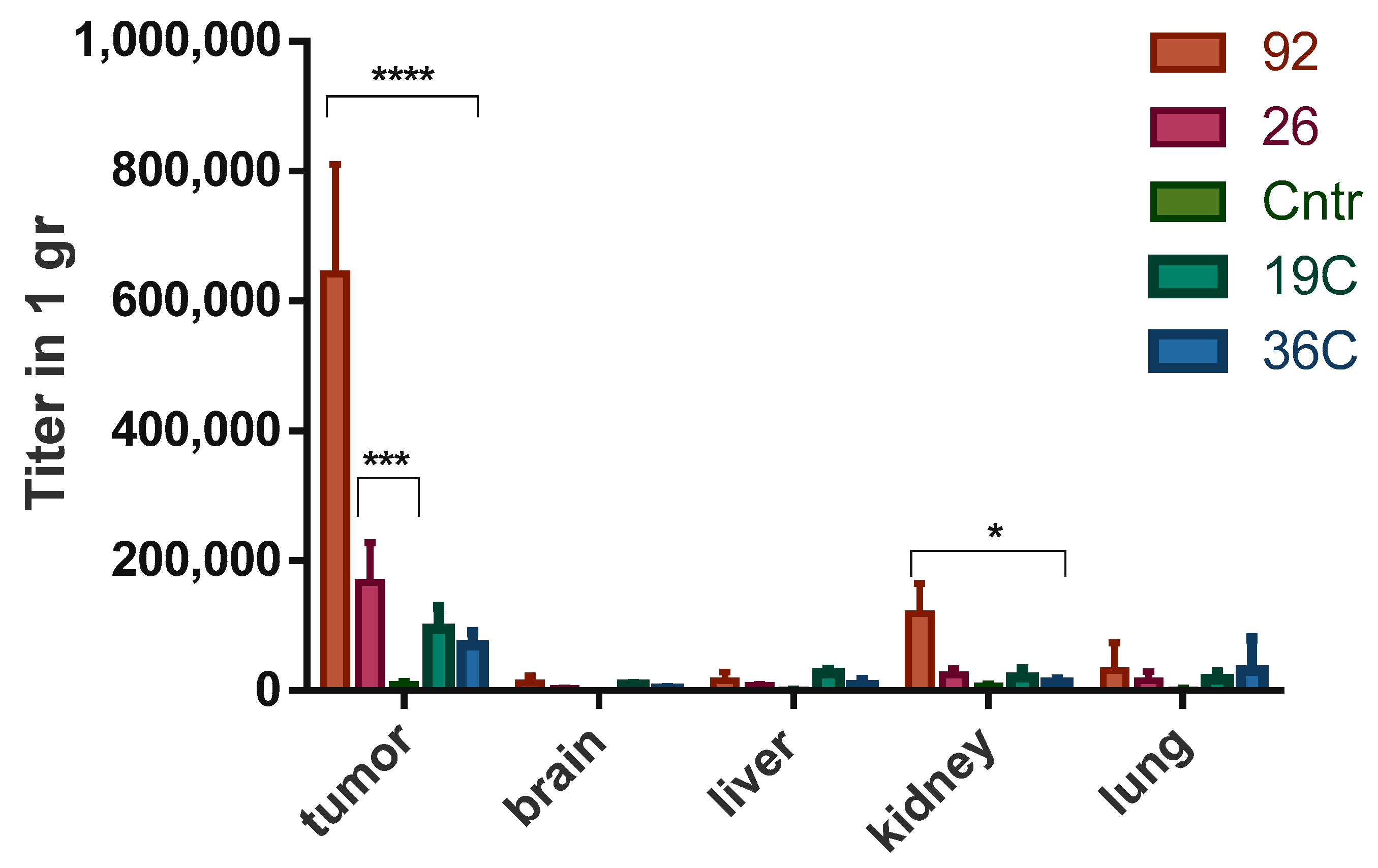
2.5. Analysis of Biodistribution and Specificity of Accumulation of Bacteriophages, Displaying Selected Tumor-Targeting Peptides, in U87 MG Tumor Tissue
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Animals
4.3. In Vivo and In Vitro Biopanning
4.4. Tumor Preparation for Cell Sorting
4.5. Cell Sorting
4.6. Immunocytochemistry
4.7. Analysis of the Specificity of Accumulation of Bacteriophages Displayed Selected Peptides in Glioblastoma Xenograft U-87 Mg
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taylor, O.G.; Brzozowski, J.S.; Skelding, K.A. Glioblastoma multiforme: An overview of emerging therapeutic targets. Front. Oncol. 2019, 9, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badr, C.E.; Silver, D.J.; Siebzehnrubl, F.A.; Deleyrolle, L.P. Metabolic heterogeneity and adaptability in brain tumors. Cell. Mol. Life Sci. 2020, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Saw, P.E.; Song, E.-W. Phage display screening of therapeutic peptide for cancer targeting and therapy. Protein Cell 2019, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voitova, A.A.; Dmitrieva, M.D.; Dymova, M.A.; Vasileva, N.S.; Nushtaeva, A.A.; Richter, V.A.; Kuligina, E.V. Tumor Specific Peptides Selected for Targeted Delivery of Therapeutic Agents to Glioma Human Cells. Russ. J. Bioorg. Chem. 2019, 45, 783–792. [Google Scholar] [CrossRef]
- Nemudraya, A.A.; Makartsova, A.A.; Fomin, A.S.; Nushtaeva, A.A.; Koval, O.A.; Richter, V.A.; Kuligina, E.V. Tumor-Specific Peptide, Selected from a Phage Peptide Library, Enhances Antitumor Activity of Lactaptin. PLoS ONE 2016, 11, e0160980. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zuo, X.; Wei, D. Concise Review: Emerging Role of CD44 in Cancer Stem Cells: A Promising Biomarker and Therapeutic Target. Stem Cells Transl. Med. 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.; Motiani, K.; Giridhar, P.V.; Kasper, S. CD44 integrates signaling in normal stem cell, cancer stem cell and (pre)metastatic niches. Exp. Biol. Med. 2013, 238, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Lingala, S.; Cui, Y.Y.; Chen, X.; Ruebner, B.H.; Qian, X.F.; Zern, M.A.; Wu, J. Immunohistochemical staining of cancer stem cell markers in hepatocellular carcinoma. Exp. Mol. Pathol. 2010, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutella, S.; Bonanno, G.; Procoli, A.; Mariotti, A.; Corallo, M.; Prisco, M.G.; Eramo, A.; Napoletano, C.; Gallo, D.; Perillo, A.; et al. Cells with characteristics of cancer stem/progenitor cells express the CD133 antigen in human endometrial tumors. Clin. Cancer Res. 2009, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haraguchi, N.; Ohkuma, M.; Sakashita, H.; Matsuzaki, S.; Tanaka, F.; Mimori, K.; Kamohara, Y.; Inoue, H.; Mori, M. CD133+CD44+ population efficiently enriches colon cancer initiating cells. Ann. Surg. Oncol. 2008, 15. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.V.; Filiz, G.; Daniel, P.M.; Hollande, F.; Dworkin, S.; Amiridis, S.; Kountouri, N.; Ng, W.; Morokoff, A.P.; Mantamadiotis, T. Expression of CD133 and CD44 in glioblastoma stem cells correlates with cell proliferation, phenotype stability and intratumor heterogeneity. PLoS ONE 2017, 12, e172791. [Google Scholar] [CrossRef] [PubMed]
- Griguer, C.E.; Oliva, C.R.; Gobin, E.; Marcorelles, P.; Benos, D.J.; Lancaster, J.R.; Gillespie, G.Y. CD133 is a marker of bioenergetic stress in human glioma. PLoS ONE 2008, 3, e3655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, M.; Hoque, M.O. Targeting cancer stem cells: A strategy for effective eradication of cancer. Cancers 2019, 11, 732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sioud, M. Phage Display Libraries: From Binders to Targeted Drug Delivery and Human Therapeutics. Mol. Biotechnol. 2019, 61, 286–303. [Google Scholar] [CrossRef] [PubMed]
- Roxin, Á.; Zheng, G. Flexible or fixed: A comparative review of linear and cyclic cancer-targeting peptides. Future Med. Chem. 2012, 4, 1601–1618. [Google Scholar] [CrossRef] [PubMed]
- Lau, J.L.; Dunn, M.K. Therapeutic peptides: Historical perspectives, current development trends, and future directions. Bioorg. Med. Chem. 2018, 26, 2700–2707. [Google Scholar] [CrossRef] [PubMed]
- Marsh, W.; Witten, A.; Stabenfeldt, S.E. Exploiting Phage Display for Development of Novel Cellular Targeting Strategies; Humana Press: New York, NY, USA, 2018; pp. 71–94. [Google Scholar]
- Soendergaard, M.; Newton-Northup, J.R.; Deutscher, S.L. In vivo phage display selection of an ovarian cancer targeting peptide for SPECT/CT imaging. Am. J. Nucl. Med. Mol. Imaging 2014, 4, 561–570. [Google Scholar] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dmitrieva, M.D.; Voitova, A.A.; Dymova, M.A.; Richter, V.A.; Kuligina, E.V. Tumor-Targeting Peptides Search Strategy for the Delivery of Therapeutic and Diagnostic Molecules to Tumor Cells. Int. J. Mol. Sci. 2021, 22, 314. https://doi.org/10.3390/ijms22010314
Dmitrieva MD, Voitova AA, Dymova MA, Richter VA, Kuligina EV. Tumor-Targeting Peptides Search Strategy for the Delivery of Therapeutic and Diagnostic Molecules to Tumor Cells. International Journal of Molecular Sciences. 2021; 22(1):314. https://doi.org/10.3390/ijms22010314
Chicago/Turabian StyleDmitrieva, Maria D., Anna A. Voitova, Maya A. Dymova, Vladimir A. Richter, and Elena V. Kuligina. 2021. "Tumor-Targeting Peptides Search Strategy for the Delivery of Therapeutic and Diagnostic Molecules to Tumor Cells" International Journal of Molecular Sciences 22, no. 1: 314. https://doi.org/10.3390/ijms22010314
APA StyleDmitrieva, M. D., Voitova, A. A., Dymova, M. A., Richter, V. A., & Kuligina, E. V. (2021). Tumor-Targeting Peptides Search Strategy for the Delivery of Therapeutic and Diagnostic Molecules to Tumor Cells. International Journal of Molecular Sciences, 22(1), 314. https://doi.org/10.3390/ijms22010314