Presynaptic Inhibition of Pain and Touch in the Spinal Cord: From Receptors to Circuits

{kind=link}
{kind=link}
{kind=link}
Abstract
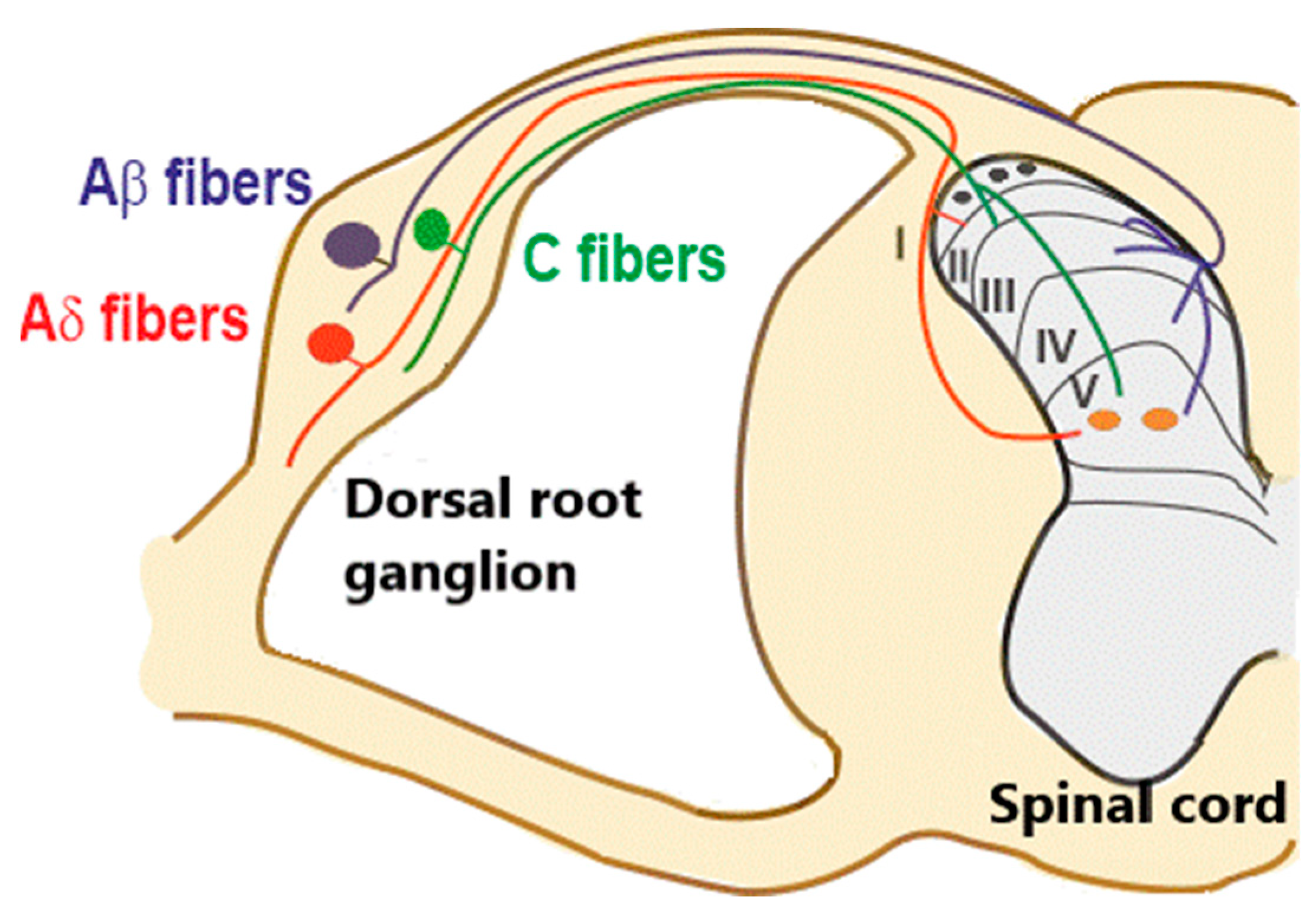
:1. Introduction
2. GABAergic Inhibition on Primary Afferent Fibers
2.1. Primary Afferent Depolarization
2.2. GABAB Receptors as Presynaptic Modulators
3. Dorsal Horn Circuits Involved in GABAergic Presynaptic Inhibition
3.1. Presynaptic Inhibition of Tactile Stimuli
3.2. Presynaptic Modulation of Nociceptive Fibers
4. GABA Release from Descending Fibers and Astrocytes
5. Modifications of Presynaptic Inhibition in Chronic Pain
6. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Koch, S.C.; Acton, D.; Goulding, M. Spinal Circuits for Touch, Pain, and Itch. Annu. Rev. Physiol. 2018, 80, 189–217. [Google Scholar] [CrossRef]
- Merighi, A. The Histology, Physiology, Neurochemistry and Circuitry of the Substantia Gelatinosa Rolandi (Lamina II) in Mammalian Spinal Cord. Prog. Neurobiol. 2018, 169, 91–134. [Google Scholar] [CrossRef] [PubMed]
- Gatto, G.; Smith, K.M.; Ross, S.E.; Goulding, M. Neuronal Diversity in the Somatosensory System: Bridging the Gap between Cell Type and Function. Curr. Opin. Neurobiol. 2019, 56, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.I.; Todd, A.J. Central Nervous System Targets: Inhibitory Interneurons in the Spinal Cord. Neurotherapeutics 2020, 17, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Komisaruk, R. Nociceptive responses to altered GABAergic activity at the spinal cord. Life Sci. 1986, 39, 1667–1674. [Google Scholar] [CrossRef]
- Melzack, R.; Wall, P.D. Pain mechanisms: A new theory. Science 1965, 150, 971–979. [Google Scholar] [CrossRef]
- Paul, J.; Zeilhofer, H.U.; Fritschy, J.M. Selective Distribution of GABAA Receptor Subtypes in Mouse Spinal Dorsal Horn Neurons and Primary Afferents. J. Comp. Neurol. 2012, 520, 3895–3911. [Google Scholar] [CrossRef]
- Lorenzo, L.E.; Godin, A.G.; Wang, F.; St-Louis, M.; Carbonetto, S.; Wiseman, P.W.; Ribeiro-da-Silva, A.; De Koninck, Y. Gephyrin Clusters Are Absent from Small Diameter Primary Afferent Terminals despite the Presence of GABAA Receptors. J. Neurosci. 2014, 34, 8300–8317. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Saunders, P.A.; Somogyi, R.; Poulter, M.O.; Barker, J.L. Ontogeny of GABAA Receptor Subunit MRNAs in Rat Spinal Cord and Dorsal Root Ganglia. J. Comp. Neurol. 1993, 338, 337–359. [Google Scholar] [CrossRef]
- Chen, J.T.C.; Guo, D.; Campanelli, D.; Frattini, F.; Mayer, F.; Zhou, L.; Kuner, R.; Heppenstall, P.A.; Knipper, M.; Hu, J. Presynaptic GABAergic Inhibition Regulated by BDNF Contributes to Neuropathic Pain Induction. Nat. Commun. 2014, 5, 5331. [Google Scholar] [CrossRef] [Green Version]
- Frank, K.; Fuortes, M.G.F. Presynaptic and postsynaptic inhibition of monosynaptic reflexes. Fed. Proc. 1957, 16, 39–40. [Google Scholar]
- Gallagher, J.P.; Higashi, H.; Nishi, S. Characterization and Ionic Basis of GABA-Induced Depolarizations Recorded in Vitro from Cat Primary Afferent Neurones. J. Physiol. 1978, 275, 263–282. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Leefmans, F.J.; Gamiño, S.M.; Giraldez, F.; Noguerón, I. Intracellular Chloride Regulation in Amphibian Dorsal Root Ganglion Neurones Studied with Ion-Selective Microelectrodes. J. Physiol. 1988, 406, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Sung, K.W.; Kirby, M.; McDonald, M.P.; Lovinger, D.M.; Delpire, E. Abnormal GABAA Receptor-Mediated Currents in Dorsal Root Ganglion Neurons Isolated from Na-K-2Cl Cotransporter Null Mice. J. Neurosci. 2000, 20, 7531–7538. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, D.; Franjic-Würtz, C.; Funk, K.; Gensch, T.; Frings, S.; Möhrlen, F. Differential Maturation of Chloride Homeostasis in Primary Afferent Neurons of the Somatosensory System. Int. J. Dev. Neurosci. 2007, 25, 479–489. [Google Scholar] [CrossRef]
- Mao, S.; Garzon-Muvdi, T.; Di Fulvio, M.; Chen, Y.; Delpire, E.; Alvarez, F.J.; Alvarez-Leefmans, F.J. Molecular and Functional Expression of Cation-Chloride Cotransporters in Dorsal Root Ganglion Neurons during Postnatal Maturation. J. Neurophysiol. 2012, 108, 834–852. [Google Scholar] [CrossRef] [Green Version]
- Rocha-González, H.I.; Mao, S.; Alvarez-Leefmans, F.J. Na+,K+,2Cl- Cotransport and Intracellular Chloride Regulation in Rat Primary Sensory Neurons: Thermodynamic and Kinetic Aspects. J. Neurophysiol. 2008, 100, 169–184. [Google Scholar] [CrossRef] [Green Version]
- Graham, B.; Redman, S. A Simulation of Action Potentials in Synaptic Boutons during Presynaptic Inhibition. J. Neurophysiol. 1994, 71, 538–549. [Google Scholar] [CrossRef] [Green Version]
- Walmsley, B.; Graham, B.; Nicol, M.J. Serial E-M and Simulation Study of Presynaptic Inhibition along a Group Ia Collateral in the Spinal Cord. J. Neurophysiol. 1995, 74, 616–623. [Google Scholar] [CrossRef] [Green Version]
- Rudomin, P.; Schmidt, R.F. Presynaptic Inhibition in the Vertebrate Spinal Cord Revisited. Exp. Brain Res. 1999, 129, 1–37. [Google Scholar] [CrossRef]
- French, A.S.; Panek, I.; Torkkeli, P.H. Shunting versus Inactivation: Simulation of GABAergic Inhibition in Spider Mechanoreceptors Suggests That Either Is Sufficient. Neurosci. Res. 2006, 55, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Takkala, P.; Zhu, Y.; Prescott, S.A. Combined Changes in Chloride Regulation and Neuronal Excitability Enable Primary Afferent Depolarization to Elicit Spiking without Compromising Its Inhibitory Effects. PLoS Comput. Biol. 2016, 12, e1005215. [Google Scholar] [CrossRef] [PubMed]
- Segev, I. Computer Study of Presynaptic Inhibition Controlling the Spread of Action Potentials into Axonal Terminals. J. Neurophysiol. 1990, 63, 987–998. [Google Scholar] [CrossRef] [PubMed]
- Cattaert, D.; El Manira, A. Shunting versus Inactivation: Analysis of Presynaptic Inhibitory Mechanisms in Primary Afferents of the Crayfish. J. Neurosci. 1999, 19, 6079–6089. [Google Scholar] [CrossRef]
- Cattaert, D.; Libersat, F.; El Manira, A. A Presynaptic Inhibition and Antidromic Spikes in Primary Afferents of the Crayfish: A Computational and Experimental Analysis. J. Neurosci. 2001, 21, 1007–1021. [Google Scholar] [CrossRef]
- Verdier, D.; Lund, J.P.; Kolta, A. GABAergic Control of Action Potential Propagation along Axonal Branches of Mammalian Sensory Neurons. J. Neurosci. 2003, 23, 2002–2007. [Google Scholar] [CrossRef] [Green Version]
- Willis, W.D. Dorsal Root Potentials and Dorsal Root Reflexes: A Double-Edged Sword. Exp. Brain. Res. 1999, 124, 395–421. [Google Scholar] [CrossRef]
- Prescott, S.A.; Sejnowski, T.J.; De Koninck, Y. Reduction of Anion Reversal Potential Subverts the Inhibitory Control of Firing Rate in Spinal Lamina I Neurons: Towards a Biophysical Basis for Neuropathic Pain. Mol. Pain. 2006, 2, 32. [Google Scholar] [CrossRef] [Green Version]
- Eccles, J.C.; Schmidt, R.; Willis, W.D. Pharmacological studies on presynaptic inhibition. J. Physiol. 1963, 168, 500–530. [Google Scholar] [CrossRef]
- Rudomín, P.; Engberg, I.; Jiménez, I. Mechanisms Involved in Presynaptic Depolarization of Group I and Rubrospinal Fibers in Cat Spinal Cord. J. Neurophysiol. 1981, 46, 532–548. [Google Scholar] [CrossRef]
- Witschi, R.; Punnakkal, P.; Paul, J.; Walczak, J.S.; Cervero, F.; Fritschy, J.M.; Kuner, R.; Keist, R.; Rudolph, U.; Zeilhofer, H.U. Presynaptic α2-GABAA Receptors in Primary Afferent Depolarization and Spinal Pain Control. J. Neurosci. 2011, 31, 8134–8142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardoni, R. Role of Presynaptic Glutamate Receptors in Pain Transmission at the Spinal Cord Level. Curr. Neuropharmacol. 2013, 11, 477–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas-Osma, A.M.; Li, Y.; Lin, S.; Black, S.; Singla, R.; Fouad, K.; Fenrich, K.K.; Bennett, D.J. Extrasynaptic α5-GABAA Receptors on Proprioceptive Afferents Produce a Tonic Depolarization That Modulates Sodium Channel Function in the Rat Spinal Cord. J. Neurophysiol. 2018, 120, 2953–2974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudomin, P.; Jiménez, I.; Chávez, D. Differential Presynaptic Control of the Synaptic Effectiveness of Cutaneous Afferents Evidenced by Effects Produced by Acute Nerve Section. J. Physiol. 2013, 591, 2629–2645. [Google Scholar] [CrossRef] [PubMed]
- Fink, A.J.P.; Croce, K.R.; Huang, Z.J.; Abbott, L.F.; Jessell, T.M.; Azim, E. Presynaptic Inhibition of Spinal Sensory Feedback Ensures Smooth Movement. Nature 2014, 508, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Betelli, C.; MacDermott, A.B.; Bardoni, R. Transient, Activity Dependent Inhibition of Transmitter Release from Low Threshold Afferents Mediated by GABAA Receptors in Spinal Cord Lamina III/IV. Mol. Pain. 2015, 11, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, E.C.; Pechincha, C.; Luz, L.L.; Kokai, E.; Szucs, P.; Safronov, B.V. Primary Afferent-Driven Presynaptic Inhibition of C-Fiber Inputs to Spinal Lamina I Neurons. Prog. Neurobiol. 2020, 188, 101786. [Google Scholar] [CrossRef]
- Malcangio, M. GABAB Receptors and Pain. Neuropharmacology 2018, 136, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Towers, S.; Princivalle, A.; Billinton, A.; Edmunds, M.; Bettler, B.; Urban, L.; Castro-Lopes, J.; Bowery, N.G. GABAB Receptor Protein and MRNA Distribution in Rat Spinal Cord and Dorsal Root Ganglia. Eur. J. Neurosci. 2000, 12, 3201–3210. [Google Scholar] [CrossRef]
- Yang, K.; Wang, D.; Li, Y.Q. Distribution and Depression of the GABA(B) Receptor in the Spinal Dorsal Horn of Adult Rat. Brain. Res. Bull. 2001, 55, 479–485. [Google Scholar] [CrossRef]
- Salio, C.; Merighi, A.; Bardoni, R. GABAB Receptors-Mediated Tonic Inhibition of Glutamate Release from Aβ Fibers in Rat Laminae III/IV of the Spinal Cord Dorsal Horn. Mol. Pain. 2017, 13, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuhara, K.; Katafuchi, T.; Yoshimura, M. Effects of Baclofen on Mechanical Noxious and Innocuous Transmission in the Spinal Dorsal Horn of the Adult Rat: In Vivo Patch-Clamp Analysis. Eur. J. Neurosci. 2013, 38, 3398–3407. [Google Scholar] [CrossRef] [PubMed]
- Kangrga, I.; Jiang, M.C.; Randić, M. Actions of (-)-Baclofen on Rat Dorsal Horn Neurons. Brain. Res 1991, 562, 265–275. [Google Scholar] [CrossRef]
- Malcangio, M.; Bowery, N.G. Gamma-Aminobutyric AcidB, but Not Gamma-Aminobutyric AcidA Receptor Activation, Inhibits Electrically Evoked Substance P-Like Immunoreactivity Release from the Rat Spinal Cord In Vitro. J. Pharmacol. Exp. Ther. 1993, 266, 1490–1496. [Google Scholar] [PubMed]
- Ataka, T.; Kumamoto, E.; Shimoji, K.; Yoshimura, M. Baclofen Inhibits More Effectively C-Afferent than Adelta-Afferent Glutamatergic Transmission in Substantia Gelatinosa Neurons of Adult Rat Spinal Cord Slices. Pain 2000, 86, 273–282. [Google Scholar] [CrossRef]
- Papon, M.-A.; Le Feuvre, Y.; Barreda-Gómez, G.; Favereaux, A.; Farrugia, F.; Bouali-Benazzouz, R.; Nagy, F.; Rodríguez-Puertas, R.; Landry, M. Spinal Inhibition of GABAB Receptors by the Extracellular Matrix Protein Fibulin-2 in Neuropathic Rats. Front Cell Neurosci. 2020, 14, 214. [Google Scholar] [CrossRef]
- Désarmenien, M.; Feltz, P.; Occhipinti, G.; Santangelo, F.; Schlichter, R. Coexistence of GABAA and GABAB Receptors on A Delta and C Primary Afferents. Br. J. Pharmacol. 1984, 81, 327–333. [Google Scholar] [CrossRef]
- Huang, D.; Huang, S.; Peers, C.; Du, X.; Zhang, H.; Gamper, N. GABAB Receptors Inhibit Low-Voltage Activated and High-Voltage Activated Ca(2+) Channels in Sensory Neurons via Distinct Mechanisms. Biochem. Biophys. Res. Commun. 2015, 465, 188–193. [Google Scholar] [CrossRef]
- Dittman, J.S.; Regehr, W.G. Contributions of Calcium-Dependent and Calcium-Independent Mechanisms to Presynaptic Inhibition at a Cerebellar Synapse. J. Neurosci. 1996, 16, 1623–1633. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Ma, H. Blockade of GABAB Receptors Facilitates Evoked Neurotransmitter Release at Spinal Dorsal Horn Synapse. Neuroscience 2011, 193, 411–420. [Google Scholar] [CrossRef]
- Todd, A.J. GABA and Glycine in Synaptic Glomeruli of the Rat Spinal Dorsal Horn. Eur. J. Neurosci. 1996, 8, 2492–2498. [Google Scholar] [CrossRef] [PubMed]
- Szucs, P.; Luz, L.L.; Pinho, R.; Aguiar, P.; Antal, Z.; Tiong, S.Y.X.; Todd, A.J.; Safronov, B.V. Axon Diversity of Lamina I Local-Circuit Neurons in the Lumbar Spinal Cord. J. Comp. Neurol. 2013, 521, 2719–2741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, D.I.; Sikander, S.; Kinnon, C.M.; Boyle, K.A.; Watanabe, M.; Callister, R.J.; Graham, B.A. Morphological, Neurochemical and Electrophysiological Features of Parvalbumin-Expressing Cells: A Likely Source of Axo-Axonic Inputs in the Mouse Spinal Dorsal Horn. J. Physiol. 2012, 590, 3927–3951. [Google Scholar] [CrossRef] [PubMed]
- Boyle, K.A.; Gradwell, M.A.; Yasaka, T.; Dickie, A.C.; Polgár, E.; Ganley, R.P.; Orr, D.P.H.; Watanabe, M.; Abraira, V.E.; Kuehn, E.D.; et al. Defining a Spinal Microcircuit That Gates Myelinated Afferent Input: Implications for Tactile Allodynia. Cell Rep. 2019, 28, 526–540. [Google Scholar] [CrossRef]
- Petitjean, H.; Pawlowski, S.A.; Fraine, S.L.; Sharif, B.; Hamad, D.; Fatima, T.; Berg, J.; Brown, C.M.; Jan, L.Y.; Ribeiro-da-Silva, A.; et al. Dorsal Horn Parvalbumin Neurons Are Gate-Keepers of Touch-Evoked Pain after Nerve Injury. Cell Rep. 2015, 13, 1246–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Miao, X.; Liang, L.; Abdus-Saboor, I.; Olson, W.; Fleming, M.S.; Ma, M.; Tao, Y.X.; Luo, W. Identification of Early RET+ Deep Dorsal Spinal Cord Interneurons in Gating Pain. Neuron 2016, 91, 1137–1153. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, A.L.; Kovatsis, E.M.; Pozsgai, R.Y.; Tasnim, A.; Zhang, Q.; Ginty, D.D. Distinct Modes of Presynaptic Inhibition of Cutaneous Afferents and Their Functions in Behavior. Neuron 2019, 102, 420–434. [Google Scholar] [CrossRef] [Green Version]
- Garcá-Ramrez, D.L.; Calvo, J.R.; Hochman, S.; Quevedo, J.N. Serotonin, Dopamine and Noradrenaline Adjust Actions of Myelinated Afferents via Modulation of Presynaptic Inhibition in the Mouse Spinal Cord. PLoS ONE 2014, 9, e89999. [Google Scholar] [CrossRef]
- Bardoni, R. Serotonergic Modulation of Nociceptive Circuits in Spinal Cord Dorsal Horn. Curr. Neuropharmacol. 2019, 17, 1133–1145. [Google Scholar] [CrossRef]
- Mena-Avila, E.; Milla-Cruz, J.J.; Calvo, J.R.; Hochman, S.; Villalón, C.M.; Arias-Montaño, J.A.; Quevedo, J.N. Activation of α-Adrenoceptors Depresses Synaptic Transmission of Myelinated Afferents and Inhibits Pathways Mediating Primary Afferent Depolarization (PAD) in the In Vitro Mouse Spinal Cord. Exp. Brain. Res. 2020, 238, 1293–1303. [Google Scholar] [CrossRef]
- Milla-Cruz, J.J.; Mena-Avila, E.; Calvo, J.R.; Hochman, S.; Villalón, C.M.; Quevedo, J.N. The Activation of D2 and D3 Receptor Subtypes Inhibits Pathways Mediating Primary Afferent Depolarization (PAD) in the Mouse Spinal Cord. Neurosci. Lett. 2020, 736, 135257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, S.; Rodriguez, E.; Takatoh, J.; Han, B.X.; Zhou, X.; Wang, F. Identifying Local and Descending Inputs for Primary Sensory Neurons. J. Clin. Invest. 2015, 125, 3782–3794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- François, A.; Low, S.A.; Sypek, E.I.; Christensen, A.J.; Sotoudeh, C.; Beier, K.T.; Ramakrishnan, C.; Ritola, K.D.; Sharif-Naeini, R.; Deisseroth, K.; et al. A Brainstem-Spinal Cord Inhibitory Circuit for Mechanical Pain Modulation by GABA and Enkephalins. Neuron 2017, 93, 822–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-López, Y.; Pérez-Sánchez, J.; Martínez-Lorenzana, G.; Condés-Lara, M.; Rojas-Piloni, G. Cortical Presynaptic Control of Dorsal Horn C-Afferents in the Rat. PLoS ONE 2013, 8, 1–10. [Google Scholar] [CrossRef]
- Sekiguchi, K.J.; Shekhtmeyster, P.; Merten, K.; Arena, A.; Cook, D.; Hoffman, E.; Ngo, A.; Nimmerjahn, A. Imaging Large-Scale Cellular Activity in Spinal Cord of Freely Behaving Mice. Nat. Commun. 2016, 7, 11450. [Google Scholar] [CrossRef]
- Bardoni, R.; Ghirri, A.; Zonta, M.; Betelli, C.; Vitale, G.; Ruggieri, V.; Sandrini, M.; Carmignoto, G. Glutamate-Mediated Astrocyte-to-Neuron Signalling in the Rat Dorsal Horn. J. Physiol. 2010, 588, 831–846. [Google Scholar] [CrossRef]
- Ficker, C.; Rozmer, K.; Kató, E.; Andó, R.D.; Schumann, L.; Krügel, U.; Franke, H.; Sperlágh, B.; Riedel, T.; Illes, P. Astrocyte-Neuron Interaction in the Substantia Gelatinosa of the Spinal Cord Dorsal Horn via P2X7 Receptor-Mediated Release of Glutamate and Reactive Oxygen Species. Glia 2014, 62, 1671–1686. [Google Scholar] [CrossRef] [Green Version]
- Christensen, R.K.; Delgado-Lezama, R.; Russo, R.E.; Lind, B.L.; Alcocer, E.L.; Rath, M.F.; Fabbiani, G.; Schmitt, N.; Lauritzen, M.; Petersen, A.V.; et al. Spinal Dorsal Horn Astrocytes Release GABA in Response to Synaptic Activation. J. Physiol. 2018, 596, 4983–4994. [Google Scholar] [CrossRef] [Green Version]
- Russo, R.E.; Delgado-Lezama, R.; Hounsgaard, J. Dorsal Root Potential Produced by a TTX-Insensitive Micro-Circuitry in the Turtle Spinal Cord. J. Physiol. 2000, 528, 115–122. [Google Scholar] [CrossRef]
- Guo, D.; Hu, J. Spinal Presynaptic Inhibition in Pain Control. Neuroscience 2014, 283, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Gradwell, M.A.; Callister, R.J.; Graham, B.A. Reviewing the Case for Compromised Spinal Inhibition in Neuropathic Pain. J. Neural. Transm. 2020, 127, 481–503. [Google Scholar] [CrossRef]
- Cervero, F.; Laird, J.M. Mechanisms of Touch-Evoked Pain (Allodynia): A New Model. Pain 1996, 68, 13–23. [Google Scholar] [CrossRef]
- Cervero, F.; Laird, J.M.A.; García-Nicas, E. Secondary Hyperalgesia and Presynaptic Inhibition: An Update. Eur. J. Pain 2003, 7, 345–351. [Google Scholar] [CrossRef]
- Rees, H.; Sluka, K.A.; Westlund, K.N.; Willis, W.D. Do Dorsal Root Reflexes Augment Peripheral Inflammation? Neuroreport 1994, 5, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Sluka, K.A.; Rees, H.; Westlund, K.N.; Willis, W.D. Fiber Types Contributing to Dorsal Root Reflexes Induced by Joint Inflammation in Cats and Monkeys. J. Neurophysiol. 1995, 74, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Wu, J.; Willis, W.D. Dorsal Root Reflexes and Cutaneous Neurogenic Inflammation after Intradermal Injection of Capsaicin in Rats. J. Neurophysiol. 1999, 82, 2602–2611. [Google Scholar] [CrossRef]
- Funk, K.; Woitecki, A.; Franjic-Würtz, C.; Gensch, T.; Möhrlen, F.; Frings, S. Modulation of Chloride Homeostasis by Inflammatory Mediators in Dorsal Root Ganglion Neurons. Mol. Pain. 2008, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Galan, A.; Cervero, F. Painful Stimuli Induce in Vivo Phosphorylation and Membrane Mobilization of Mouse Spinal Cord NKCC1 Co-Transporter. Neuroscience 2005, 133, 245–252. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Pattison, L.A.; Singhal, K.; Hockley, J.R.F.; Callejo, G.; Smith, E.S.J. Acute Inflammation Sensitizes Knee-Innervating Sensory Neurons and Decreases Mouse Digging Behavior in a TRPV1-Dependent Manner. Neuropharmacology 2018, 143, 49–62. [Google Scholar] [CrossRef]
- Bráz, J.M.; Sharif-Naeini, R.; Vogt, D.; Kriegstein, A.; Alvarez-Buylla, A.; Rubenstein, J.L.; Basbaum, A.I. Forebrain GABAergic Neuron Precursors Integrate into Adult Spinal Cord and Reduce Injury-Induced Neuropathic Pain. Neuron 2012, 74, 663–675. [Google Scholar] [CrossRef] [Green Version]
- Pieraut, S.; Lucas, O.; Sangari, S.; Sar, C.; Boudes, M.; Bouffi, C.; Noel, D.; Scamps, F. An Autocrine Neuronal Interleukin-6 Loop Mediates Chloride Accumulation and NKCC1 Phosphorylation in Axotomized Sensory Neurons. J. Neurosci. 2011, 31, 13516–13526. [Google Scholar] [CrossRef]
- Mòdol, L.; Cobianchi, S.; Navarro, X. Prevention of NKCC1 Phosphorylation Avoids Downregulation of KCC2 in Central Sensory Pathways and Reduces Neuropathic Pain after Peripheral Nerve Injury. Pain 2014, 155, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, T.; Tokunaga, A.; Kondo, E.; Miki, K.; Tachibana, T.; Noguchi, K. Change in mRNAs for Neuropeptides and the GABA(A) Receptor in Dorsal Root Ganglion Neurons in a Rat Experimental Neuropathic Pain Model. Pain 1998, 78, 13–26. [Google Scholar] [CrossRef]
- Obata, K.; Yamanaka, H.; Fukuoka, T.; Yi, D.; Tokunaga, A.; Hashimoto, N.; Yoshikawa, H.; Noguchi, K. Contribution of Injured and Uninjured Dorsal Root Ganglion Neurons to Pain Behavior and the Changes in Gene Expression Following Chronic Constriction Injury of the Sciatic Nerve in Rats. Pain 2003, 101, 65–77. [Google Scholar] [CrossRef]
- Lian, Y.; Wang, Y.; Ma, K.; Zhao, L.; Zhang, Z.; Shang, Y.; Si, J.; Li, L. Expression of Gamma-Aminobutyric Acid Type A Receptor A2 Subunit in the Dorsal Root Ganglion of Rats with Sciatic Nerve Injury. Neural. Regen. Res. 2012, 7, 2492–2499. [Google Scholar]
- Obradovic, A.L.; Scarpa, J.; Osuru, H.P.; Weaver, J.L.; Park, J.-Y.; Pathirathna, S.; Peterkin, A.; Lim, Y.; Jagodic, M.M.; Todorovic, S.M.; et al. Silencing the A2 Subunit of γ-Aminobutyric Acid Type A Receptors in Rat Dorsal Root Ganglia Reveals Its Major Role in Antinociception Posttraumatic Nerve Injury. Anesthesiology 2015, 123, 654–667. [Google Scholar] [CrossRef] [Green Version]
- Baba, H.; Ji, R.-R.; Kohno, T.; Moore, K.A.; Ataka, T.; Wakai, A.; Okamoto, M.; Woolf, C.J. Removal of GABAergic Inhibition Facilitates Polysynaptic A Fiber-Mediated Excitatory Transmission to the Superficial Spinal Dorsal Horn. Mol. Cell Neurosci. 2003, 24, 818–830. [Google Scholar] [CrossRef]
- Torsney, C.; MacDermott, A.B. Disinhibition Opens the Gate to Pathological Pain Signaling in Superficial Neurokinin 1 Receptor-Expressing Neurons in Rat Spinal Cord. J. Neurosci. 2006, 26, 1833–1843. [Google Scholar] [CrossRef] [Green Version]
- Duan, B.; Cheng, L.; Bourane, S.; Britz, O.; Padilla, C.; Garcia-Campmany, L.; Krashes, M.; Knowlton, W.; Velasquez, T.; Ren, X.; et al. Identification of Spinal Circuits Transmitting and Gating Mechanical Pain. Cell 2014, 159, 1417–1432. [Google Scholar] [CrossRef] [Green Version]
- Peirs, C.; Williams, S.-P.G.; Zhao, X.; Arokiaraj, C.M.; Ferreira, D.W.; Noh, M.-C.; Smith, K.M.; Halder, P.; Corrigan, K.A.; Gedeon, J.Y.; et al. Mechanical Allodynia Circuitry in the Dorsal Horn Is Defined by the Nature of the Injury. Neuron 2020. [Google Scholar] [CrossRef]
- Leitner, J.; Westerholz, S.; Heinke, B.; Forsthuber, L.; Wunderbaldinger, G.; Jäger, T.; Gruber-Schoffnegger, D.; Braun, K.; Sandkühler, J. Impaired Excitatory Drive to Spinal Gabaergic Neurons of Neuropathic Mice. PLoS ONE 2013, 8, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Chen, S.R.; Pan, H.L. Presynaptic NMDA Receptors Control Nociceptive Transmission at the Spinal Cord Level in Neuropathic Pain. Cell. Mol. Life Sci. 2019, 76, 1889–1899. [Google Scholar] [CrossRef] [PubMed]
- Nickolls, S.A.; Gurrell, R.; van Amerongen, G.; Kammonen, J.; Cao, L.; Brown, A.R.; Stead, C.; Mead, A.; Watson, C.; Hsu, C.; et al. Pharmacology in Translation: The Preclinical and Early Clinical Profile of the Novel A2/3 Functionally Selective GABAA Receptor Positive Allosteric Modulator PF-06372865. Br. J. Pharmacol. 2018, 175, 708–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeilhofer, H.U.; Neumann, E.; Munro, G. Spinal GABAA Receptors for Pain Control: Back to the Future? Br. J. Anaesth. 2019, 123, e176–e179. [Google Scholar] [CrossRef]
- Brennan, P.M.; Whittle, I.R. Intrathecal Baclofen Therapy for Neurological Disorders: A Sound Knowledge Base but Many Challenges Remain. Br. J. Neurosurg. 2008, 22, 508–519. [Google Scholar] [CrossRef]
- Kalinichev, M.; Donovan-Rodriguez, T.; Girard, F.; Haddouk, H.; Royer-Urios, I.; Schneider, M.; Bate, S.T.; Marker, C.; Pomonis, J.D.; Poli, S. ADX71943 and ADX71441, Novel Positive Allosteric Modulators of the GABAB Receptor with Distinct Central/Peripheral Profiles, Show Efficacy in the Monosodium Iodoacetate Model of Chronic Osteoarthritis Pain in the Rat. Eur. J. Pharmacol. 2017, 795, 43–49. [Google Scholar] [CrossRef]
- Zemoura, K.; Ralvenius, W.T.; Malherbe, P.; Benke, D. The Positive Allosteric GABAB Receptor Modulator Rac-BHFF Enhances Baclofen-Mediated Analgesia in Neuropathic Mice. Neuropharmacology 2016, 108, 172–178. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Comitato, A.; Bardoni, R. Presynaptic Inhibition of Pain and Touch in the Spinal Cord: From Receptors to Circuits. Int. J. Mol. Sci. 2021, 22, 414. https://doi.org/10.3390/ijms22010414
Comitato A, Bardoni R. Presynaptic Inhibition of Pain and Touch in the Spinal Cord: From Receptors to Circuits. International Journal of Molecular Sciences. 2021; 22(1):414. https://doi.org/10.3390/ijms22010414
Chicago/Turabian StyleComitato, Antonella, and Rita Bardoni. 2021. "Presynaptic Inhibition of Pain and Touch in the Spinal Cord: From Receptors to Circuits" International Journal of Molecular Sciences 22, no. 1: 414. https://doi.org/10.3390/ijms22010414
APA StyleComitato, A., & Bardoni, R. (2021). Presynaptic Inhibition of Pain and Touch in the Spinal Cord: From Receptors to Circuits. International Journal of Molecular Sciences, 22(1), 414. https://doi.org/10.3390/ijms22010414