The Many Faces of CD4+ T Cells: Immunological and Structural Characteristics


 and
and 
Abstract
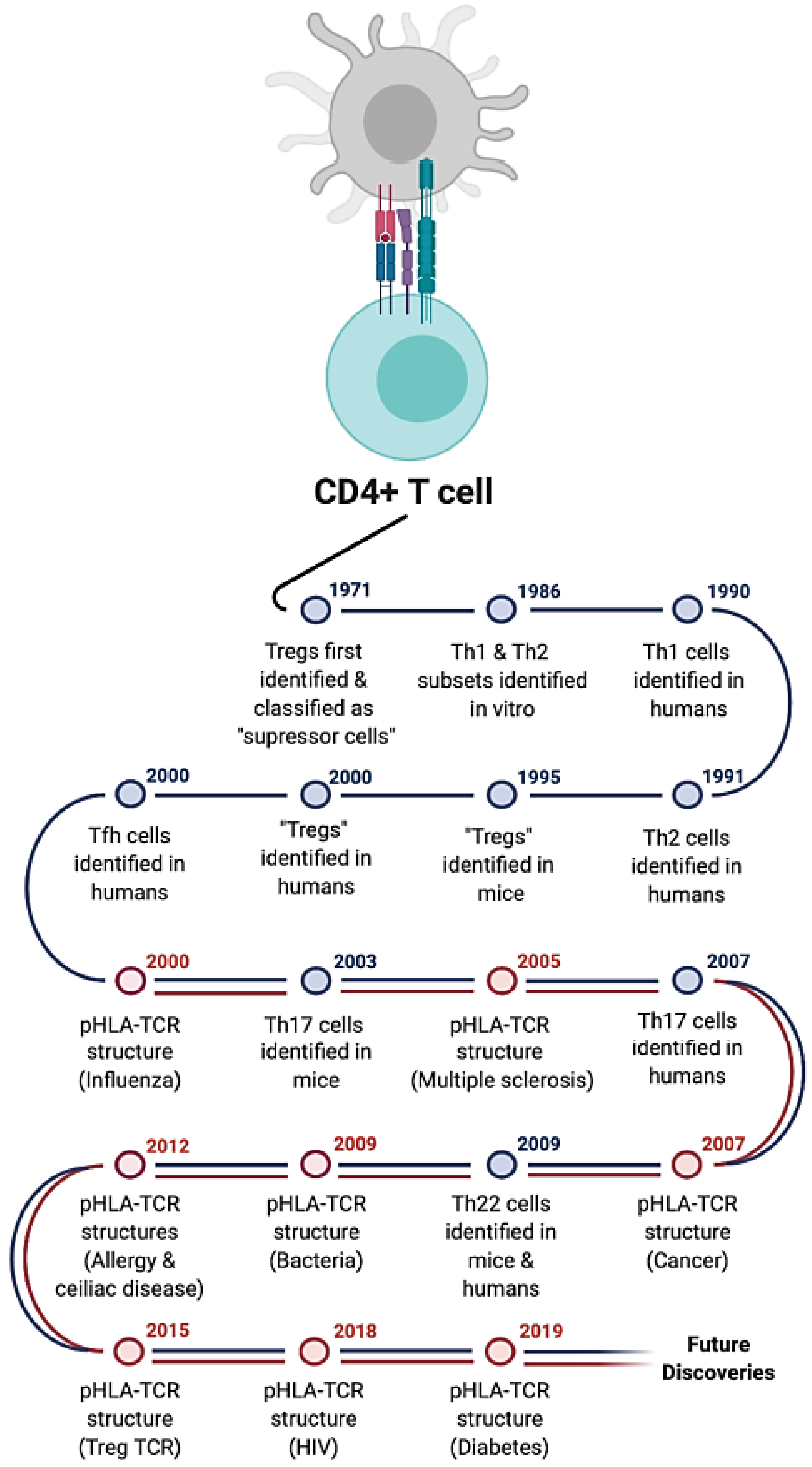
:1. Introduction
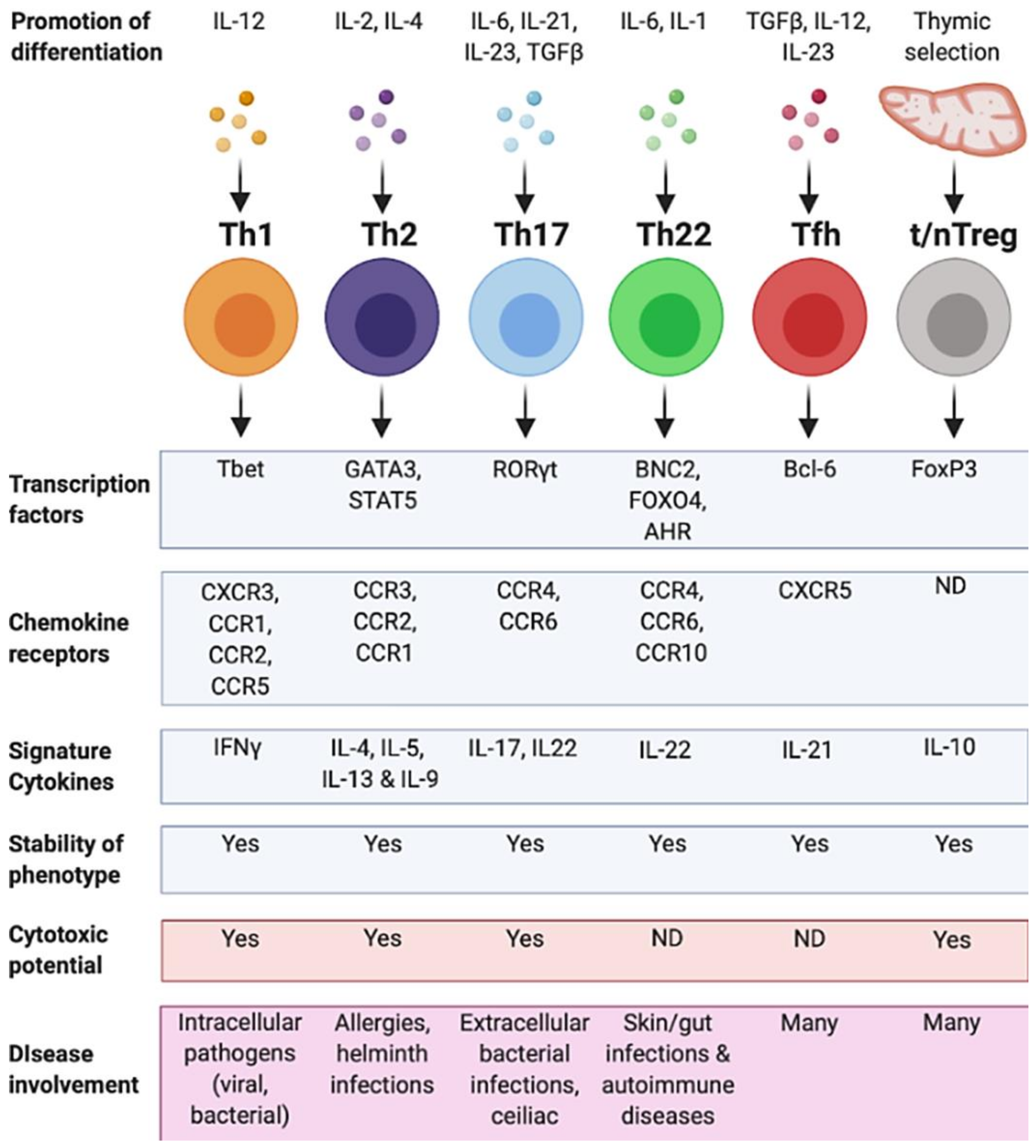
2. T Helper 1 Cells (Th1)
2.1. Overview of Th1 Cell Features
2.2. Antiviral Role of CD4+ Th1 Cells
2.3. Antibacterial Role of CD4+ Th1 Cells
2.4. Role of Th1 CD4+ T Cells in Autoimmunity
2.5. Role of CD4+ Th1 Cells in Cancer
2.6. CD4+ Th1 Cells with Cytotoxic Properties
3. T Helper 2 Cells (Th2)
3.1. Defining Features of Th2 Cells
3.2. Role of Th2 Cells in Atopic Disease
3.3. IL-9 Producing Th2 Cells
4. T Helper 17 Cells (Th17)
4.1. Defining Features of Th17 Cells
4.2. Role of Th17 Cells in Autoimmune Diseases
4.3. Role of Th17 Cells in Infectious Diseases
4.4. The Role of Th17 Cells in Other Types of Disease
5. T Helper 22 Cells (Th22)
5.1. Features of Th22 Cells
5.2. Role of Th22 Cells in the Immune System
6. Regulatory T Cells (Tregs)
6.1. Defining Features of TREGS
6.2. Tregs Are Critical to the Immune System
6.3. Tregs Are an Attractive Target for Immunotherapy
6.4. The Dark Side of TREGS, How Much Is too Much?
7. T Follicular Helper Cells (Tfh)
7.1. Defining Features of Tfh Cells
7.2. Role of Tfh Cells in Autoimmune Diseases
7.3. Role of Tfh Cells in Infectious Diseases
7.4. Role of Tfh Cells in Cancer
8. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Rossjohn, J.; Gras, S.; Miles, J.J.; Turner, S.J.; Godfrey, D.I.; McCluskey, J. T cell antigen receptor recognition of antigen-presenting molecules. Annu. Rev. Immunol. 2015, 33, 169–200. [Google Scholar] [CrossRef]
- Ohno, Y.; Kitamura, H.; Takahashi, N.; Ohtake, J.; Kaneumi, S.; Sumida, K.; Homma, S.; Kawamura, H.; Minagawa, N.; Shibasaki, S.; et al. IL-6 down-regulates HLA class II expression and IL-12 production of human dendritic cells to impair activation of antigen-specific CD4(+) T cells. Cancer Immunol. Immunother. 2016, 65, 193–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijvers, L.; Melief, M.-J.; Van Langelaar, J.; Vries, R.M.V.D.V.D.; Wierenga-Wolf, A.F.; Koetzier, S.C.; Priatel, J.; Jorritsma, T.; Van Ham, S.M.; Hintzen, R.Q.; et al. The Role of Autoimmunity-Related Gene CLEC16A in the B Cell Receptor-Mediated HLA Class II Pathway. J. Immunol. 2020, 205, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Codolo, G.; Toffoletto, M.; Chemello, F.; Coletta, S.; Teixidor, G.S.; Battaggia, G.; Munari, G.; Fassan, M.; Cagnin, S.; De Bernard, M. Helicobacter pylori Dampens HLA-II Expression on Macrophages via the Up-Regulation of miRNAs Targeting CIITA. Front. Immunol. 2020, 10, 2923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wosen, J.E.; Mukhopadhyay, D.; Macaubas, C.; Mellins, E.D. Epithelial MHC Class II Expression and Its Role in Antigen Presentation in the Gastrointestinal and Respiratory Tracts. Front. Immunol. 2018, 9, 2144. [Google Scholar] [CrossRef]
- Davis, M.M.; Boniface, J.J.; Reich, Z.; Lyons, D.; Hampl, J.; Arden, B.; Chien, Y.H. Ligand recognition by alpha beta T cell receptors. Annu Rev. Immunol. 1998, 16, 523–544. [Google Scholar] [CrossRef]
- Ridge, J.P.; Di Rosa, F.; Matzinger, P. A conditioned dendritic cell can be a temporal bridge between a CD4+ T-helper and a T-killer cell. Nature 1998, 393, 474–478. [Google Scholar] [CrossRef]
- Claman, H.N.; Chaperon, E.A.; Triplett, R.F. Thymus-marrow cell combinations. Synergism in antibody production. Exp. Biol. Med. 1966, 122, 1167–1171. [Google Scholar] [CrossRef]
- Appay, V.; Zaunders, J.J.; Papagno, L.; Sutton, J.; Jaramillo, A.; Waters, A.; Easterbrook, P.; Grey, P.; Smith, D.; McMichael, A.J.; et al. Characterization of CD4(+) CTLs ex vivo. J. Immunol. 2002, 168, 5954–5958. [Google Scholar] [CrossRef] [Green Version]
- Galperin, M.; Farenc, C.; Mukhopadhyay, M.; Jayasinghe, D.; Decroos, A.; Benati, D.; Tan, L.L.; Ciacchi, L.; Reid, H.H.; Rossjohn, J.; et al. CD4(+) T cell-mediated HLA class II cross-restriction in HIV controllers. Sci. Immunol. 2018, 3, eaat0687. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, T.M.; Li, C.K.F.; Chui, C.S.C.; Huang, A.K.Y.; Perkins, M.; Liebner, J.C.; Lambkin-Williams, R.; Gilbert, A.; Oxford, J.; Nicholas, B.; et al. Preexisting influenza-specific CD4+ T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 2012, 18, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Pachnio, A.; Ciaurriz, M.; Begum, J.; Lal, N.; Zuo, J.; Beggs, A.; Moss, P. Cytomegalovirus Infection Leads to Development of High Frequencies of Cytotoxic Virus-Specific CD4+ T Cells Targeted to Vascular Endothelium. PLoS Pathog. 2016, 12, e1005832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juno, J.A.; Van Bockel, D.; Kent, S.J.; Kelleher, A.D.; Zaunders, J.J.; Munier, C.M.L. Cytotoxic CD4 T Cells-Friend or Foe during Viral Infection? Front. Immunol. 2017, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of effector CD4 T cell populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenshields-Watson, A.; Attaf, M.; MacLachlan, B.J.; Whalley, T.; Rius, C.; Wall, A.; Lloyd, A.; Hughes, H.; Strange, K.E.; Mason, G.H.; et al. CD4(+) T Cells Recognize Conserved Influenza A Epitopes through Shared Patterns of V-Gene Usage and Complementary Biochemical Features. Cell Rep. 2020, 32, 107885. [Google Scholar] [CrossRef] [PubMed]
- Hennecke, J.; Carfi, A.; Wiley, D.C. Structure of a covalently stabilized complex of a human alphabeta T-cell receptor, influenza HA peptide and MHC class II molecule, HLA-DR1. EMBO J. 2000, 19, 5611–5624. [Google Scholar] [CrossRef] [Green Version]
- Gras, S.; Van Rhijn, I.; Shahine, A.; Cheng, T.Y.; Bhati, M.; Tan, L.L.; Halim, H.; Tuttle, K.D.; Gapin, L.; Le Nours, J.; et al. T cell receptor recognition of CD1b presenting a mycobacterial glycolipid. Nat. Commun. 2016, 7, 13257. [Google Scholar] [CrossRef]
- Gras, S.; Van Rhijn, I.; Shahine, A.; Cheng, T.-Y.; Bhati, M.; Tan, L.L.; Halim, H.; Tuttle, K.D.; Gapin, L.; Le Nours, J.; et al. How C-terminal additions to insulin B-chain fragments create superagonists for T cells in mouse and human type 1 diabetes. Nat. Commun. 2016, 7, 13257. [Google Scholar] [CrossRef]
- Petersen, J.; Montserrat, V.; Mujico, J.R.; Loh, K.L.; Beringer, D.X.; Van Lummel, M.; Thompson, A.; Mearin, M.L.; Schweizer, J.; Kooy-Winkelaar, Y.; et al. T-cell receptor recognition of HLA-DQ2-gliadin complexes associated with celiac disease. Nat. Struct. Mol. Biol. 2014, 21, 480–488. [Google Scholar] [CrossRef]
- Ting, Y.T.; Dahal-Koirala, S.; Kim, H.S.K.; Qiao, S.-W.; Neumann, R.S.; Lundin, K.E.A.; Petersen, J.; Reid, H.H.; Sollid, L.M.; Rossjohn, J. A molecular basis for the T cell response in HLA-DQ2.2 mediated celiac disease. Proc. Natl. Acad. Sci. USA 2020, 117, 3063–3073. [Google Scholar] [CrossRef]
- Petersen, J.; Van Bergen, J.; Loh, K.L.; Kooy-Winkelaar, Y.; Beringer, D.X.; Thompson, A.; Bakker, S.F.; Mulder, C.J.J.; Ladell, K.; McLaren, J.E.; et al. Determinants of gliadin-specific T cell selection in celiac disease. J. Immunol. 2015, 194, 6112–6122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, J.; Kooy-Winkelaar, Y.; Loh, K.L.; Tran, M.; Van Bergen, J.; Koning, F.; Rossjohn, J.; Reid, H.H. Diverse T Cell Receptor Gene Usage in HLA-DQ8-Associated Celiac Disease Converges into a Consensus Binding Solution. Structure 2016, 24, 1643–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broughton, S.E.; Petersen, J.; Theodossis, A.; Scally, S.W.; Loh, K.L.; Thompson, A.; Van Bergen, J.; Kooy-Winkelaar, Y.; Henderson, K.N.; Beddoe, T.; et al. Biased T cell receptor usage directed against human leukocyte antigen DQ8-restricted gliadin peptides is associated with celiac disease. Immunity 2012, 37, 611–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Langley, R.J.; Wang, Q.; Topalian, S.L.; Mariuzza, R.A. Structural insights into the editing of germ-line-encoded interactions between T-cell receptor and MHC class II by Valpha CDR3. Proc. Natl. Acad. Sci. USA 2012, 109, 14960–14965. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Langley, R.J.; Brown, P.H.; Xu, G.; Teng, L.; Wang, Q.; I Gonzales, M.; Callender, G.G.; I Nishimura, M.; Topalian, S.L.; et al. Structural basis for the recognition of mutant self by a tumor-specific, MHC class II-restricted T cell receptor. Nat. Immunol. 2007, 8, 398–408. [Google Scholar] [CrossRef]
- Yin, L.; Crawford, F.; Marrack, P.; Kappler, J.W.; Dai, S. T-cell receptor (TCR) interaction with peptides that mimic nickel offers insight into nickel contact allergy. Proc. Natl. Acad. Sci. USA 2012, 109, 18517–18522. [Google Scholar] [CrossRef] [Green Version]
- Clayton, G.M.; Wang, Y.; Crawford, F.; Novikov, A.; Wimberly, B.T.; Kieft, J.S.; Falta, M.T.; Bowerman, N.A.; Marrack, P.; Fontenot, A.P.; et al. Structural basis of chronic beryllium disease: Linking allergic hypersensitivity and autoimmunity. Cell 2014, 158, 132–142. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.; Nicholson, M.J.; Pyrdol, J.; Wucherpfennig, K.W. Unconventional topology of self peptide-major histocompatibility complex binding by a human autoimmune T cell receptor. Nat. Immunol. 2005, 6, 490–496. [Google Scholar] [CrossRef]
- Li, Y.; Huang, Y.; Lue, J.; Quandt, J.A.; Martin, R.; Mariuzza, R.A. Structure of a human autoimmune TCR bound to a myelin basic protein self-peptide and a multiple sclerosis-associated MHC class II molecule. EMBO J. 2005, 24, 2968–2979. [Google Scholar] [CrossRef] [Green Version]
- Harkiolaki, M.; Holmes, S.L.; Svendsen, P.; Gregersen, J.W.; Jensen, L.T.; McMahon, R.; Friese, M.A.; Van Boxel, G.; Etzensperger, R.; Tzartos, J.S.; et al. T cell-mediated autoimmune disease due to low-affinity crossreactivity to common microbial peptides. Immunity 2009, 30, 348–357. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Li, Y.; Kerzic, M.C.; Martin, R.; A Mariuzza, R. Structure of a TCR with high affinity for self-antigen reveals basis for escape from negative selection. EMBO J. 2011, 30, 1137–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethi, D.K.; Schubert, D.A.; Anders, A.-K.; Heroux, A.; Bonsor, D.A.; Thomas, C.P.; Sundberg, E.J.; Pyrdol, J.; Wucherpfennig, K.W. A highly tilted binding mode by a self-reactive T cell receptor results in altered engagement of peptide and MHC. J. Exp. Med. 2011, 208, 91–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, J.; Ciacchi, L.; Tran, M.T.; Loh, K.L.; Kooy-Winkelaar, Y.; Croft, N.P.; Hardy, M.Y.; Chen, Z.; McCluskey, J.; Anderson, R.P.; et al. T cell receptor cross-reactivity between gliadin and bacterial peptides in celiac disease. Nat. Struct. Mol. Biol. 2019, 27, 49–61. [Google Scholar] [CrossRef]
- Beringer, D.X.; Kleijwegt, F.S.; Wiede, F.; Van Der Slik, A.R.; Loh, K.L.; Petersen, J.; Dudek, N.L.; Duinkerken, G.; Laban, S.; Joosten, A.M.; et al. T cell receptor reversed polarity recognition of a self-antigen major histocompatibility complex. Nat. Immunol. 2015, 16, 1153–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosmann, T.R.; Cherwinski, H.; Bond, M.W.; A Giedlin, M.; Coffman, R.L. Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 1986, 136, 2348–2357. [Google Scholar] [PubMed]
- Wierenga, E.A.; Snoek, M.; De Groot, C.; Chrétien, I.; Bos, J.D.; Jansen, H.M.; Kapsenberg, M.L. Evidence for compartmentalization of functional subsets of CD2+ T lymphocytes in atopic patients. J. Immunol. 1990, 144, 4651–4656. [Google Scholar]
- Pace, J.L.; Russell, S.W.; Schreiber, R.D.; Altman, A.; Katz, D.H. Macrophage activation: Priming activity from a T-cell hybridoma is attributable to interferon-gamma. Proc. Natl. Acad. Sci. USA 1983, 80, 3782–3786. [Google Scholar] [CrossRef] [Green Version]
- Keller, R.; Fischer, W.; Keist, R.; Bassetti, S. Macrophage response to bacteria: Induction of marked secretory and cellular activities by lipoteichoic acids. Infect. Immun. 1992, 60, 3664–3672. [Google Scholar] [CrossRef] [Green Version]
- Muller, E.; Christopoulos, P.F.; Halder, S.; Lunde, A.; Beraki, K.; Speth, M.; Oynebraten, I.; Corthay, A. Toll-Like Receptor Ligands and Interferon-gamma Synergize for Induction of Antitumor M1 Macrophages. Front. Immunol. 2017, 8, 1383. [Google Scholar] [CrossRef]
- Mullen, A.C.; High, F.A.; Hutchins, A.S.; Lee, H.W.; Villarino, A.V.; Livingston, D.M.; Kung, A.L.; Cereb, N.; Yao, T.-P.; Yang, S.Y.; et al. Role of T-bet in commitment of TH1 cells before IL-12-dependent selection. Science 2001, 292, 1907–1910. [Google Scholar] [CrossRef]
- Usui, T.; Preiss, J.C.; Kanno, Y.; Yao, Z.J.; Bream, J.H.; O′Shea, J.J.; Strober, W. T-bet regulates Th1 responses through essential effects on GATA-3 function rather than on IFNG gene acetylation and transcription. J. Exp. Med. 2006, 203, 755–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usui, T.; Nishikomori, R.; Kitani, A.; Strober, W. GATA-3 suppresses Th1 development by downregulation of Stat4 and not through effects on IL-12Rbeta2 chain or T-bet. Immunity 2003, 18, 415–428. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.S.; E Macatonia, S.; Tripp, C.S.; Wolf, S.F.; O’Garra, A.; Murphy, K.M. Development of TH1 CD4+ T cells through IL-12 produced by Listeria-induced macrophages. Science 1993, 260, 547–549. [Google Scholar] [CrossRef] [PubMed]
- Gagliani, N.; Huber, S. Basic Aspects of T Helper Cell Differentiation. Methods Mol. Biol. 2016, 1514, 19–30. [Google Scholar] [CrossRef]
- Szabo, S.J.; Kim, S.T.; Costa, G.L.; Zhang, X.; Fathman, C.; Glimcher, L.H. A novel transcription factor, T-bet, directs Th1 lineage commitment. Cell 2000, 100, 655–669. [Google Scholar] [CrossRef] [Green Version]
- Szabo, S.J.; Kim, S.T.; Costa, G.L.; Zhang, X.; Fathman, C.G.; Glimcher, L.H. Pillars article: A novel transcription factor, T-bet, directs Th1 lineage commitment. J. Immunol. 2015, 194, 2961–2975. [Google Scholar]
- Szabo, S.J.; Sullivan, B.M.; Stemmann, C.; Satoskar, A.R.; Sleckman, B.P.; Glimcher, L.H. Distinct effects of T-bet in TH1 lineage commitment and IFN-gamma production in CD4 and CD8 T cells. Science 2002, 295, 338–342. [Google Scholar] [CrossRef]
- Deng, N.; Weaver, J.M.; Mosmann, T.R. Cytokine diversity in the Th1-dominated human anti-influenza response caused by variable cytokine expression by Th1 cells, and a minor population of uncommitted IL-2+IFNgamma- Thpp cells. PLoS ONE 2014, 9, e95986. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, M.; Christensen, J.R.; Veldhoen, M.; Murphy, T.L.; Murphy, K.M.; Ogarra, A. Interleukin-10 production by Th1 cells requires interleukin-12-induced STAT4 transcription factor and ERK MAP kinase activation by high antigen dose. Immunity 2009, 31, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.E.; Hassan, M.; Burton, B.R.; Britton, G.; Hill, E.V.; Verhagen, J.; Wraith, D.C. IL-4 enhances IL-10 production in Th1 cells: Implications for Th1 and Th2 regulation. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Austin, L.M.; Ozawa, M.; Kikuchi, T.; Walters, I.B.; Krueger, J.G. The majority of epidermal T cells in Psoriasis vulgaris lesions can produce type 1 cytokines, interferon-gamma, interleukin-2, and tumor necrosis factor-alpha, defining TC1 (cytotoxic T lymphocyte) and TH1 effector populations: A type 1 differentiation bias is also measured in circulating blood T cells in psoriatic patients. J. Investig. Dermatol. 1999, 113, 752–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hondowicz, B.D.; Kim, K.S.; Ruterbusch, M.J.; Keitany, G.J.; Pepper, M. IL-2 is required for the generation of viral-specific CD4(+) Th1 tissue-resident memory cells and B cells are essential for maintenance in the lung. Eur. J. Immunol. 2018, 48, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.D.N.T.; Olsen, A.W.; Lorenzen, E.; Andersen, P.; Hvid, M.; Follmann, F.; Dietrich, J. Parenteral vaccination protects against transcervical infection with Chlamydia trachomatis and generate tissue-resident T cells post-challenge. NPJ Vaccines 2020, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, O.; Depil, S.; Mrizak, D.; Martin, N.; Ndour, P.A.; Dufossé, F.; Miroux, C.; Coll, J.; De Launoit, Y.; Auriault, C.; et al. EBV Latency II-derived peptides induce a specific CD4+ cytotoxic T-cell activity and not a CD4+ regulatory T-cell response. J. Immunother. 2012, 35, 254–266. [Google Scholar] [CrossRef]
- Meckiff, B.J.; Ladell, K.; McLaren, J.E.; Ryan, G.B.; Leese, A.M.; James, E.; A Price, D.; Long, H.M. Primary EBV Infection Induces an Acute Wave of Activated Antigen-Specific Cytotoxic CD4(+) T Cells. J. Immunol. 2019, 203, 1276–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.E.; Sedikides, G.X.; Mason, G.M.; Okecha, G.; Wills, M.R. Human Cytomegalovirus (HCMV)-Specific CD4(+) T Cells Are Polyfunctional and Can Respond to HCMV-Infected Dendritic Cells In Vitro. J. Virol. 2017, 91, e02128-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beňová, K.; Hancková, M.; Koči, K.; Kúdelová, M.; Betáková, T. T cells and their function in the immune response to viruses. Acta Virol. 2019, 64, 131–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassotta, A.; Paparoditis, P.; Geiger, R.; Mettu, R.R.; Landry, S.J.; Donati, A.; Benevento, M.; Foglierini, M.; Lewis, D.J.; Lanzavecchia, A.; et al. Deciphering and predicting CD4+ T cell immunodominance of influenza virus hemagglutinin. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Weaver, J.M.; Yang, H.; Roumanes, D.; Lee, F.E.; Wu, H.; Treanor, J.J.; Mosmann, T.R. Increase in IFNgamma(-)IL-2(+) cells in recent human CD4 T cell responses to 2009 pandemic H1N1 influenza. PLoS ONE 2013, 8, e57275. [Google Scholar] [CrossRef]
- Chen, L.; Zanker, D.; Xiao, K.; Wu, C.; Zou, Q.; Chen, W. Immunodominant CD4+ T-cell responses to influenza A virus in healthy individuals focus on matrix 1 and nucleoprotein. J. Virol. 2014, 88, 11760–11773. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Anthony, A.; Oveissi, S.; Huang, M.; Zanker, D.; Xiao, K.; Wu, C.; Zou, Q.; Chen, W. Broad-Based CD4(+) T Cell Responses to Influenza A Virus in a Healthy Individual Who Lacks Typical Immunodominance Hierarchy. Front. Immunol. 2017, 8, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roche, P.A.; Cresswell, P. High-affinity binding of an influenza hemagglutinin-derived peptide to purified HLA-DR. J. Immunol. 1990, 144, 1849–1856. [Google Scholar] [PubMed]
- O’Sullivan, D.; Arrhenius, T.; Sidney, J.; Del Guercio, M.F.; Albertson, M.; Wall, M.; Oseroff, C.; Southwood, S.; Colón, S.M.; Gaeta, F.C.; et al. On the interaction of promiscuous antigenic peptides with different DR alleles. Identification of common structural motifs. J. Immunol. 1991, 147, 2663–2669. [Google Scholar] [PubMed]
- Holland, C.J.; Rizkallah, P.J.; Vollers, S.; Calvo-Calle, J.M.; Madura, F.; Fuller, A.; Sewell, A.K.; Stern, L.J.; Godkin, A.; Cole, D.K. Minimal conformational plasticity enables TCR cross-reactivity to different MHC class II heterodimers. Sci. Rep. 2012, 2, 629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchtenhagen, H.; Rims, C.; Blahnik, G.; Chow, I.-T.; Kwok, W.W.; Buckner, J.H.; James, E. Efficient ex vivo analysis of CD4+ T-cell responses using combinatorial HLA class II tetramer staining. Nat. Commun. 2016, 7, 12614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, F.R.; McLaren, J.E. T Cell Immunity to Bacterial Pathogens: Mechanisms of Immune Control and Bacterial Evasion. Int. J. Mol. Sci. 2020, 21, 6144. [Google Scholar] [CrossRef]
- Green, A.M.; Difazio, R.; Flynn, J.L. IFN-gamma from CD4 T cells is essential for host survival and enhances CD8 T cell function during Mycobacterium tuberculosis infection. J. Immunol. 2013, 190, 270–277. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xiong, X.; Wu, D.; Wang, X.; Wen, B. Efficient activation of T cells by human monocyte-derived dendritic cells (HMDCs) pulsed with Coxiella burnetii outer membrane protein Com1 but not by HspB-pulsed HMDCs. BMC Immunol. 2011, 12, 52. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.F.; Murphy, A.G.; Lalor, S.J.; Leech, J.M.; O’Keeffe, K.M.; Mac Aogáin, M.; O’Halloran, D.P.; Lacey, K.A.; Tavakol, M.; Hearnden, C.H.; et al. Memory Th1 Cells Are Protective in Invasive Staphylococcus aureus Infection. PLoS Pathog. 2015, 11, e1005226. [Google Scholar] [CrossRef]
- Van Rhijn, I.; Kasmar, A.; De Jong, A.; Gras, S.; Bhati, M.; Doorenspleet, M.E.; De Vries, N.; Godfrey, D.I.; Altman, J.D.; De Jager, W.; et al. A conserved human T cell population targets mycobacterial antigens presented by CD1b. Nat. Immunol. 2013, 14, 706–713. [Google Scholar] [CrossRef]
- Chujo, D.; Kawabe, A.; Matsushita, M.; Takahashi, N.; Tsutsumi, C.; Haseda, F.; Imagawa, A.; Hanafusa, T.; Ueki, K.; Kajio, H.; et al. Distinct Phenotypes of Islet Antigen-Specific CD4+ T Cells Among the 3 Subtypes of Type 1 Diabetes. J. Clin. Endocrinol. Metab. 2020, 105, 3141–3151. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yang, C.; Wang, Y.; Song, W.; Jia, L.; Peng, X.; Zhao, R. The Expression of P2X7 Receptor on Th1, Th17, and Regulatory T Cells in Patients with Systemic Lupus Erythematosus or Rheumatoid Arthritis and Its Correlations with Active Disease. J. Immunol. 2020, ji2000222. [Google Scholar] [CrossRef] [PubMed]
- Todd, J.A.; Bell, J.I.; McDevitt, H.O. HLA-DQ beta gene contributes to susceptibility and resistance to insulin-dependent diabetes mellitus. Nature 1987, 329, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Hitman, G.A. The immunogenetics of insulin-dependent diabetes. Eye 1993, 7, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Alleva, D.G.; Crowe, P.D.; Jin, L.; Kwok, W.W.; Ling, N.; Gottschalk, M.; Conlon, P.J.; Gottlieb, P.A.; Putnam, A.L.; Gaur, A. A disease-associated cellular immune response in type 1 diabetics to an immunodominant epitope of insulin. J. Clin. Investig. 2001, 107, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chow, I.T.; Sosinowski, T.; Torres-Chinn, N.; Greenbaum, C.J.; James, E.A.; Kappler, J.W.; Davidson, H.W.; Kwok, W.W. Autoreactive T cells specific for insulin B:11-23 recognize a low-affinity peptide register in human subjects with autoimmune diabetes. Proc. Natl. Acad. Sci. USA 2014, 111, 14840–14845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzarella, G. Effector and suppressor T cells in celiac disease. World J. Gastroenterol. 2015, 21, 7349–7356. [Google Scholar] [CrossRef]
- Festen, E.A.M.; Szperl, A.M.; Weersma, R.K.; Wijmenga, C.; Wapenaar, M.C. Inflammatory bowel disease and celiac disease: Overlaps in the pathology and genetics, and their potential drug targets. Endocr. Metab. Immune Disord. Drug Targets 2009, 9, 199–218. [Google Scholar] [CrossRef]
- Spurkland, A.; Sollid, L.M.; Polanco, I.; Vartdal, F.; Thorsby, E. HLA-DR and -DQ genotypes of celiac disease patients serologically typed to be non-DR3 or non-DR5/7. Hum. Immunol. 1992, 35, 188–192. [Google Scholar] [CrossRef]
- Karell, K.; Louka, A.S.; Moodie, S.J.; Ascher, H.; Clot, F.; Greco, L.; Ciclitira, P.J.; Sollid, L.M.; Partanen, J. European Genetics Cluster on Celiac, D. HLA types in celiac disease patients not carrying the DQA1*05-DQB1*02 (DQ2) heterodimer: Results from the European Genetics Cluster on Celiac Disease. Hum. Immunol. 2003, 64, 469–477. [Google Scholar] [CrossRef]
- Tay, R.E.; Richardson, E.K.; Toh, H.C. Revisiting the role of CD4(+) T cells in cancer immunotherapy-new insights into old paradigms. Cancer Gene Ther. 2020, 1–13. [Google Scholar] [CrossRef]
- Kagi, D.; Vignaux, F.; Ledermann, B.; Burki, K.; Depraetere, V.; Nagata, S.; Hengartner, H.; Golstein, P. Fas and perforin pathways as major mechanisms of T cell-mediated cytotoxicity. Science 1994, 265, 528–530. [Google Scholar] [CrossRef] [PubMed]
- Benati, D.; Galperin, M.; Lambotte, O.; Gras, S.; Lim, A.; Mukhopadhyay, M.; Nouël, A.; Campbell, K.-A.; Lemercier, B.; Claireaux, M.; et al. Public T cell receptors confer high-avidity CD4 responses to HIV controllers. J. Clin. Investig. 2016, 126, 2093–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Prete, G.F.; De Carli, M.; Mastromauro, C.; Biagiotti, R.; Macchia, D.; Falagiani, P.; Ricci, M.; Romagnani, S. Purified protein derivative of Mycobacterium tuberculosis and excretory-secretory antigen(s) of Toxocara canis expand in vitro human T cells with stable and opposite (type 1 T helper or type 2 T helper) profile of cytokine production. J. Clin. Investig. 1991, 88, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Kay, A.B.; Ying, S.; Varney, V.; Gaga, M.; Durham, S.R.; Moqbel, R.; Wardlaw, A.J.; Hamid, Q. Messenger RNA expression of the cytokine gene cluster, interleukin 3 (IL-3), IL-4, IL-5, and granulocyte/macrophage colony-stimulating factor, in allergen-induced late-phase cutaneous reactions in atopic subjects. J. Exp. Med. 1991, 173, 775–778. [Google Scholar] [CrossRef]
- Walker, J.A.; McKenzie, A.N.J. TH2 cell development and function. Nat. Rev. Immunol. 2018, 18, 121–133. [Google Scholar] [CrossRef]
- Cote-Sierra, J.; Foucras, G.; Guo, L.; Chiodetti, L.; Young, H.A.; Hu-Li, J.; Zhu, J.; Paul, W.E. Interleukin 2 plays a central role in Th2 differentiation. Proc. Natl. Acad. Sci. USA 2004, 101, 3880–3885. [Google Scholar] [CrossRef] [Green Version]
- Morawetz, R.A.; Gabriele, L.; Rizzo, L.V.; Noben-Trauth, N.; Kühn, R.; Rajewsky, K.; Müller, W.; Doherty, T.M.; Finkelman, F.; Coffman, R.L.; et al. Interleukin (IL)-4-independent immunoglobulin class switch to immunoglobulin (Ig)E in the mouse. J. Exp. Med. 1996, 184, 1651–1661. [Google Scholar] [CrossRef]
- Sehmi, R.; Wood, L.J.; Watson, R.; Foley, R.; Hamid, Q.; O’Byrne, P.M.; A Denburg, J. Allergen-induced increases in IL-5 receptor alpha-subunit expression on bone marrow-derived CD34+ cells from asthmatic subjects. A novel marker of progenitor cell commitment towards eosinophilic differentiation. J. Clin. Investig. 1997, 100, 2466–2475. [Google Scholar] [CrossRef]
- Robinson, D.S.; Damia, R.; Zeibecoglou, K.; Molet, S.; North, J.; Yamada, T.; Kay, A.B.; Hamid, Q. CD34(+)/interleukin-5Ralpha messenger RNA+ cells in the bronchial mucosa in asthma: Potential airway eosinophil progenitors. Am. J. Respir. Cell Mol. Biol. 1999, 20, 9–13. [Google Scholar] [CrossRef]
- Fulkerson, P.C.; Schollaert, K.L.; Bouffi, C.; Rothenberg, M.E. IL-5 triggers a cooperative cytokine network that promotes eosinophil precursor maturation. J. Immunol. 2014, 193, 4043–4052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.Z.; Wang, X.; Wang, Y.; Niu, A.; Wang, S.; Zou, C.; Harris, R.C. IL-4/IL-13-mediated polarization of renal macrophages/dendritic cells to an M2a phenotype is essential for recovery from acute kidney injury. Kidney Int. 2017, 91, 375–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Homer, R.J.; Wang, Z.; Chen, Q.; Geba, G.P.; Wang, J.; Zhang, Y.; Elias, J.A. Pulmonary expression of interleukin-13 causes inflammation, mucus hypersecretion, subepithelial fibrosis, physiologic abnormalities, and eotaxin production. J. Clin. Investig. 1999, 103, 779–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.S.; Kim, Y.U.; Lee, S.; Jang, S.W.; Kim, M.K.; Koh, B.H.; Lee, W.; Kim, J.; Souabni, A.; Busslinger, M.; et al. Transcription factor YY1 is essential for regulation of the Th2 cytokine locus and for Th2 cell differentiation. Proc. Natl. Acad. Sci. USA 2012, 110, 276–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J. Transcriptional regulation of Th2 cell differentiation. Immunol. Cell Biol. 2010, 88, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.E.; Sutherland, T.E. Host protective roles of type 2 immunity: Parasite killing and tissue repair, flip sides of the same coin. Semin. Immunol. 2014, 26, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Zaiss, M.M.; Maslowski, K.M.; Mosconi, I.; Guenat, N.; Marsland, B.J.; Harris, N.L. IL-1beta suppresses innate IL-25 and IL-33 production and maintains helminth chronicity. PLoS Pathog. 2013, 9, e1003531. [Google Scholar] [CrossRef] [Green Version]
- Bamias, G.; Cominelli, F. Role of type 2 immunity in intestinal inflammation. Curr. Opin. Gastroenterol. 2015, 31, 471–476. [Google Scholar] [CrossRef] [Green Version]
- Harris, N.L.; Loke, P. Recent Advances in Type-2-Cell-Mediated Immunity: Insights from Helminth Infection. Immunity 2017, 47, 1024–1036. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, T.; Hirahara, K.; Onodera, A.; Endo, Y.; Hosokawa, H.; Shinoda, K.; Tumes, D.J.; Okamoto, Y. Th2 Cells in Health and Disease. Annu. Rev. Immunol. 2017, 35, 53–84. [Google Scholar] [CrossRef]
- Mutalithas, K.; Guillen, C.; Day, C.; Brightling, C.; Pavord, I.D.; Wardlaw, A.J. CRTH2 expression on T cells in asthma. Clin. Exp. Immunol. 2010, 161, 34–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seumois, G.; Zapardiel-Gonzalo, J.; White, B.; Singh, D.; Schulten, V.; Dillon, M.; Hinz, D.; Broide, D.H.; Sette, A.; Peters, B.; et al. Transcriptional Profiling of Th2 Cells Identifies Pathogenic Features Associated with Asthma. J. Immunol. 2016, 197, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.W.; Hojer, C.D.; Zhou, M.; Wu, X.; Wuster, A.; Lee, W.P.; Yaspan, B.L.; Chan, A.C. Regulation of T Cell Receptor Signaling by DENND1B in TH2 Cells and Allergic Disease. Cell 2016, 164, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Fiuza, B.S.; Silva, M.D.J.; Alcântara-Neves, N.M.; Costa, R.D.S.; Figueiredo, C.A.; Barreto, M.L. Polymorphisms in DENND1B gene are associated with asthma and atopy phenotypes in Brazilian children. Mol. Immunol. 2017, 90, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Wambre, E.; Bajzik, V.; DeLong, J.H.; O’Brien, K.; Nguyen, Q.A.; Speake, C.; Gersuk, V.H.; DeBerg, H.A.; Whalen, E.; Ni, C.; et al. A phenotypically and functionally distinct human TH2 cell subpopulation is associated with allergic disorders. Sci. Transl. Med. 2017, 9, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlström, M.G.; Thyssen, J.P.; Wennervaldt, M.; Menné, T.; Johansen, J.D. Nickel allergy and allergic contact dermatitis: A clinical review of immunology, epidemiology, exposure, and treatment. Contact Dermat. 2019, 81, 227–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diepgen, T.L.; Ofenloch, R.F.; Bruze, M.; Bertuccio, P.; Cazzaniga, S.; Coenraads, P.J.; Elsner, P.; Goncalo, M.; Svensson, A.; Naldi, L. Prevalence of contact allergy in the general population in different European regions. Br. J. Dermatol. 2016, 174, 319–329. [Google Scholar] [CrossRef]
- Dai, S.; Falta, M.T.; Bowerman, N.A.; McKee, A.S.; Fontenot, A.P. T cell recognition of beryllium. Curr. Opin. Immunol. 2013, 25, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Goswami, R.; Kaplan, M.H. A brief history of IL-9. J. Immunol. 2011, 186, 3283–3288. [Google Scholar] [CrossRef] [Green Version]
- Dardalhon, V.; Awasthi, A.; Kwon, H.; Galileos, G.; Gao, W.; Sobel, R.A.; Mitsdoerffer, M.; Strom, T.B.; Elyaman, W.; Ho, I.C.; et al. IL-4 inhibits TGF-beta-induced Foxp3+ T cells and, together with TGF-beta, generates IL-9+ IL-10+ Foxp3(−) effector T cells. Nat. Immunol. 2008, 9, 1347–1355. [Google Scholar] [CrossRef] [Green Version]
- Veldhoen, M.; Uyttenhove, C.; Snick, J.V.; Helmby, H.; Westendorf, A.; Buer, J.; Martin, B.; Wilhelm, C.; Stockinger, B. Transforming growth factor-beta ‘reprograms’ the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat. Immunol. 2008, 9, 1341–1346. [Google Scholar] [CrossRef] [PubMed]
- Gessner, A.; Blum, H.; Röllinghoff, M. Differential regulation of IL-9-expression after infection with Leishmania major in susceptible and resistant mice. Immunobiology 1993, 189, 419–435. [Google Scholar] [CrossRef]
- Schlapbach, C.; Gehad, A.; Yang, C.; Watanabe, R.; Guenova, E.; Teague, J.E.; Campbell, L.; Yawalkar, N.; Kupper, T.S.; Clark, R.A. Human TH9 cells are skin-tropic and have autocrine and paracrine proinflammatory capacity. Sci. Transl. Med. 2014, 6, 219ra8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micosse, C.; Meyenn, L.V.; Steck, O.; Kipfer, E.; Adam, C.; Simillion, C.; Jafari, S.M.S.; Olah, P.; Yawlkar, N.; Simon, D.; et al. Human “TH9” cells are a subpopulation of PPAR-gamma(+) TH2 cells. Sci. Immunol. 2019, 4, eaat5943. [Google Scholar] [CrossRef]
- Chang, H.C.; Sehra, S.; Goswami, R.; Yao, W.; Yu, Q.; Stritesky, G.L.; Jabeen, R.; McKinley, C.; Ahyi, A.N.; Han, L.; et al. The transcription factor PU.1 is required for the development of IL-9-producing T cells and allergic inflammation. Nat. Immunol. 2010, 11, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Neurath, M.F.; Kaplan, M.H. Th9 cells in immunity and immunopathological diseases. Semin. Immunopathol. 2016, 39, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Nonomura, Y.; Otsuka, A.; Nakashima, C.; Seidel, J.A.; Kitoh, A.; Dainichi, T.; Nakajima, S.; Sawada, Y.; Matsushita, S.; Aoki, M.; et al. Peripheral blood Th9 cells are a possible pharmacodynamic biomarker of nivolumab treatment efficacy in metastatic melanoma patients. OncoImmunology 2016, 5, e1248327. [Google Scholar] [CrossRef]
- Diani, M.; Altomare, G.; Reali, E. T Helper Cell Subsets in Clinical Manifestations of Psoriasis. J. Immunol. Res. 2016, 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Vyas, S.P.; Goswami, R. A Decade of Th9 Cells: Role of Th9 Cells in Inflammatory Bowel Disease. Front. Immunol. 2018, 9, 1139. [Google Scholar] [CrossRef] [Green Version]
- Defendenti, C.; Sarzi-Puttini, P.; Saibeni, S.; Bollani, S.; Bruno, S.; Almasio, P.L.; Declich, P.; Atzeni, F. Significance of serum Il-9 levels in inflammatory bowel disease. Int. J. Immunopathol. Pharmacol. 2015, 28, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Harrington, L.E.; Hatton, R.D.; Mangan, P.R.; Turner, H.; Murphy, T.L.; Murphy, K.M.; Weaver, C.T. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immunol. 2005, 6, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Gagliani, N.; Huber, S.; Flavell, R. Chapter 38: Intestinal Regulatory CD4+ T cells. Mucosal Immunol. 2015, 4, 777–785. [Google Scholar] [CrossRef]
- Sallusto, F. Heterogeneity of Human CD4(+) T Cells Against Microbes. Annu. Rev. Immunol. 2016, 34, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 2006, 24, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.O.; Panopoulos, A.D.; Nurieva, R.; Chang, S.H.; Wang, D.; Watowich, S.S.; Dong, C. STAT3 regulates cytokine-mediated generation of inflammatory helper T cells. J. Biol. Chem. 2007, 282, 9358–9363. [Google Scholar] [CrossRef] [Green Version]
- Luckheeram, R.V.; Zhou, R.; Verma, A.D.; Xia, B. CD4(+)T cells: Differentiation and functions. Clin. Dev. Immunol. 2012, 2012, 925135. [Google Scholar] [CrossRef] [Green Version]
- Hirahara, K.; Nakayama, T. CD4+ T-cell subsets in inflammatory diseases: Beyond the Th1/Th2 paradigm. Int. Immunol. 2016, 28, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Peters, A.; Pitcher, L.A.; Sullivan, J.M.; Mitsdoerffer, M.; Acton, S.E.; Franz, B.; Wucherpfennig, K.; Turley, S.; Carroll, M.C.; Sobel, R.A.; et al. Th17 cells induce ectopic lymphoid follicles in central nervous system tissue inflammation. Immunity 2011, 35, 986–996. [Google Scholar] [CrossRef] [Green Version]
- Kolls, J.K.; Khader, S.A. The role of Th17 cytokines in primary mucosal immunity. Cytokine Growth Factor Rev. 2010, 21, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Zielinski, C.E.; Mele, F.; Aschenbrenner, D.; Jarrossay, D.; Ronchi, F.; Gattorno, M.; Monticelli, S.; Lanzavecchia, A.; Sallusto, F. Pathogen-induced human TH17 cells produce IFN-gamma or IL-10 and are regulated by IL-1beta. Nature 2012, 484, 514–518. [Google Scholar] [CrossRef]
- McGeachy, M.J.; Bak-Jensen, K.S.; Chen, Y.; Tato, C.M.; Blumenschein, W.; McClanahan, T.; Cua, D.J. TGF-beta and IL-6 drive the production of IL-17 and IL-10 by T cells and restrain T(H)-17 cell-mediated pathology. Nat. Immunol. 2007, 8, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Schorer, M.; Kuchroo, V.K.; Joller, N. Role of Co-stimulatory Molecules in T Helper Cell Differentiation. Adv. Exp. Med. Biol. 2019, 1189, 153–177. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Kaveri, S.V.; Bayry, J. Th17 cells, pathogenic or not? TGF-beta3 imposes the embargo. Cell. Mol. Immunol. 2013, 10, 101–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Tian, J.; Wang, S. Insight Into Non-Pathogenic Th17 Cells in Autoimmune Diseases. Front. Immunol. 2018, 9, 1112. [Google Scholar] [CrossRef]
- Ramesh, R.; Kozhaya, L.; McKevitt, K.; Djuretic, I.M.; Carlson, T.J.; Quintero, M.A.; McCauley, J.L.; Abreu, M.T.; Unutmaz, D.; Sundrud, M.S. Pro-inflammatory human Th17 cells selectively express P-glycoprotein and are refractory to glucocorticoids. J. Exp. Med. 2014, 211, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Zambrano-Zaragoza, J.F.; Romo-Martinez, E.J.; Durán-Avelar, M.D.J.; García-Magallanes, N.; Vibanco-Pérez, N. Th17 cells in autoimmune and infectious diseases. Int. J. Inflamm. 2014, 2014, 1–12. [Google Scholar] [CrossRef]
- Patel, D.D.; Kuchroo, V.K. Th17 Cell Pathway in Human Immunity: Lessons from Genetics and Therapeutic Interventions. Immunity 2015, 43, 1040–1051. [Google Scholar] [CrossRef] [Green Version]
- Angelou, C.C.; Wells, A.C.; Vijayaraghavan, J.; Dougan, C.E.; Lawlor, R.; Iverson, E.; Lazarevic, V.; Kimura, M.Y.; Peyton, S.R.; Minter, L.M.; et al. Differentiation of Pathogenic Th17 Cells Is Negatively Regulated by Let-7 MicroRNAs in a Mouse Model of Multiple Sclerosis. Front. Immunol. 2020, 10, 3125. [Google Scholar] [CrossRef] [Green Version]
- Falivene, J.; Ghiglione, Y.; Laufer, N.; Socías, M.E.; Holgado, M.P.; Ruiz, M.J.; Maeto, C.; Figueroa, M.I.; Giavedoni, L.D.; Cahn, P.; et al. Th17 and Th17/Treg ratio at early HIV infection associate with protective HIV-specific CD8(+) T-cell responses and disease progression. Sci. Rep. 2015, 5, 11511. [Google Scholar] [CrossRef] [Green Version]
- Caruso, M.P.; Falivene, J.; Holgado, M.P.; Zurita, D.H.; Laufer, N.; Castro, C.; Nico, Á.; Maeto, C.; Salido, J.; Pérez, H.; et al. Impact of HIV-ART on the restoration of Th17 and Treg cells in blood and female genital mucosa. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef]
- Pandrea, E.; Landay, A.L. Chapter 3: Implications for Therapy. In Models of Protection Against HIV/SIV; Elsevier: Amsterdam, The Netherlands, 2012; pp. 81–132. [Google Scholar]
- Li, Y.; Wei, C.; Xu, H.; Jia, J.; Wei, Z.; Guo, R.; Jia, Y.; Wu, Y.; Li, Y.; Qi, X.; et al. The Immunoregulation of Th17 in Host against Intracellular Bacterial Infection. Mediat. Inflamm. 2018, 2018, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wucherpfennig, K.W.; Strominger, J.L. Molecular mimicry in T cell-mediated autoimmunity: Viral peptides activate human T cell clones specific for myelin basic protein. Cell 1995, 80, 695–705. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, S.; Martin, M.; Gauthier, L.; Wucherpfennig, K.W. Structural features of autoreactive TCR that determine the degree of degeneracy in peptide recognition. J. Immunol. 1999, 162, 338–344. [Google Scholar] [PubMed]
- Socie, G.; Zeiser, R.; Blazar, B.R. Chapter 1: Overview of the Immune Biology of Allogenic HSCT. In Immune Biology of Allogeneic Hematopoietic Stem Celll Transplantation, 2nd ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 1–14. [Google Scholar]
- Buggert, M. Resident TH17 cells "break bad" in kidney autoimmunity. Sci. Immunol. 2020, 5, eabd3638. [Google Scholar] [CrossRef]
- Trifari, S.; Kaplan, C.D.; Tran, E.H.; Crellin, N.K.; Spits, H. Identification of a human helper T cell population that has abundant production of interleukin 22 and is distinct from T(H)-17, T(H)1 and T(H)2 cells. Nat. Immunol. 2009, 10, 864–871. [Google Scholar] [CrossRef]
- Duhen, T.; Geiger, R.; Jarrossay, D.; Lanzavecchia, A.; Sallusto, F. Production of interleukin 22 but not interleukin 17 by a subset of human skin-homing memory T cells. Nat. Immunol. 2009, 10, 857–863. [Google Scholar] [CrossRef]
- Yang, X.; Jiang, H.; Lei, R.; Lu, W.; Tan, S.; Qin, S. Recruitment and significance of Th22 cells and Th17 cells in malignant ascites. Oncol. Lett. 2018, 16, 5389–5397. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, Y.-G.; Li, Y.-H.; Qi, L.; Liu, X.-G.; Yuan, C.-Z.; Hu, N.-W.; Ma, D.-X.; Li, Z.-F.; Yang, Q.; et al. Increased frequencies of Th22 cells as well as Th17 cells in the peripheral blood of patients with ankylosing spondylitis and rheumatoid arthritis. PLoS ONE 2012, 7, e31000. [Google Scholar] [CrossRef]
- Gittler, J.K.; Shemer, A.; Suarez-Farinas, M.; Fuentes-Duculan, J.; Gulewicz, K.J.; Wang, C.Q.; Mitsui, H.; Cardinale, I.; De Guzman, C.S.; Krueger, J.G.; et al. Progressive activation of T(H)2/T(H)22 cytokines and selective epidermal proteins characterizes acute and chronic atopic dermatitis. J. Allergy Clin. Immunol. 2012, 130, 1344–1354. [Google Scholar] [CrossRef] [Green Version]
- Nograles, K.E.; Zaba, L.C.; Shemer, A.; Fuentes-Duculan, J.; Cardinale, I.; Kikuchi, T.; Ramon, M.; Bergman, R.; Krueger, J.G.; Guttman-Yassky, E. IL-22-producing “T22” T cells account for upregulated IL-22 in atopic dermatitis despite reduced IL-17-producing TH17 T cells. J. Allergy Clin. Immunol. 2009, 123, 1244–1252. [Google Scholar] [CrossRef] [Green Version]
- Eyerich, S.; Eyerich, K.; Pennino, D.; Carbone, T.; Nasorri, F.; Pallotta, S.; Cianfarani, F.; Odorisio, T.; Traidl-Hoffmann, C.; Behrendt, H.; et al. Th22 cells represent a distinct human T cell subset involved in epidermal immunity and remodeling. J. Clin. Investig. 2009, 119, 3573–3585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Yin, H.; Zhang, K.; Wang, T.; Yang, Y.; Liu, X.; Chang, X.; Zhang, M.; Yan, X.; Ren, Y.; et al. Effector T helper cell populations are elevated in the bone marrow of rheumatoid arthritis patients and correlate with disease severity. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Nakayamada, S.; Kubo, S.; Nakano, K.; Iwata, S.; Miyagawa, I.; Ma, X.; Trimova, G.; Sakata, K.; Tanaka, Y. Th22 Cells Promote Osteoclast Differentiation via Production of IL-22 in Rheumatoid Arthritis. Front. Immunol. 2018, 9, 2901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jong, A.; Pena-Cruz, V.; Cheng, T.Y.; Clark, R.A.; Van Rhijn, I.; Moody, D.B. CD1a-autoreactive T cells are a normal component of the human alphabeta T cell repertoire. Nat. Immunol. 2010, 11, 1102–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkinshaw, R.W.; Pellicci, D.G.; Cheng, T.Y.; Keller, A.N.; Sandoval-Romero, M.; Gras, S.; de Jong, A.; Uldrich, A.P.; Moody, D.B.; Godfrey, D.I.; et al. Alphabeta T cell antigen receptor recognition of CD1a presenting self lipid ligands. Nat. Immunol. 2015, 16, 258–266. [Google Scholar] [CrossRef]
- Gershon, R.K.; Kondo, K. Cell interactions in the induction of tolerance: The role of thymic lymphocytes. Immunology 1970, 18, 723–737. [Google Scholar]
- Taussig, M.J. Demonstration of suppressor T cells in a population of ‘educated’ T cells. Nature 1974, 248, 236–238. [Google Scholar] [CrossRef]
- Stephens, L.A.; Mottet, C.; Mason, D.; Powrie, F. Human CD4(+)CD25(+) thymocytes and peripheral T cells have immune suppressive activity in vitro. Eur J. Immunol. 2001, 31, 1247–1254. [Google Scholar] [CrossRef]
- Baecher-Allan, C.; Brown, J.A.; Freeman, G.J.; Hafler, D.A. CD4+CD25high regulatory cells in human peripheral blood. J. Immunol. 2001, 167, 1245–1253. [Google Scholar] [CrossRef] [Green Version]
- Dieckmann, D.; Plottner, H.; Berchtold, S.; Berger, T.; Schuler, G. Ex vivo isolation and characterization of CD4(+)CD25(+) T cells with regulatory properties from human blood. J. Exp. Med. 2001, 193, 1303–1310. [Google Scholar] [CrossRef]
- Jonuleit, H.; Schmitt, E.; Stassen, M.; Tuettenberg, A.; Knop, J.; Enk, A.H. Identification and functional characterization of human CD4(+)CD25(+) T cells with regulatory properties isolated from peripheral blood. J. Exp. Med. 2001, 193, 1285–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 1995, 155, 1151–1164. [Google Scholar] [PubMed]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Pillars article: Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 2011, 186, 3808–3821. [Google Scholar] [PubMed]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 2003, 4, 330–336. [Google Scholar] [CrossRef]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Pillars Article: Foxp3 Programs the Development and Function of CD4+CD25+ Regulatory T Cells. Nat. Immunol. 2003. 4: 330–336. Nat. Immunol. 2017, 198, 986–992, reprinted. [Google Scholar]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [Green Version]
- Roncador, G.; Brown, P.J.; Maestre, L.; Hue, S.; Martinez-Torrecuadrada, J.L.; Ling, K.L.; Pratap, S.; Toms, C.; Fox, B.C.; Cerundolo, V.; et al. Analysis of FOXP3 protein expression in human CD4+CD25+ regulatory T cells at the single-cell level. Eur. J. Immunol. 2005, 35, 1681–1691. [Google Scholar] [CrossRef]
- Pillai, V.; Ortega, S.B.; Wang, C.K.; Karandikar, N.J. Transient regulatory T-cells: A state attained by all activated human T-cells. Clin. Immunol. 2007, 123, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Mandapathil, M.; Lang, S.; Gorelik, E.; Whiteside, T.L. Isolation of functional human regulatory T cells (Treg) from the peripheral blood based on the CD39 expression. J. Immunol. Methods 2009, 346, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.; Kretz, C.C.; Krammer, P.H.; Kuhn, A. CD127(low/-) and FoxP3(+) expression levels characterize different regulatory T-cell populations in human peripheral blood. J. Investig. Dermatol. 2010, 130, 492–499. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Li, X.; Song, W.; Li, D.; Yu, D.; Zeng, X.; Li, M.; Leng, X.; Li, X. CD4(+)CD25 (+)CD127 (low/-) T cells: A more specific Treg population in human peripheral blood. Inflammation 2012, 35, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Mason, G.M.; Lowe, K.; Melchiotti, R.; Ellis, R.; De Rinaldis, E.; Peakman, M.; Heck, S.; Lombardi, G.; Tree, T.I.M. Phenotypic Complexity of the Human Regulatory T Cell Compartment Revealed by Mass Cytometry. J. Immunol. 2015, 195, 2030–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krop, J.; Heidt, S.; Claas, F.H.J.; Eikmans, M. Regulatory T Cells in Pregnancy: It Is Not All About FoxP3. Front. Immunol. 2020, 11, 1182. [Google Scholar] [CrossRef] [PubMed]
- Shevach, E.M.; Thornton, A.M. tTregs, pTregs, and iTregs: Similarities and differences. Immunol. Rev. 2014, 259, 88–102. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Jin, W.; Hardegen, N.; Lei, K.J.; Li, L.; Marinos, N.; McGrady, G.; Wahl, S.M. Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J. Exp. Med. 2003, 198, 1875–1886. [Google Scholar] [CrossRef]
- Atwany, N.Z.; Hashemi, S.-K.; Jayakumar, M.N.; Nagarkatti, M.; Nagarkatti, P.; Hassuneh, M.R. Induction of CD4(+)CD25(+) Regulatory T Cells from In Vitro Grown Human Mononuclear Cells by Sparteine Sulfate and Harpagoside. Biology 2020, 9, 211. [Google Scholar] [CrossRef]
- Zhao, L.; Zhou, X.; Zhou, X.; Wang, H.; Gu, L.; Ke, Y.; Zhang, M.; Ji, X.; Yang, X. Low expressions of PD-L1 and CTLA-4 by induced CD4(+)CD25(+) Foxp3(+) Tregs in patients with SLE and their correlation with the disease activity. Cytokine 2020, 133, 155119. [Google Scholar] [CrossRef]
- Bennett, C.L.; Christie, J.; Ramsdell, F.; Brunkow, M.E.; Ferguson, P.J.; Whitesell, L.; Kelly, T.E.; Saulsbury, F.T.; Chance, P.F.; Ochs, H.D. The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nat. Genet. 2001, 27, 20–21. [Google Scholar] [CrossRef]
- Agakidis, C.; Agakidou, E.; Sarafidis, K.; Papoulidis, I.; Xinias, I.; Farmaki, E. Immune Dysregulation, Polyendocrinopathy, Enteropathy, X-Linked Syndrome Associated With a Novel Mutation of FOXP3 Gene. Front. Pediatr. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Bennett, C.L.; Ochs, H.D. IPEX is a unique X-linked syndrome characterized by immune dysfunction, polyendocrinopathy, enteropathy, and a variety of autoimmune phenomena. Curr. Opin. Pediatr. 2001, 13, 533–538. [Google Scholar] [CrossRef]
- Vignali, D.A.A.; Collison, L.W.; Workman, C.J. How regulatory T cells work. Nat. Rev. Immunol. 2008, 8, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josefowicz, S.Z.; Lu, L.-F.; Rudensky, A.Y. Regulatory T cells: Mechanisms of differentiation and function. Annu Annu. Rev. Immunol. 2012, 30, 531–564. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xue, H.; Gao, T.; Gao, M.; Zhang, Y. Notch1 Signaling Regulates the Th17/Treg Immune Imbalance in Patients with Psoriasis Vulgaris. Mediat. Inflamm. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamehic, J.; Zecevic, L.; Resic, H.; Jukic, M.; Jukic, T.; Ridjic, O.; Panjeta, M.; Coric, J. Immunophenotype lymphocyte of peripheral blood in patients with psoriasis. Med Arch. 2014, 68, 236–238. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, Y.; Yang, X.; Wei, J.; Zhou, S.; Zhao, Z.; Cheng, J.; Duan, H.; Jia, T.; Lei, Q.; et al. Characterization of Th17 and FoxP3(+) Treg Cells in Paediatric Psoriasis Patients. Scand. J. Immunol. 2016, 83, 174–180. [Google Scholar] [CrossRef]
- Nussbaum, L.; Chen, Y.; Ogg, G.S. Role of regulatory T cells in psoriasis pathogenesis and treatment. Br. J. Dermatol. 2020. [Google Scholar] [CrossRef]
- Huan, J.; Culbertson, N.; Spencer, L.; Bartholomew, R.; Burrows, G.G.; Chou, Y.K.; Bourdette, D.; Ziegler, S.F.; Offner, H.; Vandenbark, A.A. Decreased FOXP3 levels in multiple sclerosis patients. J. Neurosci. Res. 2005, 81, 45–52. [Google Scholar] [CrossRef]
- Ciccocioppo, F.; Lanuti, P.; Pierdomenico, L.; Simeone, P.; Bologna, G.; Ercolino, E.; Buttari, F.; Fantozzi, R.; Thomas, A.; Onofrj, M.; et al. The Characterization of Regulatory T-Cell Profiles in Alzheimer's Disease and Multiple Sclerosis. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Sánchez, N.; Cruz-Chamorro, I.; Díaz-Sánchez, M.; Lardone, P.J.; Guerrero, J.M.; Carrillo-Vico, A. Peripheral CD39-expressing T regulatory cells are increased and associated with relapsing-remitting multiple sclerosis in relapsing patients. Sci. Rep. 2019, 9, 2302. [Google Scholar] [CrossRef]
- Antiga, E.; Quaglino, P.; Bellandi, S.; Volpi, W.; Del Bianco, E.; Comessatti, A.; Osella-Abate, S.; De Simone, C.; Marzano, A.; Bernengo, M.; et al. Regulatory T cells in the skin lesions and blood of patients with systemic sclerosis and morphoea. Br. J. Dermatol. 2010, 162, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gao, N.; Li, M.; Xu, D.; Hou, Y.; Wang, Q.; Zhang, G.; Sun, Q.; Zhang, H.; Zeng, X. Elevated levels of CD4(+)CD25(+)FoxP3(+) T cells in systemic sclerosis patients contribute to the secretion of IL-17 and immunosuppression dysfunction. PLoS ONE 2013, 8, e64531. [Google Scholar] [CrossRef] [PubMed]
- Miyara, M.; Amoura, Z.; Parizot, C.; Badoual, C.; Dorgham, K.; Trad, S.; Nochy, D.; Debré, P.; Piette, J.-C.; Gorochov, G. Global natural regulatory T cell depletion in active systemic lupus erythematosus. J. Immunol. 2005, 175, 8392–8400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, A.; Wittekind, P.S.; Kotschenreuther, K.; Schiller, J.; Tresckow, J.V.; Haak, T.H.; Kofler, D.M. Regulatory T cell frequencies in patients with rheumatoid arthritis are increased by conventional and biological DMARDs but not by JAK inhibitors. Ann. Rheum Dis. 2019, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Sun, X.; Zhao, J.; Zhang, J.; Zhu, H.; Li, C.; Gao, N.; Jia, Y.; Xu, D.; Huang, F.-P.; et al. Regulatory T cells in rheumatoid arthritis showed increased plasticity toward Th17 but retained suppressive function in peripheral blood. Ann. Rheum. Dis. 2014, 74, 1293–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viisanen, T.; Gazali, A.M.; Ihantola, E.-L.; Ekman, I.; Näntö-Salonen, K.; Veijola, R.; Toppari, J.; Knip, M.; Ilonen, J.; Kinnunen, T. FOXP3+ Regulatory T Cell Compartment Is Altered in Children With Newly Diagnosed Type 1 Diabetes but Not in Autoantibody-Positive at-Risk Children. Front. Immunol. 2019, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Lindley, S.; Dayan, C.M.; Bishop, A.; Roep, B.O.; Peakman, M.; Tree, T.I. Defective suppressor function in CD4(+)CD25(+) T-cells from patients with type 1 diabetes. Diabetes 2005, 54, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Putnam, A.L.; Vendrame, F.; Dotta, F.; Gottlieb, P.A. CD4+CD25high regulatory T cells in human autoimmune diabetes. J. Autoimmun. 2005, 24, 55–62. [Google Scholar] [CrossRef]
- Brusko, T.; Wasserfall, C.H.; McGrail, K.; Schatz, R.; Viener, H.L.; Schatz, D.A.; Haller, M.; Rockell, J.; Gottlieb, P.; Clare-Salzler, M.; et al. No alterations in the frequency of FOXP3+ regulatory T-cells in type 1 diabetes. Diabetes 2007, 56, 604–612. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, A.; Socci, C.; Stabilini, A.; Valle, A.; Monti, P.; Piemonti, L.; Nano, R.; Olek, S.; Maffi, P.; Scavini, M.; et al. Expansion of Th17 cells and functional defects in T regulatory cells are key features of the pancreatic lymph nodes in patients with type 1 diabetes. Diabetes 2011, 60, 2903–2913. [Google Scholar] [CrossRef] [Green Version]
- Okubo, Y.; Torrey, H.; Butterworth, J.; Zheng, H.; Faustman, D.L. Treg activation defect in type 1 diabetes: Correction with TNFR2 agonism. Clin. Transl. Immunol. 2016, 5, e56. [Google Scholar] [CrossRef] [PubMed]
- Hamari, S.; Kirveskoski, T.; Glumoff, V.; Kulmala, P.; Simell, O.; Knip, M.; Veijola, R. Analyses of regulatory CD4+ CD25+ FOXP3+ T cells and observations from peripheral T cell subpopulation markers during the development of type 1 diabetes in children. Scand. J. Immunol. 2016, 83, 279–287. [Google Scholar] [CrossRef] [PubMed]
- El Essawy, B.; Li, X.C. Type 1 diabetes and T regulatory cells. Pharmacol. Res. 2015, 98, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Gras, S.; Chadderton, J.; Del Campo, C.M.; Farenc, C.; Wiede, F.; Josephs, T.M.; Sng, X.Y.; Mirams, M.; Watson, K.A.; Tiganis, T.; et al. Reversed T Cell Receptor Docking on a Major Histocompatibility Class I Complex Limits Involvement in the Immune Response. Immunity 2016, 45, 749–760. [Google Scholar] [CrossRef] [Green Version]
- Stadinski, B.D.; Blevins, S.J.; Spidale, N.A.; Duke, B.R.; Huseby, P.G.; Stern, L.J.; Huseby, E.S. A temporal thymic selection switch and ligand binding kinetics constrain neonatal Foxp3(+) Treg cell development. Nat. Immunol. 2019, 20, 1046–1058. [Google Scholar] [CrossRef]
- Scherm, M.G.; Daniel, C. miRNA Regulation of T Cells in Islet Autoimmunity and Type 1 Diabetes. Curr. Diabetes Rep. 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Rosenzwajg, M.; Salet, R.; Lorenzon, R.; Tchitchek, N.; Roux, A.; Bernard, C.; Carel, J.-C.; Storey, C.; Polak, M.; Beltrand, J.; et al. Low-dose IL-2 in children with recently diagnosed type 1 diabetes: A Phase I/II randomised, double-blind, placebo-controlled, dose-finding study. Diabetology 2020, 63, 1808–1821. [Google Scholar] [CrossRef]
- Rosenzwajg, M.; Lorenzon, R.; Cacoub, P.; Pham, H.P.; Pitoiset, F.; El Soufi, K.; Bernard, C.; Aractingi, S.; Banneville, B.; Beaugerie, L.; et al. Immunological and clinical effects of low-dose interleukin-2 across 11 autoimmune diseases in a single, open clinical trial. Ann. Rheum Dis. 2019, 78, 209–217. [Google Scholar] [CrossRef]
- Tahvildari, M.; Dana, R. Low-Dose IL-2 Therapy in Transplantation, Autoimmunity, and Inflammatory Diseases. J. Immunol. 2019, 203, 2749–2755. [Google Scholar] [CrossRef]
- Hull, C.M.; Nickolay, L.E.; Estorninho, M.; Richardson, M.W.; Riley, J.L.; Peakman, M.; Maher, J.; Tree, T. Generation of human islet-specific regulatory T cells by TCR gene transfer. J. Autoimmun. 2017, 79, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Marek-Trzonkowska, N.; Myśliwiec, M.; Dobyszuk, A.; Grabowska, M.; Derkowska, I.; Juścińska, J.; Owczuk, R.; Szadkowska, A.; Witkowski, P.; Młynarski, W.; et al. Therapy of type 1 diabetes with CD4(+)CD25(high)CD127-regulatory T cells prolongs survival of pancreatic islets-results of one year follow-up. Clin. Immunol. 2014, 153, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Imura, Y.; Ando, M.; Kondo, T.; Ito, M.; Yoshimura, A. CD19-targeted CAR regulatory T cells suppress B cell pathology without GvHD. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Roth-Walter, F.; Adcock, I.M.; Benito-Villalvilla, C.; Bianchini, R.; Bjermer, L.; Boyman, O.; Caramori, G.; Cari, L.; Chung, K.F.; Diamant, Z.; et al. Immune modulation via T regulatory cell enhancement: Disease-modifying therapies for autoimmunity and their potential for chronic allergic and inflammatory diseases—An EAACI position paper of the Task Force on Immunopharmacology (TIPCO). Allergy. 2020. [Google Scholar] [CrossRef] [PubMed]
- Saleh, R.; Elkord, E. FoxP3(+) T regulatory cells in cancer: Prognostic biomarkers and therapeutic targets. Cancer Lett. 2020, 490, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Bates, G.J.; Fox, S.B.; Han, C.; Leek, R.D.; Garcia, J.F.; Harris, A.L.; Banham, A.H. Quantification of regulatory T cells enables the identification of high-risk breast cancer patients and those at risk of late relapse. J. Clin. Oncol. 2006, 24, 5373–5380. [Google Scholar] [CrossRef]
- Miller, A.M.; Lundberg, K.; Özenci, V.; Banham, A.H.; Hellström, M.; Egevad, L.; Pisa, P. CD4+CD25high T cells are enriched in the tumor and peripheral blood of prostate cancer patients. J. Immunol. 2006, 177, 7398–7405. [Google Scholar] [CrossRef] [Green Version]
- Perrone, G.; Ruffini, P.A.; Catalano, V.; Spino, C.; Santini, D.; Muretto, P.; Spoto, C.; Zingaretti, C.; Sisti, V.; Alessandroni, P.; et al. Intratumoural FOXP3-positive regulatory T cells are associated with adverse prognosis in radically resected gastric cancer. Eur. J. Cancer 2008, 44, 1875–1882. [Google Scholar] [CrossRef]
- O’Callaghan, D.S.; Rexhepaj, E.; Gately, K.; Coate, L.; Delaney, D.; O’Donnell, D.M.; Kay, E.; O’Connell, F.; Gallagher, W.M.; O’Byrne, K.J. Tumour islet Foxp3+ T-cell infiltration predicts poor outcome in nonsmall cell lung cancer. Eur. Respir. J. 2015, 46, 1762–1772. [Google Scholar] [CrossRef] [Green Version]
- Liotta, F.; Gacci, M.; Frosali, F.; Querci, V.; Vittori, G.; Lapini, A.; Santarlasci, V.; Serni, S.; Cosmi, L.; Maggi, L.; et al. Frequency of regulatory T cells in peripheral blood and in tumour-infiltrating lymphocytes correlates with poor prognosis in renal cell carcinoma. BJU Int. 2010, 107, 1500–1506. [Google Scholar] [CrossRef]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med. 2004, 10, 942–949. [Google Scholar] [CrossRef]
- Shah, W.; Yan, X.; Jing, L.; Zhou, Y.; Chen, H.; Wang, Y. A reversed CD4/CD8 ratio of tumor-infiltrating lymphocytes and a high percentage of CD4(+)FOXP3(+) regulatory T cells are significantly associated with clinical outcome in squamous cell carcinoma of the cervix. Cell. Mol. Immunol. 2010, 8, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitfeld, D.; Ohl, L.; Kremmer, E.; Ellwart, J.; Sallusto, F.; Lipp, M.; Förster, R. Follicular B helper T cells express CXC chemokine receptor 5, localize to B cell follicles, and support immunoglobulin production. J. Exp. Med. 2000, 192, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Rott, L.S.; Clark-Lewis, I.; Campbell, D.J.; Wu, L.; Butcher, E.C. Subspecialization of CXCR5+ T cells: B helper activity is focused in a germinal center-localized subset of CXCR5+ T cells. J. Exp. Med. 2001, 193, 1373–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaerli, P.; Willimann, K.; Lang, A.B.; Lipp, M.; Loetscher, P.; Moser, B. CXC chemokine receptor 5 expression defines follicular homing T cells with B cell helper function. J. Exp. Med. 2000, 192, 1553–1562. [Google Scholar] [CrossRef] [Green Version]
- Fazilleau, N.; Mark, L.; McHeyzer-Williams, L.J.; McHeyzer-Williams, M.G. Follicular helper T cells: Lineage and location. Immunity 2009, 30, 324–335. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Waseem, T.C.; Sahoo, A.; Bieerkehazhi, S.; Zhou, H.; Galkina, E.V.; Nurieva, R. Insights Into the Molecular Mechanisms of T Follicular Helper-Mediated Immunity and Pathology. Front. Immunol. 2018, 9, 1884. [Google Scholar] [CrossRef]
- Karnowski, A.; Chevrier, S.; Belz, G.T.; Mount, A.; Emslie, D.; D’Costa, K.; Tarlinton, D.M.; Kallies, A.; Corcoran, L.M. B and T cells collaborate in antiviral responses via IL-6, IL-21, and transcriptional activator and coactivator, Oct2 and OBF-1. J. Exp. Med. 2012, 209, 2049–2064. [Google Scholar] [CrossRef]
- Nurieva, R.I.; Chung, Y.; Hwang, D.; Yang, X.O.; Kang, H.S.; Ma, L.; Wang, Y.H.; Watowich, S.S.; Jetten, A.M.; Tian, Q.; et al. Generation of T follicular helper cells is mediated by interleukin-21 but independent of T helper 1, 2, or 17 cell lineages. Immunity 2008, 29, 138–149. [Google Scholar] [CrossRef] [Green Version]
- Nurieva, R.; Chung, Y.; Martinez, G.J.; Yang, X.O.; Tanaka, S.; Matskevitch, T.D.; Wang, Y.-H.; Dong, C. Bcl6 mediates the development of T follicular helper cells. Science 2009, 325, 1001–1005. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, N.; Liu, Y.; Bentebibel, S.E.; Munagala, I.; Bourdery, L.; Venuprasad, K.; Banchereau, J.; Ueno, H. The cytokine TGF-beta co-opts signaling via STAT3-STAT4 to promote the differentiation of human TFH cells. Nat. Immunol. 2014, 15, 856–865. [Google Scholar] [CrossRef] [Green Version]
- Locci, M.; E Wu, J.; Arumemi, F.; Mikulski, Z.; Dahlberg, C.; Miller, C.D.A.T.; Crotty, S. Activin A programs the differentiation of human TFH cells. Nat. Immunol. 2016, 17, 976–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballesteros-Tato, A.; Leon, B.; Graf, B.A.; Moquin, A.; Adams, P.S.; Lund, F.E.; Randall, T.D. Interleukin-2 inhibits germinal center formation by limiting T follicular helper cell differentiation. Immunity 2012, 36, 847–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiToro, D.; Winstead, C.J.; Pham, D.; Witte, S.; Andargachew, R.; Singer, J.R.; Wilson, C.G.; Zindl, C.L.; Luther, R.J.; Silberger, D.J.; et al. Differential IL-2 expression defines developmental fates of follicular versus nonfollicular helper T cells. Science 2018, 361, eaao2933. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S. T Follicular Helper Cell Biology: A Decade of Discovery and Diseases. Immunity 2019, 50, 1132–1148. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.L.; Axtell, R.C. Emerging Role of Follicular T Helper Cells in Multiple Sclerosis and Experimental Autoimmune Encephalomyelitis. Int. J. Mol. Sci. 2018, 19, 3233. [Google Scholar] [CrossRef] [Green Version]
- Gensous, N.; Charrier, M.; Duluc, D.; Contin-Bordes, C.; Truchetet, M.-E.; Lazaro, E.; Duffau, P.; Blanco, P.; Richez, C. T Follicular Helper Cells in Autoimmune Disorders. Front. Immunol. 2018, 9, 1637. [Google Scholar] [CrossRef]
- Serr, I.; Furst, R.W.; Ott, V.B.; Scherm, M.G.; Nikolaev, A.; Gokmen, F.; Kalin, S.; Zillmer, S.; Bunk, M.; Weigmann, B.; et al. miRNA92a targets KLF2 and the phosphatase PTEN signaling to promote human T follicular helper precursors in T1D islet autoimmunity. Proc. Natl. Acad. Sci. USA 2016, 113, E6659–E6668. [Google Scholar] [CrossRef] [Green Version]
- Viisanen, T.; Ihantola, E.-L.; Näntö-Salonen, K.; Hyöty, H.; Nurminen, N.; Selvenius, J.; Juutilainen, A.; Moilanen, L.; Pihlajamäki, J.; Veijola, R.; et al. Circulating CXCR5+PD-1+ICOS+ Follicular T Helper Cells Are Increased Close to the Diagnosis of Type 1 Diabetes in Children With Multiple Autoantibodies. Diabetes 2016, 66, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Corsiero, E.; Nerviani, A.; Bombardieri, M.; Pitzalis, C. Ectopic Lymphoid Structures: Powerhouse of Autoimmunity. Front. Immunol. 2016, 7, 430. [Google Scholar] [CrossRef] [Green Version]
- Smolen, J.S.; Aletaha, D.; Barton, A.; Burmester, G.R.; Emery, P.; Firestein, G.S.; Kavanaugh, A.; McInnes, I.B.; Solomon, D.H.; Strand, V.; et al. Rheumatoid arthritis. Nat. Rev. Dis. Primers 2018, 4, 18001. [Google Scholar] [CrossRef]
- Szabo, K.; Papp, G.; Barath, S.; Gyimesi, E.; Szanto, A.; Zeher, M. Follicular helper T cells may play an important role in the severity of primary Sjogren′s syndrome. Clin. Immunol. 2013, 147, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.-Y.; Seth, A.; Kashgarian, M.; Terrillon, S.; Fung, E.; Huang, L.; Wang, L.C.; Craft, J. Disruption of Pathogenic Cellular Networks by IL-21 Blockade Leads to Disease Amelioration in Murine Lupus. J. Immunol. 2017, 198, 2578–2588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Lee, K.; Diamond, B. Follicular Helper T Cells in Systemic Lupus Erythematosus. Front. Immunol. 2018, 9, 1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dan, J.M.; Havenar-Daughton, C.; Kendric, K.; Al-Kolla, R.; Kaushik, K.; Rosales, S.L.; Anderson, E.L.; LaRock, C.N.; Vijayanand, P.; Seumois, G.; et al. Recurrent group A Streptococcus tonsillitis is an immunosusceptibility disease involving antibody deficiency and aberrant TFH cells. Sci. Transl. Med. 2019, 11, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banga, R.; Procopio, F.A.; Noto, A.; Pollakis, G.; Cavassini, M.; Ohmiti, K.; Corpataux, J.-M.; De Leval, L.; Pantaleo, G.; Perreau, M. PD-1(+) and follicular helper T cells are responsible for persistent HIV-1 transcription in treated aviremic individuals. Nat. Med. 2016, 22, 754–761. [Google Scholar] [CrossRef]
- Moody, M.A.; Pedroza-Pacheco, I.; Vandergrift, N.A.; Chui, C.; Lloyd, K.E.; Parks, R.; Soderberg, K.A.; Ogbe, A.T.; Cohen, M.S.; Liao, H.-X.; et al. Immune perturbations in HIV-1-infected individuals who make broadly neutralizing antibodies. Sci. Immunol. 2016, 1, aag0851. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Lynch, R.M.; Gautam, R.; Matus-Nicodemos, R.; Schmidt, S.D.; Boswell, K.L.; Darko, S.; Wong, P.; Sheng, Z.; Petrovas, C.; et al. Quality and quantity of TFH cells are critical for broad antibody development in SHIVAD8 infection. Sci. Transl. Med. 2015, 7, 298ra120. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Tosolini, M.; Kirilovsky, A.; Waldner, M.; Obenauf, A.C.; Angell, H.; Fredriksen, T.; Lafontaine, L.; Berger, A.; et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity 2013, 39, 782–795. [Google Scholar] [CrossRef] [Green Version]
- Gu-Trantien, C.; Migliori, E.; Buisseret, L.; De Wind, R.; Brohée, S.; Garaud, S.; Noël, G.; Dang Chi, V.L.; Lodewyckx, J.N.J.; Naveaux, C.; et al. CXCL13-producing TFH cells link immune suppression and adaptive memory in human breast cancer. JCI Insight 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Havenar-Daughton, C.; Lindqvist, M.; Heit, A.; Wu, J.E.; Reiss, S.M.; Kendric, K.; Bélanger, S.; Kasturi, S.P.; Landais, E.; Akondy, R.S.; et al. CXCL13 is a plasma biomarker of germinal center activity. Proc. Natl. Acad. Sci. USA 2016, 113, 2702–2707. [Google Scholar] [CrossRef] [Green Version]
- Garaud, S.; Zayakin, P.; Buisseret, L.; Rulle, U.; Siliņa, K.; De Wind, A.; Eyden, G.V.D.; Larsimont, D.; Willard-Gallo, K.; Linē, A. Antigen Specificity and Clinical Significance of IgG and IgA Autoantibodies Produced in situ by Tumor-Infiltrating B Cells in Breast Cancer. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
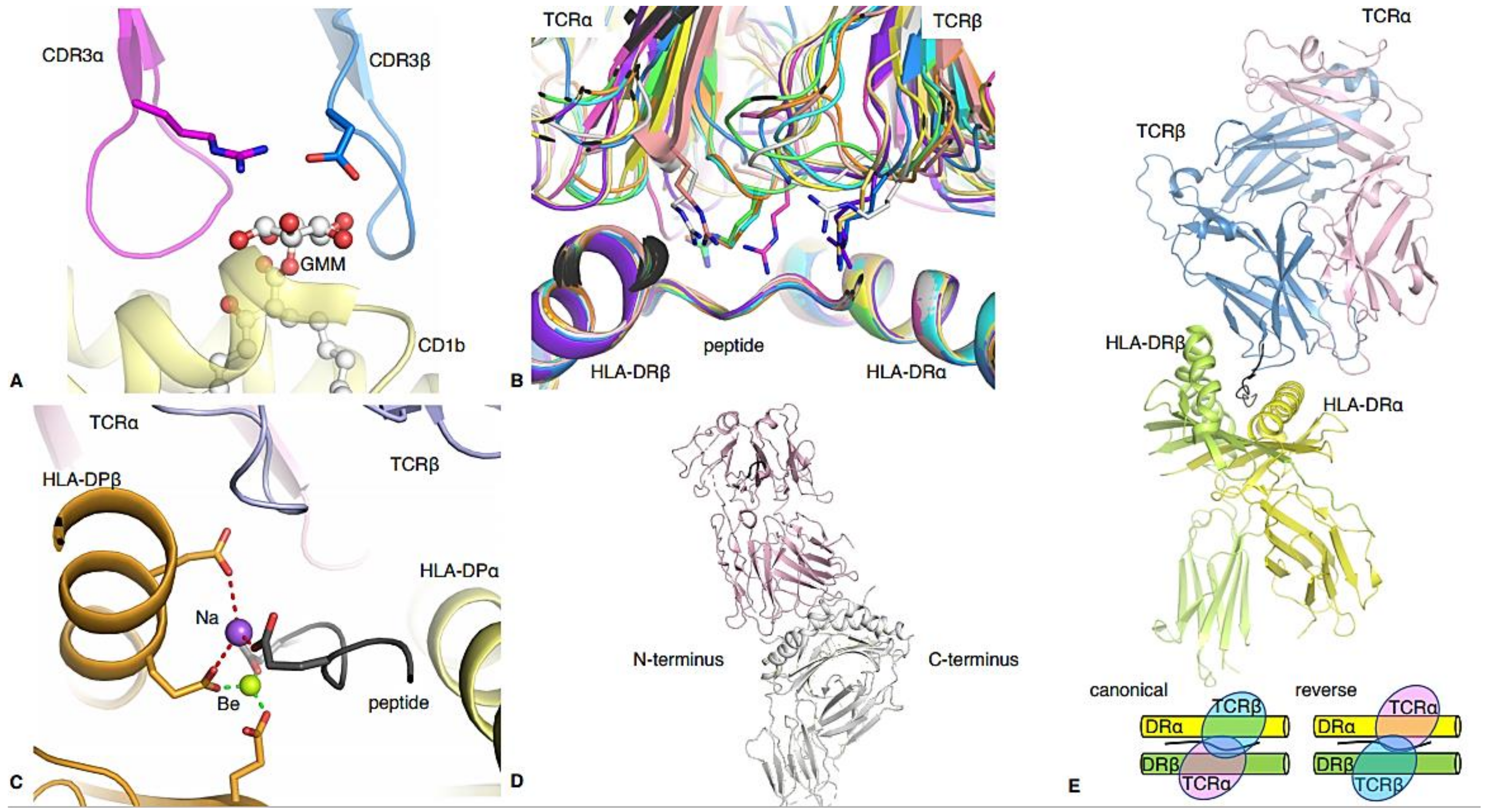
| HLA or CD1 | Peptide Sequence | TCR | TRAV | TRBV | Kd (μM) | Role in Disease | PBD | Reference |
|---|---|---|---|---|---|---|---|---|
| HLA-DR1 | PKYVKQNTLKLAT | F11 | 8-4*01 | 24-1*01 | 26.7 | Antiviral | 6R0E | [15] |
| HLA-DR1 | PKYVKQNTLKLAT | HA1.7 | 8-4*01 | 28*01 | 37 | Antiviral | 1FYT | [16] |
| HLA-DR1 | RFYKTLRAEQAS | F24 | 24*01 | 2*01 | 10.56 | Antiviral | 6CQR | [10] |
| HLA-DR11 | RFYKTLRAEQAS | F24 | 24*01 | 2*01 | 1.16 | Antiviral | 6CQL | [10] |
| HLA-DR15 | RFYKTLRAEQAS | F24 | 24*01 | 2*01 | 6.9 | Antiviral | 6CQQ | [10] |
| CD1b | GMM-C32 | GEM42 | 1-2*01 | 6-2*01 | 0.85 | Anti-bacterial | 5L2K | [17] |
| HLA-DQ8 | VEELYLVAGEEGC | T1D-3 | 17*01 | 5-1*01 | 21.4 | Diabetes | 6DFX | [18] |
| HLA-DQ2 | PQPELPYPQ | D2 | 26-1*01 | 7-2*01 | 15.8 | Coeliac disease | 4OZH | [19] |
| HLA-DQ2 | PQPELPYPQ | JR5.1 | 26-1*01 | 7-2*01 | 79.4 | Coeliac disease | 4OZG | [19] |
| HLA-DQ2 | PQPELPYPQ | S16 | 26-1*01 | 7-2*01 | 24.8 | Coeliac disease | 4OZF | [19] |
| HLA-DQ2 | PFPQPELPY | S2 | 4*01 | 20-1*01 | 70 | Coeliac disease | 4OZI | [19] |
| HLA-DQ2.2 | PFSEQEQPV | T1005.2.56 | 21*01 | 7-3*01 | 22.1 | Coeliac disease | 6PX6 | [20] |
| HLA-DQ2.2 | PFSEQEQPV | T594 | 9-2*01 | 11-2*01 | 20.9 | Coeliac disease | 6PY2 | [20] |
| HLA-DQ8 | SGEGSFQPSQENP | S13 | 26-2*01 | 9*01 | 1.05 | Coeliac disease | 4Z7U | [21] |
| HLA-DQ8 | SGEGSFQPSQENP | L3-12 | 26-2*01 | 9*01 | 7 | Coeliac disease | 4Z7V | [21] |
| HLA-DQ8 | SGEGSFQPSQENP | T316 | 8-3*01 | 6-1*01 | 2.1 | Coeliac disease | 4Z7W | [21] |
| HLA-DQ8 | SGEGSFQPSQENP | Bel502 | 20*01 | 9*01 | 2.8 | Coeliac disease | 5KS9 | [22] |
| HLA-DQ8 | QPQQSFPEQEA | Bel602 | 20*01 | 9*01 | 4.7 | Coeliac disease | 5KSA | [22] |
| HLA-DQ8 | SGEGSFQPSQENP | SP3.4 | 26-2*01 | 9*01 | 11.4 | Coeliac disease | 4GG6 | [23] |
| HLA-DQ8.5 | GPQQSFPEQEA | T15 | 20*01 | 9*01 | 2 | Coeliac disease | 5KSB | [22] |
| HLA-DR1 | GELIGILNAAKVPAD | G4 | 22*01 | 5-8*01 | low $ | Cancer | 4E41 | [24] |
| HLA-DR1 | GELIGILNAAKVPAD | E8 | 22*01 | 6-6*01 | low $ | Cancer | 2IAM | [25] |
| HLA-DR3 # | QHIRCNIPKRISA | ANi2.3 | 8-3*01 | 19*01 | 38.3 | Allergy | 4H1L | [26] |
| HLA-DP2 | QAFWIDLFETIG | AV22 | 9-2*01 | 5-1*01 | 6 | Allergy | 4P4K | [27] |
| HLA-DR2 | ENPVVHFFKNIVTP | OB.1A12 | 17*01 | 20-1*01 | 47 | Multiple sclerosis | 1YMM | [28] |
| HLA-DR2a | VHFFKNIVTPRTPGG | 3A6 | 9-2*02 | 5-1*01 | low $ | Multiple sclerosis | 1ZGL | [29] |
| HLA-DR2b | DFARVHFISALHGSG | OB.1A12 | 17*01 | 20-1*01 | 300 | Anti-bacterial | 2WBJ | [30] |
| HLA-DR4 | FSWGAEGQRPGFG | MS2-3C8 | 26-2*01 | 20-1*01 | 5.5 | Multiple sclerosis | 3O6F | [31] |
| HLA-DQ1 | ENPVVHFFKNIVTPR | Hy.1B11 | 13-1*02 | 7-3*01 | 14.3 | Multiple sclerosis | 3PL6 | [32] |
| HLA-DQ2 | APMPMPELPYP | LS2.8/3.15 | 8-3*01 | 5-5*01 | 39.6 | Anti-bacterial | 6U3N | [33] |
| HLA-DQ2 | AVVQSELPYPEGS | JR5.1 | 26-1*01 | 7-2*01 | 132 | Anti-bacterial | 6U3O | [33] |
| HLA-DR4 | GSLQPLALEGSLQKRGIV | FS18 | 29/DV5*01 | 6-2*01 | low $ | Treg | 4Y19 | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chatzileontiadou, D.S.M.; Sloane, H.; Nguyen, A.T.; Gras, S.; Grant, E.J. The Many Faces of CD4+ T Cells: Immunological and Structural Characteristics. Int. J. Mol. Sci. 2021, 22, 73. https://doi.org/10.3390/ijms22010073
Chatzileontiadou DSM, Sloane H, Nguyen AT, Gras S, Grant EJ. The Many Faces of CD4+ T Cells: Immunological and Structural Characteristics. International Journal of Molecular Sciences. 2021; 22(1):73. https://doi.org/10.3390/ijms22010073
Chicago/Turabian StyleChatzileontiadou, Demetra S. M., Hannah Sloane, Andrea T. Nguyen, Stephanie Gras, and Emma J. Grant. 2021. "The Many Faces of CD4+ T Cells: Immunological and Structural Characteristics" International Journal of Molecular Sciences 22, no. 1: 73. https://doi.org/10.3390/ijms22010073
APA StyleChatzileontiadou, D. S. M., Sloane, H., Nguyen, A. T., Gras, S., & Grant, E. J. (2021). The Many Faces of CD4+ T Cells: Immunological and Structural Characteristics. International Journal of Molecular Sciences, 22(1), 73. https://doi.org/10.3390/ijms22010073