
Molecular and Functional Characterization of MobK Protein—A Novel-Type Relaxase Involved in Mobilization for Conjugational Transfer of Klebsiella pneumoniae Plasmid pIGRK

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
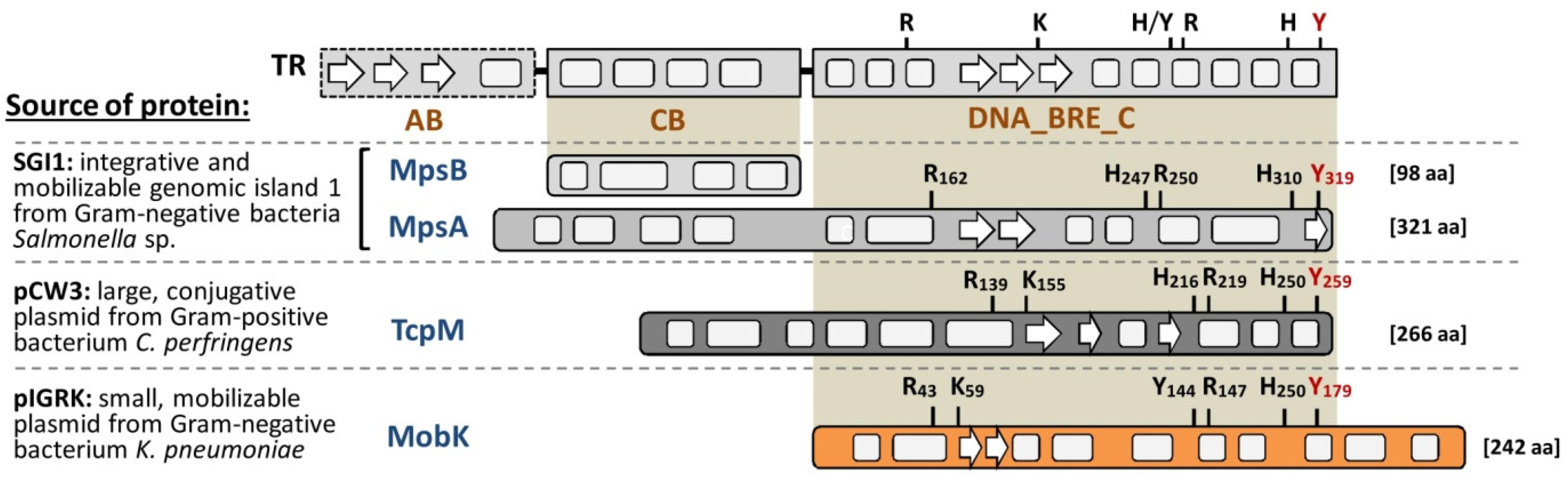
2.1. Comparative Analysis of Amino Acid Sequences of MobK and Other TR—Like Relaxases
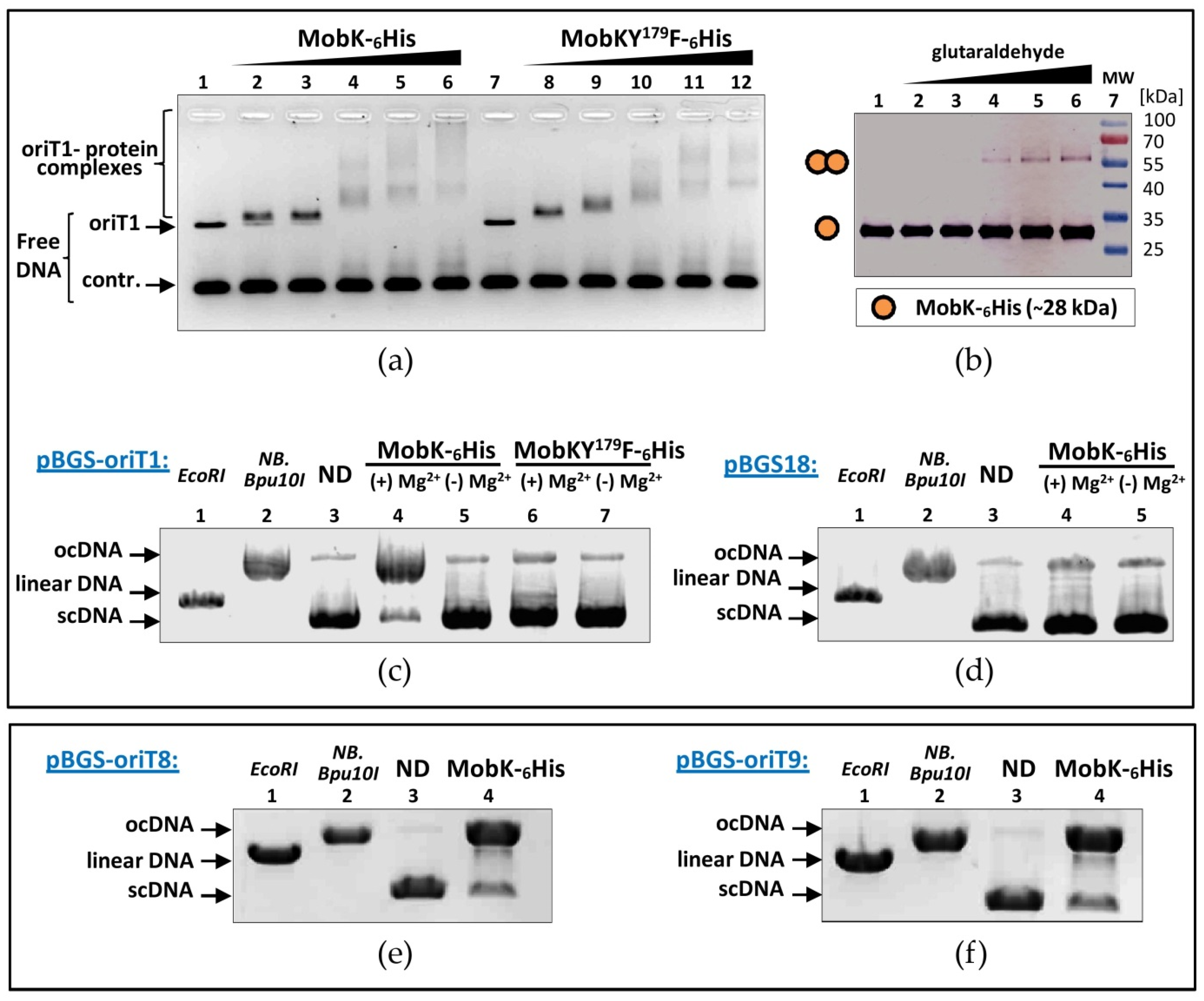
2.2. Tyrosine Residue (Y179) Is Crucial for the MobK Relaxase Activity
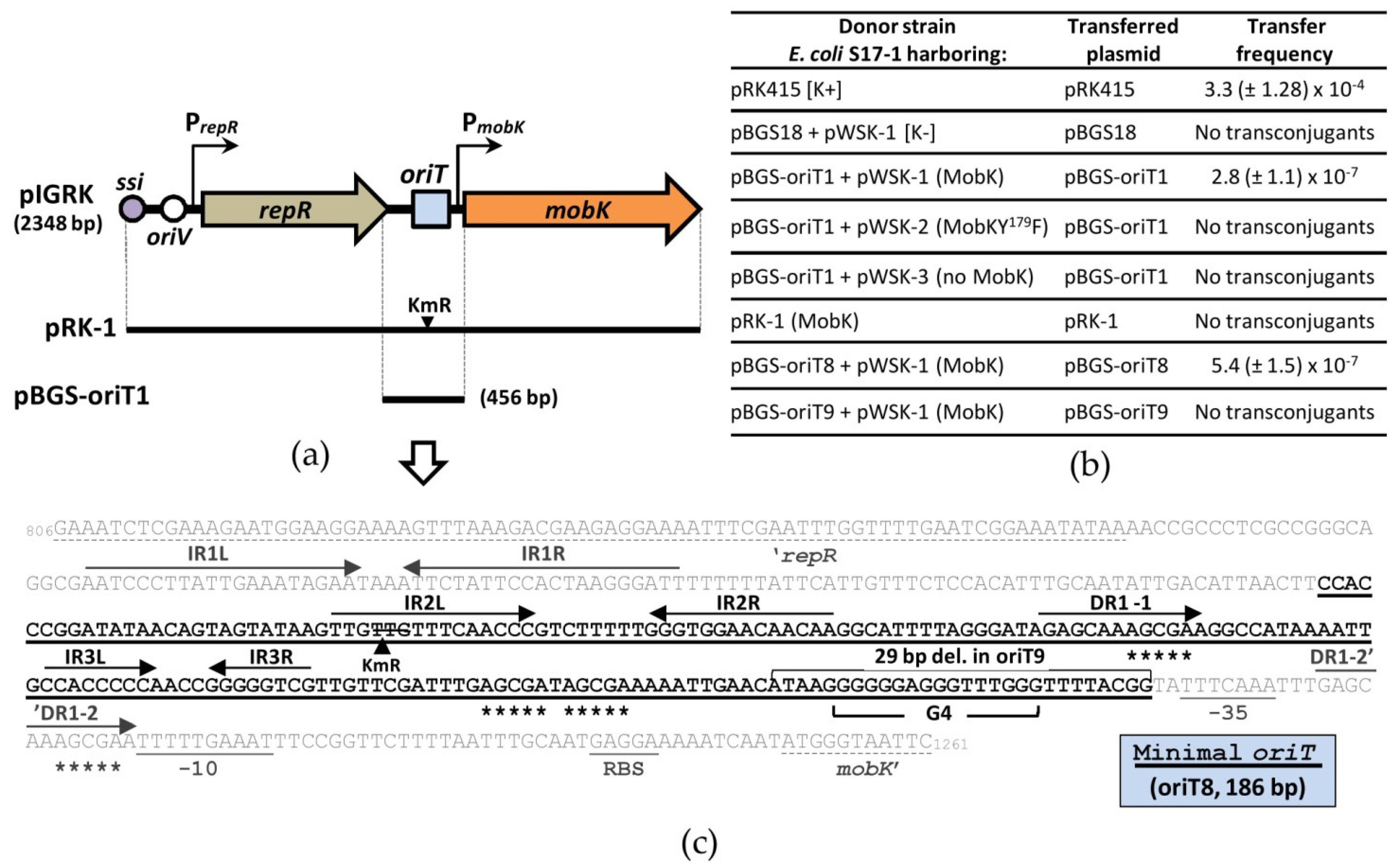
2.3. MobK Interacts Specifically with oriT1 Region of pIGRK In Vitro
2.4. MobK Forms Homodimers
2.5. MobKY179 Residue and Mg2+ Cations Are Critical for MobK Site-Specific Nuclease Activity
2.6. Mapping of pIGRK Minimal oriT Sequence
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids and Culture Conditions
4.2. DNA Manipulations
4.3. Western Blot Analysis
4.4. Overexpression and Purification of 6His-Tagged MobK and MobKY179F Proteins
4.5. Electrophoretic Mobility Shift Assay
4.6. Bacterial Mating Procedure
4.7. In Vitro MobK Nuclease Activity Assay
4.8. Glutaraldehyde Cross-Linking
4.9. Bioinformatics Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, J.P.J.; Brockhurst, M.A.; Harrison, E.; Philos Trans, R.; Soc Lond, B. Sampling the mobile gene pool: Innovation via horizontal gene transfer in bacteria. Biol. Sci. 2017, 372, 20160424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.; Nielsen, K. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Wawrzyniak, P.; Płucienniczak, G.; Bartosik, D. The different faces of rolling-circle replication and its multifunctional initiator proteins. Front. Microbiol. 2017, 8, 2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamczuk, M.; Zaleski, P.; Dziewit, L.; Wolinowska, R.; Nieckarz, M.; Wawrzyniak, P.; Kieryl, P.; Plucienniczak, A.; Bartosik, D. Diversity and global distribution of IncL/M plasmids enabling horizontal dissemination of β-lactam resistance genes among the Enterobacteriaceae. BioMed Res. Intern. 2015, 2015, 414681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, N.; Connery, S.; Dujeancourt, A.; Lu, F.; Redzej, A.; Fronzes, R.; Orlova, E.V.; Waksman, G. Structure of a type IV secretion system. Nature 2014, 508, 550–553. [Google Scholar] [CrossRef]
- Alderliesten, J.B.; Duxbury, S.J.N.; Zwart, M.P.; de Visser, J.A.G.M.; Stegeman, A.; Fischer, E.A.J. Effect of donor-recipient relatedness on the plasmid conjugation frequency: A meta-analysis. BMC Microbiol. 2020, 20, 135. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef]
- Cabezón, E.; Ripoll-Rozada, J.; Peña, A.; de la Cruz, F.; Arechaga, I. Towards an integrated model of bacterial conjugation. FEMS Microbiol. Rev. 2015, 39, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Waksman, G. From conjugation to T4S systems in Gram-negative bacteria: A mechanistic biology perspective. EMBO Rep. 2019, 20, e47012. [Google Scholar] [CrossRef] [PubMed]
- Dmowski, M.; Gołębiewski, M.; Kern-Zdanowicz, I. Characteristics of the conjugative transfer system of the IncM plasmid pCTX-M3 and identification of its putative regulators. J. Bacteriol. 2018, 200, e00234–e00318. [Google Scholar] [CrossRef] [Green Version]
- Dmowski, M.; Kern-Zdanowicz, I. A novel mobilizing tool based on the conjugative transfer system of the IncM plasmid pCTX-M3. Appl. Environ. Microbiol. 2020, 86, e01205-20. [Google Scholar] [CrossRef] [PubMed]
- Getino, M.; Sanabria-Ríos, D.J.; Fernández-López, R.; Campos-Gómez, J.; Sánchez-López, J.M.; Fernández, A.; Carballeira, N.M.; de la Cruz, F. Synthetic fatty acids prevent plasmid-mediated horizontal gene transfer. mBio. 2015, 6, e01032-15. [Google Scholar] [CrossRef] [Green Version]
- Cabezón, E.; de la Cruz, F.; Arechaga, I.; Cabezón, E.; de la Cruz, F.; Arechaga, I. Conjugation inhibitors and their potential use to prevent dissemination of antibiotic resistance genes in bacteria. Front. Microbiol. 2017, 8, 2329. [Google Scholar] [CrossRef] [Green Version]
- Boudaher, E.; Shaffer, C.L. Inhibiting bacterial secretion systems in the fight against antibiotic resistance. Medchemcomm 2019, 10, 682–692. [Google Scholar] [CrossRef]
- Ripoll-Rozada, J.; García-Cazorla, Y.; Getino, M.; Machón, C.; Sanabria-Ríos, D.; de la Cruz, F.; Cabezón, E.; Arechaga, I. Type IV traffic ATPase TrwD as molecular target to inhibit bacterial conjugation. Mol. Microbiol. 2016, 100, 912–921. [Google Scholar] [CrossRef]
- García-Cazorla, Y.; Getino, M.; Sanabria-Ríos, D.J.; Carballeira, N.M.; de la Cruz, F.; Arechaga, I.; Cabezón, E. Conjugation inhibitors compete with palmitic acid for binding to the conjugative traffic ATPase TrwD, providing a mechanism to inhibit bacterial conjugation. J. Biol. Chem. 2018, 293, 16923–16930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez-Rodríguez, I.; Arana, L.; Ugarte-Uribe, B.; Gómez-Rubio, E.; Martín-Santamaría, S.; Garbisu, C.; Alkorta, I. Type IV coupling proteins as potential targets to control the dissemination of antibiotic resistance. Front. Mol. Biosci. 2020, 7, 201. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Herrador, D.L.; Llosa, M. The secret life of conjugative relaxases. Plasmid. 2019, 104, 102415. [Google Scholar] [CrossRef] [PubMed]
- Garcillan-Barcia, M.P.; Francia, M.V.; de la Cruz, F. The diversity of conjugative relaxases and its application in plasmid classification. FEMS Microbiol. Rev. 2009, 33, 657–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, M.; de la Cruz, F.; Dyda, F.; Hickman, A.B.; Moncalian, G.; Ton-Hoang, B. Breaking and joining single-stranded DNA: The HUH endonuclease superfamily. Nat. Rev. Microbiol. 2013, 11, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Francia, M.V.; Clewell, D.B.; de la Cruz, F.; Moncalián, G. Catalytic domain of plasmid pAD1 relaxase TraX defines a group of relaxases related to restriction endonucleases. Proc. Natl. Acad. Sci. USA. 2013, 110, 13606–13611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, S.B.; Phillips, S.E.; Thomas, C.D. Structures of replication initiation proteins from staphylococcal antibiotic resistance plasmids reveal protein asymmetry and flexibility are necessary for replication. Nucleic Acids Res. 2016, 44, 2417–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godziszewska, J.; Moncalián, G.; Cabezas, M.; Bartosik, A.A.; de la Cruz, F.; Jagura-Burdzy, G. Concerted action of NIC relaxase and auxiliary protein MobC in RA3 plasmid conjugation. Mol. Microbiol. 2016, 101, 439–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisniewski, J.A.; Traore, D.A.; Bannam, T.L.; Lyras, D.; Whisstock, J.C.; Rood, J.I. TcpM: A novel relaxase that mediates transfer of large conjugative plasmids from Clostridium perfringens. Mol. Microbiol. 2016, 99, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, J.A.; Rood, J.I. The Tcp conjugation system of Clostridium perfringens. Plasmid 2017, 91, 28–36. [Google Scholar] [CrossRef]
- Kiss, J.; Szabó, M.; Hegyi, A.; Douard, G.; Praud, K.; Nagy, I.; Olasz, F.; Cloeckaert, A.; Doublet, B. Identification and characterization of oriT and two mobilization genes required for conjugative transfer of Salmonella genomic island 1. Front. Microbiol. 2019, 10, 457. [Google Scholar] [CrossRef] [Green Version]
- Smorawińska, M.; Szuplewska, M.; Zaleski, P.; Wawrzyniak, P.; Maj, A.; Płucienniczak, A.; Bartosik, D. Mobilizable narrow host range plasmids as natural suicide vectors enabling horizontal gene transfer among distantly related bacterial species. FEMS Microbiol. Lett. 2012, 326, 76–82. [Google Scholar] [CrossRef]
- Meinke, G.; Bohm, A.; Hauber, J.; Pisabarro, M.T.; Buchholz, F. Cre recombinase and other tyrosine recombinases. Chem. Rev. 2016, 116, 12785–12820. [Google Scholar] [CrossRef]
- Smyshlyaev, G.; Barabas, O.; Bateman, A. Sequence analysis allows functional annotation of tyrosine recombinases in prokaryotic genomes. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Drozdetskiy, A.; Cole, C.; Procter, J.; Barton, G.J. JPred4: A protein secondary structure prediction server. Nucleic Acids Res. 2015, 43, 389–394. [Google Scholar] [CrossRef]
- Goodwin, T.J.; Butler, M.I.; Poulter, R.T. Cryptons: A group of tyrosine-recombinase-encoding DNA transposons from pathogenic fungi. Microbiology 2003, 149, 3099–3109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wawrzyniak, P.; Sobolewska-Ruta, A.; Zaleski, P.; Łukasiewicz, N.; Kabaj, P.; Kierył, P.; Gościk, A.; Bierczyńska-Krzysik, A.; Baran, P.; Mazurkiewicz-Pisarek, A.; et al. Molecular dissection of the replication system of plasmid pIGRK encoding two in-frame Rep proteins with antagonistic functions. BMC Microbiol. 2019, 19, 254. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Tawani, A.; Mishra, A.; Kumar, A. G4IPDB. A database for G-quadruplex structure forming nucleic acid interacting proteins. Sci. Rep. 2016, 6, 38144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastrana, C.L.; Carrasco, C.; Akhtar, P.; Leuba, S.H.; Khan, S.A.; Moreno-Herrero, F. Force and twist dependence of RepC nicking activity on torsionally-constrained DNA molecules. Nucleic Acids Res. 2016, 44, 8885–8896. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Ruiz-Cruz, S.; Espinosa, M.; Zhang, J.R.; Bravo, A. In vitro DNA inversions mediated by the PsrA site-specific tyrosine recombinase of Streptococcus pneumoniae. Front. Mol. Biosci. 2020, 7, 43. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, P.; Kim, N.; Kumari, M.; Verma, S.; Sharma, T.K.; Yadav, V.; Kumar, A. G-quadruplex structures in bacteria - biological relevance and potential as antimicrobial target. J. Bacteriol. 2021, 1, 00577–005820. [Google Scholar] [CrossRef]
- Matilla, I.; Alfonso, C.; Rivas, G.; Bolt, E.L.; de la Cruz, F.; Cabezon, E. The conjugative DNA translocase TrwB is a structure-specific DNA binding protein. J. Biol. Chem. 2010, 285, 17537–17544. [Google Scholar] [CrossRef] [Green Version]
- Rozhon, W.; Khan, M.; Poppenberger, B. Identification of the region required for maintaining pHW126 in its monomeric form. FEMS Microbiol. Lett. 2012, 2, 89–96. [Google Scholar] [CrossRef]
- Thomas, J.; Hecht, D.W. Interaction of Bacteroides fragilis pLV22a relaxase and transfer DNA with Escherichia coli RP4-TraG coupling protein. Mol. Microbiol. 2007, 66, 948–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szpirer, C.Y.; Faelen, M.; Couturier, M. Interaction between the RP4 coupling protein TraG and the pBHR1 mobilization protein Mob. Mol. Microbiol. 2000, 37, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Moncalián, G.; Cabezón, E.; Alkorta, I.; Valle, M.; Moro, F.; Valpuesta, J.M.; Goñi, F.M.; de la Cruz, F. Characterization of ATP and DNA binding activities of TrwB, the coupling protein essential in plasmid R388 conjugation. J. Biol. Chem. 1999, 274, 36117–36124. [Google Scholar] [CrossRef] [Green Version]
- Abajy, M.Y.; Kope’c, J.; Schiwon, K.; Burzynski, M.; Döring, M.; Bohn, C.; Grohmann, E. A type IV-secretion-like system is required for conjugative DNA transport of broad-host-range plasmid pIP501 in gram-positive bacteria. J. Bacteriol. 2007, 189, 2487–2496. [Google Scholar] [CrossRef] [Green Version]
- Amado, E.; Muth, G.; Arechaga, I.; Cabezón, E. The FtsK-like motor TraB is a DNA-dependent ATPase that forms higher-order assemblies. J. Biol. Chem. 2019, 294, 5050–5059. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NJ, USA, 2001. [Google Scholar]
- Wulfing, C.; Lombardero, J.; Pluckthun, A. An Escherichia coli protein consisting of a domain homologous to FK506-binding proteins (FKBP) and a new metal binding motif. J. Biol. Chem. 1994, 269, 2895–2901. [Google Scholar] [CrossRef]
- Simon, R.; Preifer, U.; Puhler, A. A broad host range mobilization system for in vivo genetic engineering: Transposon mutagenesis in gram negative bacteria. Biotechnology 1983, 9, 184–191. [Google Scholar] [CrossRef]
- Bartosik, D.; Baj, J.; Sochacka, M.; Piechucka, E.; Wlodarczyk, M. Molecular characterization of functional modules of plasmid pWKS1 of Paracoccus pantotrophus DSM 11072. Microbiology 2002, 148, 2847–2856. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.F.; Kushner, S.R. Construction of versatile low-copy-number vectors for cloning, sequencing and gene expression in Escherichia coli. Gene 1991, 100, 195–199. [Google Scholar] [CrossRef]
- Keen, N.T.; Tamaki, S.; Kobayashi, D.; Trollinger, D. Improved broad-host-range plasmids for DNA cloning in Gram-negative bacteria. Gene 1988, 70, 191–197. [Google Scholar] [CrossRef]
- Spratt, B.G.; Hedge, P.J.; te Heesen, S.; Edelman, A.; Broome-Smith, J.K. Kanamycin-resistant vectors that are analogues of plasmids pUC8, pUC9, pEMBL8 and pEMBL9. Gene 1986, 41, 337–342. [Google Scholar] [CrossRef]
- Rozhon, W.; Khan, M.; Petutschnig, E.; Poppenberger, B. Identification of cis- and trans-acting elements in pHW126, a representative of a novel group of rolling circle plasmids. Plasmid 2011, 65, 70–76. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowak, K.P.; Sobolewska-Ruta, A.; Jagiełło, A.; Bierczyńska-Krzysik, A.; Kierył, P.; Wawrzyniak, P. Molecular and Functional Characterization of MobK Protein—A Novel-Type Relaxase Involved in Mobilization for Conjugational Transfer of Klebsiella pneumoniae Plasmid pIGRK. Int. J. Mol. Sci. 2021, 22, 5152. https://doi.org/10.3390/ijms22105152
Nowak KP, Sobolewska-Ruta A, Jagiełło A, Bierczyńska-Krzysik A, Kierył P, Wawrzyniak P. Molecular and Functional Characterization of MobK Protein—A Novel-Type Relaxase Involved in Mobilization for Conjugational Transfer of Klebsiella pneumoniae Plasmid pIGRK. International Journal of Molecular Sciences. 2021; 22(10):5152. https://doi.org/10.3390/ijms22105152
Chicago/Turabian StyleNowak, Katarzyna Paulina, Agnieszka Sobolewska-Ruta, Agata Jagiełło, Anna Bierczyńska-Krzysik, Piotr Kierył, and Paweł Wawrzyniak. 2021. "Molecular and Functional Characterization of MobK Protein—A Novel-Type Relaxase Involved in Mobilization for Conjugational Transfer of Klebsiella pneumoniae Plasmid pIGRK" International Journal of Molecular Sciences 22, no. 10: 5152. https://doi.org/10.3390/ijms22105152
APA StyleNowak, K. P., Sobolewska-Ruta, A., Jagiełło, A., Bierczyńska-Krzysik, A., Kierył, P., & Wawrzyniak, P. (2021). Molecular and Functional Characterization of MobK Protein—A Novel-Type Relaxase Involved in Mobilization for Conjugational Transfer of Klebsiella pneumoniae Plasmid pIGRK. International Journal of Molecular Sciences, 22(10), 5152. https://doi.org/10.3390/ijms22105152