
Effects of Androgen Receptor Inhibition on Kanamycin-Induced Hearing Loss in Rats


Abstract
:1. Introduction
2. Results
3. Discussion
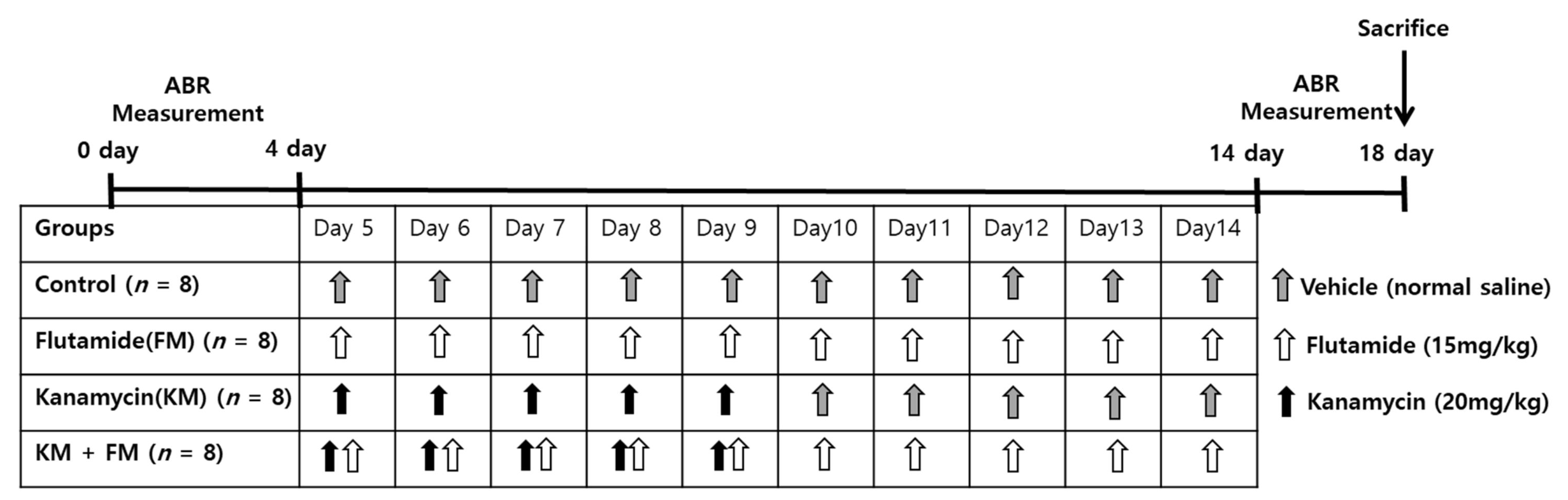
4. Materials and Methods
4.1. Auditory Function Tests
4.2. Cochlear Histologic Examinations
4.3. mRNA Expression Levels of Megalin, Cytochrome P450 1a1 (cyp1a1), and cyp1b1
4.4. Protein Expression Levels of Megalin, MT1A, MT2A, TNFα, Caspase 3, and Cleaved Caspase 3
4.5. Statistical Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bas, E.; Van De Water, T.R.; Gupta, C.; Dinh, J.; Vu, L.; Martinez-Soriano, F.; Lainez, J.M.; Marco, J. Efficacy of three drugs for protecting against gentamicin-induced hair cell and hearing losses. Br. J. Pharmacol. 2012, 166, 1888–1904. [Google Scholar] [CrossRef] [Green Version]
- Alharazneh, A.; Luk, L.; Huth, M.; Monfared, A.; Steyger, P.S.; Cheng, A.G.; Ricci, A.J. Functional hair cell mechanotransducer channels are required for aminoglycoside ototoxicity. PLoS ONE 2011, 6, e22347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choung, Y.H.; Taura, A.; Pak, K.; Choi, S.J.; Masuda, M.; Ryan, A.F. Generation of highly-reactive oxygen species is closely related to hair cell damage in rat organ of Corti treated with gentamicin. Neuroscience 2009, 161, 214–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanvers-Kaminsky, C.; Zehnhoff-Dinnesen, A.A.; Parfitt, R.; Ciarimboli, G. Drug-induced ototoxicity: Mechanisms, Pharmacogenetics, and protective strategies. Clin. Pharmacol. Ther. 2017, 101, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Ciarimboli, G.; Deuster, D.; Knief, A.; Sperling, M.; Holtkamp, M.; Edemir, B.; Pavenstadt, H.; Lanvers-Kaminsky, C.; am Zehnhoff-Dinnesen, A.; Schinkel, A.H.; et al. Organic cation transporter 2 mediates cisplatin-induced oto- and nephrotoxicity and is a target for protective interventions. Am. J. Pathol. 2010, 176, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.I.; Birn, H. Megalin and cubilin: Multifunctional endocytic receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 256–266. [Google Scholar] [CrossRef]
- Konig, O.; Ruttiger, L.; Muller, M.; Zimmermann, U.; Erdmann, B.; Kalbacher, H.; Gross, M.; Knipper, M. Estrogen and the inner ear: Megalin knockout mice suffer progressive hearing loss. FASEB J. 2008, 22, 410–417. [Google Scholar] [CrossRef]
- Moestrup, S.K.; Verroust, P.J. Megalin- and cubilin-mediated endocytosis of protein-bound vitamins, lipids, and hormones in polarized epithelia. Annu. Rev. Nutr. 2001, 21, 407–428. [Google Scholar] [CrossRef]
- McWilliam, S.J.; Antoine, D.J.; Smyth, R.L.; Pirmohamed, M. Aminoglycoside-induced nephrotoxicity in children. Pediatr. Nephrol. 2017, 32, 2015–2025. [Google Scholar] [CrossRef]
- Hosokawa, S.; Hosokawa, K.; Ishiyama, G.; Ishiyama, A.; Lopez, I.A. Immunohistochemical localization of megalin and cubilin in the human inner ear. Brain Res. 2018, 1701, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Elsakka, E.G.E.; Elsisi, A.M.; Mansour, O.A.A.; Elsadek, B.E.M.; Abd Elaziz, A.I.; Salama, S.A.; Allam, S. Androgen/androgen receptor affects gentamicin-induced nephrotoxicity through regulation of megalin expression. Life Sci. 2020, 251, 117628. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.W.; Chang, K.H.; Park, S.N.; Suh, B.D. Effects of testosterone in the treatment of immune-mediated sensorineural hearing loss. Eur. Arch. Oto-Rhino-Laryngol. 2003, 260, 316–319. [Google Scholar] [CrossRef]
- Axelstad, M.; Boberg, J.; Hougaard, K.S.; Christiansen, S.; Jacobsen, P.R.; Mandrup, K.R.; Nellemann, C.; Lund, S.P.; Hass, U. Effects of pre- and postnatal exposure to the UV-filter octyl methoxycinnamate (OMC) on the reproductive, auditory and neurological development of rat offspring. Toxicol. Appl. Pharmacol. 2011, 250, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Tauris, J.; Christensen, E.I.; Nykjaer, A.; Jacobsen, C.; Petersen, C.M.; Ovesen, T. Cubilin and megalin co-localize in the neonatal inner ear. Audiol. Neurootol. 2009, 14, 267–278. [Google Scholar] [CrossRef]
- Park, Y.H. Do Women Have Better Hearing Than Men? Clin. Exp. Otorhinolaryngol. 2021, 14, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.T.; Lee, J.H.; Carpena, N.T.; Lee, M.Y.; Chung, P.S.; Jung, J.Y. Estrogen Replacement Reduces Hearing Threshold Shifts and Cochlear Hair Cell Loss After Acoustic Overexposure in Ovariectomized Rats. Clin. Exp. Otorhinolaryngol. 2021, 14, 61–68. [Google Scholar] [CrossRef]
- Ambjorn, M.; Asmussen, J.W.; Lindstam, M.; Gotfryd, K.; Jacobsen, C.; Kiselyov, V.V.; Moestrup, S.K.; Penkowa, M.; Bock, E.; Berezin, V. Metallothionein and a peptide modeled after metallothionein, EmtinB, induce neuronal differentiation and survival through binding to receptors of the low-density lipoprotein receptor family. J. Neurochem. 2008, 104, 21–37. [Google Scholar] [CrossRef]
- Cai, L.; Wang, Y.; Zhou, G.; Chen, T.; Song, Y.; Li, X.; Kang, Y.J. Attenuation by metallothionein of early cardiac cell death via suppression of mitochondrial oxidative stress results in a prevention of diabetic cardiomyopathy. J. Am. Coll. Cardiol. 2006, 48, 1688–1697. [Google Scholar] [CrossRef] [Green Version]
- Mazurek, B.; Amarjargal, N.; Haupt, H.; Fuchs, J.; Olze, H.; Machulik, A.; Gross, J. Expression of genes implicated in oxidative stress in the cochlea of newborn rats. Hear. Res. 2011, 277, 54–60. [Google Scholar] [CrossRef]
- West, A.K.; Hidalgo, J.; Eddins, D.; Levin, E.D.; Aschner, M. Metallothionein in the central nervous system: Roles in protection, regeneration and cognition. Neurotoxicology 2008, 29, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Jakovac, H.; Kezele, T.G.; Radosevic-Stasic, B. Expression Profiles of Metallothionein I/II and Megalin in Cuprizone Model of De- and Remyelination. Neuroscience 2018, 388, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Hsu, C.C.; Lin, M.T.; Chen, S.H. Flutamide, an androgen receptor antagonist, improves heatstroke outcomes in mice. Eur. J. Pharmacol. 2012, 688, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Raju, R.; Chaudry, I.H. Sex steroids/receptor antagonist: Their use as adjuncts after trauma-hemorrhage for improving immune/cardiovascular responses and for decreasing mortality from subsequent sepsis. Anesth. Analg. 2008, 107, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Asaka, J.; Terada, T.; Okuda, M.; Katsura, T.; Inui, K. Androgen receptor is responsible for rat organic cation transporter 2 gene regulation but not for rOCT1 and rOCT3. Pharm. Res. 2006, 23, 697–704. [Google Scholar] [CrossRef]
- Nyquist, M.D.; Corella, A.; Burns, J.; Coleman, I.; Gao, S.; Tharakan, R.; Riggan, L.; Cai, C.; Corey, E.; Nelson, P.S.; et al. Exploiting AR-Regulated Drug Transport to Induce Sensitivity to the Survivin Inhibitor YM155. Mol. Cancer Res. 2017, 15, 521–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.P.; Yang, S.; Choudhry, M.A.; Hsieh, Y.C.; Bland, K.I.; Chaudry, I.H. Mechanism responsible for the salutary effects of flutamide on cardiac performance after trauma-hemorrhagic shock: Upregulation of cardiomyocyte estrogen receptors. Surgery 2005, 138, 85–92. [Google Scholar] [CrossRef]
- Lee, C.H.; Kim, K.W.; Lee, S.M.; Kim, S.Y. Dose-Dependent Effects of Resveratrol on Cisplatin-Induced Hearing Loss. Int. J. Mol. Sci. 2020, 22. [Google Scholar] [CrossRef]
- Alvarado, J.C.; Fuentes-Santamaria, V.; Jareno-Flores, T.; Blanco, J.L.; Juiz, J.M. Normal variations in the morphology of auditory brainstem response (ABR) waveforms: A study in Wistar rats. Neurosci. Res. 2012, 73, 302–311. [Google Scholar] [CrossRef]
- Lee, C.H.; Park, S.S.; Lee, D.H.; Lee, S.M.; Kim, M.Y.; Choi, B.Y.; Kim, S.Y. Tauroursodeoxycholic acid attenuates cisplatin-induced hearing loss in rats. Neurosci. Lett. 2020, 722, 134838. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Jung, G.; Shim, Y.J.; Koo, J.W. The Novel Peptide Vaccine GV1001 Protects Hearing in a Kanamycin-induced Ototoxicity Mouse Model. Otol. Neurotol. 2018, 39, e731–e737. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Frequencies | Pre-Treatment | Post-Treatment | |||||
|---|---|---|---|---|---|---|---|
| Mean | Standard error | p-value | Mean | Standard error | p-value | ||
| 4 kHz | 0.883 | 0.003 * | |||||
| Control | 37.50 | 2.50 | 38.33 | 1.67 | |||
| FM | 35.00 | 3.78 | 37.50 | 1.64 | |||
| KM | 35.00 | 1.89 | 53.75 | 6.80 | 0.042 † (0.019 **) | ||
| KM + FM | 36.25 | 1.55 | 36.67 | 1.81 | 0.002 ‡ | ||
| 8 kHz | 0.659 | 0.014 * | |||||
| Control | 37.50 | 4.53 | 43.33 | 2.11 | |||
| FM | 36.25 | 5.96 | 41.25 | 5.15 | |||
| KM | 42.50 | 1.64 | 56.25 | 4.98 | 0.033 † (0.020 **) | ||
| KM + FM | 39.38 | 1.70 | 42.14 | 1.14 | 0.021 ‡ | ||
| 16 kHz | 0.320 | 0.005 * | |||||
| Control | 31.25 | 2.95 | 33.33 | 2.11 | |||
| FM | 41.25 | 2.95 | 36.25 | 3.75 | |||
| KM | 30.00 | 3.78 | 47.50 | 5.90 | 0.048 † (0.026 **) | ||
| KM + FM | 30.63 | 1.93 | 30.00 | 1.48 | 0.003 ‡ | ||
| 32 kHz | 0.741 | 0.014 * | |||||
| Control | 51.25 | 5.49 | 61.67 | 3.07 | |||
| FM | 51.25 | 3.98 | 57.50 | 4.53 | |||
| KM | 53.75 | 2.63 | 75.00 | 5.00 | 0.049 † (0.002 **) | ||
| KM + FM | 56.25 | 3.40 | 59.29 | 2.67 | 0.023 ‡ | ||
| Gene | Primer Sequence (Forward) | Primer Sequence (Reverse) | Annealing Temperature (°C) | Product Size (bp) | Sequence Number |
|---|---|---|---|---|---|
| Megalin | 5′- TAGCGATTTGGTTCTCCACC -3′ | 5′- ACTTGTTGGCCTGCATAACC -3′ | 60 | 101 | NM_030827.2 |
| Cyp1a1 | 5′-ATGTCCAGCTCTCAGATGATAAGGTC-3′ | 5′-ATCCCTGCCAATCACTGTGTCTAAC-3′ | 60 | 167 | NM_012540.3 |
| Cyp1b1 | 5′-AAGTCTTGCAGAGTCTGGGC-3′ | 5′-TGGCCATGCTGCGATGAA-3′ | 60 | 92 | NM_012940.2 |
| Gapdh | 5′-AGTGCCAGCCTCGTCTCATA-3′ | 5′-AAGAGAAGGCAGCCCTGGTA-3′ | 60 | 93 | NM_017008.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chun, K.-J.; Lee, C.-H.; Kim, K.-W.; Lee, S.-M.; Kim, S.-Y. Effects of Androgen Receptor Inhibition on Kanamycin-Induced Hearing Loss in Rats. Int. J. Mol. Sci. 2021, 22, 5307. https://doi.org/10.3390/ijms22105307
Chun K-J, Lee C-H, Kim K-W, Lee S-M, Kim S-Y. Effects of Androgen Receptor Inhibition on Kanamycin-Induced Hearing Loss in Rats. International Journal of Molecular Sciences. 2021; 22(10):5307. https://doi.org/10.3390/ijms22105307
Chicago/Turabian StyleChun, Kyung-Ju, Chang-Ho Lee, Kyung-Woon Kim, So-Min Lee, and So-Young Kim. 2021. "Effects of Androgen Receptor Inhibition on Kanamycin-Induced Hearing Loss in Rats" International Journal of Molecular Sciences 22, no. 10: 5307. https://doi.org/10.3390/ijms22105307
APA StyleChun, K. -J., Lee, C. -H., Kim, K. -W., Lee, S. -M., & Kim, S. -Y. (2021). Effects of Androgen Receptor Inhibition on Kanamycin-Induced Hearing Loss in Rats. International Journal of Molecular Sciences, 22(10), 5307. https://doi.org/10.3390/ijms22105307