
EMS Derived Wheat Mutant BIG8-1 (Triticum aestivum L.)—A New Drought Tolerant Mutant Wheat Line

 ,
,  and
and
Abstract
:1. Introduction
2. Results
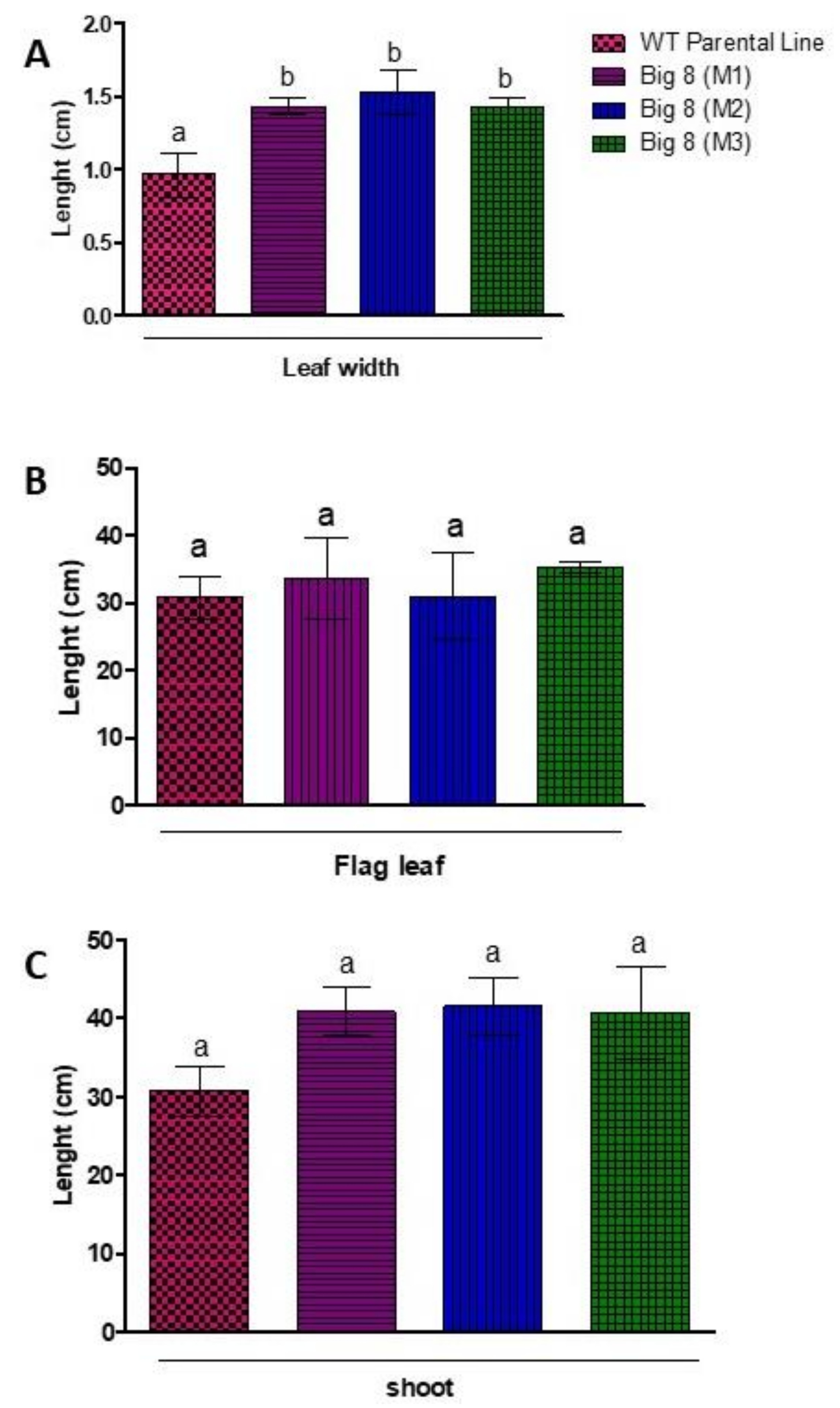
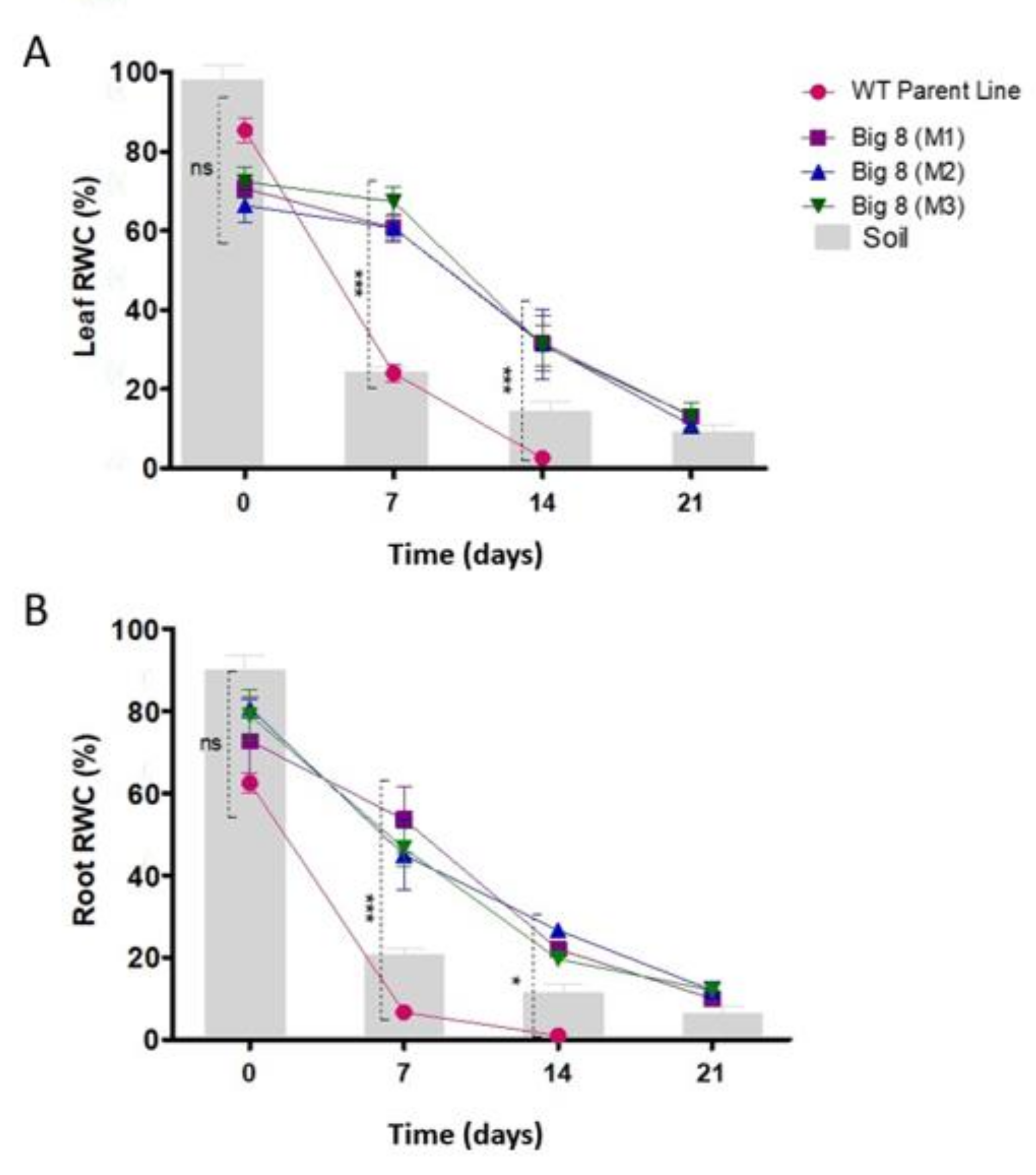
2.1. Plant Phenotypes and Relative Moisture Content
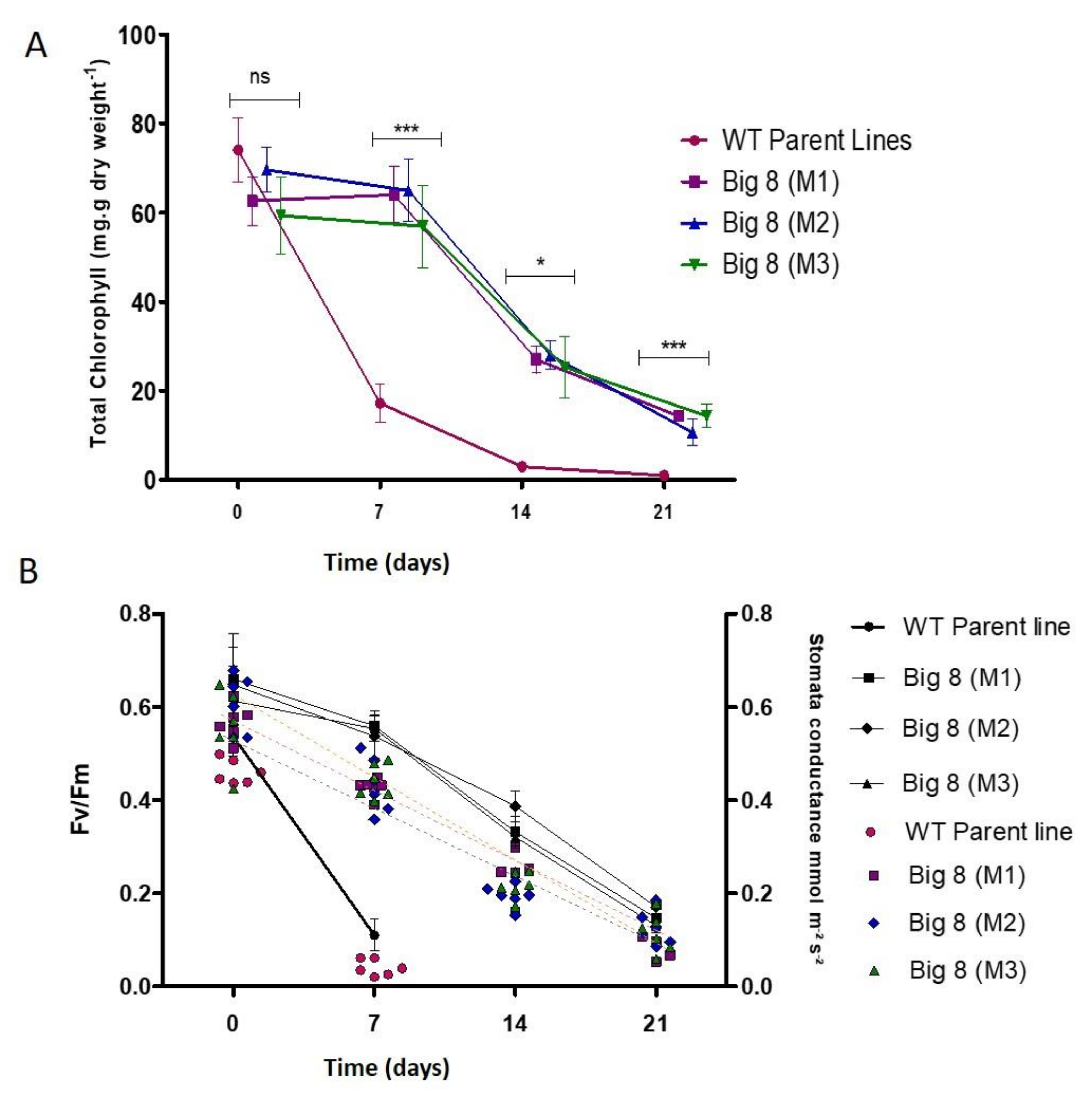
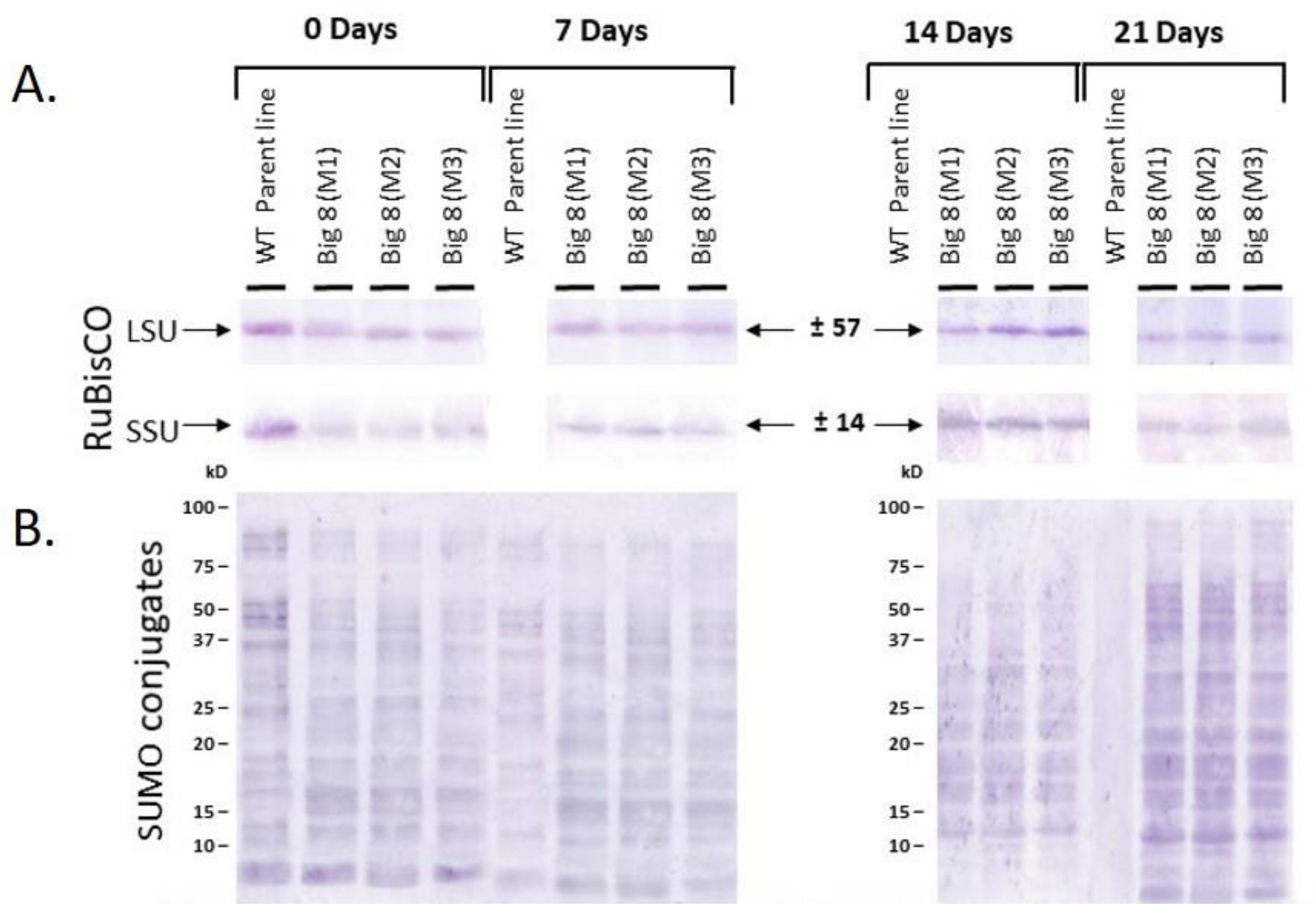
2.2. Chlorophyll, Photosynthesis, Stomatal Conductance and RuBisCo Response
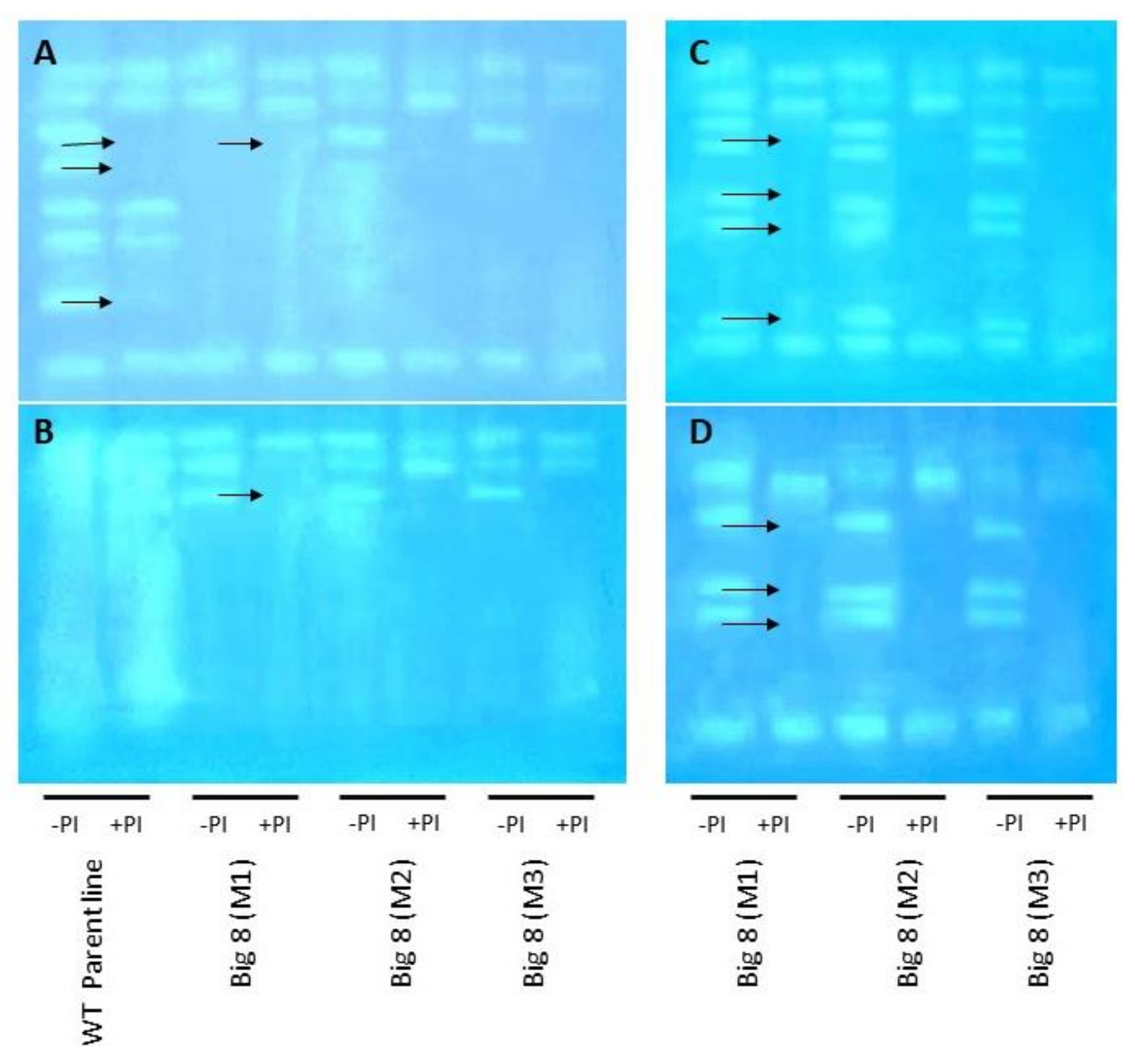
2.3. SUMOylation and Protease Activity under Water Stress Conditions
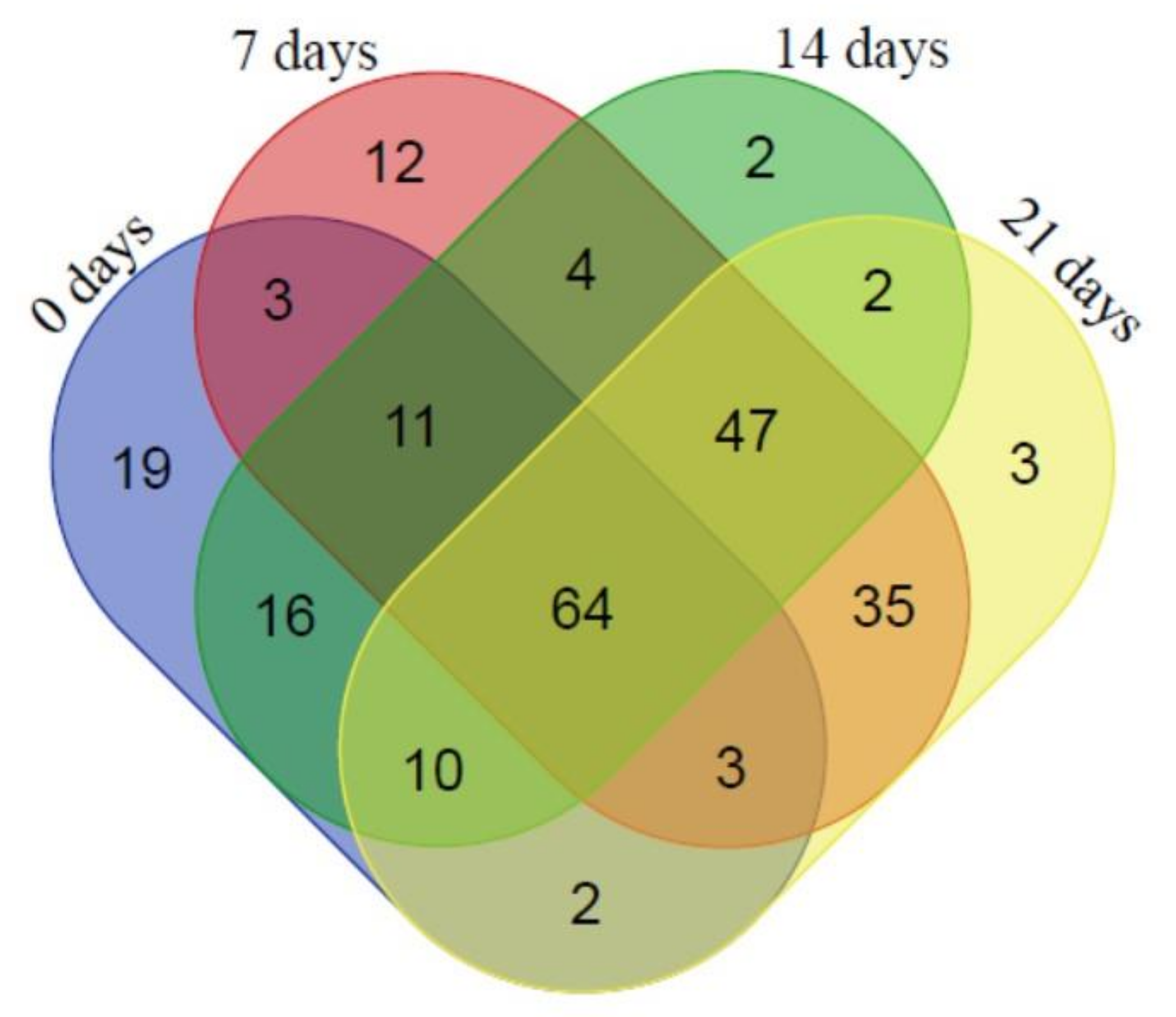
2.4. Changes in the Proteome under Water Deficit Stress Conditions
2.5. Changes in Free Amino Acid under Water Stress
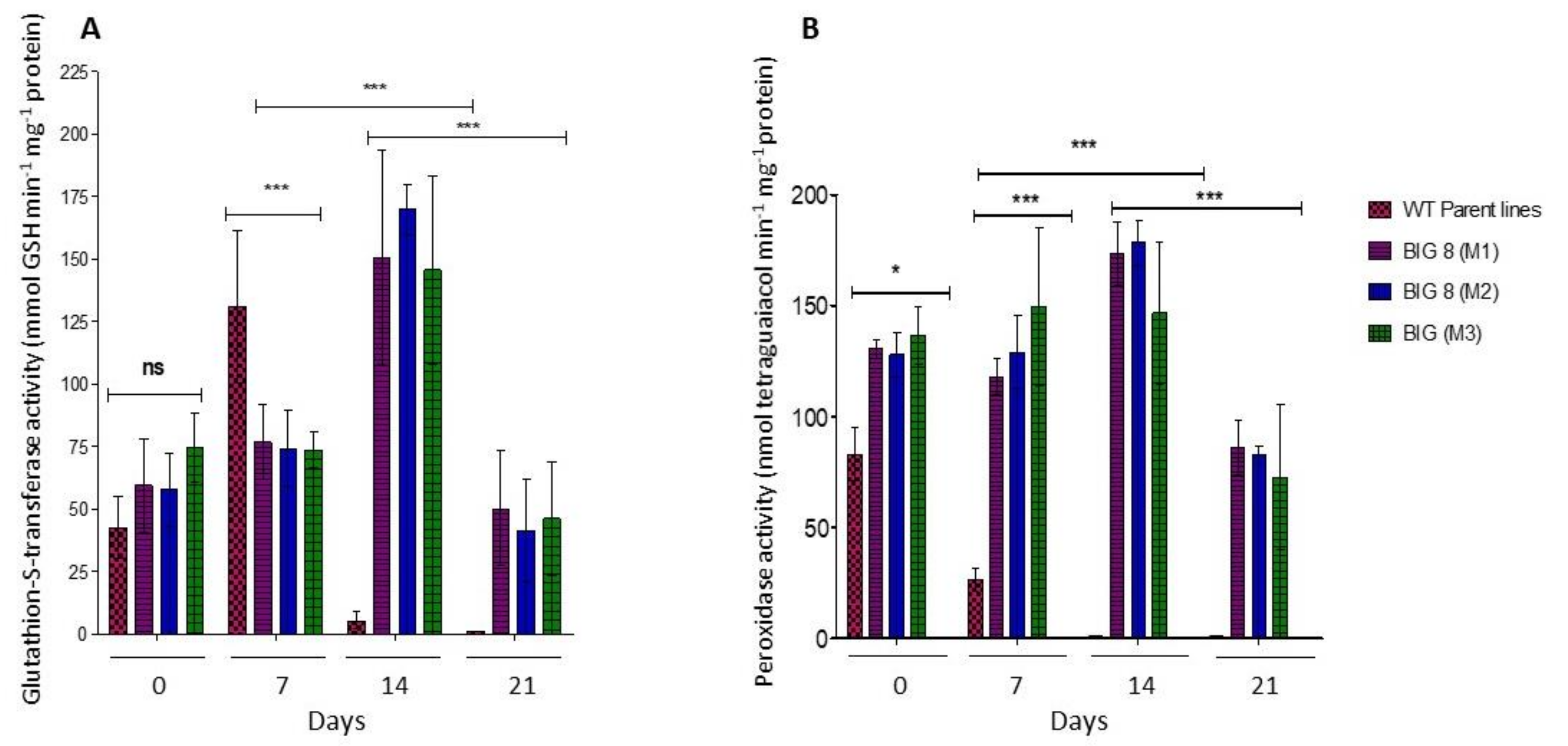
2.6. Oxidative Defense
3. Discussion
4. Material and Methods
4.1. Plant Materials, Growth Conditions Water Deficit Stress Treatments
4.2. Measurement of Plant Growth and Relative Water Content
4.3. Chlorophyll Fluorescence (Fv/Fm)Stomatal Conductance Chlorophyll Content
4.4. SDS-PAGE Electrophoresis and Western Blot Analysis
4.5. Cysteine Proteases Activity
4.6. Amino Acid Extraction and Quantification
4.7. Protein Extraction, Quantification Digestion
4.8. Enzyme Measurements
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gill, B.S.; Appels, R.; Botha-Oberholster, A.-M.; Buell, C.R.; Bennetzen, J.L.; Chalhoub, B.; Chumley, F.; Dvořák, J.; Iwanaga, M.; Keller, B.; et al. A workshop report on wheat genome sequencing: International genome research on wheat consortium. Genetics 2004, 168, 1087–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallerstein, D. Food-energy-water (FEW) nexus: Rearchitecting the planet to accommodate 10 billion humans by 2050. Resour. Conserv. Recycl. 2020, 155, 104658. [Google Scholar] [CrossRef]
- Borisjuk, N.; Kishchenko, O.; Eliby, S.; Schramm, C.; Anderson, P.; Jatayev, S.; Kurishbayev, A.; Shavrukov, Y. Genetic modification for wheat improvement: From transgenesis to genome editing. Biomed. Res. Int. 2019, 2019, 18. [Google Scholar] [CrossRef] [PubMed]
- Lampl, N.; Budai-Hadrian, O.; Davydov, O.; Joss, T.V.; Harrop, S.J.; Curmi, P.M.G.; Roberts, T.H.; Fluhr, R. Arabidopsis atserpin1, crystal structure and in vivo interaction with its target protease responsive to desiccation-21 (RD21). J. Biol. Chem. 2010, 285, 13550–13560. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M. Stress-induced changes in wheat grain composition and quality. Crit. Rev. Food Sci. Nutr. 2014, 54, 1576–1583. [Google Scholar] [CrossRef]
- Bazargani, M.M.; Sarhadi, E.; Bushehri, A.-A.S.; Matros, A.; Mock, H.-P.; Naghavi, M.-R.; Hajihoseini, V.; Mardi, M.; Hajirezaei, M.-R.; Moradi, F.; et al. A proteomics view on the role of drought-induced senescence and oxidative stress defense in enhanced stem reserves remobilization in wheat. J. Proteom. 2011, 74, 1959–1973. [Google Scholar] [CrossRef]
- Ford, K.L.; Cassin, A.; Bacic, A.F. Quantitative proteomic analysis of wheat cultivars with differing drought stress tolerance. Front. Plant Sci. 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Michaletti, A.; Naghavi, M.R.; Toorchi, M.; Zolla, L.; Rinalducci, S. Metabolomics and proteomics reveal drought-stress responses of leaf tissues from spring-wheat. Sci. Rep. 2018, 8, 5710. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Fountain, J.C.; Ji, P.; Ni, X.; Chen, S.; Lee, R.D.; Kemerait, R.C.; Guo, B. Deciphering drought-induced metabolic responses and regulation in developing maize kernels. Plant Biotechnol. J. 2018. [Google Scholar] [CrossRef] [Green Version]
- Zörb, C.; Becker, E.; Merkt, N.; Kafka, S.; Schmidt, S.; Schmidhalter, U. Shift of grain protein composition in bread wheat under summer drought events. J. Plant Nutr. Soil Sci. 2017, 180, 49–55. [Google Scholar] [CrossRef]
- Gul, F.; Jan, D.; Ashfaq, M. Assessing the socio-economic impact of climate change on wheat production in khyber pakhtunkhwa, Pakistan. Environ. Sci. Pollut. Res. 2019, 26, 6576–6585. [Google Scholar] [CrossRef]
- Hansen, J.; Hellin, J.; Rosenstock, T.; Fisher, E.; Cairns, J.; Stirling, C.; Lamanna, C.; van Etten, J.; Rose, A.; Campbell, B. Climate risk management and rural poverty reduction. Agric. Syst. 2019, 172, 28–46. [Google Scholar] [CrossRef]
- Karatay, Y.N.; Meyer-Aurich, A. Profitability and downside risk implications of site-specific nitrogen management with respect to wheat grain quality. Precis. Agric. 2020, 21, 449–472. [Google Scholar] [CrossRef]
- Luo, L.; Xia, H.; Lu, B.-R. Editorial: Crop breeding for drought resistance. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Arabi, M.I.E.; Shoaib, A.; Al-Shehadah, E.; Jawhar, M. Genetic diversity within local and introduced cultivars of wheat (Triticum aestivum L.) grown under mediterranean environment as revealed by AFLP markers. Acta Biologica Szegediensis 2019, 63, 25–30. [Google Scholar] [CrossRef]
- Eid, M. RAPD fingerprinting and genetic relationships of some wheat genotypes. Int. J. Genet. Genom. 2019, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Joshi, C.P.; Nguyen, H.T. RAPD (random amplified polymorphic DNA) analysis based intervarietal genetic relationships among hexaploid wheats. Plant Sci. 1993, 93, 95–103. [Google Scholar] [CrossRef]
- Reif, J.C.; Zhang, P.; Dreisigacker, S.; Warburton, M.L.; van Ginkel, M.; Hoisington, D.; Bohn, M.; Melchinger, A.E. Wheat genetic diversity trends during domestication and breeding. Theor. Appl. Genet. 2005, 110, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Mwadzingeni, L.; Shimelis, H.; Tesfay, S.; Tsilo, T.J. Screening of bread wheat genotypes for drought tolerance using phenotypic and proline analyses. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Chai, L.; Chen, Z.; Xu, L.; Zhai, H.; Zhao, A.; Peng, H.; Yao, Y.; You, M.; Sun, Q. Identification and characterization of a high kernel weight mutant induced by gamma radiation in wheat (Triticum aestivum L.). BMC Genet. 2015, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Iqbal, M.A.; Till, B.J.; Rahman, M. Identification of induced mutations in hexaploid wheat genome using exome capture assay. PLoS ONE 2018, 13, e0201918. [Google Scholar] [CrossRef] [Green Version]
- Adamu, A.K.; Aliyu, H. Morphogical effects of sodium azide on tomato (Lycopersicon esculentum Mill). Sci. World J. 2007, 2. [Google Scholar] [CrossRef]
- Ahmed, H.G.M.-D.; Khan, A.S.; Li, M.; Khan, S.H.; Kashif, M. Early selection of bread wheat genotypes using morphological and photosynthetic attributes conferring drought tolerance. J. Integr. Agric. 2019, 18, 2483–2491. [Google Scholar] [CrossRef]
- Baloch, A.W.; Soomro, A.M.; Javed, M.A.; Bughio, M.S.; Mastoi, N.N. Impact of reduced culm length on yieldand yield parameters in rice. Asian J. Plant Sci. 2002, 1, 39–40. [Google Scholar]
- Erdem, G.; Oldacay, S. Employment of RAPD technique to assess the genetic stability of helianthus annuus treated with different mutagenic agents. J. Appl. Sci. 2004, 4, 277–281. [Google Scholar] [CrossRef]
- Ilbas, A.I.; Eroglu, Y.; Eroglu, H.E. Effects of the application of different concentrations of Na N3 for different times on the morphological and cytogenetic characteristics of barley (Hordeum vulgare L.). J. Integr. Plant Biol. 2005, 47, 1101–1106. [Google Scholar] [CrossRef]
- Khan, S.; Goyal, S. Improvement of mungbean varieties through induced mutations. AJPS 2009, 3, 174–180. [Google Scholar] [CrossRef]
- Kozgar, M.I.; Goyal, S.; Khan, S. EMS induced mutational variability in vigna radiata and vigna mungo. Res. J. Bot. 2011, 6, 31. [Google Scholar]
- Mostafa, G. Effect of sodium azide on the growth and variability induction in Helianthus annuus L. Int. J. Plant Breed. Genet. 2011, 5, 76–85. [Google Scholar] [CrossRef]
- Rachovska, G.; Dimova, D. Effect of sodium azide and gamma rays on M1 quantitative characteristics of the productivity and their connection with M2 mutation changes in winter common wheat. Rasteniev’dni Nauki 2000, 37, 413–419. [Google Scholar]
- Rime, J.; Dinesh, M.R.; Sankaran, M.; Shivashankara, K.S.; Rekha, A.; Ravishankar, K.V. Evaluation and characterization of EMS derived mutant populations in mango. Scientia Horticulturae 2019, 254, 55–60. [Google Scholar] [CrossRef]
- Singh, S.; Singh, R.; Prasad, J.; Agrawal, R.; Shahi, J. Induced genetic variability for protein content, yield and yield components in microsperma lentil (Lens culinaris Medik.). Madras Agric. J. 2006, 93, 155–159. [Google Scholar]
- Tah, P.R. Induced macromutation in mungbean [Vigna radiata (L.) Wilczek]. Int. J. Bot. 2006, 3, 219–228. [Google Scholar]
- Wani, M.R.; Khan, S. Estimates of genetic variability in mutated populations and the scope of selection for yield attributes in Vigna radiata (L.) Wilczek. Egypt. J. Biol. 2006, 8. [Google Scholar] [CrossRef]
- Nazarenko, M. Parameters of winter wheat growing and development after mutagen action. Agric. Food Eng. 2016, 9, 8. [Google Scholar]
- Nazarenko, M.; Lykholat, Y.; Grygoryuk, I.; Khromikh, N. Optimal doses and concentrations of mutagens for winter wheat breeding purposes. Part I. Grain productivity. J. Cent. Eur. Agric. 2018, 19, 194–205. [Google Scholar] [CrossRef]
- Le Roux, M.-S.; Burger, N.F.V.; Vlok, M.; Kunert, K.J.; Cullis, C.A.; Botha, A.-M. Wheat line “RYNO3936” is associated with delayed water stress-induced leaf senescence and rapid water-deficit stress recovery. Front. Plant Sci. 2020, 11, 1053. [Google Scholar] [CrossRef] [PubMed]
- Levitt, E. Responses of plants to environmental stresses. In Drought Avoidance, 2nd ed.; Academic Press: Cambridge, MA, USA, 1980; Volume 1. [Google Scholar]
- Kozlowski, T.T. Water supply and tree growth. Part I. Water deficits. Bot. Rev. 1982, 43, 57–95. [Google Scholar]
- Rochaix, J.-D. Assembly of the photosynthetic apparatus. Plant Physiol. 2011, 155, 1493–1500. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Kouzmanova, M.; Kalaji, H.M.; Yordanov, I.; Krasteva, V.; Alexandrov, V.; Stefanov, D.; Allakhverdiev, S.I.; et al. Drought-induced modifications of photosynthetic electron transport in intact leaves: Analysis and use of neural networks as a tool for a rapid non-invasive estimation. Biochimica et Biophysica Acta Bioenerg. 2012, 1817, 1490–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huseynova, I.M.; Suleymanov, S.Y.; Aliyev, J.A. Structural-functional state of thylakoid membranes of wheat genotypes under water stress. Biochimica et Biophysica Acta Bioenerg. 2007, 1767, 869–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ögren, E.; Sjöström, M. Estimation of the effect of photoinhibition on the carbon gain in leaves of a willow canopy. Planta 1990, 181, 560–567. [Google Scholar] [CrossRef]
- Roháček, K. Chlorophyll fluorescence parameters: The Definitions, photosynthetic meaning, and mutual relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Shah, N.H.; Paulsen, G.M. Interaction of drought and high temperature on photosynthesis and grain-filling of wheat. Plant Soil 2003, 257, 219–226. [Google Scholar] [CrossRef]
- Galmés, J.; Capó-Bauçà, S.; Niinemets, Ü.; Iñiguez, C. Potential improvement of photosynthetic CO2 assimilation in crops by exploiting the natural variation in the temperature response of rubisco catalytic traits. Curr. Opin. Plant Biol. 2019, 49, 60–67. [Google Scholar] [CrossRef]
- Iñiguez, C.; Capó-Bauçà, S.; Niinemets, Ü.; Stoll, H.; Aguiló-Nicolau, P.; Galmés, J. Evolutionary trends in RuBisCO kinetics and their co-evolution with CO2 concentrating mechanisms. Plant J. 2020, 101, 897–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degl’Innocenti, E.; Hafsi, C.; Guidi, L.; Navari-Izzo, F. The effect of salinity on photosynthetic activity in potassium-deficient barley species. J. Plant Physiol. 2009, 166, 1968–1981. [Google Scholar] [CrossRef]
- Misson, L.; Limousin, J.-M.; Rodriguez, R.; Letts, M.G. Leaf physiological responses to extreme droughts in Mediterranean quercus ilex forest. Plant Cell Environ. 2010, 33, 1898–1910. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Murata, N. How do environmental stresses accelerate photoinhibition? Trends Plant Sci. 2008, 13, 178–182. [Google Scholar] [CrossRef]
- Janků, M.; Luhová, L.; Petřivalský, M. On the origin and fate of reactive oxygen species in plant cell compartments. Antioxidants 2019, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought responses of leaf tissues from wheat cultivars of differing drought tolerance at the metabolite level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Shi, L.; Jiao, Y.; Li, M.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Metabolic responses to drought stress in the tissues of drought-tolerant and drought-sensitive wheat genotype seedlings. AoB Plants 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Ullah, N.; Yüce, M.; Gökçe, Z.N.O.; Budak, H. Comparative metabolite profiling of drought stress in roots and leaves of seven triticeae species. BMC Genom. 2017, 18, 969. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Wang, M.; Li, F.; Lv, H.; Li, C.; Xia, G. A proteomic study of the response to salinity and drought stress in an introgression strain of bread wheat. Mol. Cell. Proteom. 2009, 8, 2676–2686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Sultan, M.A.R.F.; Liu, X.L.; Zhang, J.; Yu, F.; Zhao, H.X. Physiological and comparative proteomic analysis reveals different drought responses in roots and leaves of drought-tolerant wild wheat (Triticum boeoticum). PLoS ONE 2015, 10, e0121852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, D.; Crosatti, C.; Khoshro, H.H.; Mastrangelo, A.M.; Mica, E.; Mazzucotelli, E. Post-transcriptional and post-translational regulations of drought and heat response in plants: A spider’s web of mechanisms. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Hashiguchi, A.; Komatsu, S. Impact of post-translational modifications of crop proteins under abiotic stress. Proteomes 2016, 4, 42. [Google Scholar] [CrossRef]
- Su, S.; Zhang, Y.; Liu, P. Roles of ubiquitination and SUMOylation in DNA damage reponse. Curr. Issues Mol. Biol. 2019, 35, 59–84. [Google Scholar] [CrossRef] [Green Version]
- Morrell, R.; Sadanandom, A. Dealing with stress: A review of plant SUMO proteases. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Li, Y.; Wang, G.; Xu, Z.; Li, J.; Sun, M.; Guo, J.; Ji, W. Organization and regulation of soybean SUMOylation system under abiotic stress conditions. Front. Plant Sci. 2017, 8, 1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, A.K.; Zhang, C.; Yates, G.; Bailey, M.; Brown, A.; Sadanandom, A. SUMO is a critical regulator of salt stress responses in rice. Plant Physiol. 2016, 170, 2378–2391. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.K.; Zhang, C.; Caine, R.S.; Gray, J.; Sadanandom, A. Rice SUMO protease overly tolerant to salt 1 targets the transcription factor, OsbZIP23 to promote drought tolerance in rice. Plant J. 2017, 92, 1031–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [Green Version]
- Saldanha, A.J. Java treeview—Extensible visualization of microarray data. Bioinformatics 2004, 20, 3246–3248. [Google Scholar] [CrossRef] [Green Version]
- Kooyers, N.J. The evolution of drought escape and avoidance in natural herbaceous populations. Plant Sci. 2015, 234, 155–162. [Google Scholar] [CrossRef]
- Kramer, P.J. The relation between rate of transpiration and rate of absorption of water in plants. Am. J. Bot. 1937, 24, 10–15. [Google Scholar] [CrossRef]
- Barber, S.A.; Silberbush, M. Plant root morphology and nutrient uptake. In Roots, Nutrient and Water Influx, and Plant Growth; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1984; pp. 65–87. [Google Scholar] [CrossRef]
- Eissenstat, D.M. Trade-offs in root form and function. In Ecology andAgriculture; Academic Press: San Diego, CA, USA, 1997; pp. 173–199. [Google Scholar]
- Vesala, T.; Sevanto, S.; Grönholm, T.; Salmon, Y.; Nikinmaa, E.; Hari, P.; Hölttä, T. Effect of leaf water potential on internal humidity and CO2 dissolution: Reverse transpiration and improved water use efficiency under negative pressure. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ries, L.L.; Purcell, L.C.; Carter, T.E.; Edwards, J.T.; King, C.A. Physiological traits contributing to differential canopy wilting in soybean under drought. Crop Sci. 2012, 52, 272–281. [Google Scholar] [CrossRef]
- Li, R.; Guo, P.-G.; Michael, B.; Stefania, G.; Salvatore, C. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Nikolaeva, M.K.; Maevskaya, S.N.; Shugaev, A.G.; Bukhov, N.G. Effect of drought on chlorophyll content and antioxidant enzyme activities in leaves of three wheat cultivars differing in productivity. Russ. J. Plant Physiol. 2010, 57, 87–95. [Google Scholar] [CrossRef]
- Nowsherwan, I.; Shabbir, G.; Malik, S.; Ilyas, M.; Iqbal, M.; Musa, M. Effect of drought stress on different physiological traits in bread wheat. SAARC J. Agric. 2018, 16, 1–6. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defence mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Pour-Aboughadareh, A.; Ahmadi, J.; Mehrabi, A.A.; Etminan, A.; Moghaddam, M.; Siddique, K.H.M. Physiological responses to drought stress in wild relatives of wheat: Implications for wheat improvement. Acta Physiol. Plant. 2017, 39, 106. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Jabran, K.; Cheema, M.A. Terminal drought and heat stress alter physiological and biochemical attributes in flag leaf of bread wheat. PLoS ONE 2020, 15, e0232974. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E. Plant Physiology; Sinauer Associates: Sunderland, MA, USA, 2006. [Google Scholar]
- Liu, J.; Guo, Y.Y.; Bai, Y.W.; Camberato, J.J.; Xue, J.Q.; Zhang, R.H. Effects of drought stress on the photosynthesis in maize. Russ. J. Plant. Physiol. 2018, 65, 849–856. [Google Scholar] [CrossRef]
- Xu, Q.; Ma, X.; Lv, T.; Bai, M.; Wang, Z.; Niu, J. Effects of water stress on fluorescence parameters and photosynthetic characteristics of drip irrigation in rice. Water 2020, 12, 289. [Google Scholar] [CrossRef] [Green Version]
- Mathobo, R.; Marais, D.; Steyn, J.M. The effect of drought stress on yield, leaf gaseous exchange and chlorophyll fluorescence of dry beans (Phaseolus vulgaris L.). Agric. Water Manag. 2017, 180, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.K.; Yang, Z.Q.; Li, Y.X.; Liu, Q.; Han, W. Effects of different levels of water stress on leaf photosynthetic characteristics and antioxidant enzyme activities of greenhouse tomato. Photosynthetica 2016, 54, 28–39. [Google Scholar] [CrossRef]
- Nakamura, S.; Izumi, M. Regulation of chlorophagy during photoinhibition and senescence: Lessons from mitophagy. Plant Cell Physiol. 2018, 59, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Amreen; Tantray, A.Y.; Khan, N.A.; Ahmad, A. Reactive oxygen species detection-approaches in plants: Insights into genetically encoded FRET-based sensors. J. Biotechnol. 2020, 308, 108–117. [Google Scholar] [CrossRef]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Trivedi, P.K. Glutathione S-transferases: Role in combating abiotic stresses including arsenic detoxification in plants. Front. Plant Sci. 2018, 9, 751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noctor, G.; Reichheld, J.-P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef] [PubMed]
- Batista-Silva, W.; da Fonseca-Pereira, P.; Martins, A.O.; Zsögön, A.; Nunes-Nesi, A.; Araújo, W.L. Engineering improved photosynthesis in the era of synthetic biology. Plant Commun. 2020, 1, 100032. [Google Scholar] [CrossRef]
- De Lomana, A.L.G.; Schäuble, S.; Valenzuela, J.; Imam, S.; Carter, W.; Bilgin, D.D.; Yohn, C.B.; Turkarslan, S.; Reiss, D.J.; Orellana, M.V. Transcriptional program for nitrogen starvation-induced lipid accumulation in chlamydomonas reinhardtii. Biotechnol. Biofuels 2015, 8, 1–18. [Google Scholar]
- Feller, U.; Anders, I.; Mae, T. Rubiscolytics: Fate of rubisco after its enzymatic function in a cell is terminated. J. Exp. Bot. 2008, 59, 1615–1624. [Google Scholar] [CrossRef] [Green Version]
- Recuenco-Muñoz, L.; Offre, P.; Valledor, L.; Lyon, D.; Weckwerth, W.; Wienkoop, S. Targeted quantitative analysis of a diurnal RuBisCO subunit expression and translation profile in Chlamydomonas reinhardtii introducing a novel mass western approach. J. Proteom. 2015, 113, 143–153. [Google Scholar] [CrossRef]
- González-Rábade, N.; Badillo-Corona, J.A.; Aranda-Barradas, J.S.; del Carmen Oliver-Salvador, M. Production of plant proteases in vivo and in vitro—A review. Biotechnol. Adv. 2011, 29, 983–996. [Google Scholar] [CrossRef] [PubMed]
- Botha, A.-M.; Kunert, K.J.; Cullis, C.A. Cysteine proteases and wheat (Triticum aestivum L.) under drought: A still greatly unexplored association. Plant Cell Environ. 2017, 40, 1679–1690. [Google Scholar] [CrossRef]
- Gur, E.; Biran, D.; Ron, E.Z. Regulated proteolysis in gram-negative bacteria—How and when? Nat. Rev. Microbiol. 2011, 9, 839–848. [Google Scholar] [CrossRef]
- Zhang, Y.; Zeng, L. Crosstalk between ubiquitination and other posttranslational protein modifications in plant immunity. Plant Commun. 2020, 1, 100041. [Google Scholar] [CrossRef]
- Gareau, J.R.; Lima, C.D. The SUMO pathway: Emerging mechanisms that shape specificity, conjugation and recognition. Nat. Rev. Mol. Cell Biol. 2010, 11, 861–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerscher, O.; Felberbaum, R.; Hochstrasser, M. Modification of proteins by ubiquitin and ubiquitin-like proteins. Annu. Rev. Cell Dev. Biol. 2006, 22, 159–180. [Google Scholar] [CrossRef] [Green Version]
- Benlloch, R.; Lois, L.M. Sumoylation in plants: Mechanistic insights and its role in drought stress. J. Exp. Bot. 2018, 69, 4539–4554. [Google Scholar] [CrossRef]
- Kozak, M. Comparison of initiation of protein synthesis in procaryotes, eucaryotes, and organelles. Microbiol. Rev. 1983, 47, 1. [Google Scholar] [CrossRef] [PubMed]
- Varshney, U.; RajBhandary, U.L. Role of methionine and formylation of initiator TRNA in initiation of protein synthesis in Escherichia coli. J. Bacteriol. 1992, 174, 7819–7826. [Google Scholar] [CrossRef] [Green Version]
- Ding, P.; Fang, L.; Wang, G.; Li, X.; Huang, S.; Gao, Y.; Zhu, J.; Xiao, L.; Tong, J.; Chen, F.; et al. Wheat methionine sulfoxide reductase A4.1 interacts with heme oxygenase 1 to enhance seedling tolerance to salinity or drought stress. Plant Mol. Biol. 2019, 101, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.C.; Gladyshev, V.N. The biological significance of methionine sulfoxide stereochemistry. Free Radic. Biol. Med. 2011, 50, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, T.H.; Hejgaard, J. Serpins in plants and green algae. Funct. Integr. Genom. 2008, 8, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Botër, M.; Amigues, B.; Peart, J.; Breuer, C.; Kadota, Y.; Casais, C.; Moore, G.; Kleanthous, C.; Ochsenbein, F.; Shirasu, K.; et al. Structural and functional analysis of SGT1 reveals that its interaction with HSP90 is required for the accumulation of Rx, an R protein involved in plant immunity. Plant Cell 2007, 19, 3791–3804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, V.Y.; Shearman, C.W.; Cohen, M.P. Identification of calnexin as a binding protein for amadori-modified glycated albumin. Biochem. Biophys. Res. Commun. 2001, 284, 602–606. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, J.; Xia, L.; Xiang, B.; Li, E. Investigating biological nitrogen cycling in lacustrine systems by FT-ICR-MS analysis of nitrogen-containing compounds in petroleum. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 556, 109887. [Google Scholar] [CrossRef]
- Fierens, E.; Gebruers, K.; Courtin, C.M.; Delcour, J.A. Xylanase inhibitors bind to nonstarch polysaccharides. J. Agric. Food Chem. 2008, 56, 564–570. [Google Scholar] [CrossRef]
- Mbwanji, K. Inducing Mutations in Bread Wheat (Triticum aestivum L.) Using Chemical Treatments; Stellenbosch University: Stellenbosch, South Africa, 2014. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Vendruscolo, E.C.G.; Schuster, I.; Pileggi, M.; Scapim, C.A.; Molinari, H.; Marur, C.J.; Vieira, L.G. Stress-induced synthesis of proline confers tolerance to water deficit in transgenic wheat. J. Plant Physiol. 2007. [Google Scholar] [CrossRef]
- Sade, D.; Sade, N.; Shriki, O.; Lerner, S.; Gebremedhin, A.; Karavani, A.; Brotman, Y.; Osorio, S.; Fernie, A.R.; Willmitzer, L.; et al. Water balance, hormone homeostasis, and sugar signaling are all involved in tomato resistance to tomato yellow leaf curl virus. Plant Physiol. 2014, 165, 1684–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sade, N.; Galkin, E.; Moshelion, M. Measuring arabidopsis, tomato and barley leaf relative water content (RWC). BioProtocols 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Black, A.L.; Power, J.F. Effect of chemical and mechanical fallow methods on moisture storage, wheat yields, and soil erodibility. Soil Sci. Soc. Am. J. 1965, 29, 465–468. [Google Scholar] [CrossRef]
- Engelbrecht, B.M.; Tyree, M.T.; Kursar, T.A. Visual assessment of wilting as a measure of leaf water potential and seedling drought survival. J. Trop. Ecol. 2007, 23, 497–500. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Advances in Photosynthesis and Respiration; Springer: Berlin/Heidelberg, Germany, 2004; pp. 321–362. [Google Scholar] [CrossRef]
- Arnon, D.I. Determination of chlorophyll concentration in leaf tissues of plants. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Rylatt, D.B.; Parish, C.R. Protein determination on an automatic spectrophotometer. Anal. Biochem. 1982, 121, 213–214. [Google Scholar] [CrossRef]
- Botha, F.C.; Small, J.C. Control of ribulose 1, 5-bisphosphate carboxylase synthesis in the cotyledons of citrullus lanatus. Plant Sci. 1987, 53, 121–129. [Google Scholar] [CrossRef]
- Barrett, A.J.; Kembhavi, A.A.; Brown, M.A.; Kirschke, H.; Knight, C.G.; Tamai, M.; Hanada, K. L-trans-epoxysuccinyl-leucylamido(4-Guanidino)butane (E-64) and its analogues as inhibitors of cysteine proteinases including cathepsins B, H and L. Biochem. J. 1982, 201, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Mizoue, K.; Kitamura, K.; Tse, W.-C.; Huber, C.P.; Ishida, T. Structural basis of inhibition of cysteine proteases by E-64 and its derivatives. Pept. Sci. 1999, 51, 99–107. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Searle, B.C. Scaffold: A bioinformatic tool for validating MS/MS-based proteomic studies. Proteomics 2010, 10, 1265–1269. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Botha, A.-M.; van Eck, L.; Burger, N.F.V.; Swanevelder, Z.H. Near-isogenic lines of Triticum aestivum with distinct modes of resistance exhibit dissimilar transcriptional regulation during Diuraphis noxia feeding. Biol. Open 2014, 3, 1116–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zieslin, N.; Ben Zaken, R. Peroxidase activity and presence of phenolic substances in peduncles of rose flowers. Plant Physiol. Biochem. 1993, 31, 333–339. [Google Scholar]
- Venisse, J.-S.; Gullner, G.; Brisset, M.-N. Evidence for the involvement of an oxidative stress in the initiation of infection of pear by Erwinia amylovora. Plant Physiol. 2001, 125, 2164–2172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motulsky, H. Prism 5 Statistics Guide; GraphPad Prism: San Diego, CA, USA, 2007. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Treatment | Free Amino Acid (FAA) Level | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| His | Ser | Arg | Gly | Asp | Glu | Thr | Ala | Pro | Lys | Tyr | Met | Val | Ile | Leu | Phe | ||
| BIG8 (WT) * | Day 0 | 0.10 | 0.33 | 0.32 | 0.31 | 0.55 | 0.59 | 0.24 | 0.39 | 0.29 | 0.39 | 0.21 | 0.54 | 0.31 | 0.18 | 0.42 | 0.34 |
| Day 7 | n.d | 0.20 | n.d | 0.27 | n.d | 0.56 | 0.31 | 0.42 | 0.88 | 0.50 | 0.14 | 2.54 | 0.14 | 0.33 | 0.59 | 1.65 | |
| BIG8-1 (Mutant) | Day 0 | 0.16 | 0.35 | 0.45 | 0.34 | 0.84 | 0.69 | 0.32 | 0.37 | 0.33 | 0.50 | 0.24 | 0.43 | 0.36 | 0.19 | 0.80 | 0.59 |
| Day 7 | 0.10 | 0.14 | n.d | 0.12 | 0.24 | 0.33 | 0.06 | 0.13 | 0.31 | 0.18 | 0.08 | 0.75 | 0.08 | 0.06 | n.d | 0.42 | |
| Day 14 | 0.14 | 0.26 | 0.27 | 0.30 | 0.80 | 0.68 | 0.22 | 0.36 | 0.88 | 0.34 | 0.17 | 0.54 | 0.36 | 0.20 | 0.66 | 0.41 | |
| Day 21 | 0.31 | 0.41 | 0.59 | 0.57 | 1.71 | 2.37 | 0.32 | 0.51 | 4.60 | 0.38 | 0.39 | 0.46 | 0.77 | 0.43 | 0.76 | 0.84 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
le Roux, M.-S.L.; Burger, N.F.V.; Vlok, M.; Kunert, K.J.; Cullis, C.A.; Botha, A.-M. EMS Derived Wheat Mutant BIG8-1 (Triticum aestivum L.)—A New Drought Tolerant Mutant Wheat Line. Int. J. Mol. Sci. 2021, 22, 5314. https://doi.org/10.3390/ijms22105314
le Roux M-SL, Burger NFV, Vlok M, Kunert KJ, Cullis CA, Botha A-M. EMS Derived Wheat Mutant BIG8-1 (Triticum aestivum L.)—A New Drought Tolerant Mutant Wheat Line. International Journal of Molecular Sciences. 2021; 22(10):5314. https://doi.org/10.3390/ijms22105314
Chicago/Turabian Stylele Roux, Marlon-Schylor L., Nicolas Francois V. Burger, Maré Vlok, Karl J. Kunert, Christopher A. Cullis, and Anna-Maria Botha. 2021. "EMS Derived Wheat Mutant BIG8-1 (Triticum aestivum L.)—A New Drought Tolerant Mutant Wheat Line" International Journal of Molecular Sciences 22, no. 10: 5314. https://doi.org/10.3390/ijms22105314
APA Stylele Roux, M. -S. L., Burger, N. F. V., Vlok, M., Kunert, K. J., Cullis, C. A., & Botha, A. -M. (2021). EMS Derived Wheat Mutant BIG8-1 (Triticum aestivum L.)—A New Drought Tolerant Mutant Wheat Line. International Journal of Molecular Sciences, 22(10), 5314. https://doi.org/10.3390/ijms22105314