
Virtual Screening of FDA-Approved Drugs against Triose Phosphate Isomerase from Entamoeba histolytica and Giardia lamblia Identifies Inhibitors of Their Trophozoite Growth Phase

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
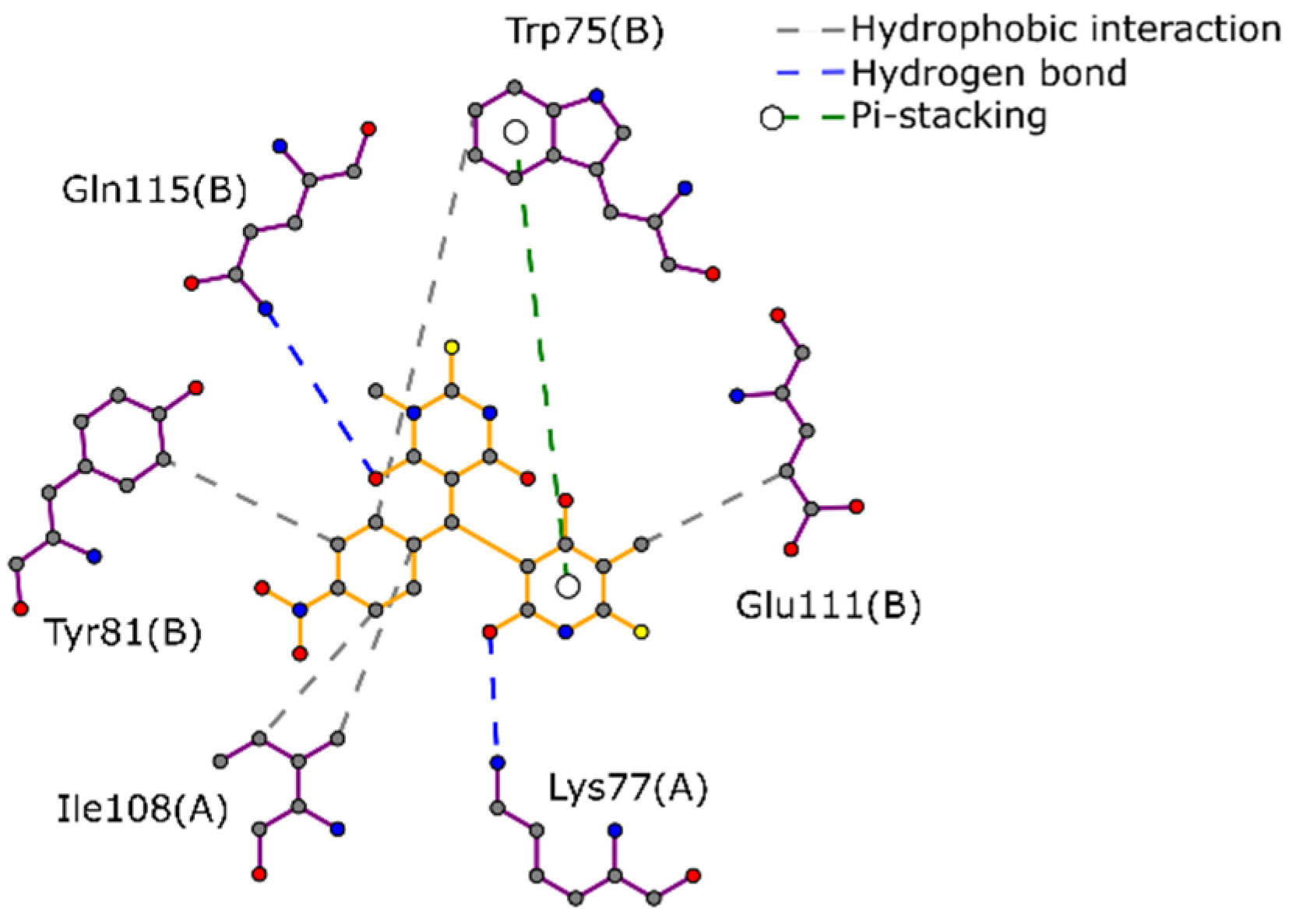
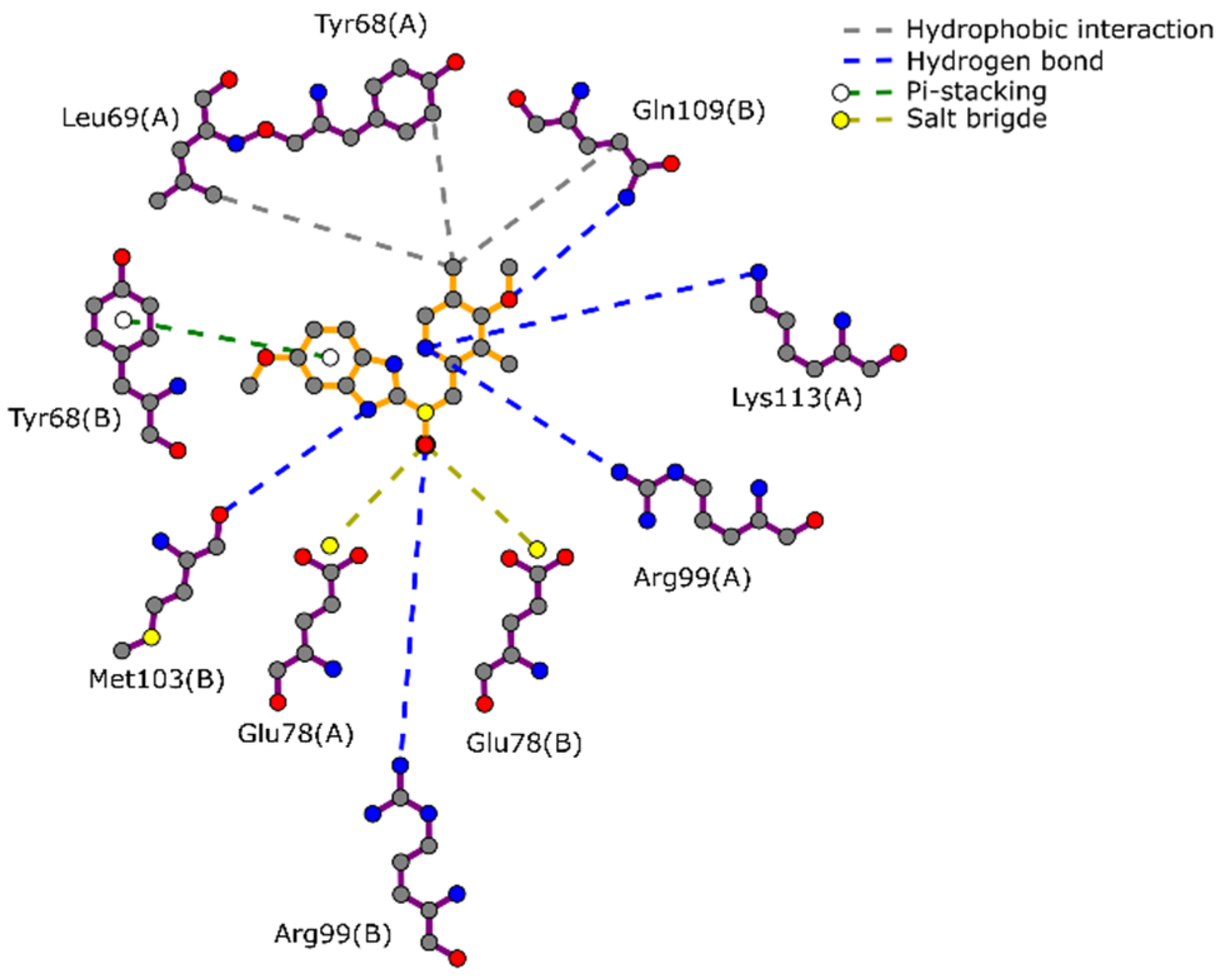
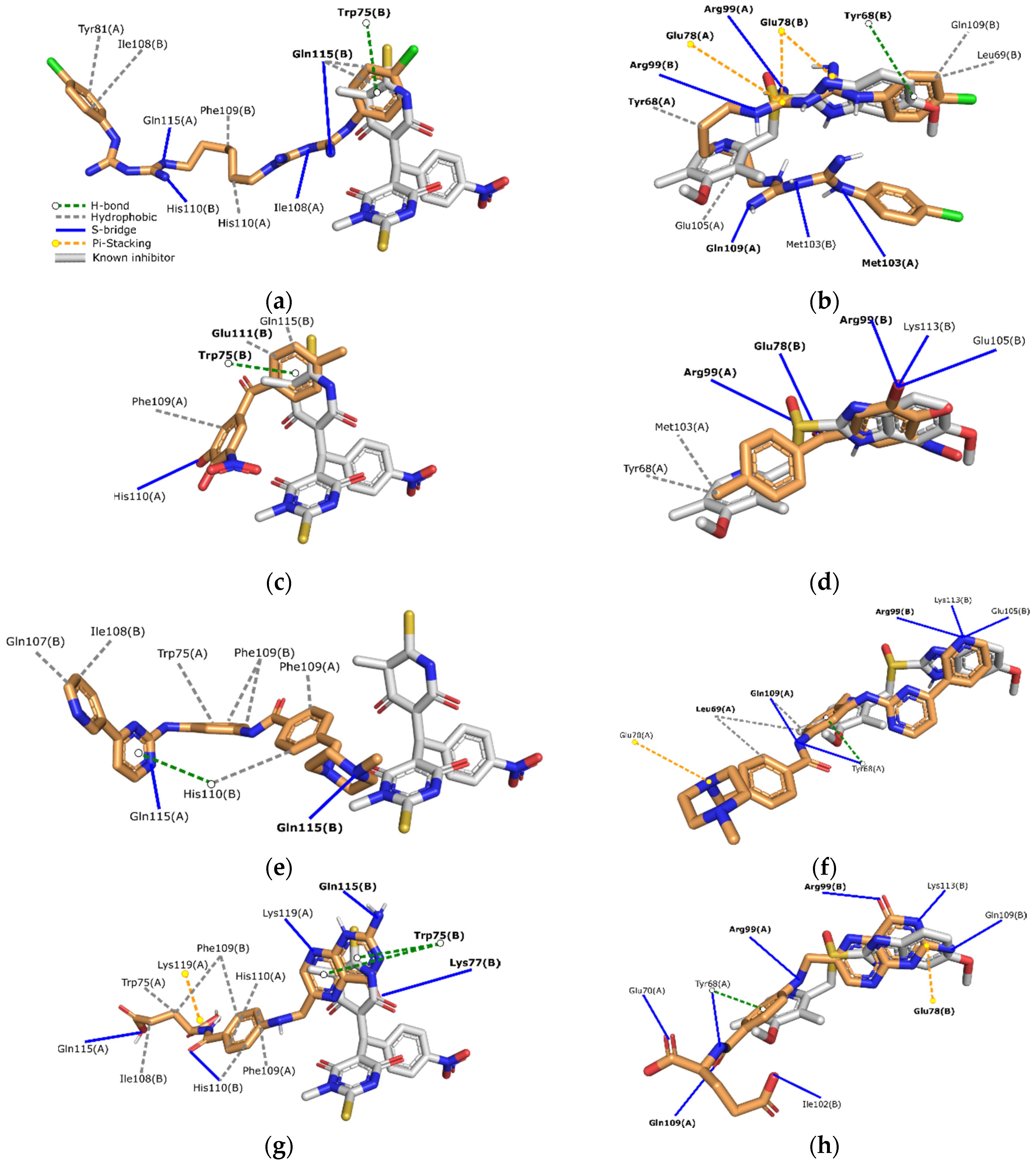
2.1. Docking-Based Virtual Screening on TIMEh and TIMGl
2.2. In Vitro Activity
3. Materials and Methods
3.1. Docking-Based Virtual Screening
3.2. In Vitro Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kantor, M.; Abrantes, A.; Estevez, A.; Schiller, A.; Torrent, J.; Gascon, J.; Hernandez, R.; Ochner, C. Entamoeba histolytica: Updates in Clinical Manifestation, Pathogenesis, and Vaccine Development. Can. J. Gastroenterol. Hepatol. 2018, 2018, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, A.; Austin, R.L. Entamoeba histolytica. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Ryan, U.; Hijjawi, N.; Feng, Y.; Xiao, L. Giardia: An under-reported foodborne parasite. Int. J. Parasitol. 2019, 49, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Castillo, M.; Pacheco-Yepez, J.; Flores-Huerta, N.; Guzmán-Téllez, P.; Jarillo-Luna, R.A.; Cárdenas-Jaramillo, L.M.; Campos-Rodríguez, R.; Shibayama, M. Flavonoids as a Natural Treatment Against Entamoeba histolytica. Front. Cell. Infect. Microbiol. 2018, 8, 209. [Google Scholar] [CrossRef] [PubMed]
- Tharmaratnam, T.; Kumanan, T.; Iskandar, M.A.; D’Urzo, K.; Gopee-Ramanan, P.; Loganathan, M.; Tabobondung, T.; Tabobondung, T.A.; Sivagurunathan, S.; Patel, M.; et al. Entamoeba histolytica and amoebic liver abscess in northern Sri Lanka: A public health problem. Trop. Med. Health 2020, 48, 2–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooshyar, H.; Rostamkhani, P.; Arbabi, M.; Delavari, M. Giardia lamblia infection: Review of current diagnostic strategies. Gastroenterol. Hepatol. Bed Bench 2019, 12, 3–12. [Google Scholar] [PubMed]
- Rumsey, P.; Waseem, M. Giardia Lamblia Enteritis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Hemphill, A.; Müller, N.; Müller, J. Comparative Pathobiology of the Intestinal Protozoan Parasites Giardia lamblia, Entamoeba histolytica, and Cryptosporidium parvum. Pathogens 2019, 8, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatei, W.; Wamae, C.N.; Mbae, C.; Waruru, A.; Mulinge, E.; Waithera, T.; Gatika, S.M.; Kamwati, S.K.; Revathi, G.; Hart, C.A. Cryptosporidiosis: Prevalence, genotype analysis, and symptoms associated with infections in children in Kenya. Am. J. Trop. Med. Hyg. 2006, 75, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbinger, K.-H.; Fleischmann, E.; Weber, C.; Perona, P.; Loscher, T.; Bretzel, G. Epidemiological, clinical, and diagnostic data on intestinal infections with Entamoeba histolytica and Entamoeba dispar among returning travelers. Infection 2011, 39, 527–535. [Google Scholar] [CrossRef]
- Ashburn, T.T.; Thor, K.B. Drug repositioning: Identifying and developing new uses for existing drugs. Nat. Rev. Drug Discov. 2004, 3, 673–683. [Google Scholar] [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Adasme, M.F.; Parisi, D.; Sveshnikova, A.; Schroeder, M. Structure-based drug repositioning: Potential and limits. Semin. Cancer Biol. 2021, 68, 192–198. [Google Scholar] [CrossRef]
- Juárez-Saldivar, A.; Schroeder, M.; Salentin, S.; Haupt, J.; Saavedra, E.; Vázquez, C.; Reyes-Espinosa, F.; Herrera-Mayorga, V.; Villalobos-Rocha, J.C.; García-Pérez, C.A.; et al. Computational Drug Repositioning for Chagas Disease Using Protein-Ligand Interaction Profiling. Int. J. Mol. Sci. 2020, 21, 4270. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhu, F.; Ma, X.H.; Shi, Z.; Yang, S.Y.; Wei, Y.Q.; Chen, Y.Z. Predicting targeted polypharmacology for drug repositioning and multi-target drug discovery. Curr. Med. Chem. 2013, 20, 1646–1661. [Google Scholar] [CrossRef] [PubMed]
- Trosset, J.-Y.; Cavé, C. In Silico Drug–Target Profiling. Methods Mol. Biol. 2019, 1953, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Jiménez, L.K.; Paz-González, A.D.; Juárez-Saldivar, A.; Uhrig, M.L.; Agusti, R.; Reyes-Arellano, A.; Nogueda-Torres, B.; Rivera, G. Structure-Based Virtual Screening of New Benzoic Acid Derivatives as Trypanosoma cruzi Trans-sialidase Inhibitors. Med. Chem. 2020, 16, 1–9. [Google Scholar] [CrossRef]
- Delgado-Maldonado, T.; Nogueda-Torres, B.; Espinoza-Hicks, J.C.; Vázquez-Jiménez, L.K.; Paz-González, A.D.; Juárez-Saldívar, A.; Rivera, G. Synthesis and biological evaluation in vitro and in silico of N-propionyl-N′-benzeneacylhydrazone derivatives as cruzain inhibitors of Trypanosoma cruzi. Mol. Divers. 2020, 24, 1–12. [Google Scholar] [CrossRef]
- García-Torres, I.; Mora, I.D.L.M.-D.L.; Marcial-Quino, J.; Gómez-Manzo, S.; Vanoye-Carlo, A.; Navarrete-Vázquez, G.; Colín-Lozano, B.; Gutiérrez-Castrellón, P.; Sierra-Palacios, E.; López-Velázquez, G.; et al. Proton pump inhibitors drastically modify triosephosphate isomerase from Giardia lamblia at functional and structural levels, providing molecular leads in the design of new antigiardiasic drugs. Biochim. Biophys. Acta 2016, 1860, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Leitsch, D.; Müller, J.; Müller, N. Evaluation of Giardia lamblia thioredoxin reductase as drug activating enzyme and as drug target. Int. J. Parasitol. Drugs Drug Resist. 2016, 6, 148–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Herrera, A.; Cortez-Maya, S.; Bocanegra-Garcia, V.; Banik, B.K.; Rivera, G. Recent Advances in the Development of Broad-Spectrum Antiprotozoal Agents. Curr. Med. Chem. 2021, 28, 583–606. [Google Scholar] [CrossRef] [PubMed]
- Nurkanto, A.; Jeelani, G.; Yamamoto, T.; Naito, Y.; Hishiki, T.; Mori, M.; Suematsu, M.; Shiomi, K.; Hashimoto, T.; Nozaki, T. Characterization and validation of Entamoeba histolytica pantothenate kinase as a novel anti-amebic drug target. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Tarique, K.F.; Devi, S.; Tomar, P.; Ali, M.F.; Rehman, S.A.A.; Gourinath, S. Characterization and functional insights into the Entamoeba histolytica pyridoxal kinase, an enzyme essential for its survival. J. Struct. Biol. 2020, 212, 107645. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, M.; Balaram, H.; Balaram, P. Structural effects of a dimer interface mutation on catalytic activity of triosephosphate isomerase. The role of conserved residues and complementary mutations. FEBS J. 2009, 276, 4169–4183. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Illana, V.; Perez-Montfort, R.; López-Calahorra, F.; Costas, M.; Rodríguez-Romero, A.; de Gómez-Puyou, M.T.; Puyou, A.G. Structural Differences in Triosephosphate Isomerase from Different Species and Discovery of a Multitrypanosomatid Inhibitor. Biochemistry 2006, 45, 2556–2560. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Romero, A.; Hernández-Santoyo, A.; del Pozo-Yauner, L.; Kornhauser, A.; Fernández-Velasco, D. Structure and Inactivation of Triosephosphate Isomerase from Entamoeba histolytica. J. Mol. Biol. 2002, 322, 669–675. [Google Scholar] [CrossRef]
- Vique--Sánchez, J.L.; Jiménez--Pineda, A.; Benítez--Cardoza, C.G. Amoebicidal effect of 5,5′--[(4--nitrophenyl)methylene]bis--6--hydroxy--2--mercapto--3--methyl--4(3H)--pyrimidinone), a new drug against Entamoeba histolytica. Arch. Pharm. 2021, 354, e2000263. [Google Scholar] [CrossRef] [PubMed]
- López-Velázquez, G.; Fernández-Lainez, C.; de la Mora, J.I.; de la Portilla, D.C.; Robles, R.R.; González-Maciel, A.; Ridaura, C.; García-Torres, I.; Gutiérrez-Castrellón, P.; Olivos-García, A.; et al. On the molecular and cellular effects of omeprazole to further support its effectiveness as an antigiardial drug. Sci. Rep. 2019, 9, 8922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurkcuoglu, Z.; Findik, D.; Akten, E.D.; Doruker, P. How an Inhibitor Bound to Subunit Interface Alters Triosephosphate Isomerase Dynamics. Biophys. J. 2015, 109, 1169–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejman-Yarden, N.; Miyamoto, Y.; Leitsch, D.; Santini, J.; Debnath, A.; Gut, J.; McKerrow, J.H.; Reed, S.L.; Eckmann, L. A Reprofiled Drug, Auranofin, Is Effective against Metronidazole-Resistant Giardia lamblia. Antimicrob. Agents Chemother. 2013, 57, 2029–2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivares, J.L.; Fernández, R.; Fleta, J.; Ruiz, M.Y.; Clavel, A. Vitamin B12and Folic Acid in Children with Intestinal Parasitic Infection. J. Am. Coll. Nutr. 2002, 21, 109–113. [Google Scholar] [CrossRef]
- Mulenga, M.; Malunga, P.; Bennett, S.; Thuma, P.; Shulman, C.; Fielding, K.; Greenwood, B. Folic acid treatment of Zambian children with moderate to severe malaria anemia. Am. J. Trop. Med. Hyg. 2006, 74, 986–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A Major Update to the DrugBank Database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminf. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein–ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Compound. | IC50 E. histolytica (μg/mL) | IC50 G. lamblia (μg/mL) |
|---|---|---|
| Metronidazole | 0.205 | 7.8 |
| D4 | 8.306 ± 1.616 1 | - |
| Omeprazol | - | 0.025 2 |
| Chlorhexidine | > 100 | 4.93 ± 0.005 |
| Tolcapone | > 100 | 0.05 ± 0.002 |
| Imatinib | > 100 | 3.46 ± 0.005 |
| Folic acid | 0.186 ± 0.003 | 5.34 ± 0.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juárez-Saldivar, A.; Barbosa-Cabrera, E.; Lara-Ramírez, E.E.; Paz-González, A.D.; Martínez-Vázquez, A.V.; Bocanegra-García, V.; Palos, I.; Campillo, N.E.; Rivera, G. Virtual Screening of FDA-Approved Drugs against Triose Phosphate Isomerase from Entamoeba histolytica and Giardia lamblia Identifies Inhibitors of Their Trophozoite Growth Phase. Int. J. Mol. Sci. 2021, 22, 5943. https://doi.org/10.3390/ijms22115943
Juárez-Saldivar A, Barbosa-Cabrera E, Lara-Ramírez EE, Paz-González AD, Martínez-Vázquez AV, Bocanegra-García V, Palos I, Campillo NE, Rivera G. Virtual Screening of FDA-Approved Drugs against Triose Phosphate Isomerase from Entamoeba histolytica and Giardia lamblia Identifies Inhibitors of Their Trophozoite Growth Phase. International Journal of Molecular Sciences. 2021; 22(11):5943. https://doi.org/10.3390/ijms22115943
Chicago/Turabian StyleJuárez-Saldivar, Alfredo, Elizabeth Barbosa-Cabrera, Edgar E. Lara-Ramírez, Alma D. Paz-González, Ana V. Martínez-Vázquez, Virgilio Bocanegra-García, Isidro Palos, Nuria E. Campillo, and Gildardo Rivera. 2021. "Virtual Screening of FDA-Approved Drugs against Triose Phosphate Isomerase from Entamoeba histolytica and Giardia lamblia Identifies Inhibitors of Their Trophozoite Growth Phase" International Journal of Molecular Sciences 22, no. 11: 5943. https://doi.org/10.3390/ijms22115943
APA StyleJuárez-Saldivar, A., Barbosa-Cabrera, E., Lara-Ramírez, E. E., Paz-González, A. D., Martínez-Vázquez, A. V., Bocanegra-García, V., Palos, I., Campillo, N. E., & Rivera, G. (2021). Virtual Screening of FDA-Approved Drugs against Triose Phosphate Isomerase from Entamoeba histolytica and Giardia lamblia Identifies Inhibitors of Their Trophozoite Growth Phase. International Journal of Molecular Sciences, 22(11), 5943. https://doi.org/10.3390/ijms22115943