
Altered Expression of ESR1, ESR2, PELP1 and c-SRC Genes Is Associated with Ovarian Cancer Manifestation


 , , ,
, , ,
Abstract
:1. Introduction
2. Results
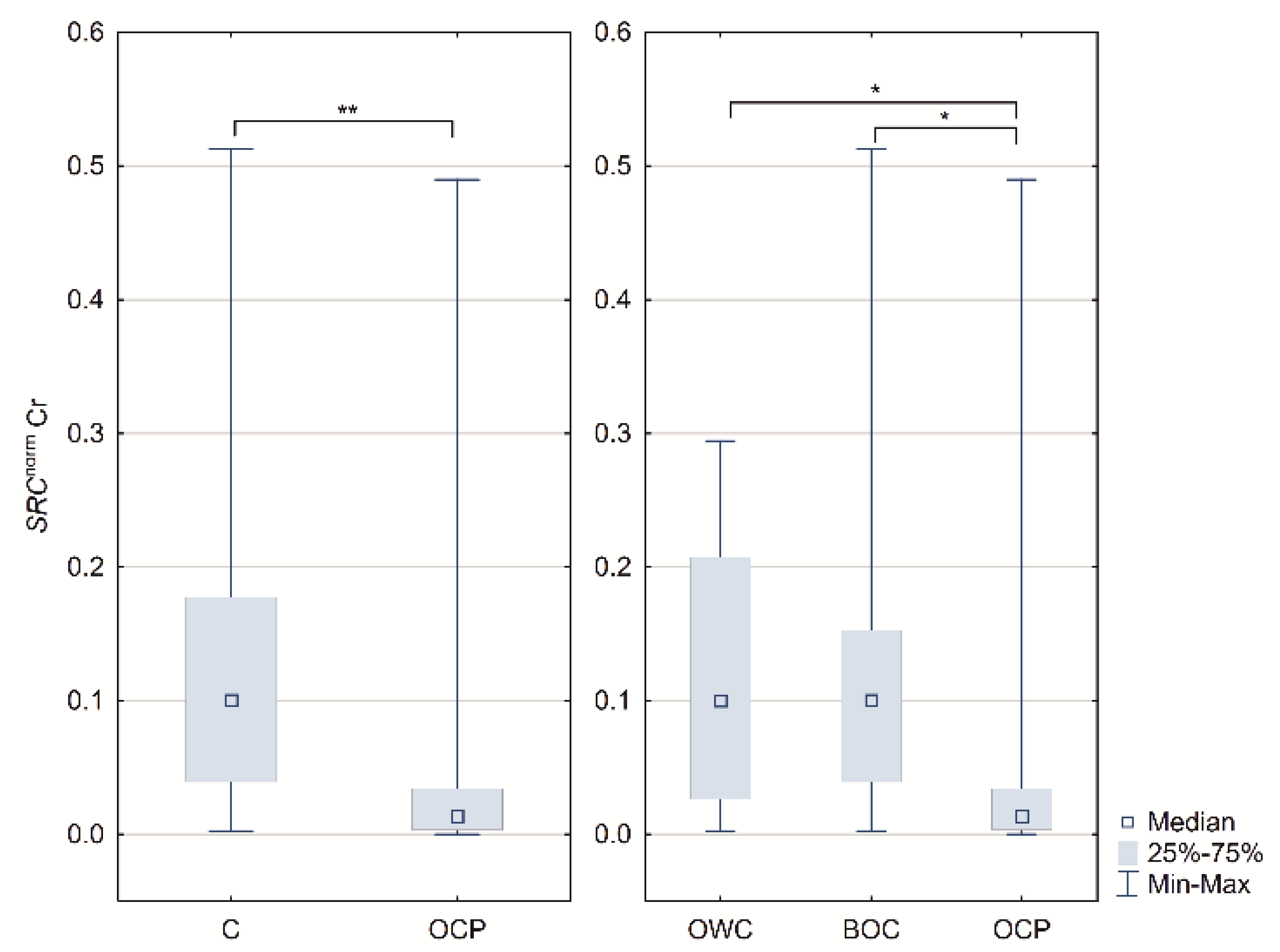
2.1. ESR1, ESR2 PELP1 and SRC Relative Expression
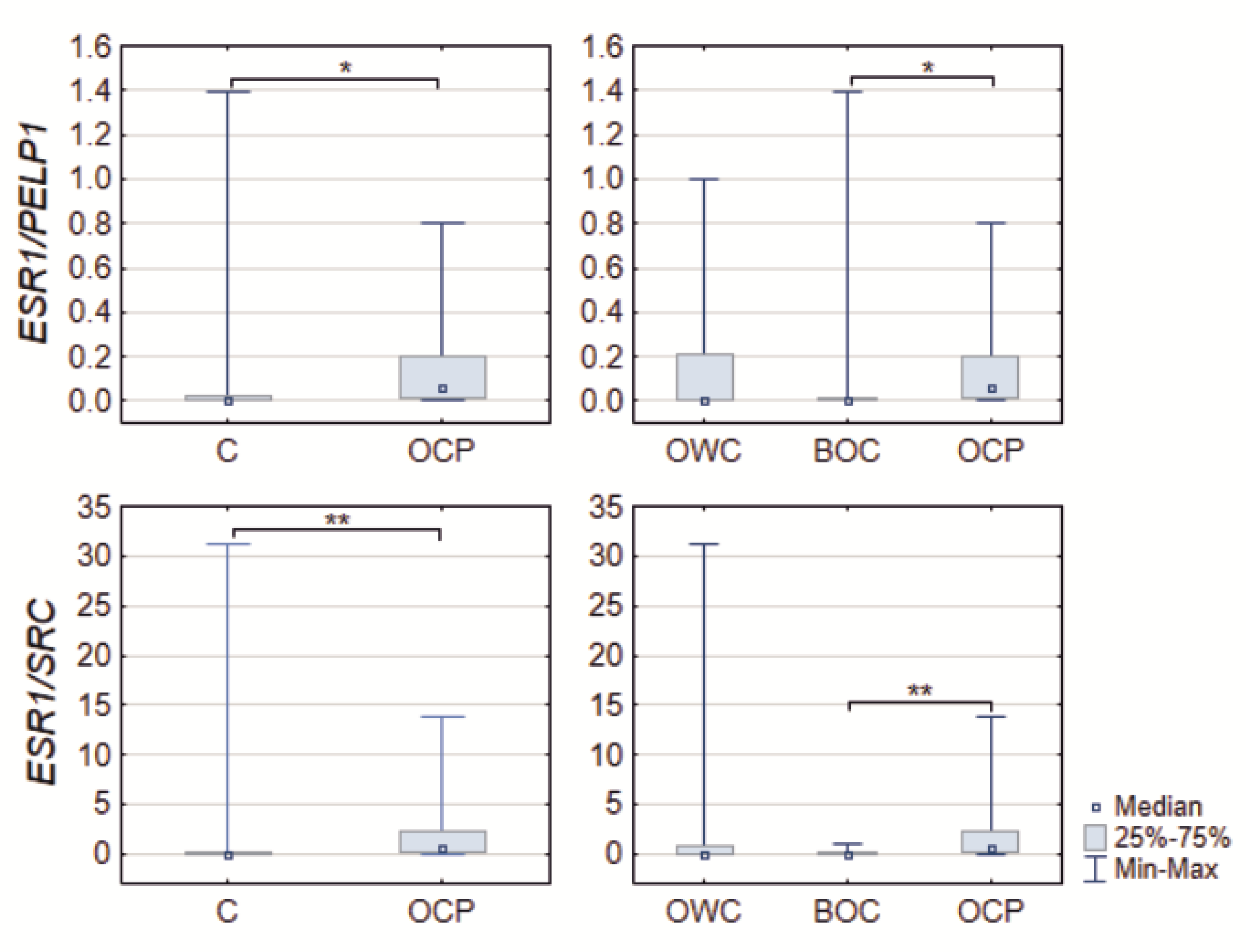
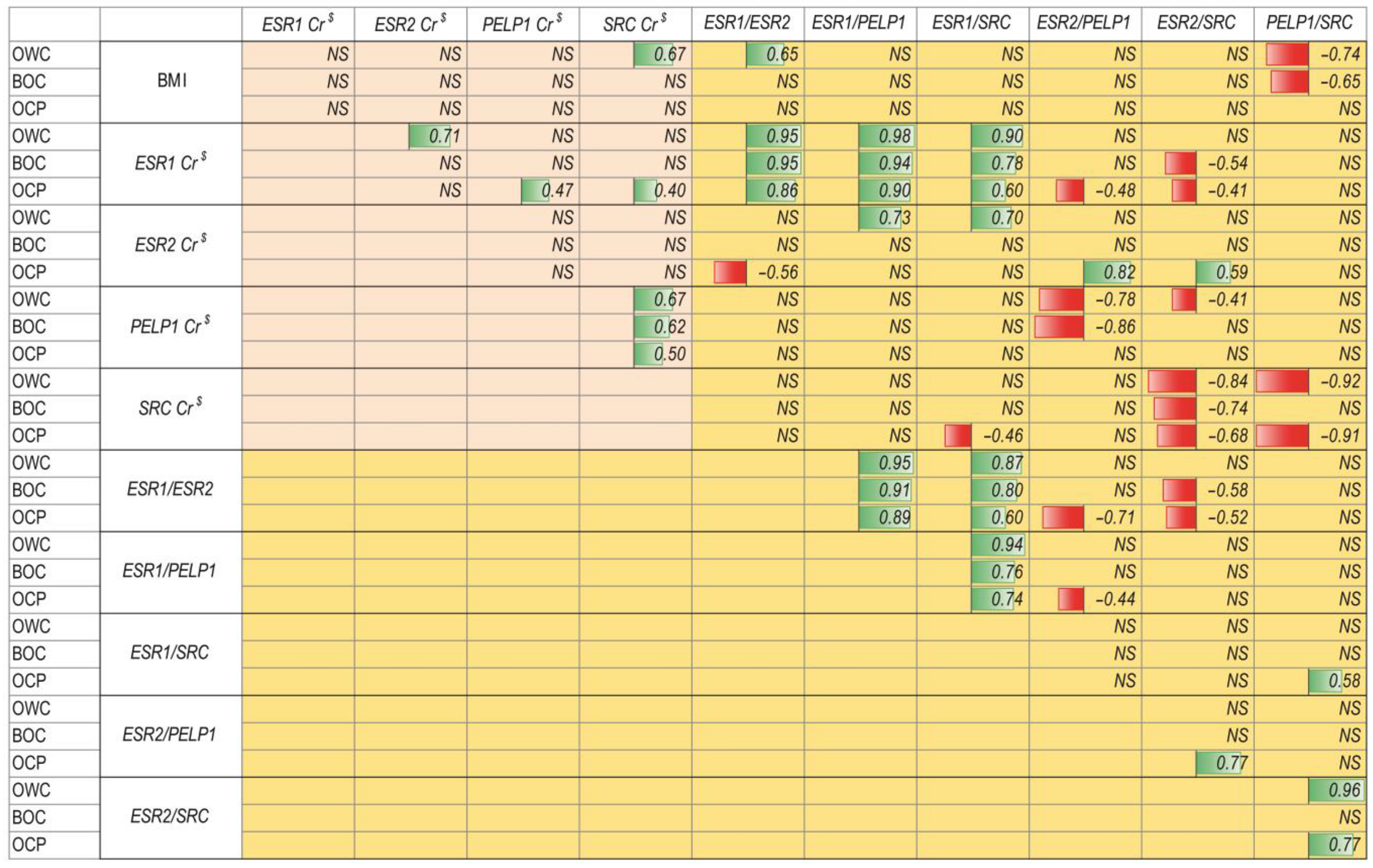
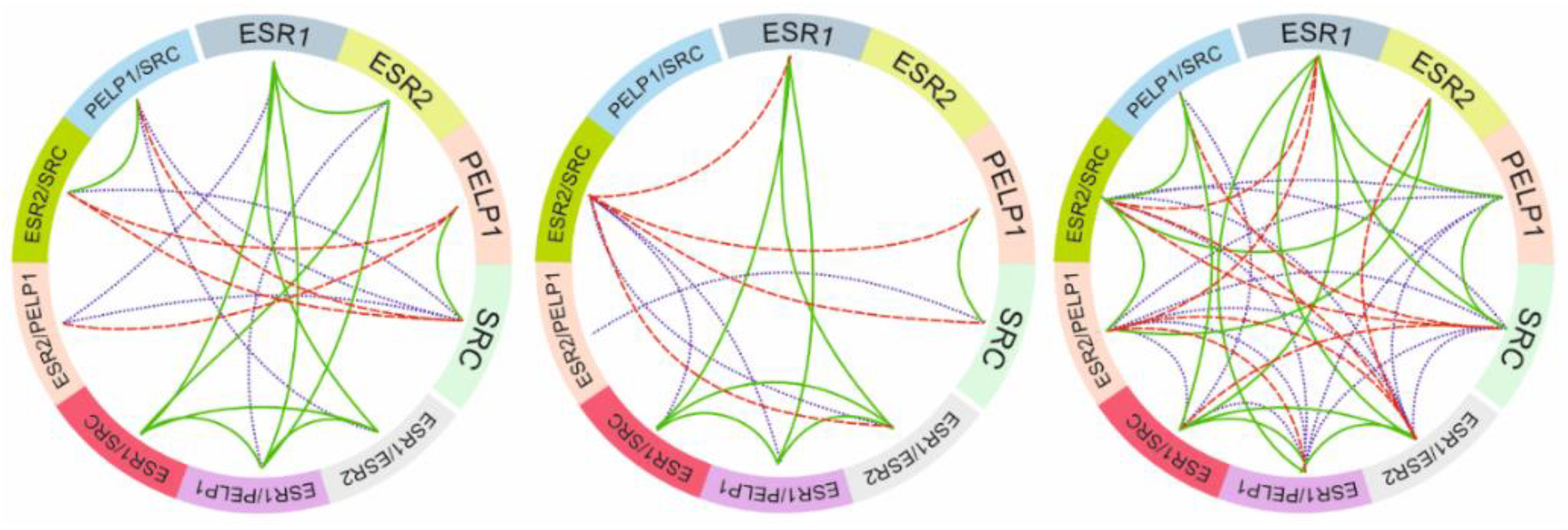
2.2. Relationship between ER Signaling and Ovarian Cancer
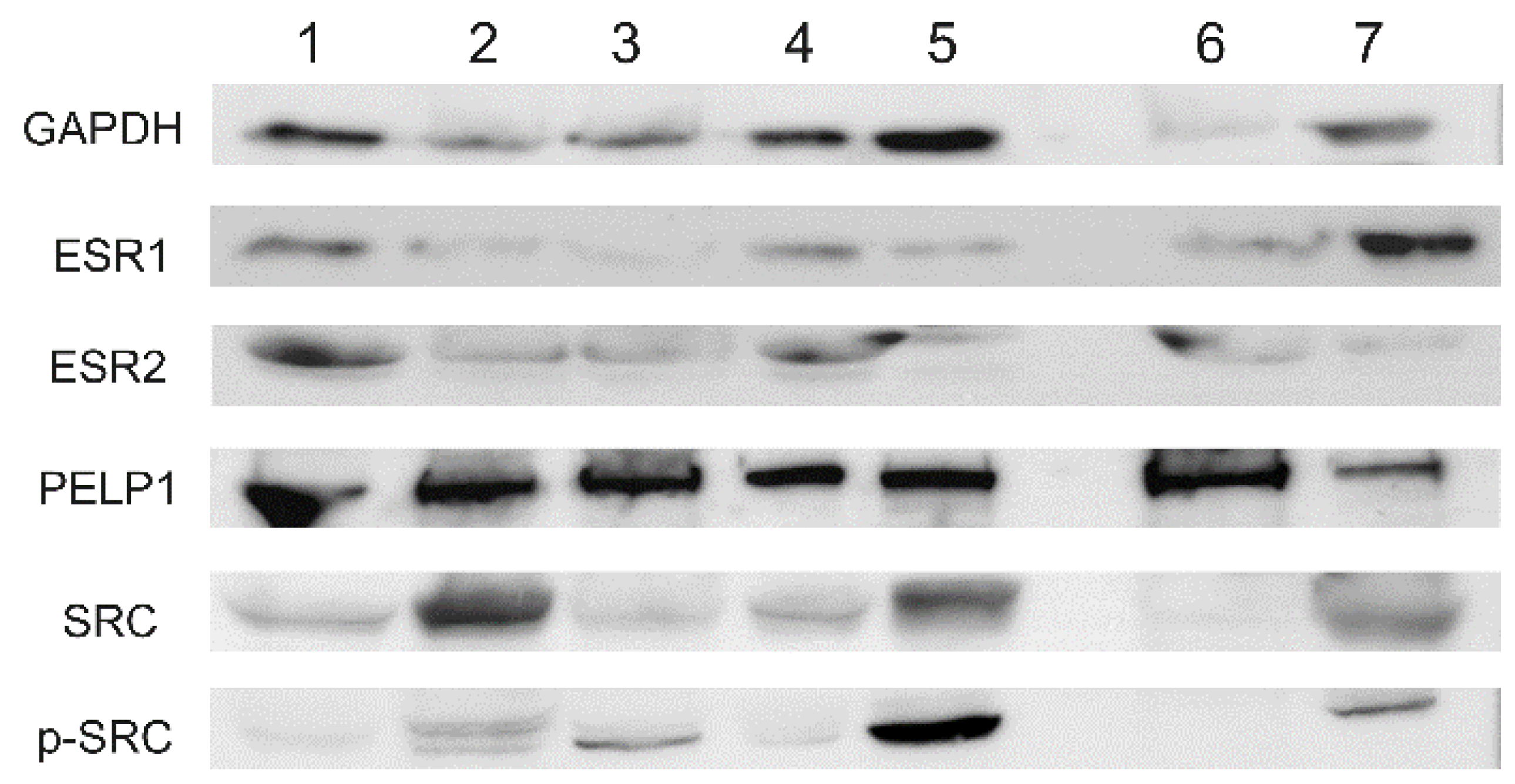
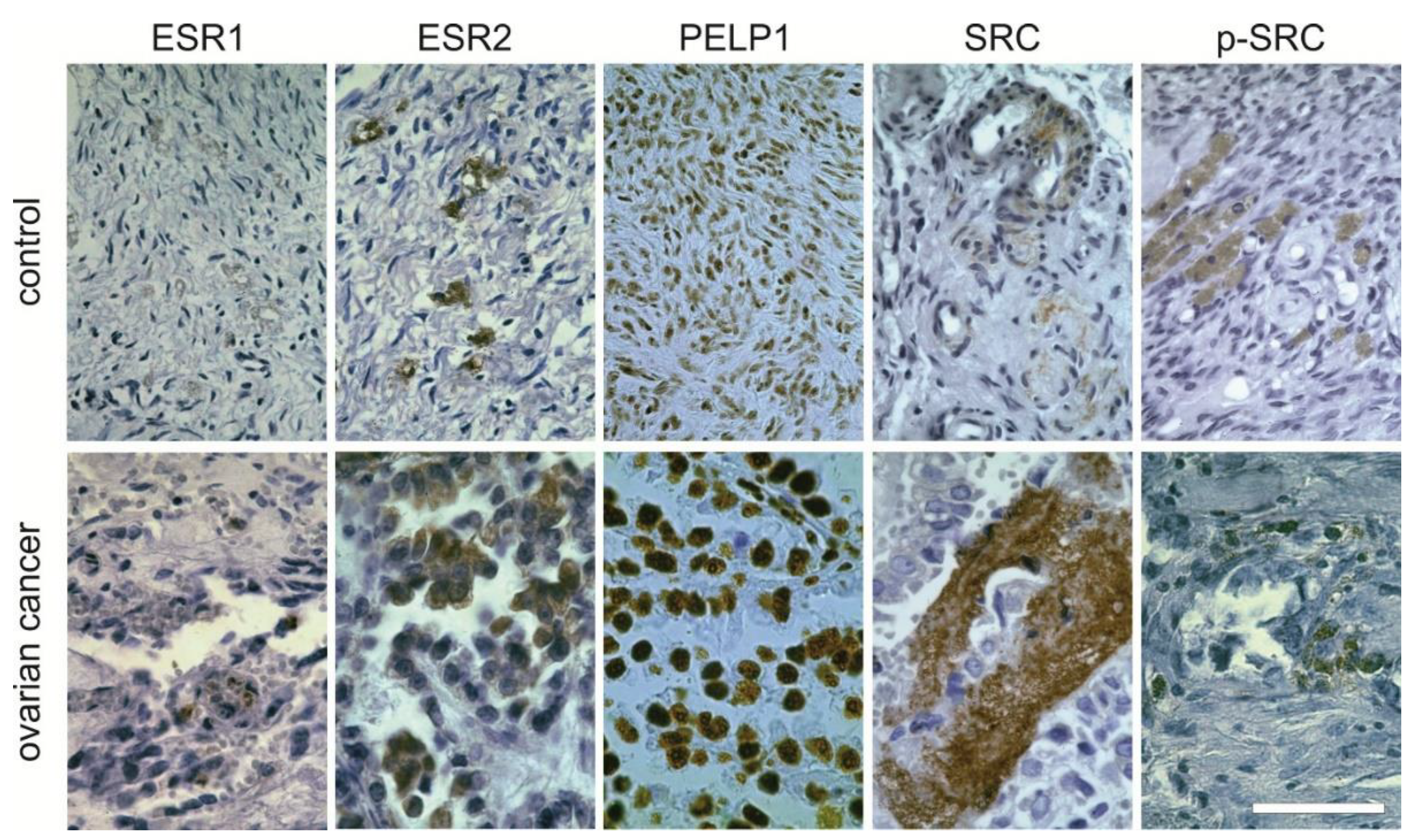
2.3. Protein Expression
3. Discussion
4. Materials and Methods
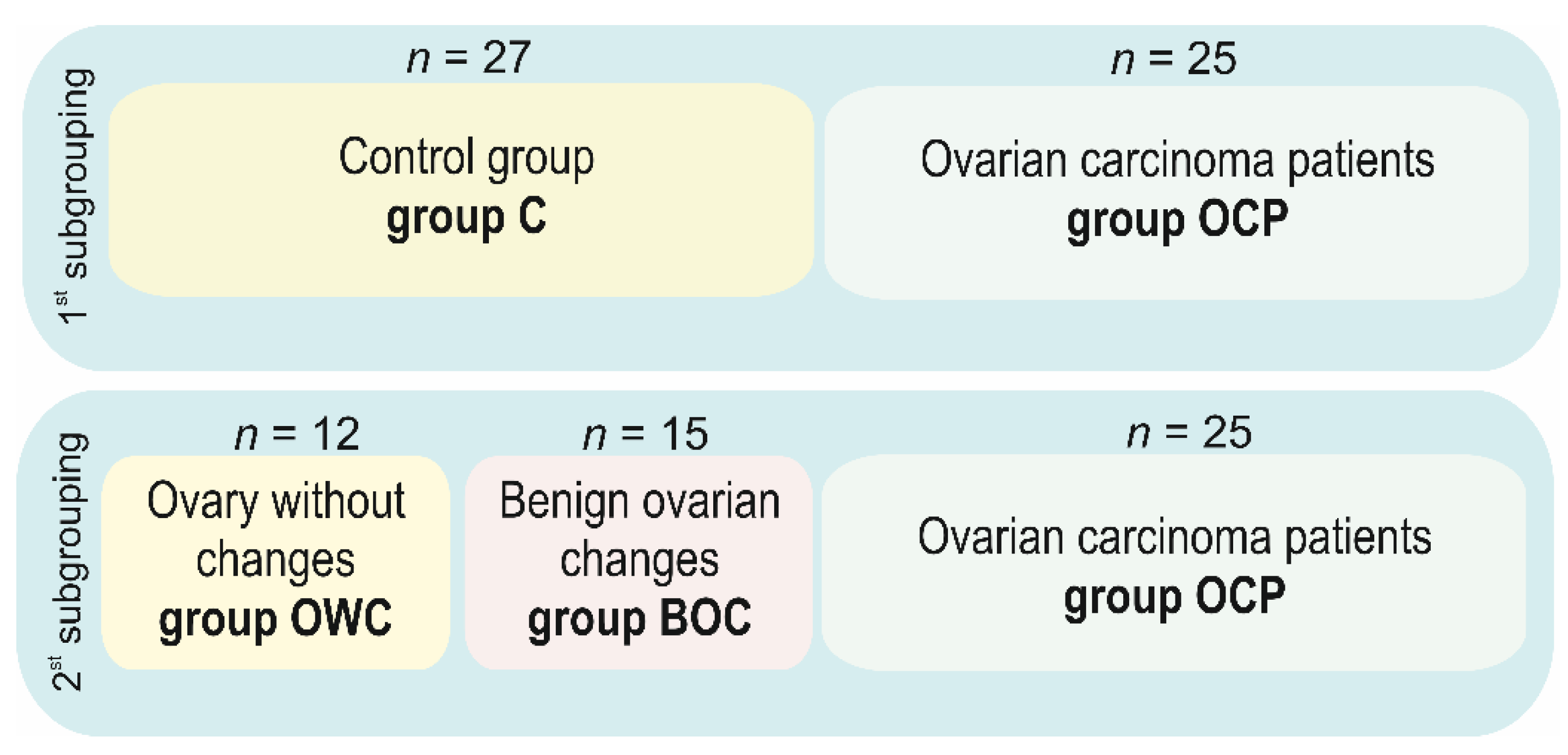
4.1. Patients
4.1.1. Clinical Markers Data Evaluation
4.1.2. FIGO Staging and Grading System
4.1.3. Histopatologic Classification
4.2. Nucleic Acid Extraction and Validation
4.3. Reverse Transcription and Quantitative PCR
4.3.1. RNA Extraction and Validation
4.3.2. Real Time Polymerase Chain Reaction
4.4. Protein Expression
4.4.1. Western Blot
4.4.2. Immunohistochemistry
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reid, B.M.; Permuth, J.B.; Sellers, T.A. Epidemiology of ovarian cancer: A review. Cancer Biol. Med. 2017, 14, 9–32. [Google Scholar]
- Boussios, S.; Karihtala, P.; Moschetta, M.; Abson, C.; Karathanasi, A.; Zakynthinakis-Kyriakou, N.; Ryan, J.E.; Sheriff, M.; Rassy, E.; Pavlidis, N. Veliparib in ovarian cancer: A new synthetically lethal therapeutic approach. Investig. New Drugs 2020, 38, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Dochez, V.; Caillon, H.; Vaucel, E.; Dimet, J.; Winer, N.; Ducarme, G. Biomarkers and algorithms for diagnosis of ovarian cancer: CA125, HE4, RMI and ROMA, a review. J. Ovarian Res. 2019, 12, 28. [Google Scholar] [CrossRef] [Green Version]
- Jelovac, D.; Armstrong, D.K. Recent progress in the diagnosis and treatment of ovarian cancer. CA Cancer J. Clin. 2011, 61, 183–203. [Google Scholar] [CrossRef]
- Modugno, F.; Laskey, R.; Smith, A.L.; Andersen, C.L.; Haluska, P.; Oesterreich, S. Hormone response in ovarian cancer: Time to reconsider as a clinical target? Endocr. Relat. Cancer 2012, 19, R255–R279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schüler, S.; Ponnath, M.; Engel, J.; Ortmann, O. Ovarian epithelial tumors and reproductive factors: A systematic review. Arch. Gynecol. Obstet. 2013, 287, 1187–1204. [Google Scholar] [CrossRef]
- Labrie, F. All sex steroids are made intracellularly in peripheral tissues by the mechanisms of intracrinology after menopause. J. Steroid Biochem. Mol. Biol. 2015, 145, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Folkerd, E.J.; Dowsett, M. Influence of sex hormones on cancer progression. J. Clin. Oncol. 2010, 28, 4038–4044. [Google Scholar] [CrossRef] [PubMed]
- Mungenast, F.; Thalhammer, T. Estrogen biosynthesis and action in ovarian cancer. Front. Endocrinol. 2014, 5, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadlamudi, R.K.; Rajhans, R.; Chakravarty, D.; Nair, B.C.; Nair, S.S.; Evans, D.B.; Chen, S.; Tekmal, R.R. Regulation of aromatase induction by nuclear receptor coregulator PELP1. J. Steroid Biochem. Mol. Biol. 2010, 118, 211–218. [Google Scholar]
- Sareddy, G.R.; Vadlamudi, R.K. PELP1: Structure, biological function and clinical significance. Gene 2016, 585, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Thakkar, R.; Sareddy, G.R.; Zhang, Q.; Wang, R.; Vadlamudi, R.K.; Brann, D. PELP1: A key mediator of oestrogen signalling and actions in the brain. J. Neuroendocrinol. 2018, 30, e12484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.K.; Balasenthil, S.; Nguyen, D.; Vadlamudi, R.K. Cloning and functional characterization of PELP1/MNAR promoter. Gene 2004, 330, 115–122. [Google Scholar] [CrossRef]
- Rayala, S.K.; den Hollander, P.; Balasenthil, S.; Molli, P.R.; Bean, A.J.; Vadlamudi, R.K.; Wang, R.-A.; Kumar, R. Hepatocyte growth factor-regulated tyrosine kinase substrate (HRS) interacts with PELP1 and activates MAPK. J. Biol. Chem. 2006, 281, 4395–4403. [Google Scholar] [CrossRef] [Green Version]
- Samartzis, E.P.; Labidi-Galy, S.I.; Moschetta, M.; Uccello, M.; Kalaitzopoulos, D.R.; Perez-Fidalgo, J.A.; Boussios, S. Endometriosis-associated ovarian carcinomas: Insights into pathogenesis, diagnostics, and therapeutic targets—A narrative review. Ann. Transl. Med. 2020, 8, 1712. [Google Scholar] [CrossRef]
- Vadlamudi, R.K.; Manavathi, B.; Balasenthil, S.; Nair, S.S.; Yang, Z.; Sahin, A.A.; Kumar, R. Functional implications of altered subcellular localization of PELP1 in breast cancer cells. Cancer Res. 2005, 65, 7724–7732. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.W.; McNally, C.; Nickbarg, E.; Komm, B.S.; Cheskis, B.J. Estrogen receptor-interacting protein that modulates its nongenomic activity-crosstalk with Src/Erk phosphorylation cascade. Proc. Natl. Acad. Sci. USA 2002, 99, 14783–14788. [Google Scholar] [CrossRef] [Green Version]
- Dimple, C.; Nair, S.S.; Rajhans, R.; Pitcheswara, P.R.; Liu, J.; Balasenthil, S.; Le, X.-F.; Burow, M.E.; Auersperg, N.; Tekmal, R.R.; et al. Role of PELP1/MNAR signaling in ovarian tumorigenesis. Cancer Res. 2008, 68, 4902–4909. [Google Scholar] [CrossRef] [Green Version]
- Manavathi, B.; Nair, S.S.; Wang, R.-A.; Kumar, R.; Vadlamudi, R.K. Proline-, glutamic acid-, and leucine-rich protein-1 is essential in growth factor regulation of signal transducers and activators of transcription 3 activation. Cancer Res. 2005, 65, 5571–5577. [Google Scholar] [CrossRef] [Green Version]
- Chakravarty, D.; Nair, S.S.; Santhamma, B.; Nair, B.C.; Wang, L.; Bandyopadhyay, A.; Agyin, J.K.; Brann, D.; Sun, L.Z.; Yeh, I.T.; et al. Extranuclear functions of ER impact invasive migration and metastasis by breast cancer cells. Cancer Res. 2010, 70, 4092–4101. [Google Scholar] [CrossRef] [Green Version]
- Rajhans, R.; Nair, S.; Holden, A.H.; Kumar, R.; Tekmal, R.R.; Vadlamudi, R.K. Oncogenic potential of the nuclear receptor coregulator proline-, glutamic acid-, leucine-rich protein 1/modulator of the nongenomic actions of the estrogen receptor. Cancer Res. 2007, 67, 5505–5512. [Google Scholar] [CrossRef] [Green Version]
- Gonugunta, V.K.; Sareddy, G.R.; Krishnan, S.R.; Cortez, V.; Roy, S.S.; Tekmal, R.R.; Vadlamudi, R.K. Inhibition of mTOR signaling reduces PELP1-mediated tumor growth and therapy resistance. Mol. Cancer Ther. 2014, 13, 1578–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallabhaneni, S.; Nair, B.C.; Cortez, V.; Challa, R.; Chakravarty, D.; Tekmal, R.R.; Vadlamudi, R.K. Significance of ER-Src axis in hormonal therapy resistance. Breast Cancer Res. Treat. 2011, 130, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Chakravarty, D.; Roy, S.S.; Babu, C.R.; Dandamudi, R.; Curiel, T.J.; Vivas-Mejia, P.; Lopez-Berestein, G.; Sood, A.K.; Vadlamudi, R.K. Therapeutic targeting of PELP1 prevents ovarian cancer growth and metastasis. Clin. Cancer Res. 2011, 17, 2250–2259. [Google Scholar] [CrossRef] [Green Version]
- Nair, B.C.; Vallabhaneni, S.; Tekmal, R.R.; Vadlamudi, R.K. Roscovitine confers tumor suppressive effect on therapy-resistant breast tumor cells. Breast Cancer Res. 2011, 13, R80. [Google Scholar] [CrossRef] [Green Version]
- Gregorio, K.C.R.; Laurindo, C.P.; Machado, U.F. Estrogen and Glycemic Homeostasis: The Fundamental Role of Nuclear Estrogen Receptors ESR1/ESR2 in Glucose Transporter GLUT4 Regulation. Cells 2021, 10, 99. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, P.M.; Jordan, S.J. Epidemiology of epithelial ovarian cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 41, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Kim, Y.-H.; Kim, Y.B.; Kim, J.; Kim, J.-W.; Park, M.H.; Park, J.H.; Rhee, J.H.; Lim, M.C.; Hong, J.-S. Bilateral salpingectomy to reduce the risk of ovarian/fallopian/peritoneal cancer in women at average risk: A position statement of the Korean Society of Obstetrics and Gynecology (KSOG). Obstet. Gynecol. Sci. 2018, 61, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Stratton, J.F.; Pharoah, P.; Smith, S.K.; Easton, D.; Ponder, B.A. A systematic review and meta-analysis of family history and risk of ovarian cancer. Br. J. Obstet. Gynaecol. 1998, 105, 493–499. [Google Scholar] [CrossRef]
- Jervis, S.; Song, H.; Lee, A.; Dicks, E.; Tyrer, J.; Harrington, P.; Easton, D.F.; Jacobs, I.J.; Pharoah, P.P.D.; Antoniou, A.C. Ovarian cancer familial relative risks by tumour subtypes and by known ovarian cancer genetic susceptibility variants. J. Med. Genet. 2014, 51, 108–113. [Google Scholar] [CrossRef]
- Boussios, S.; Mikropoulos, C.; Samartzis, E.; Karihtala, P.; Moschetta, M.; Sheriff, M.; Karathanasi, A.; Sadauskaite, A.; Rassy, E.; Pavlidis, N. Wise management of ovarian cancer: On the cutting edge. J. Pers. Med. 2020, 10, 41. [Google Scholar] [CrossRef]
- Arruda, J. Salpingectomy to reduce risk of ovarian cancer in women at average risk. Menopause 2019, 26, 1059–1061. [Google Scholar] [CrossRef]
- La Vecchia, C. Ovarian cancer: Epidemiology and risk factors. Eur. J. Cancer Prev. 2017, 26, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.K.L.; Siu, M.K.Y.; Jiang, Y.-X.; Wang, J.-J.; Wang, Y.; Leung, T.H.Y.; Liu, S.S.; Cheung, A.N.Y.; Ngan, H.Y.S. Differential expression of estrogen receptor subtypes and variants in ovarian cancer: Effects on cell invasion, proliferation and prognosis. BMC Cancer 2017, 17, 606. [Google Scholar] [CrossRef]
- Aust, S.; Horak, P.; Pils, D.; Pils, S.; Grimm, C.; Horvat, R.; Tong, D.; Schmid, B.; Speiser, P.; Reinthaller, A.; et al. The prognostic value of estrogen receptor beta and proline-, glutamic acid- and leucine-rich protein 1 (PELP1) expression in ovarian cancer. BMC Cancer 2013, 13, 115. [Google Scholar] [CrossRef] [Green Version]
- Grivas, P.D.; Tzelepi, V.; Sotiropoulou-Bonikou, G.; Kefalopoulou, Z.; Papavassiliou, A.G.; Kalofonos, H. Expression of ERalpha, ERbeta and co-regulator PELP1/MNAR in colorectal cancer: Prognostic significance and clinicopathologic correlations. Cell. Oncol. 2009, 31, 235–247. [Google Scholar]
- Biscardi, J.S.; Tice, D.A.; Parsons, S.J. c-Src, receptor tyrosine kinases, and human cancer. Adv. Cancer Res. 1999, 76, 61–119. [Google Scholar]
- Irby, R.B.; Yeatman, T.J. Role of Src expression and activation in human cancer. Oncogene 2000, 19, 5636–5642. [Google Scholar] [CrossRef] [Green Version]
- Camacho-Sanchez, M.; Burraco, P.; Gomez-Mestre, I.; Leonard, J.A. Preservation of RNA and DNA from mammal samples under field conditions. Mol. Ecol. Resour. 2013, 13, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Andrusiewicz, M.; Słowikowski, B.; Skibińska, I.; Wołuń-Cholewa, M.; Dera-Szymanowska, A. Selection of reliable reference genes in eutopic and ectopic endometrium for quantitative expression studies. Biomed. Pharmacother. 2016, 78, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Waligórska-Stachura, J.; Sawicka-Gutaj, N.; Zabel, M.; Andrusiewicz, M.; Gut, P.; Czarnywojtek, A.; Ruchała, M. Survivin DEx3 as a biomarker of thyroid cancers: A study at the mRNA and protein level. Oncol. Lett. 2017, 13, 2437–2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | 1st Subgrouping | 2nd Subgrouping | |||||
|---|---|---|---|---|---|---|---|
| Controls (n = 27) Median (Q1–Q3) & | Ca Ovary (n = 25) Median (Q1–Q3) & | p-Value # | Ovary without Changes (n = 12) Median (Q1–Q3) & | Benign Ovarian Changes (n = 15) Median (Q1–Q3) & | Ca Ovary (n = 25) Median (Q1–Q3) & | p-Value @ | |
| Age (years) | 53 (45–67) | 63 (54.5–69.5) | 0.0445 | 58 (49.0–69.0) | 51 (44.0–60.0) | 63.0 (54.5–69.5) | 0.0404 |
| BMI | 27.14 (22.31–28.96) | 26.22 (20.93–30.42) | 0.8103 | 24.84 (22.32–28.67) | 27.33 (21.63–31.23) | 26.22 (20.93–30.42) | 0.9062 |
| Menopause No | N = 11 (85%) | N = 2 (15%) | 0.0068 ‡ | N = 4 (31%) | N = 7 (54%) | N = 2 (15%) | 0.0151§ |
| Menopause Yes | N = 15 (40%) | N = 22 (60%) | N = 7 (19%) | N = 8 (22%) | N = 22 (59%) | ||
| ESR1 Cr $ | 0.0042 (0.0002–0.0074) | 0.0041 (0.0019–0.0370) | 0.2339 | 0.0042 (0.0006–0.0468) | 0.0038 (0.0001–0.0068) | 0.0041 (0.0019–0.0369) | 0.2579 |
| ESR2 Cr $ | 0.0675 (0.0329–0.1193) | 0.0329 (0.0157–0.1190) | 0.1996 | 0.0416 (0.0329–0.1106) | 0.1020 (0.0675–0.1365) | 0.0329 (0.0157–0.1193) | 0.0997 |
| PELP1 Cr $ | 0.2880 (0.1863–0.5932) | 0.1863 (0.0846–0.2880) | 0.0602 | 0.3389 (0.1863–0.5932) | 0.2880 (0.0846–0.5932) | 0.1863 (0.0846–0.2880) | 0.1193 |
| SRC Cr $ | 0.1003 (0.0388–0.1770) | 0.0136 (0.0035–0.0340) | 0.0070 | 0.1000 (0.0266–0.2077) | 0.1003 (0.0379–0.1530) | 0.0136 (0.0036–0.0340) | 0.0247 |
| ESR1/ESR2 | 0.0380 (0.0034–0.1273) | 0.1805 (0.0225–0.8400) | 0.0842 | 0.0658 (0.0195–0.4106) | 0.0343 (0.0007–0.0653) | 0.1805 (0.0225–0.8403) | 0.0711 |
| ESR1/PELP1 | 0.0076 (0.0021–0.0183) | 0.0598 (0.0100–0.1970) | 0.0189 | 0.0076 (0.0034–0.2070) | 0.0078 (0.0021–0.0131) | 0.0598 (0.0100–0.1969) | 0.0468 |
| ESR1/SRC | 0.0282 (0.0103–0.0762) | 0.4791 (0.0649–2.1490) | 0.0048 | 0.0419 (0.0236–0.7292) | 0.0261 (0.0095–0.0604) | 0.4791 (0.0649–2.1491) | 0.0079 |
| ESR2/PELP1 | 0.2156 (0.1720–0.5340) | 0.2694 (0.1026–0.5950) | 0.9249 | 0.1744 (0.1240–0.1809) | 0.3881 (0.2302–0.7934) | 0.2694 (0.1026–0.5953) | 0.0842 |
| ESR2/SRC | 0.7612 (0.4080–3.0752) | 3.0844 (0.3503–21.790) | 0.2308 | 0.6691 (0.3079–2.7817) | 0.8509 (0.5823–3.0752) | 3.0844 (0.3503–21.790) | 0.3796 |
| PELP1/SRC | 3.8765 (1.3271–7.0129) | 7.2099 (2.2466–34.277) | 0.0808 | 4.0065 (3.3745–15.903) | 2.0548 (0.8433–5.2661) | 7.2099 (2.2466–34.277) | 0.1167 |
| Gene | Manufacturer’s Designation | Cat. No. | Primer Sequence 5’→3’ | Amplicon Length (bp) | Manufacturer | |
|---|---|---|---|---|---|---|
| ESR1 | #69 | 04688686001 | F | ccttcttcaagagaagtattcaagg | 160 | Roche |
| R | attcccacttcgtagcatttg | |||||
| ESR2 | dHsaCPE5037392 | 10041596 | * | 87 | BioRad | |
| PELP1 | #24 | 04686985001 | F | caaggaggagactcacaggag | 131 | Roche |
| R | caaggaggagactcacaggag | |||||
| SRC | #21 | 04686942001 | F | gccatgttcactccggttt | 100 | Roche |
| R | cagcgtcctcatctggtttc | |||||
| HPRT | 102079 | 05532957001 | * | Roche | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Englert-Golon, M.; Andrusiewicz, M.; Żbikowska, A.; Chmielewska, M.; Sajdak, S.; Kotwicka, M. Altered Expression of ESR1, ESR2, PELP1 and c-SRC Genes Is Associated with Ovarian Cancer Manifestation. Int. J. Mol. Sci. 2021, 22, 6216. https://doi.org/10.3390/ijms22126216
Englert-Golon M, Andrusiewicz M, Żbikowska A, Chmielewska M, Sajdak S, Kotwicka M. Altered Expression of ESR1, ESR2, PELP1 and c-SRC Genes Is Associated with Ovarian Cancer Manifestation. International Journal of Molecular Sciences. 2021; 22(12):6216. https://doi.org/10.3390/ijms22126216
Chicago/Turabian StyleEnglert-Golon, Monika, Mirosław Andrusiewicz, Aleksandra Żbikowska, Małgorzata Chmielewska, Stefan Sajdak, and Małgorzata Kotwicka. 2021. "Altered Expression of ESR1, ESR2, PELP1 and c-SRC Genes Is Associated with Ovarian Cancer Manifestation" International Journal of Molecular Sciences 22, no. 12: 6216. https://doi.org/10.3390/ijms22126216
APA StyleEnglert-Golon, M., Andrusiewicz, M., Żbikowska, A., Chmielewska, M., Sajdak, S., & Kotwicka, M. (2021). Altered Expression of ESR1, ESR2, PELP1 and c-SRC Genes Is Associated with Ovarian Cancer Manifestation. International Journal of Molecular Sciences, 22(12), 6216. https://doi.org/10.3390/ijms22126216