
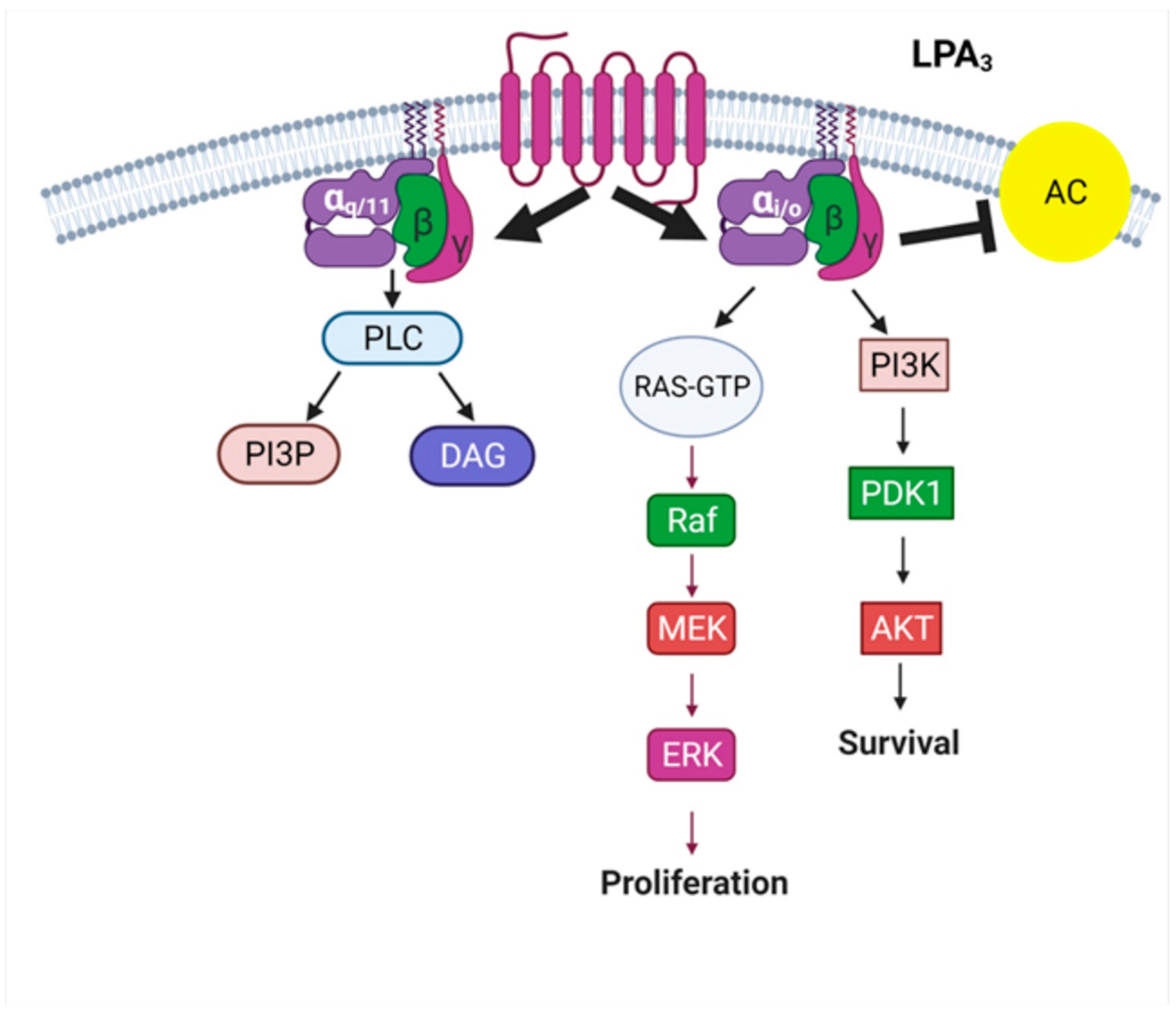
The LPA3 Receptor: Regulation and Activation of Signaling Pathways

 , and
, and
Abstract
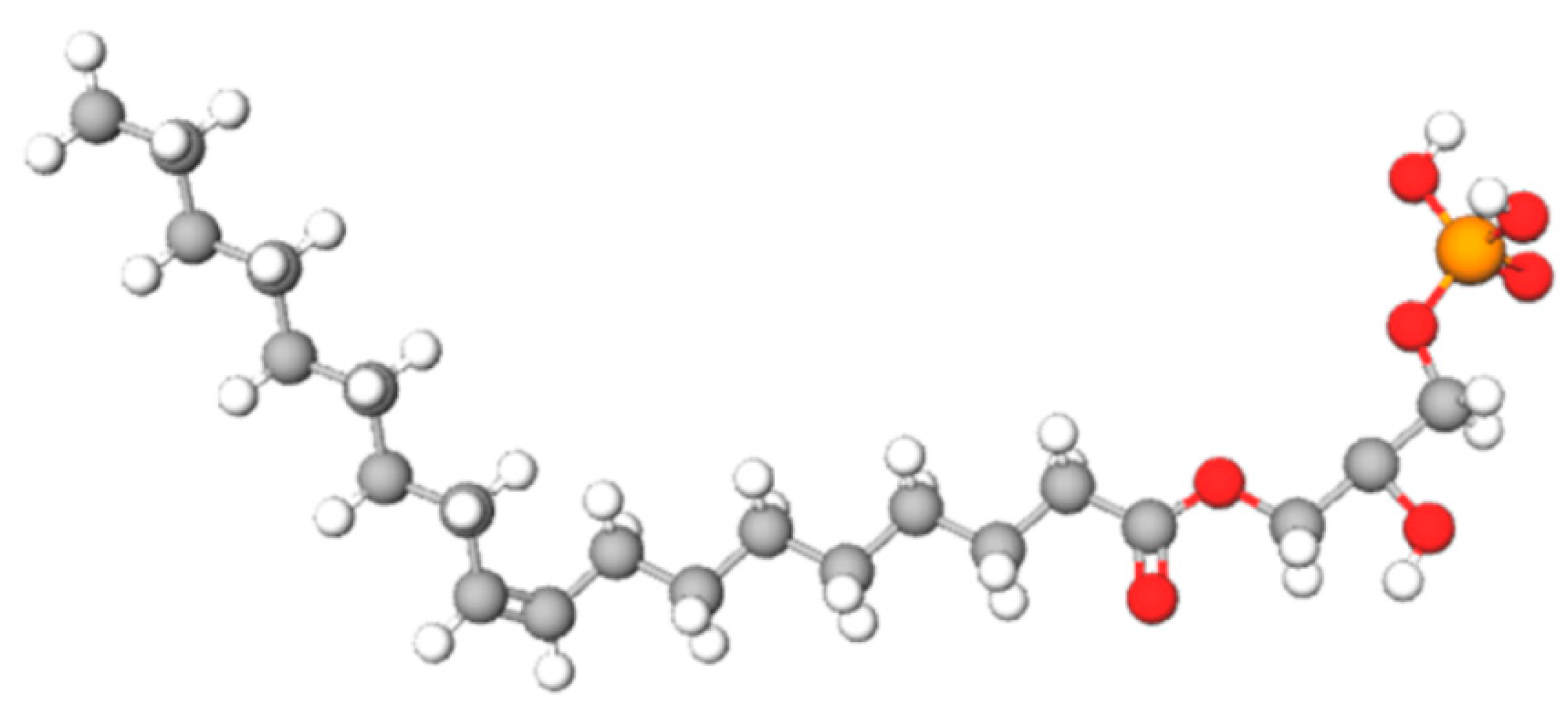
:1. Introduction
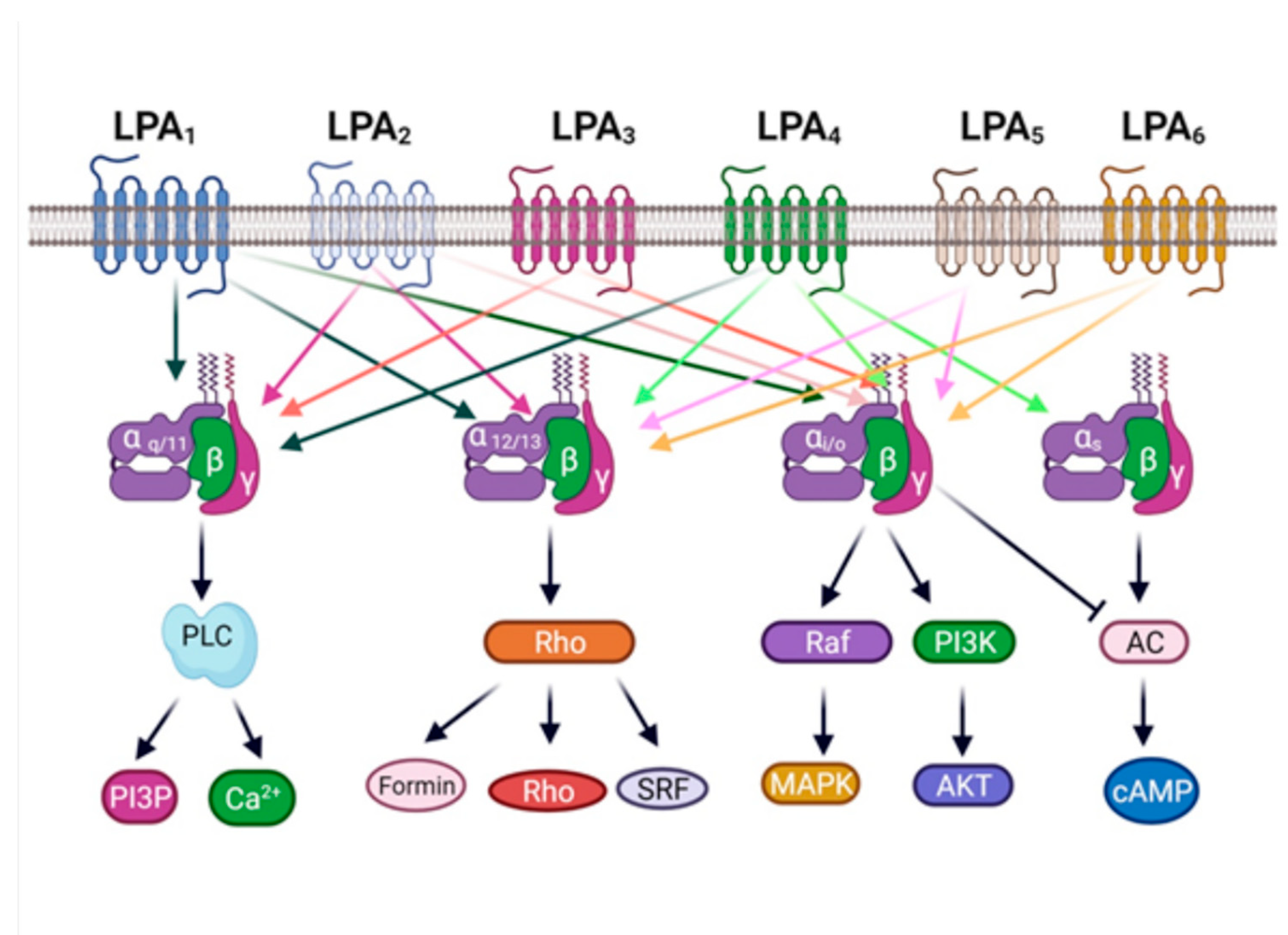
2. The LPA3 Receptor Is Involved in Physiology and Pathology
2.1. Antioxidant Enzymes Are Regulated by LPA3
2.2. LPA3 Actions in Cardiac Cells and Function
2.3. Participation of LPA3 in Fertility, Embryo Implantation, and Development
2.4. LPA3 in Cancer
2.5. Other Processes in Which LPA3 Participates
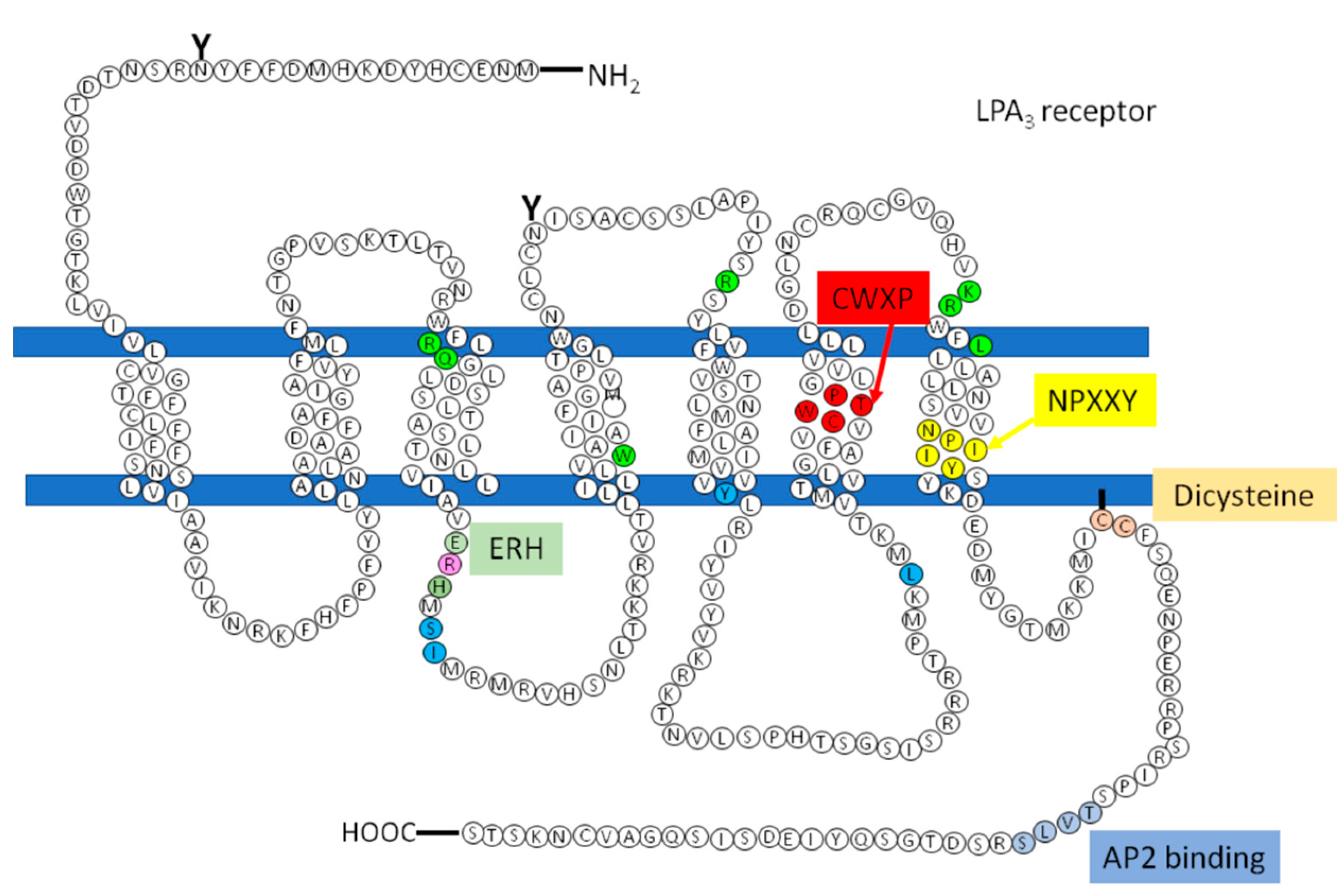
3. The LPA3 Receptor: Structure and Function
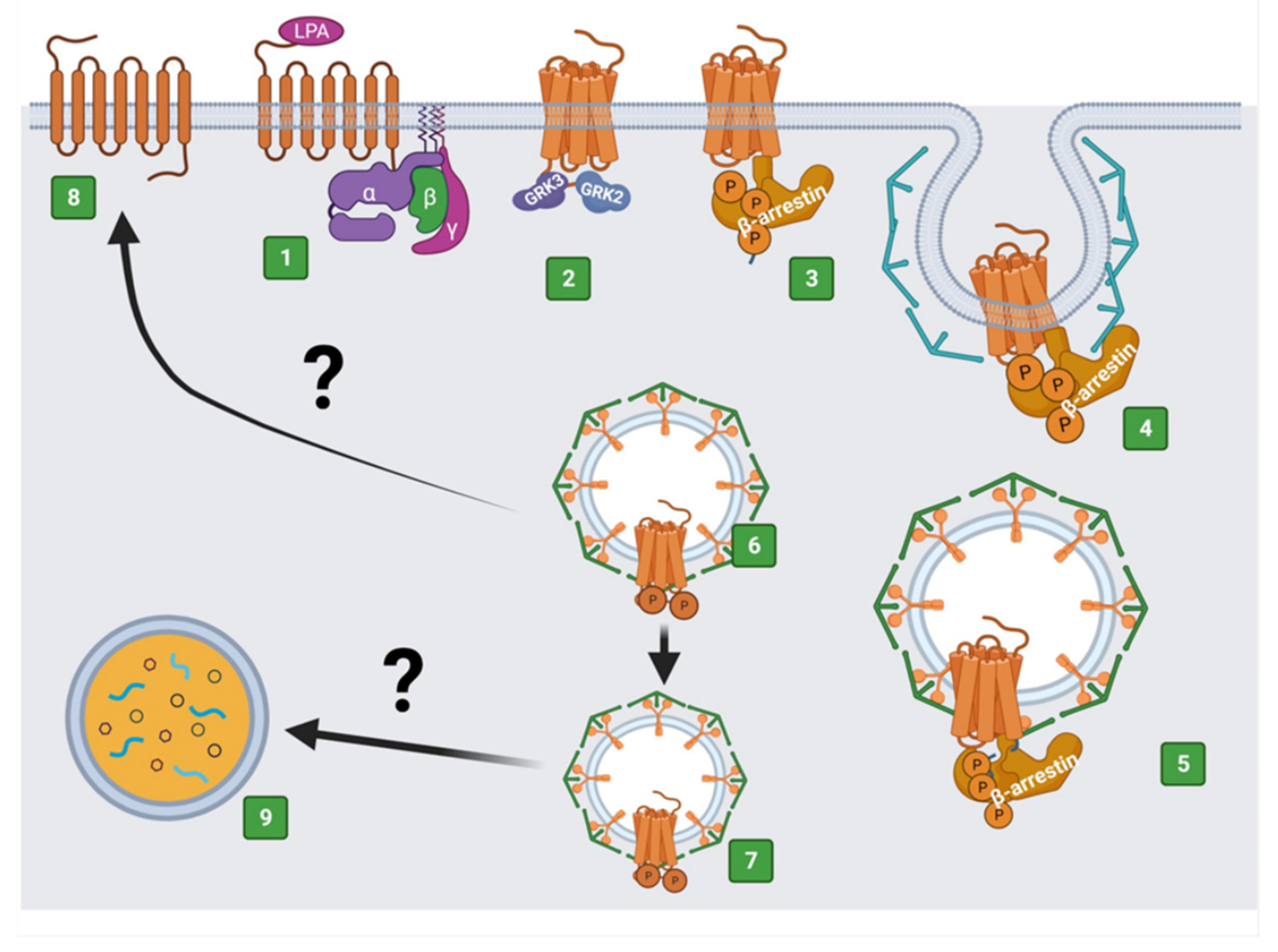
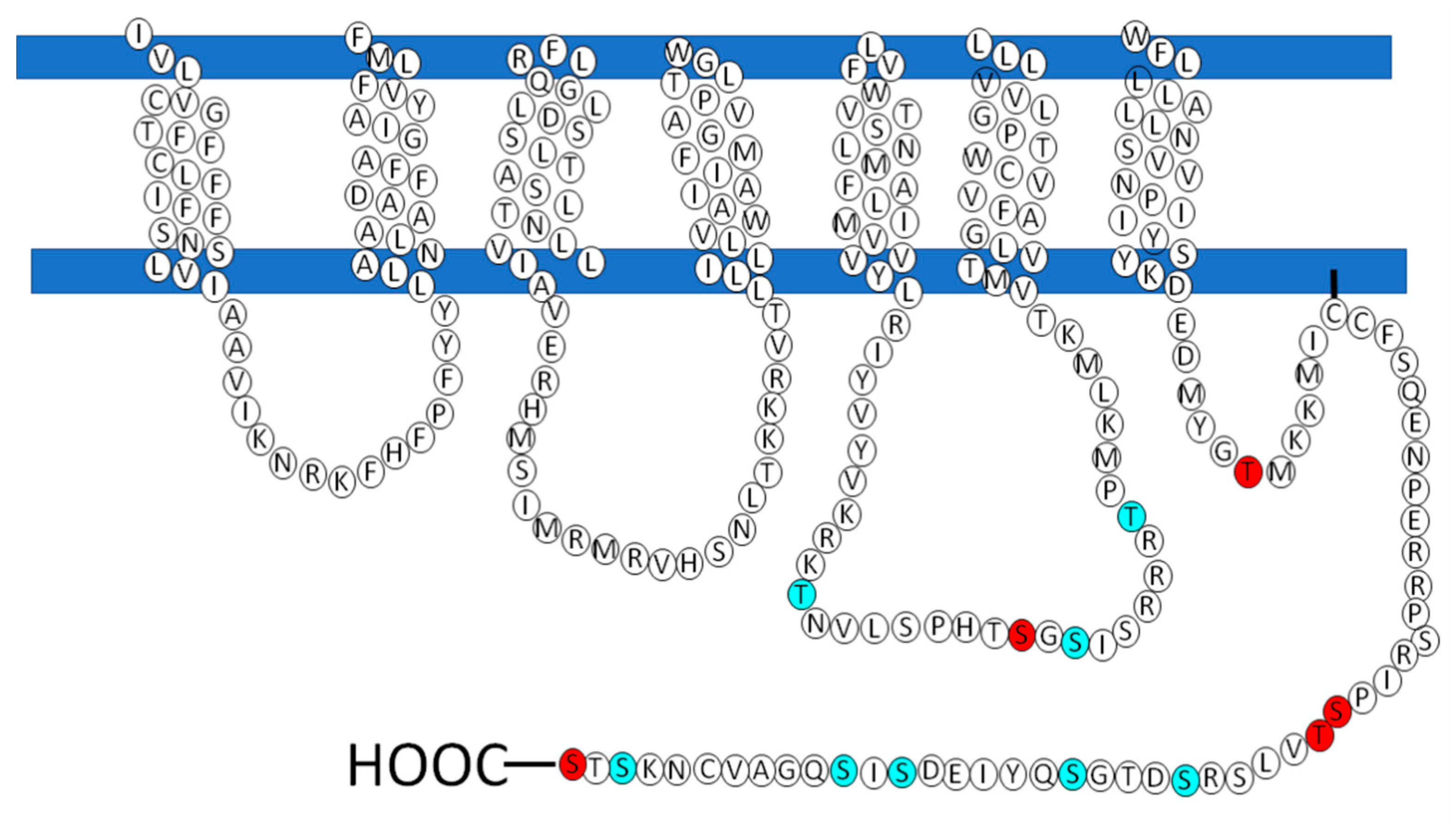
4. The Regulation of LPA3 Receptors and Its Possible Impact on Signaling
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kihara, Y.; Maceyka, M.; Spiegel, S.; Chun, J. Lysophospholipid Receptor Nomenclature Review: IUPHAR Review 8: Lysophospholipid Receptor Nomenclature. Br. J. Pharmacol. 2014, 171, 3575–3594. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Chen, G.-X. Production of Extracellular Lysophosphatidic Acid in the Regulation of Adipocyte Functions and Liver Fibrosis. World J. Gastroenterol. 2018, 24, 4132–4151. [Google Scholar] [CrossRef] [PubMed]
- Gaits, F.; Fourcade, O.; Le Balle, F.; Gueguen, G.; Gaigé, B.; Gassama-Diagne, A.; Fauvel, J.; Salles, J.P.; Mauco, G.; Simon, M.F.; et al. Lysophosphatidic Acid as a Phospholipid Mediator: Pathways of Synthesis. FEBS Lett. 1997, 410, 54–58. [Google Scholar] [CrossRef] [Green Version]
- Nakanaga, K.; Hama, K.; Aoki, J. Autotaxin-An LPA Producing Enzyme with Diverse Functions. J. Biochem. 2010, 148, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yung, Y.C.; Stoddard, N.C.; Chun, J. LPA Receptor Signaling: Pharmacology, Physiology, and Pathophysiology. J. Lipid Res. 2014, 55, 1192–1214. [Google Scholar] [CrossRef] [Green Version]
- Aoki, J. Mechanisms of Lysophosphatidic Acid Production. Semin. Cell Dev. Biol. 2004, 15, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.; Govindarajulu, M.; Suppiramaniam, V.; Moore, T.; Dhanasekaran, M. Autotaxin–Lysophosphatidic Acid Signaling in Alzheimer’s Disease. Int. J. Mol. Sci. 2018, 19, 1827. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.W.; Herr, D.R.; Noguchi, K.; Yung, Y.C.; Lee, C.-W.; Mutoh, T.; Lin, M.-E.; Teo, S.T.; Park, K.E.; Mosley, A.N.; et al. LPA Receptors: Subtypes and Biological Actions. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 157–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, J.; Hla, T.; Lynch, K.R.; Spiegel, S.; Moolenaar, W.H. International Union of Basic and Clinical Pharmacology. LXXVIII. Lysophospholipid Receptor Nomenclature: TABLE 1. Pharmacol. Rev. 2010, 62, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, D.; Kobayashi, D.; Akahoshi, N.; Ohto-Nakanishi, T.; Yoshioka, K.; Takuwa, Y.; Mizuno, S.; Takahashi, S.; Ishii, S. Lysophosphatidic Acid–Induced YAP/TAZ Activation Promotes Developmental Angiogenesis by Repressing Notch Ligand Dll4. J. Clin. Investig. 2019, 129, 4332–4349. [Google Scholar] [CrossRef]
- Rivera-Lopez, C.M.; Tucker, A.L.; Lynch, K.R. Lysophosphatidic Acid (LPA) and Angiogenesis. Angiogenesis 2008, 11, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Gross, I.; Bräuer, A. Modulation of Lysophosphatidic Acid (LPA) Receptor Activity: The Key to Successful Neural Regeneration? Neural Regen. Res. 2020, 15, 53. [Google Scholar] [CrossRef]
- Sayas, C.L.; Moreno-Flores, M.T.; Avila, J.; Wandosell, F. The Neurite Retraction Induced by Lysophosphatidic Acid Increases Alzheimer’s Disease-like Tau Phosphorylation. J. Biol. Chem. 1999, 274, 37046–37052. [Google Scholar] [CrossRef] [Green Version]
- Jang, M.; Choi, J.H.; Chang, Y.; Lee, S.J.; Nah, S.-Y.; Cho, I.-H. Gintonin, a Ginseng-Derived Ingredient, as a Novel Therapeutic Strategy for Huntington’s Disease: Activation of the Nrf2 Pathway through Lysophosphatidic Acid Receptors. Brain Behav. Immun. 2019, 80, 146–162. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, E.; Kitayoshi, M.; Yoshikawa, K.; Shibata, A.; Honoki, K.; Fukushima, N.; Tsujiuchi, T. Loss of Lysophosphatidic Acid Receptor-3 Suppresses Cell Migration Activity of Human Sarcoma Cells. J. Recept. Signal. Transduct. 2012, 32, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Okabe, K.; Kato, K.; Teranishi, M.; Okumura, M.; Fukui, R.; Mori, T.; Fukushima, N.; Tsujiuchi, T. Induction of Lysophosphatidic Acid Receptor-3 by 12-O-Tetradecanoylphorbol-13-Acetate Stimulates Cell Migration of Rat Liver Cells. Cancer Lett. 2011, 309, 236–242. [Google Scholar] [CrossRef]
- Shano, S.; Hatanaka, K.; Ninose, S.; Moriyama, R.; Tsujiuchi, T.; Fukushima, N. A Lysophosphatidic Acid Receptor Lacking the PDZ-Binding Domain Is Constitutively Active and Stimulates Cell Proliferation. Biochim. Biophys. Acta BBA Mol. Cell Res. 2008, 1783, 748–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, M.; Okabe, K.; Kato, K.; Okumura, M.; Fukui, R.; Fukushima, N.; Tsujiuchi, T. Differential Function of Lysophosphatidic Acid Receptors in Cell Proliferation and Migration of Neuroblastoma Cells. Cancer Lett. 2012, 316, 91–96. [Google Scholar] [CrossRef]
- Goldsmith, Z.G.; Ha, J.H.; Jayaraman, M.; Dhanasekaran, D.N. Lysophosphatidic Acid Stimulates the Proliferation of Ovarian Cancer Cells via the Gep Proto-Oncogene G 12. Genes Cancer 2011, 2, 563–575. [Google Scholar] [CrossRef] [Green Version]
- Anliker, B.; Choi, J.W.; Lin, M.-E.; Gardell, S.E.; Rivera, R.R.; Kennedy, G.; Chun, J. Lysophosphatidic Acid (LPA) and Its Receptor, LPA1, Influence Embryonic Schwann Cell Migration, Myelination, and Cell-to-Axon Segregation: LPA 1 Regulates Schwann Cell Physiology. Glia 2013, 61, 2009–2022. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Marín, L.; Ladrón de Guevara-Miranda, D.; Mañas-Padilla, M.C.; Alén, F.; Moreno-Fernández, R.D.; Díaz-Navarro, C.; Pérez-del Palacio, J.; García-Fernández, M.; Pedraza, C.; Pavón, F.J.; et al. Systemic Blockade of LPA1/3 Lysophosphatidic Acid Receptors by ki16425 Modulates the Effects of Ethanol on the Brain and Behavior. Neuropharmacology 2018, 133, 189–201. [Google Scholar] [CrossRef]
- Yung, Y.C.; Stoddard, N.C.; Mirendil, H.; Chun, J. Lysophosphatidic Acid Signaling in the Nervous System. Neuron 2015, 85, 669–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, H.; Neyama, H.; Sasaki, K.; Miyama, C.; Iwamoto, R. Lysophosphatidic Acid LPA1 and LPA3 Receptors Play Roles in the Maintenance of Late Tissue Plasminogen Activator-Induced Central Poststroke Pain in Mice. Neurobiol. Pain 2019, 5, 100020. [Google Scholar] [CrossRef]
- Kuwajima, K.; Sumitani, M.; Kurano, M.; Kano, K.; Nishikawa, M.; Uranbileg, B.; Tsuchida, R.; Ogata, T.; Aoki, J.; Yatomi, Y.; et al. Lysophosphatidic Acid Is Associated with Neuropathic Pain Intensity in Humans: An Exploratory Study. PLoS ONE 2018, 13, e0207310. [Google Scholar] [CrossRef] [PubMed]
- Fayyaz, S.; Japtok, L.; Schumacher, F.; Wigger, D.; Schulz, T.J.; Haubold, K.; Gulbins, E.; Völler, H.; Kleuser, B. Lysophosphatidic Acid Inhibits Insulin Signaling in Primary Rat Hepatocytes via the LPA3 Receptor Subtype and Is Increased in Obesity. Cell. Physiol. Biochem. 2017, 43, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tigyi, G.; Dacheux, M.A.; Lin, K.H.; Yue, J.; Norman, D.; Benyó, Z.; Lee, S.C. Anti-Cancer Strategies Targeting the Autotaxin-Lysophosphatidic Acid Receptor Axis: Is There a Path Forward? Cancer Metastasis Rev. 2021, 40, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Cai, H.; Duan, X.; Yang, Y.; Li, M.; Qu, J.; Zhang, X.; Wang, J. Aberrant Expression and Potential Therapeutic Target of Lysophosphatidic Acid Receptor 3 in Triple-Negative Breast Cancers. Clin. Exp. Med. 2015, 15, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Quan, M.; Cui, J.; Feng, X.; Huang, Q. The Critical Role and Potential Target of the Autotaxin/Lysophosphatidate Axis in Pancreatic Cancer. Tumor Biol. 2017, 39, 101042831769454. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, H.; Xu, W.; Guo, X. Evaluation of Serum ATX and LPA as Potential Diagnostic Biomarkers in Patients with Pancreatic Cancer. BMC Gastroenterol. 2021, 21, 58. [Google Scholar] [CrossRef]
- Lee, J.; Kim, D.; Oh, Y.; Jun, H.-S. Lysophosphatidic Acid Signaling in Diabetic Nephropathy. Int. J. Mol. Sci. 2019, 20, 2850. [Google Scholar] [CrossRef] [Green Version]
- Tigyi, G.; Miledi, R. Lysophosphatidates Bound to Serum Albumin Activate Membrane Currents in Xenopus Oocytes and Neurite Retraction in PC12 Pheochromocytoma Cells. J. Biol. Chem. 1992, 267, 21360–21367. [Google Scholar] [CrossRef]
- Panther, E.; Idzko, M.; Corinti, S.; Ferrari, D.; Herouy, Y.; Mockenhaupt, M.; Dichmann, S.; Gebicke-Haerter, P.; di Virgilio, F.; Girolomoni, G.; et al. The Influence of Lysophosphatidic Acid on the Functions of Human Dendritic Cells. J. Immunol. 2002, 169, 4129–4135. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.D.; Dhanasekaran, D.N. LPA Stimulates the Phosphorylation of P130Cas via GI2 in Ovarian Cancer Cells. Genes Cancer 2012, 3, 578–591. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xu, J.; Han, X.; Wang, H.; Zhang, Y.; Dong, J.; Deng, Y.; Wang, J. Lysophosphatidic Acid Is Associated with Cardiac Dysfunction and Hypertrophy by Suppressing Autophagy via the LPA3/AKT/MTOR Pathway. Front. Physiol. 2018, 9, 1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Mu, G.; Zhang, L.; Zhou, W.; Zhang, J.; Yu, H. Lysophosphatidic Acid Stimulates Activation of Focal Adhesion Kinase and Paxillin and Promotes Cell Motility, via LPA1–3, in Human Pancreatic Cancer. Dig. Dis. Sci. 2013, 58, 3524–3533. [Google Scholar] [CrossRef]
- Aoki, J. Two Pathways for Lysophosphatidic Acid Production. Biochim. Biophys. Acta (BBA) 2008, 1781, 513–518. [Google Scholar] [CrossRef]
- Schiöth, H.B.; Fredriksson, R. The GRAFS Classification System of G-Protein Coupled Receptors in Comparative Perspective. Gen. Comp. Endocrinol. 2005, 142, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.N.; Secker, A.; Freitas, A.A.; Mendao, M.; Timmis, J.; Flower, D.R. On the Hierarchical Classification of G Protein-Coupled Receptors. Bioinformatics 2007, 23, 3113–3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takuwa, Y.; Takuwa, N.; Sugimoto, N. The Edg Family G Protein-Coupled Receptors for Lysophospholipids: Their Signaling Properties and Biological Activities. J. Biochem. 2002, 131, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Neve, K.A.; Seamans, J.K.; Trantham-Davidson, H. Dopamine Receptor Signaling. J. Recept. Signal. Transduct. 2004, 24, 165–205. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-E.; Herr, D.R.; Chun, J. Lysophosphatidic Acid (LPA) Receptors: Signaling Properties and Disease Relevance. Prostaglandins Other Lipid Mediat. 2010, 91, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Dalle, S.; Imamura, T.; Rose, D.W.; Worrall, D.S.; Ugi, S.; Hupfeld, C.J.; Olefsky, J.M. Insulin Induces Heterologous Desensitization of G Protein-Coupled Receptor and Insulin-Like Growth Factor I Signaling by Downregulating β-Arrestin-1. Mol. Cell. Biol. 2002, 22, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calebiro, D.; Godbole, A. Internalization of G-Protein-Coupled Receptors: Implication in Receptor Function, Physiology and Diseases. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Bee, M.S.; Schonbrunn, A. Site Specificity of Agonist and Second Messenger-Activated Kinases for Somatostatin Receptor Subtype 2A (Sst2A) Phosphorylation. Mol. Pharmacol. 2009, 76, 68–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premont, R.T.; Inglese, J.; Lefkowitz, R.J. Protein Kinases That Phosphorylate Activated G Protein-coupled Receptors. FASEB J. 1995, 9, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Alfonzo-Méndez, M.A.; Carmona-Rosas, G.; Hernández-Espinosa, D.A.; Romero-Ávila, M.T.; García-Sáinz, J.A. Different Phosphorylation Patterns Regulate A1D-Adrenoceptor Signaling and Desensitization. Biochim. Biophys. Acta (BBA) 2018, 1865, 842–854. [Google Scholar] [CrossRef]
- Wootten, D.; Christopoulos, A.; Marti-Solano, M.; Babu, M.M.; Sexton, P.M. Mechanisms of Signalling and Biased Agonism in G Protein-Coupled Receptors. Nat. Rev. Mol. Cell Biol. 2018, 19, 638–653. [Google Scholar] [CrossRef]
- Mundell, S.J.; Jones, M.L.; Hardy, A.R.; Barton, J.F.; Beaucourt, S.M.; Conley, P.B.; Poole, A.W. Distinct Roles for Protein Kinase C Isoforms in Regulating Platelet Purinergic Receptor Function. Mol. Pharmacol. 2006, 70, 1132–1142. [Google Scholar] [CrossRef] [Green Version]
- Murph, M.M.; Scaccia, L.A.; Volpicelli, L.A.; Radhakrishna, H. Agonist-Induced Endocytosis of Lysophosphatidic Acid-Coupled LPA1/EDG-2 Receptors via a Dynamin2- and Rab5-Dependent Pathway. J. Cell Sci. 2003, 116, 1969–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhaes, A.C.; Dunn, H.; Ferguson, S.S. Regulation of GPCR Activity, Trafficking and Localization by GPCR-Interacting Proteins: Regulation of G-Protein-Coupled Receptor Activity. Br. J. Pharmacol. 2012, 165, 1717–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.; Khvorova, A.; Marshall, W.; Sorkin, A. Analysis of Clathrin-Mediated Endocytosis of Epidermal Growth Factor Receptor by RNA Interference. J. Biol. Chem. 2004, 279, 16657–16661. [Google Scholar] [CrossRef] [Green Version]
- Lobingier, B.T.; von Zastrow, M. When Trafficking and Signaling Mix: How Subcellular Location Shapes G Protein-Coupled Receptor Activation of Heterotrimeric G Proteins. Traffic 2019, 20, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Böhm, S.K.; Khitin, L.M.; Smeekens, S.P.; Grady, E.F.; Payan, D.G.; Bunnett, N.W. Identification of Potential Tyrosine-Containing Endocytic Motifs in the Carboxyl-Tail and Seventh Transmembrane Domain of the Neurokinin 1 Receptor. J. Biol. Chem. 1997, 272, 2363–2372. [Google Scholar] [CrossRef] [Green Version]
- Zaslavsky, A.; Singh, L.S.; Tan, H.; Ding, H.; Liang, Z.; Xu, Y. Homo- and Hetero-Dimerization of LPA/S1P Receptors, OGR1 and GPR4. Biochim. Biophys. Acta (BBA) 2006, 1761, 1200–1212. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.M.; Chiang, J.C.; Lin, Y.C.; Lin, Y.N.; Chuang, P.Y.; Chang, Y.C.; Chen, C.C.; Wu, K.Y.; Hsieh, J.C.; Chen, S.K.; et al. Lysophosphatidic Acid Receptor LPA3 Prevents Oxidative Stress and Cellular Senescence in Hutchinson–Gilford Progeria Syndrome. Aging Cell 2020, 19, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Umeno, A.; Biju, V.; Yoshida, Y. In Vivo ROS Production and Use of Oxidative Stress-Derived Biomarkers to Detect the Onset of Diseases Such as Alzheimer’s Disease, Parkinson’s Disease, and Diabetes. Free Radic. Res. 2017, 51, 413–427. [Google Scholar] [CrossRef]
- Kolodkin, A.N.; Sharma, R.P.; Colangelo, A.M.; Ignatenko, A.; Martorana, F.; Jennen, D.; Briedé, J.J.; Brady, N.; Barberis, M.; Mondeel, T.D.G.A.; et al. ROS Networks: Designs, Aging, Parkinson’s Disease and Precision Therapies. NPJ Syst. Biol. Appl. 2020, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Bobilev, I.; Novik, V.; Levi, I.; Shpilberg, O.; Levy, J.; Sharoni, Y.; Studzinski, G.P.; Danilenko, M. The Nrf2 Transcription Factor Is a Positive Regulator of Myeloid Differentiation of Acute Myeloid Leukemia Cells. Cancer Biol. Ther. 2011, 11, 317–329. [Google Scholar] [CrossRef]
- Mirzaei, S.; Zarrabi, A.; Hashemi, F.; Zabolian, A.; Saleki, H.; Azami, N.; Hamzehlou, S.; Farahani, M.V.; Hushmandi, K.; Ashrafizadeh, M.; et al. Nrf2 Signaling Pathway in Chemoprotection and Doxorubicin Resistance: Potential Application in Drug Discovery. Antioxidants 2021, 10, 349. [Google Scholar] [CrossRef]
- Wang, X.-J.; Sun, Z.; Villeneuve, N.F.; Zhang, S.; Zhao, F.; Li, Y.; Chen, W.; Yi, X.; Zheng, W.; Wondrak, G.T.; et al. Nrf2 Enhances Resistance of Cancer Cells to Chemotherapeutic Drugs, the Dark Side of Nrf2. Carcinogenesis 2008, 29, 1235–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Huang, G.; Cheng, H. Transcription Factor Nrf2 Induces the Up-Regulation of LncRNA TUG1 to Promote Progression and Adriamycin Resistance in Urothelial Carcinoma of the Bladder. Cancer Manag. Res. 2019, 11, 6079–6090. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Guo, Y.; Feng, Z.; Bergan, R.; Li, B.; Qin, Y.; Zhao, L.; Zhang, Z.; Shi, M. Involvement of the PI3K/Akt/Nrf2 Signaling Pathway in Resveratrol-Mediated Reversal of Drug Resistance in HL-60/ADR Cells. Nutr. Cancer 2019, 71, 1007–1018. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Y.; Sternberg, P.; Cai, J. Essential Roles of the PI3 Kinase/Akt Pathway in Regulating Nrf2-Dependent Antioxidant Functions in the RPE. Investig. Opthalmol. Vis. Sci. 2008, 49, 1671. [Google Scholar] [CrossRef] [Green Version]
- Hilal-Dandan, R.; Means, C.K.; Gustafsson, Å.B.; Morissette, M.R.; Adams, J.W.; Brunton, L.L.; Heller Brown, J. Lysophosphatidic Acid Induces Hypertrophy of Neonatal Cardiac Myocytes via Activation of Gi and Rho. J. Mol. Cell. Cardiol. 2004, 36, 481–493. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Y.; Zhu, W.; Han, Y.; Han, B.; Xu, R.; Deng, L.; Cai, Y.; Cong, X.; Yang, Y.; et al. Specific LPA Receptor Subtype Mediation of LPA-Induced Hypertrophy of Cardiac Myocytes and Involvement of Akt and NFκB Signal Pathways. J. Cell. Biochem. 2008, 103, 1718–1731. [Google Scholar] [CrossRef]
- Cai, L.; Fan, G.; Wang, F.; Liu, S.; Li, T.; Cong, X.; Chun, J.; Chen, X. Protective Role for LPA3 in Cardiac Hypertrophy Induced by Myocardial Infarction but Not by Isoproterenol. Front. Physiol. 2017, 8, 356. [Google Scholar] [CrossRef] [PubMed]
- Kano, K.; Matsumoto, H.; Inoue, A.; Yukiura, H.; Kanai, M.; Chun, J.; Ishii, S.; Shimizu, T.; Aoki, J. Molecular Mechanism of Lysophosphatidic Acid-Induced Hypertensive Response. Sci. Rep. 2019, 9, 2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aikawa, S.; Kano, K.; Inoue, A.; Wang, J.; Saigusa, D.; Nagamatsu, T.; Hirota, Y.; Fujii, T.; Tsuchiya, S.; Taketomi, Y.; et al. Autotaxin–Lysophosphatidic Acid–LPA3 Signaling at the Embryo-epithelial Boundary Controls Decidualization Pathways. EMBO J. 2017, 36, 2146–2160. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Seo, H.; Sung, Y.; Ka, H.; Song, G.; Kim, J. Lysophosphatidic Acid (LPA) Receptor 3-Mediated LPA Signal Transduction Pathways: A Possible Relationship with Early Development of Peri-Implantation Porcine Conceptus. Biol. Reprod. 2016, 94. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Hama, K.; Contos, J.J.A.; Anliker, B.; Inoue, A.; Skinner, K.; Suzuki, H.; Amano, T.; Kennedy, G.; Arai, H.; et al. LPA3-Mediated Lysophosphatidic Acid Signalling in Implantation and Embryo Spacing. Nature 2005, 435, 104–108. [Google Scholar] [CrossRef]
- Hama, K.; Aoki, J.; Bandoh, K.; Inoue, A.; Endo, T.; Amano, T.; Suzuki, H.; Arai, H. Lysophosphatidic Receptor, LPA3, Is Positively and Negatively Regulated by Progesterone and Estrogen in the Mouse Uterus. Life Sci. 2006, 79, 1736–1740. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, X.; Gajewiak, J.; Tsukahara, R.; Fujiwara, Y.; Liu, J.; Fells, J.I.; Perygin, D.; Parrill, A.L.; Tigyi, G.; et al. Dual Activity Lysophosphatidic Acid Receptor Pan-Antagonist/Autotaxin Inhibitor Reduces Breast Cancer Cell Migration In Vitro and Causes Tumor Regression In Vivo. Cancer Res. 2009, 69, 5441–5449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popnikolov, N.K.; Dalwadi, B.H.; Thomas, J.D.; Johannes, G.J.; Imagawa, W.T. Association of Autotaxin and Lysophosphatidic Acid Receptor 3 with Aggressiveness of Human Breast Carcinoma. Tumor Biol. 2012, 33, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Murph, M.M.; Lu, Y.; Liu, S.; Hall, H.S.; Liu, J.; Stephens, C.; Fang, X.; Mills, G.B. Lysophosphatidic Acid Receptors Determine Tumorigenicity and Aggressiveness of Ovarian Cancer Cells. JNCI J. Natl. Cancer Inst. 2008, 100, 1630–1642. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, K.; Takahashi, K.; Yamasaki, E.; Onishi, Y.; Fukushima, N.; Honoki, K.; Tsujiuchi, T. Lysophosphatidic Acid Signaling via LPA1 and LPA3 Regulates Cellular Functions during Tumor Progression in Pancreatic Cancer Cells. Exp. Cell Res. 2017, 352, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Jeon, E.S.; Heo, S.C.; Lee, I.H.; Choi, Y.J.; Park, J.H.; Choi, K.U.; Park, D.Y.; Suh, D.-S.; Yoon, M.-S.; Kim, J.H. Ovarian Cancer-Derived Lysophosphatidic Acid Stimulates Secretion of VEGF and Stromal Cell-Derived Factor-1α from Human Mesenchymal Stem Cells. Exp. Mol. Med. 2010, 42, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Jeong, K.J.; Panupinthu, N.; Yu, S.; Lee, J.; Han, J.W.; Kim, J.M.; Lee, J.-S.; Kang, J.; Park, C.G.; et al. Lysophosphatidic Acid Augments Human Hepatocellular Carcinoma Cell Invasion through LPA1 Receptor and MMP-9 Expression. Oncogene 2011, 30, 1351–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Sun, L.; Jiao, Y.; Lee, L.T.O. The Role of G Protein-Coupled Receptor Kinases in Cancer. Int. J. Biol. Sci. 2018, 14, 189–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okabe, K.; Hayashi, M.; Kato, K.; Okumura, M.; Fukui, R.; Honoki, K.; Fukushima, N.; Tsujiuchi, T. Lysophosphatidic Acid Receptor-3 Increases Tumorigenicity and Aggressiveness of Rat Hepatoma RH7777 Cells. Mol. Carcinog. 2013, 52, 247–254. [Google Scholar] [CrossRef]
- Zuo, C.; Li, X.; Huang, J.; Chen, D.; Ji, K.; Yang, Y.; Xu, T.; Zhu, D.; Yan, C.; Gao, P. Osteoglycin Attenuates Cardiac Fibrosis by Suppressing Cardiac Myofibroblast Proliferation and Migration through Antagonizing Lysophosphatidic Acid 3/Matrix Metalloproteinase 2/Epidermal Growth Factor Receptor Signalling. Cardiovasc. Res. 2018, 114, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix Metalloproteinases: Regulators of the Tumor Microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, D.; Yamane, M.; Tsujiuchi, T.; Moriyama, R.; Fukushima, N. Lysophosphatidic Acid Induces Neurite Branch Formation through LPA3. Mol. Cell. Neurosci. 2012, 50, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, N.; Ishii, S.; Tsujiuchi, T.; Kagawa, N.; Katoh, K. Comparative Analyses of Lysophosphatidic Acid Receptor-Mediated Signaling. Cell. Mol. Life Sci. 2015, 72, 2377–2394. [Google Scholar] [CrossRef]
- Chan, L.C.; Peters, W.; Xu, Y.; Chun, J.; Farese, R.V.; Cases, S. LPA3 Receptor Mediates Chemotaxis of Immature Murine Dendritic Cells to Unsaturated Lysophosphatidic Acid (LPA). J. Leukoc. Biol. 2007, 82, 1193–1200. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Fernandes, M.J.; Prestwich, G.D.; Turgeon, M.; di Battista, J.; Clair, T.; Poubelle, P.E.; Bourgoin, S.G. Regulation of Lysophosphatidic Acid Receptor Expression and Function in Human Synoviocytes: Implications for Rheumatoid Arthritis? Mol. Pharmacol. 2008, 73, 587–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nochi, H.; Tomura, H.; Tobo, M.; Tanaka, N.; Sato, K.; Shinozaki, T.; Kobayashi, T.; Takagishi, K.; Ohta, H.; Okajima, F.; et al. Stimulatory Role of Lysophosphatidic Acid in Cyclooxygenase-2 Induction by Synovial Fluid of Patients with Rheumatoid Arthritis in Fibroblast-Like Synovial Cells. J. Immunol. 2008, 181, 5111–5119. [Google Scholar] [CrossRef] [Green Version]
- Bandoh, K.; Aoki, J.; Taira, A.; Tsujimoto, M.; Arai, H.; Inoue, K. Lysophosphatidic Acid (LPA) Receptors of the EDG Family Are Differentially Activated by LPA Species. Structure-Activity Relationship of Cloned LPA Receptors. FEBS Lett. 2000, 478, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Bandoh, K.; Aoki, J.; Hosono, H.; Kobayashi, S.; Kobayashi, T.; Murakami-Murofushi, K.; Tsujimoto, M.; Arai, H.; Inoue, K. Molecular Cloning and Characterization of a Novel Human G-Protein-Coupled Receptor, EDG7, for Lysophosphatidic Acid. J. Biol. Chem. 1999, 274, 27776–27785. [Google Scholar] [CrossRef] [Green Version]
- Alcántara-Hernández, R.; Hernández-Méndez, A.; Campos-Martínez, G.A.; Meizoso-Huesca, A.; García-Sáinz, J.A. Phosphorylation and Internalization of Lysophosphatidic Acid Receptors LPA1, LPA2, and LPA3. PLoS ONE 2015, 10, e0140583. [Google Scholar] [CrossRef] [Green Version]
- Fells, J.I.; Tsukahara, R.; Liu, J.; Tigyi, G.; Parrill, A.L. Structure-Based Drug Design Identifies Novel LPA3 Antagonists. Bioorg. Med. Chem. 2009, 17, 7457–7464. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Zhang, J.; Zhang, S.; Li, J. A Single Amino Acid Mutation (R104P) in the E/DRY Motif of GPR40 Impairs Receptor Function. PLoS ONE 2015, 10, e0141303. [Google Scholar] [CrossRef]
- White, K.L.; Eddy, M.T.; Gao, Z.G.; Han, G.W.; Lian, T.; Deary, A.; Patel, N.; Jacobson, K.A.; Katritch, V.; Stevens, R.C. Structural Connection between Activation Microswitch and Allosteric Sodium Site in GPCR Signaling. Structure 2018, 26, 259.e5–269.e5. [Google Scholar] [CrossRef] [Green Version]
- Olivella, M.; Caltabiano, G.; Cordomí, A. The Role of Cysteine 6.47 in Class A GPCRs. BMC Struct. Biol. 2013, 13, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-M.; Benovic, J.L. Differential Roles of Arrestin-2 Interaction with Clathrin and Adaptor Protein 2 in G Protein-Coupled Receptor Trafficking. J. Biol. Chem. 2002, 277, 30760–30768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paing, M.M.; Johnston, C.A.; Siderovski, D.P.; Trejo, J. Clathrin Adaptor AP2 Regulates Thrombin Receptor Constitutive Internalization and Endothelial Cell Resensitization. Mol. Cell. Biol. 2006, 26, 3231–3242. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, B.L.; Trejo, J.A. Clathrin-Dependent Mechanisms of G Protein-Coupled Receptor Endocytosis. Traffic 2007, 8, 462–470. [Google Scholar] [CrossRef]
- Vickery, O.N.; Carvalheda, C.A.; Zaidi, S.A.; Pisliakov, A.V.; Katritch, V.; Zachariae, U. Intracellular Transfer of Na+ in an Active-State G-Protein-Coupled Receptor. Structure 2018, 26, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Rovati, G.E.; Capra, V.; Shaw, V.S.; Malik, R.U.; Sivaramakrishnan, S.; Neubig, R.R. The DRY Motif and the Four Corners of the Cubic Ternary Complex Model. Cell Signal. 2017, 35, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Yang, D.; Wu, M.; Guo, Y.; Guo, W.; Zhong, L.; Cai, X.; Dai, A.; Jang, W.; Shakhnovich, E.I.; et al. Common Activation Mechanism of Class A GPCRs. eLife 2019, 8, e50279. [Google Scholar] [CrossRef]
- Yuan, S.; Filipek, S.; Palczewski, K.; Vogel, H. Activation of G-Protein-Coupled Receptors Correlates with the Formation of a Continuous Internal Water Pathway. Nat. Commun. 2014, 5, 4733. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.C.; Zhou, Y.; Cao, C. Proton Transfer during Class-A GPCR Activation: Do the CWxP Motif and the Membrane Potential Act in Concert? Biophys. Rep. 2018, 4, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Moreira, I.S. Structural Features of the G-Protein/GPCR Interactions. Biochim. Biophys. Acta (BBA) 2014, 1840, 16–33. [Google Scholar] [CrossRef] [PubMed]
- Wacker, D.; Stevens, R.C.; Roth, B.L. How Ligands Illuminate GPCR Molecular Pharmacology. Cell 2017, 170, 414–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutkeviciute, I.; Vilardaga, J.-P. Structural Insights into Emergent Signaling Modes of G Protein–Coupled Receptors. J. Biol. Chem. 2020, 295, 11626–11642. [Google Scholar] [CrossRef] [PubMed]
- Ubarretxena-Belandia, I.; Engelman, D.M. Helical Membrane Proteins: Diversity of Functions in the Context of Simple Architecture. Curr. Opin. Struct. Biol. 2001, 11, 370–376. [Google Scholar] [CrossRef]
- Alewijnse, A.E.; Timmerman, H.; Jacobs, E.H.; Smit, M.J.; Roovers, E.; Cotecchia, S.; Leurs, R. The Effect of Mutations in the DRY Motif on the Constitutive Activity and Structural Instability of the Histamine H2Receptor. Mol. Pharmacol. 2000, 57, 890–898. [Google Scholar] [PubMed]
- He, R.; Browning, D.D.; Ye, R.D. Differential Roles of the NPXXY Motif in Formyl Peptide Receptor Signaling. J. Immunol. 2001, 166, 4099–4105. [Google Scholar] [CrossRef] [Green Version]
- Chung, D.A.; Wade, S.M.; Fowler, C.B.; Woods, D.D.; Abada, P.B.; Mosberg, H.I.; Neubig, R.R. Mutagenesis and Peptide Analysis of the DRY Motif in the α2A Adrenergic Receptor: Evidence for Alternate Mechanisms in G Protein-Coupled Receptors. Biochem. Biophys. Res. Commun. 2002, 293, 1233–1241. [Google Scholar] [CrossRef]
- Zhou, X.E.; He, Y.; de Waal, P.W.; Gao, X.; Kang, Y.; van Eps, N.; Yin, Y.; Pal, K.; Goswami, D.; White, T.A.; et al. Identification of Phosphorylation Codes for Arrestin Recruitment by G Protein-Coupled Receptors. Cell 2017, 170, 457.e13–469.e13. [Google Scholar] [CrossRef] [Green Version]
- Mayer, D.; Damberger, F.F.; Samarasimhareddy, M.; Feldmueller, M.; Vuckovic, Z.; Flock, T.; Bauer, B.; Mutt, E.; Zosel, F.; Allain, F.H.T.; et al. Distinct G Protein-Coupled Receptor Phosphorylation Motifs Modulate Arrestin Affinity and Activation and Global Conformation. Nat. Commun. 2019, 10, 1261. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Sardar, V.; Tokumura, A.; Baker, D.; Murakami-Murofushi, K.; Parrill, A.; Tigyi, G. Identification of Residues Responsible for Ligand Recognition and Regioisomeric Selectivity of Lysophosphatidic Acid Receptors Expressed in Mammalian Cells. J. Biol. Chem. 2005, 280, 35038–35050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentine, W.J.; Fells, J.I.; Perygin, D.H.; Mujahid, S.; Yokoyama, K.; Fujiwara, Y.; Tsukahara, R.; van Brocklyn, J.R.; Parrill, A.L.; Tigyi, G. Subtype-Specific Residues Involved in Ligand Activation of the Endothelial Differentiation Gene Family Lysophosphatidic Acid Receptors. J. Biol. Chem. 2008, 283, 12175–12187. [Google Scholar] [CrossRef] [Green Version]
- Kaye, R.G.; Saldanha, J.W.; Lu, Z.-L.; Hulme, E.C. Helix 8 of the M1 Muscarinic Acetylcholine Receptor: Scanning Mutagenesis Delineates a G Protein Recognition Site. Mol. Pharmacol. 2011, 79, 701–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, N.M.D.; Gardner, L.A.; White, S.W.; Bahouth, S.W. Characterization of the Residues in Helix 8 of the Human β1-Adrenergic Receptor That Are Involved in Coupling the Receptor to G Proteins. J. Biol. Chem. 2006, 281, 12896–12907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, J.; Thomas, W.G.; Aguilar, M.I.; Pattenden, L.K. Role of Helix 8 in G Protein-Coupled Receptors Based on Structure-Function Studies on the Type 1 Angiotensin Receptor. Mol. Cell. Endocrinol. 2009, 302, 118–127. [Google Scholar] [CrossRef]
- Dijkman, P.M.; Muñoz-García, J.C.; Lavington, S.R.; Kumagai, P.S.; dos Reis, R.I.; Yin, D.; Stansfeld, P.J.; Costa-Filho, A.J.; Watts, A. Conformational Dynamics of a G Protein–Coupled Receptor Helix 8 in Lipid Membranes. Sci. Adv. 2020, 6, eaav8207. [Google Scholar] [CrossRef] [PubMed]
- Ribas, C.; Penela, P.; Murga, C.; Salcedo, A.; García-Hoz, C.; Jurado-Pueyo, M.; Aymerich, I.; Mayor, F. The G Protein-Coupled Receptor Kinase (GRK) Interactome: Role of GRKs in GPCR Regulation and Signaling. Biochim. Biophys. Acta BBA Biomembr. 2007, 1768, 913–922. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, V.V.; Song, X.; Vishnivetskiy, S.A.; Gurevich, E.V. Enhanced Phosphorylation-Independent Arrestins and Gene Therapy. In Arrestins—Pharmacology and Therapeutic Potential; Gurevich, V.V., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 219, pp. 133–152. ISBN 978-3-642-41198-4. [Google Scholar]
- Pronin, A.N.; Carman, C.V.; Benovic, J.L. Structure-Function Analysis of G Protein-Coupled Receptor Kinase-5. J. Biol. Chem. 1998, 273, 31510–31518. [Google Scholar] [CrossRef] [Green Version]
- Watari, K.; Nakaya, M.; Kurose, H. Multiple Functions of G Protein-Coupled Receptor Kinases. J. Mol. Signal. 2014, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Drake, M.T.; Shenoy, S.K.; Lefkowitz, R.J. Trafficking of G Protein-Coupled Receptors. Circ. Res. 2006, 99, 570–582. [Google Scholar] [CrossRef]
- Reiter, E.; Lefkowitz, R.J. GRKs and β-Arrestins: Roles in Receptor Silencing, Trafficking and Signaling. Trends Endocrinol. Metab. 2006, 17, 159–165. [Google Scholar] [CrossRef]
- Baidya, M.; Kumari, P.; Dwivedi-Agnihotri, H.; Pandey, S.; Chaturvedi, M.; Stepniewski, T.M.; Kawakami, K.; Cao, Y.; Laporte, S.A.; Selent, J.; et al. Key Phosphorylation Sites in GPCR s Orchestrate the Contribution of β-Arrestin 1 in ERK 1/2 Activation. EMBO Rep. 2020, 21, e49886. [Google Scholar] [CrossRef] [PubMed]
- Sensoy, O.; Moreira, I.S.; Morra, G. Understanding the Differential Selectivity of Arrestins toward the Phosphorylation State of the Receptor. ACS Chem. Neurosci. 2016, 7, 1212–1224. [Google Scholar] [CrossRef]
- Ally, R.A.; Ives, K.L.; Traube, E.; Eltounsi, I.; Chen, P.-W.; Cahill, P.J.; Battey, J.F.; Hellmich, M.R.; Kroog, G.S. Agonist- and Protein Kinase C-Induced Phosphorylation Have Similar Functional Consequences for Gastrin-Releasing Peptide Receptor Signaling via Gq. Mol. Pharmacol. 2003, 64, 890–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Ahn, S.; Ren, X.-R.; Whalen, E.J.; Reiter, E.; Wei, H.; Lefkowitz, R.J. Functional Antagonism of Different G Protein-Coupled Receptor Kinases for -Arrestin-Mediated Angiotensin II Receptor Signaling. Proc. Natl. Acad. Sci. USA 2005, 102, 1442–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flock, T.; Ravarani, C.N.J.; Sun, D.; Venkatakrishnan, A.J.; Kayikci, M.; Tate, C.G.; Veprintsev, D.B.; Babu, M.M. Universal Allosteric Mechanism for Gα Activation by GPCRs. Nature 2015, 524, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Butcher, A.J.; Prihandoko, R.; Kong, K.C.; McWilliams, P.; Edwards, J.M.; Bottrill, A.; Mistry, S.; Tobin, A.B. Differential G-Protein-Coupled Receptor Phosphorylation Provides Evidence for a Signaling Bar Code. J. Biol. Chem. 2011, 286, 11506–11518. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Mendez, A.; Alcantara-Hernandez, R.; Garcia-Sainz, J.A. Lysophosphatidic Acid LPA1-3 Receptors: Signaling, Regulation and in Silico Analysis of Their Putative Phosphorylation Sites. Recept. Clin. Investig. 2014, 1, e193. [Google Scholar] [CrossRef]
- Gurevich, V.V.; Gurevich, E.V. GPCR Signaling Regulation: The Role of GRKs and Arrestins. Front. Pharmacol. 2019, 10, 125. [Google Scholar] [CrossRef] [Green Version]
- Black, J.B.; Premont, R.T.; Daaka, Y. Feedback Regulation of G Protein-Coupled Receptor Signaling by GRKs and Arrestins. Semin. Cell Dev. Biol. 2016, 50, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Lodowski, D.T.; Pitcher, J.A.; Capel, W.D.; Lefkowitz, R.J.; Tesmer, J.J.G. Keeping G Proteins at Bay: A Complex between G Protein-Coupled Receptor Kinase 2 and Gβγ. Science 2003, 300, 1256–1262. [Google Scholar] [CrossRef]
- Cassier, E.; Gallay, N.; Bourquard, T.; Claeysen, S.; Bockaert, J.; Crépieux, P.; Poupon, A.; Reiter, E.; Marin, P.; Vandermoere, F. Phosphorylation of β-Arrestin2 at Thr383 by MEK Underlies β-Arrestin-Dependent Activation of Erk1/2 by GPCRs. eLife 2017, 6, e23777. [Google Scholar] [CrossRef]
- Wei, H.; Ahn, S.; Shenoy, S.K.; Karnik, S.S.; Hunyady, L.; Luttrell, L.M.; Lefkowitz, R.J. Independent -Arrestin 2 and G Protein-Mediated Pathways for Angiotensin II Activation of Extracellular Signal-Regulated Kinases 1 and 2. Proc. Natl. Acad. Sci. USA 2003, 100, 10782–10787. [Google Scholar] [CrossRef] [Green Version]
- Murga, C.; Ruiz-Gómez, A.; García-Higuera, I.; Kim, C.M.; Benovic, J.L.; Mayor, F. High Affinity Binding of β-Adrenergic Receptor Kinase to Microsomal Membranes. J. Biol. Chem. 1996, 271, 985–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiina, T.; Arai, K.; Tanabe, S.; Yoshida, N.; Haga, T.; Nagao, T.; Kurose, H. Clathrin Box in G Protein-Coupled Receptor Kinase 2. J. Biol. Chem. 2001, 276, 33019–33026. [Google Scholar] [CrossRef] [Green Version]
- Peregrin, S.; Jurado-Pueyo, M.; Campos, P.M.; Sanz-Moreno, V.; Ruiz-Gomez, A.; Crespo, P.; Mayor, F.; Murga, C. Phosphorylation of P38 by GRK2 at the Docking Groove Unveils a Novel Mechanism for Inactivating P38MAPK. Curr. Biol. 2006, 16, 2042–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carman, C.V.; Barak, L.S.; Chen, C.; Liu-Chen, L.Y.; Onorato, J.J.; Kennedy, S.P.; Caron, M.G.; Benovic, J.L. Mutational Analysis of Gβγ and Phospholipid Interaction with Kinase G Protein-Coupled Receptor 2. J. Biol. Chem. 2000, 275, 10443–10452. [Google Scholar] [CrossRef] [Green Version]
- Bahouth, S.W.; Nooh, M.M. Barcoding of GPCR Trafficking and Signaling through the Various Trafficking Roadmaps by Compartmentalized Signaling Networks. Cell Signal. 2017, 36, 42–55. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, F.; Zhang, D.; Liu, Z.; Lin, A.; Liu, C.; Xiao, P.; Yu, X.; Sun, J.P. Phosphorylation of g Protein-Coupled Receptors: From the Barcode Hypothesis to the Flute Model. Mol. Pharmacol. 2017, 92, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Milligan, G. New Aspects of G-Protein-Coupled Receptor Signalling and Regulation. Trends Endocrinol. Metab. 1998, 9, 13–19. [Google Scholar] [CrossRef]
- Böttke, T.; Ernicke, S.; Serfling, R.; Ihling, C.; Burda, E.; Gurevich, V.V.; Sinz, A.; Coin, I. Exploring GPCR-arrestin Interfaces with Genetically Encoded Crosslinkers. EMBO Rep. 2020, 21. [Google Scholar] [CrossRef]
- Urs, N.M.; Kowalczyk, A.P.; Radhakrishna, H. Different Mechanisms Regulate Lysophosphatidic Acid (LPA)-Dependent versus Phorbol Ester-Dependent Internalization of the LPA1 Receptor. J. Biol. Chem. 2008, 283, 5249–5257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, G.; von Zastrow, M.; Friedman, P.A. Role of PDZ Proteins in Regulating Trafficking, Signaling, and Function of GPCRs: Means, Motif, and Opportunity. In Advances in Pharmacology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 62, pp. 279–314. ISBN 978-0-12-385952-5. [Google Scholar]
- Colgan, L.; Liu, H.; Huang, S.Y.; Liu, Y.-J. Dileucine Motif Is Sufficient for Internalization and Synaptic Vesicle Targeting of Vesicular Acetylcholine Transporter. Traffic 2007, 8, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Geisler, C.; Dietrich, J.; Nielsen, B.L.; Kastrup, J.; Lauritsen, J.P.H.; Ødum, N.; Christensen, M.D. Leucine-Based Receptor Sorting Motifs Are Dependent on the Spacing Relative to the Plasma Membrane. J. Biol. Chem. 1998, 273, 21316–21323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moo, E.V.; van Senten, J.R.; Bräuner-Osborne, H.; Møller, T.C. Arrestin-Dependent and -Independent Internalization of G Protein–Coupled Receptors: Methods, Mechanisms, and Implications on Cell Signaling. Mol. Pharmacol. 2021, 99, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Novakovic, S.; Sawai, E.T.; Radke, K. Dileucine and YXXL Motifs in the Cytoplasmic Tail of the Bovine Leukemia Virus Transmembrane Envelope Protein Affect Protein Expression on the Cell Surface. J. Virol. 2004, 78, 8301–8311. [Google Scholar] [CrossRef] [Green Version]
- Kozik, P.; Francis, R.W.; Seaman, M.N.J.; Robinson, M.S. A Screen for Endocytic Motifs. Traffic 2010, 11, 843–855. [Google Scholar] [CrossRef]
- Bonifacino, J.S.; Traub, L.M. Signals for Sorting of Transmembrane Proteins to Endosomes and Lysosomes. Annu. Rev. Biochem. 2003, 72, 395–447. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.N. Small Peptide Recognition Sequence for Intracellular Sorting. Curr. Opin. Biotechnol. 2010, 21, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Christensen, N.R.; Čalyševa, J.; Fernandes, E.F.A.; Lüchow, S.; Clemmensen, L.S.; Haugaard-Kedström, L.M.; Strømgaard, K. PDZ Domains as Drug Targets. Adv. Ther. 2019, 2, 1800143. [Google Scholar] [CrossRef] [Green Version]
- Varsano, T.; Taupin, V.; Guo, L.; Baterina, O.Y.; Farquhar, M.G. The PDZ Protein GIPC Regulates Trafficking of the LPA1 Receptor from APPL Signaling Endosomes and Attenuates the Cell’s Response to LPA. PLoS ONE 2012, 7, e49227. [Google Scholar] [CrossRef] [Green Version]
- Holcomb, J.; Jiang, Y.; Lu, G.; Trescott, L.; Brunzelle, J.; Sirinupong, N.; Li, C.; Naren, A.P.; Yang, Z. Structural Insights into PDZ-Mediated Interaction of NHERF2 and LPA2, a Cellular Event Implicated in CFTR Channel Regulation. Biochem. Biophys. Res. Commun. 2014, 446, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, D.; Sun, H.; Hall, R.A.; Yun, C.C. MAGI-3 Regulates LPA-Induced Activation of Erk and RhoA. Cell Signal. 2007, 19, 261–268. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamilies | GRKs | Domains of Interest |
|---|---|---|
| Visual GRKs | GRK1 and GRK7 | Prenylation |
| GRK2 or βARK GRK4 | GRK2 and GRK3 GRK4, GRK5 and GRK6 | Pleckstrin homology Palmitoylation, polybasic hydrophobic domains |
| Position | Amino Acid | PKC/PKA |
|---|---|---|
| 130 | S | PKA |
| 217 | T | PKCα/PKCδ/PKCγ |
| 233 | T | PKA/PKCδ/PKCι/PKCζ |
| 243 | T | PKCi/PKCζ |
| 321 | S | PKA/PKCδ/ |
| 325 | S | PKA/PKC/PKCε |
| 341 | S | PKCε |
| 351 | S | PKCε |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solís, K.H.; Romero-Ávila, M.T.; Guzmán-Silva, A.; García-Sáinz, J.A. The LPA3 Receptor: Regulation and Activation of Signaling Pathways. Int. J. Mol. Sci. 2021, 22, 6704. https://doi.org/10.3390/ijms22136704
Solís KH, Romero-Ávila MT, Guzmán-Silva A, García-Sáinz JA. The LPA3 Receptor: Regulation and Activation of Signaling Pathways. International Journal of Molecular Sciences. 2021; 22(13):6704. https://doi.org/10.3390/ijms22136704
Chicago/Turabian StyleSolís, Karina Helivier, M. Teresa Romero-Ávila, Alejandro Guzmán-Silva, and J. Adolfo García-Sáinz. 2021. "The LPA3 Receptor: Regulation and Activation of Signaling Pathways" International Journal of Molecular Sciences 22, no. 13: 6704. https://doi.org/10.3390/ijms22136704
APA StyleSolís, K. H., Romero-Ávila, M. T., Guzmán-Silva, A., & García-Sáinz, J. A. (2021). The LPA3 Receptor: Regulation and Activation of Signaling Pathways. International Journal of Molecular Sciences, 22(13), 6704. https://doi.org/10.3390/ijms22136704