
Difluoromethylornithine (DFMO), an Inhibitor of Polyamine Biosynthesis, and Antioxidant N-Acetylcysteine Potentiate Immune Response in Mice to the Recombinant Hepatitis C Virus NS5B Protein

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Induction of MDSCs and Tregs Correlates with Suppression of the Cellular Immune Response
2.2. Analysis of the Action of Compounds on T Cell Functional Activity
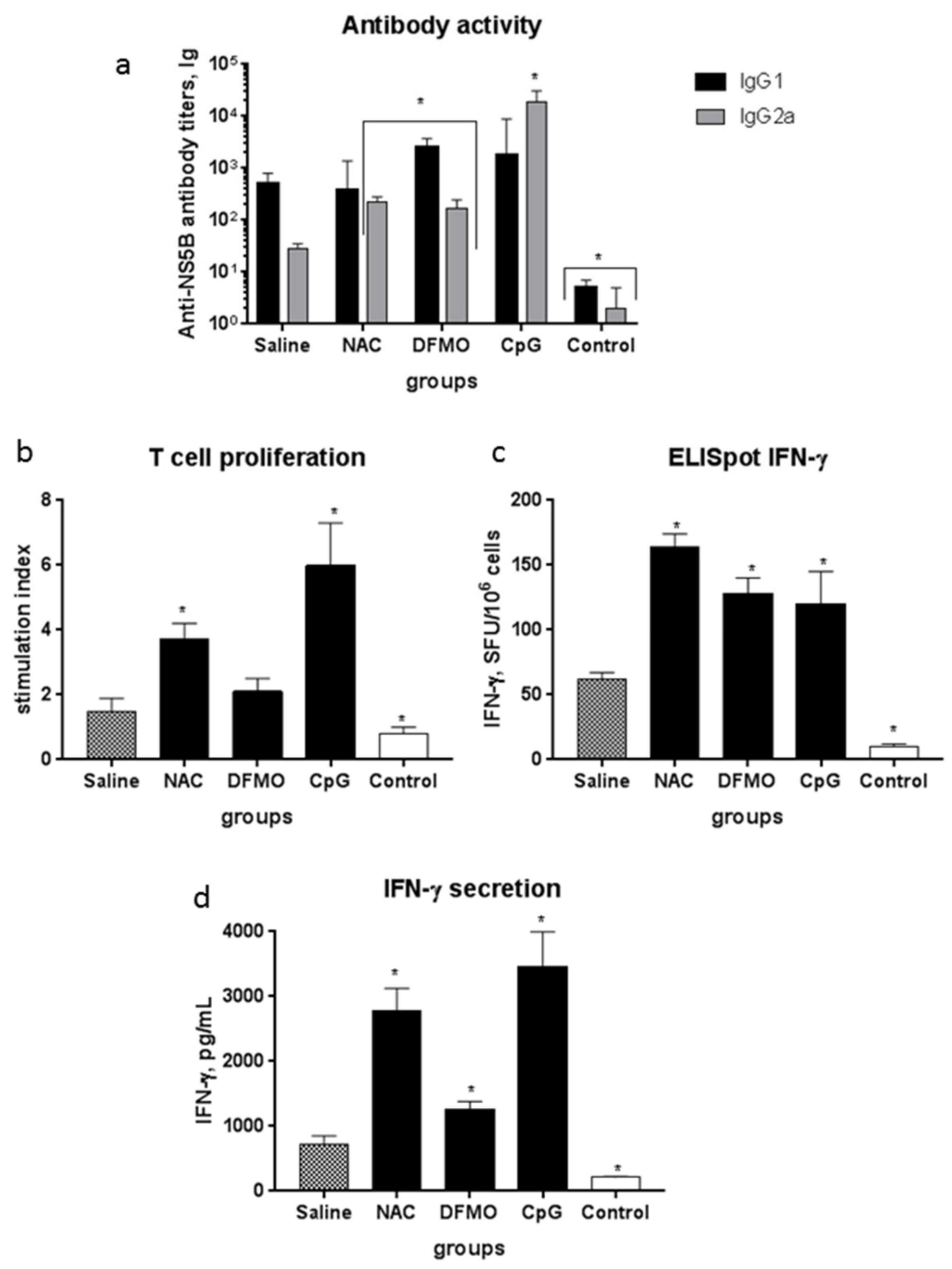
2.3. NAC and DFMO Potentiate Immune Response of Mice to the HCV NS5B Protein
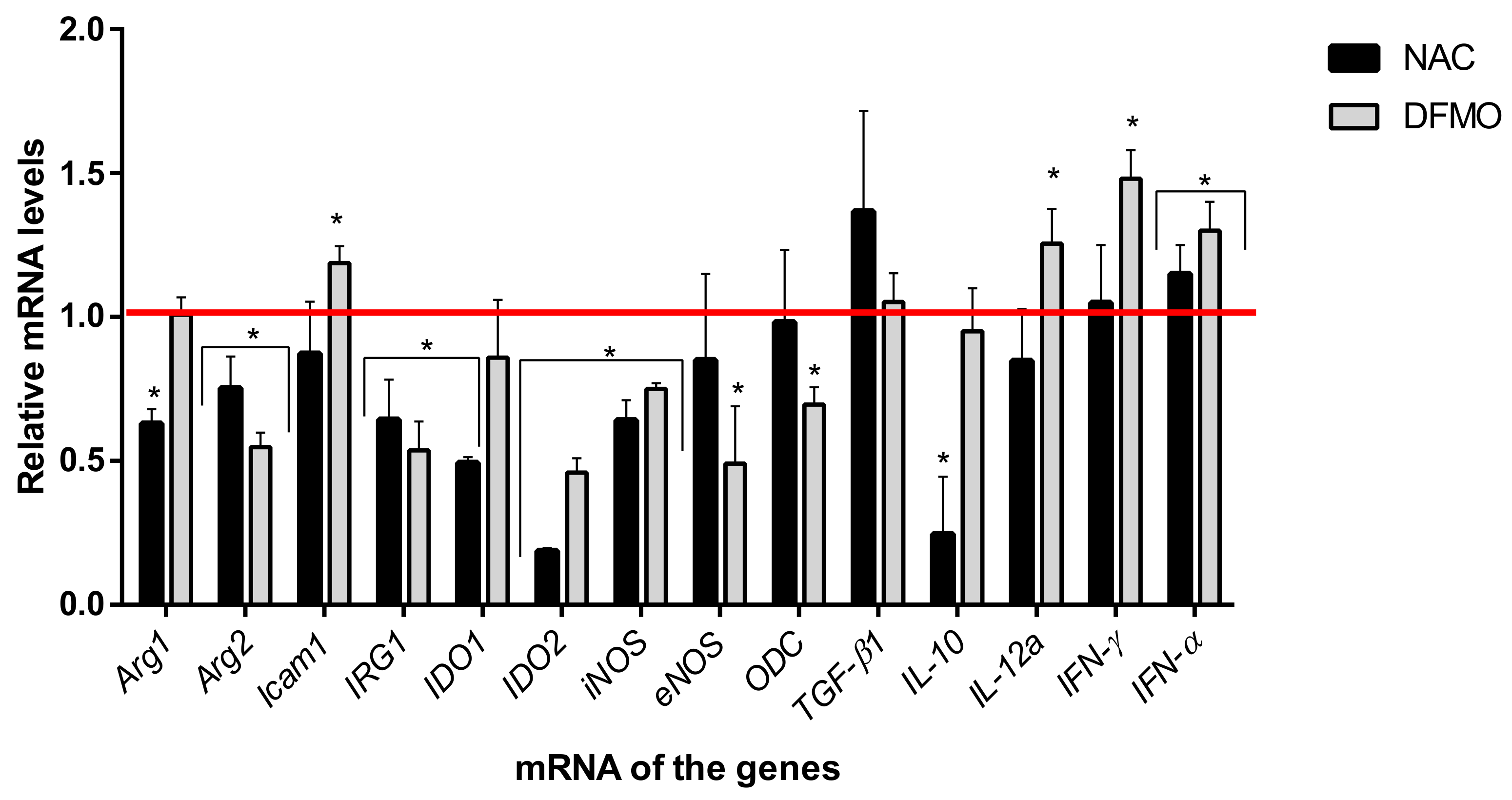
2.4. Immunoregulatory Properties of the Compounds
3. Discussion
4. Materials and Methods
4.1. The Immunogen
4.2. The Compounds
4.3. Mice
4.4. Lymphocyte Proliferation Assay
4.5. Mouse Immunization
4.6. Increasing the Level of Suppressive Cells in the Spleen
4.7. Humoral Immune Response
4.8. T Cell Proliferation and ELISpot Assays
4.9. Cytokine Quantification
4.10. Flow Cytometry
4.11. Arginase Activity Assay
4.12. Nitric Oxide Concentrations
4.13. Quantification of mRNA Levels
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Park, S.H.; Rehermann, B. Immune responses to HCV and other Hepatitis viruses. Immunity 2014, 40, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dustin, L.B. Innate and adaptive immune responses in chronic HCV infection. Curr. Drug Targets 2017, 18, 826–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Rao, H.; Chi, X.; Li, B.; Liu, H.; Wu, L.; Zhang, H.; Liu, S.; Zhou, G.; Li, N.; et al. Detection of residual HCV-RNA in patients who have achieved sustained virological response is associated with persistent histological abnormality. EBioMedicine 2019, 46, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmasry, S.; Wadhwa, S.; Bang, B.R.; Cook, L.; Chopra, S.; Kanel, G.; Kim, B.; Harper, T.; Feng, Z.; Jerome, K.R.; et al. Detection of occult Hepatitis C virus infection in patients who achieved a sustained virologic response to direct-acting antiviral agents for recurrent infection after liver transplantation. Gastroenterology 2017, 152, 550–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, J.A.; Yu, M.L.; Chung, R.T. Hepatitis B reactivation during or after direct acting antiviral therapy—Implication for susceptible individuals. Exp. Opin. Drug Saf. 2017, 16, 651–672. [Google Scholar] [CrossRef]
- Ghweil, A.A.; Helal, M.M. Reactivation of herpesvirus in patients with Hepatitis C treated with direct-acting antiviral agents. Infect. Drug Resis. 2019, 12, 759–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.F.; Yu, M.L. Unmet needs of chronic Hepatitis C in the era of direct-acting antiviral therapy. Clin. Mol. Hepatol. 2020, 26, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Todryk, S.M.; Bassendine, M.F.; Bridge, S.H. Revisiting the elusive Hepatitis C vaccine. Vaccines 2021, 9, 114. [Google Scholar] [CrossRef]
- Yokokawa, H.; Higashino, A.; Suzuki, S.; Moriyama, M.; Nakamura, N.; Suzuki, T.; Suzuki, R.; Ishii, K.; Kobiyama, K.; Ishii, K.J.; et al. Induction of humoural and cellular immunity by immunisation with HCV particle vaccine in a non-human primate model. Gut 2018, 67, 372–379. [Google Scholar] [CrossRef]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of adjuvanticity: A review on adjuvants in licensed vaccines. Semin. Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef]
- Bailey, J.R.; Barnes, E.; Cox, A.L. Approaches, progress, and challenges to Hepatitis C vaccine development. Gastroenterology 2019, 156, 418–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetta, H.F.; Zahran, A.M.; Mansor, S.G.; Abdel-Malek, M.O.; Mekky, M.A.; Abbas, W.A. Frequency and implications of myeloid-derived suppressor cells and lymphocyte subsets in Egyptian patients with Hepatitis C virus-related hepatocellular carcinoma. J. Med. Virol. 2019, 91, 1319–1328. [Google Scholar] [CrossRef]
- Pang, X.; Song, H.; Zhang, Q.; Tu, Z.; Niu, J. Hepatitis C virus regulates the production of monocytic myeloid-derived suppressor cells from peripheral blood mononuclear cells through PI3K pathway and autocrine signaling. Clin. Immunol. 2016, 164, 57–64. [Google Scholar] [CrossRef]
- Zhai, N.; Li, H.; Song, H.; Yang, Y.; Cui, A.; Li, T.; Niu, J.; Crispe, I.N.; Su, L.; Tu, Z. Hepatitis C virus induces MDSCs-like monocytes through TLR2/PI3K/AKT/STAT3 signaling. PLoS ONE 2017, 12, e0170516. [Google Scholar] [CrossRef] [PubMed]
- Telatin, V.; Nicoli, F.; Frasson, C.; Menegotto, N.; Barbaro, F.; Castelli, E.; Erne, E.; Palu, G.; Caputo, A. In chronic Hepatitis C infection, myeloid-derived suppressor cell accumulation and T cell dysfunctions revert partially and late after successful direct-acting antiviral treatment. Front. Cell. Infect. Microbiol. 2019, 9, 190. [Google Scholar] [CrossRef] [Green Version]
- Marvel, D.; Gabrilovich, D.I. Myeloid-derived suppressor cells in the tumor microenvironment: Expect the unexpected. J. Clin. Investig. 2015, 125, 3356–3364. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Jiang, M.; Yuan, H.; Ni, W.; Tai, G. Dual roles of myeloid-derived suppressor cells induced by toll-like receptor signaling in cancer. Oncol. Lett. 2021, 21, 149. [Google Scholar] [CrossRef]
- Zoglmeier, C.; Bauer, H.; Noerenberg, D.; Wedekind, G.; Bittner, P.; Sandholzer, N.; Rapp, M.; Anz, D.; Endres, S.; Bourquin, C. CpG blocks immunosuppression by myeloid-derived suppressor cells in tumor-bearing mice. Clin. Cancer Res. 2011, 17, 1765–1775. [Google Scholar] [CrossRef] [Green Version]
- Chiodetti, A.L.; Sanchez Vallecillo, M.F.; Dolina, J.S.; Crespo, M.I.; Marin, C.; Schoenberger, S.P.; Allemandi, D.A.; Palma, S.D.; Pistoresi-Palencia, M.C.; Moron, G.; et al. Class-B CpG-ODN formulated with a nanostructure induces type I interferons-dependent and CD4(+) T cell-independent CD8(+) T-cell response against unconjugated protein antigen. Front. Immunol. 2018, 9, 2319. [Google Scholar] [CrossRef]
- Sugauchi, F.; Wang, R.Y.; Qiu, Q.; Jin, B.; Alter, H.J.; Shih, J.W. Vigorous Hepatitis C virus-specific CD4+ and CD8+ T cell responses induced by protein immunization in the presence of montanide ISA720 plus synthetic oligodeoxynucleotides containing immunostimulatory cytosine-guanine dinucleotide motifs. J. Infect. Dis. 2006, 193, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Yan, M.; Tang, Y.; Diao, Y. Evaluation of immunogenicity and protective efficacy of a CpG-adjuvanted DNA vaccine against Tembusu virus. Vet. Immunol. Immunopathol. 2019, 218, 109953. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Kagan, J.C. Microbe-inducible trafficking pathways that control Toll-like receptor signaling. Traffic 2017, 18, 6–17. [Google Scholar] [CrossRef]
- Honda, K.; Ohba, Y.; Yanai, H.; Negishi, H.; Mizutani, T.; Takaoka, A.; Taya, C.; Taniguchi, T. Spatiotemporal regulation of MyD88-IRF-7 signalling for robust type-I interferon induction. Nature 2005, 434, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Foy, E.; Ferreon, J.C.; Nakamura, M.; Ferreon, A.C.; Ikeda, M.; Ray, S.C.; Gale, M., Jr.; Lemon, S.M. Immune evasion by Hepatitis C virus NS3/4A protease-mediated cleavage of the Toll-like receptor 3 adaptor protein TRIF. Proc. Natl. Acad. Sci. USA 2005, 102, 2992–2997. [Google Scholar] [CrossRef] [Green Version]
- Ghanim, H.; Mohanty, P.; Deopurkar, R.; Sia, C.L.; Korzeniewski, K.; Abuaysheh, S.; Chaudhuri, A.; Dandona, P. Acute modulation of toll-like receptors by insulin. Diabetes Care 2008, 31, 1827–1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Kumbhare, A.; Youssef, D.; McCall, C.E.; El Gazzar, M. Intracellular S100A9 promotes myeloid-derived suppressor cells during late sepsis. Front. Immunol. 2017, 8, 1565. [Google Scholar] [CrossRef] [Green Version]
- Pollizzi, K.N.; Patel, C.H.; Sun, I.H.; Oh, M.H.; Waickman, A.T.; Wen, J.; Delgoffe, G.M.; Powell, J.D. mTORC1 and mTORC2 selectively regulate CD8(+) T cell differentiation. J. Clin. Investig. 2015, 125, 2090–2108. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Pruitt, M.; Tran, D.; Du Bois, W.; Zhang, K.; Patel, R.; Hoover, S.; Simpson, R.M.; Simmons, J.; Gary, J.; et al. B cell-specific deficiencies in mTOR limit humoral immune responses. J. Immunol. 2013, 191, 1692–1703. [Google Scholar] [CrossRef] [Green Version]
- Keating, R.; McGargill, M.A. mTOR regulation of lymphoid cells in immunity to pathogens. Front. Immunol. 2016, 7, 180. [Google Scholar] [CrossRef]
- Sanchez-Jimenez, F.; Medina, M.A.; Villalobos-Rueda, L.; Urdiales, J.L. Polyamines in mammalian pathophysiology. Cell. Mol. Life Sci. 2019, 76, 3987–4008. [Google Scholar] [CrossRef]
- Latour, Y.L.; Gobert, A.P.; Wilson, K.T. The role of polyamines in the regulation of macrophage polarization and function. Amino Acids 2020, 52, 151–160. [Google Scholar] [CrossRef]
- Ye, C.; Geng, Z.; Dominguez, D.; Chen, S.; Fan, J.; Qin, L.; Long, A.; Zhang, Y.; Kuzel, T.M.; Zhang, B. Targeting ornithine decarboxylase by alpha-difluoromethylornithine inhibits tumor growth by impairing myeloid-derived suppressor cells. J. Immunol. 2016, 196, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Alexander, E.T.; Minton, A.; Peters, M.C.; Phanstiel, O.; Gilmour, S.K. A novel polyamine blockade therapy activates an anti-tumor immune response. Oncotarget 2017, 8, 84140–84152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khomich, O.A.; Kochetkov, S.N.; Bartosch, B.; Ivanov, A.V. Redox biology of respiratory viral infections. Viruses 2018, 10, 392. [Google Scholar] [CrossRef] [Green Version]
- Poe, F.L.; Corn, J. N-acetylcysteine: A potential therapeutic agent for SARS-CoV-2. Med. Hypotheses 2020, 143, 109862. [Google Scholar] [CrossRef] [PubMed]
- Chiba, T.; Takahashi, S.; Sato, N.; Ishii, S.; Kikuchi, K. Fas-mediated apoptosis is modulated by intracellular glutathione in human T cells. Eur. J. Immunol. 1996, 26, 1164–1169. [Google Scholar] [CrossRef]
- Zhang, Q.; Ju, Y.; Ma, Y.; Wang, T. N-acetylcysteine improves oxidative stress and inflammatory response in patients with community acquired pneumonia: A randomized controlled trial. Medicine 2018, 97, e13087. [Google Scholar] [CrossRef]
- Costa, F.; Sousa, D.M.; Parreira, P.; Lamghari, M.; Gomes, P.; Martins, M.C.L. N-acetylcysteine-functionalized coating avoids bacterial adhesion and biofilm formation. Sci. Rep. 2017, 7, 17374. [Google Scholar] [CrossRef] [Green Version]
- Lopez Ballester, J.A.; Penafiel, R.; Cremades, A.; Valcarcel, M.M.; Solano, F.; Lozano, J.A. Effects of treatments with alpha-difluoromethylornithine and hyperthermia on the growth and polyamine metabolism of Harding-Passey murine melanoma. Anticancer Res. 1991, 11, 691–696. [Google Scholar]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a vaccine adjuvant. Exp. Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Che, Y.; Zhao, Y.; Wang, X. Prevention and treatment of cervical cancer by a single administration of human papillomavirus peptide vaccine with CpG oligodeoxynucleotides as an adjuvant in vivo. Int. Immunopharmacol. 2019, 69, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Amaral, M.P.; Apostolico, J.S.; Tomita, N.; Coirada, F.C.; Lunardelli, V.A.S.; Fernandes, E.R.; Souza, H.F.S.; Astray, R.M.; Boscardin, S.B.; Rosa, D.S. Homologous prime-boost with Zika virus envelope protein and poly (I:C) induces robust specific humoral and cellular immune responses. Vaccine 2020, 38, 3653–3664. [Google Scholar] [CrossRef] [PubMed]
- Naarding, M.A.; Falkowska, E.; Xiao, H.; Dragic, T. Hepatitis C virus soluble E2 in combination with QuilA and CpG ODN induces neutralizing antibodies in mice. Vaccine 2011, 29, 2910–2917. [Google Scholar] [CrossRef]
- Ehrlich, A.K.; Fernandez, O.L.; Rodriguez-Pinto, D.; Castilho, T.M.; Corral Caridad, M.J.; Goldsmith-Pestana, K.; Saravia, N.G.; McMahon-Pratt, D. Local delivery of the toll-like receptor 9 ligand CpG downregulates host immune and inflammatory responses, ameliorating established Leishmania (Viannia) panamensis chronic infection. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [Green Version]
- Wingender, G.; Garbi, N.; Schumak, B.; Jungerkes, F.; Endl, E.; von Bubnoff, D.; Steitz, J.; Striegler, J.; Moldenhauer, G.; Tuting, T.; et al. Systemic application of CpG-rich DNA suppresses adaptive T cell immunity via induction of IDO. Eur. J. Immunol. 2006, 36, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Krogmann, A.O.; Lusebrink, E.; Steinmetz, M.; Asdonk, T.; Lahrmann, C.; Lutjohann, D.; Nickenig, G.; Zimmer, S. Proinflammatory stimulation of toll-like receptor 9 with high dose CpG ODN 1826 impairs endothelial regeneration and promotes atherosclerosis in mice. PLoS ONE 2016, 11, e0146326. [Google Scholar] [CrossRef]
- Krieg, A.M. Therapeutic potential of toll-like receptor 9 activation. Nat. Rev. Drug Discov. 2006, 5, 471–484. [Google Scholar] [CrossRef]
- Mutwiri, G.; van Drunen Littel-van den Hurk, S.; Babiuk, L.A. Approaches to enhancing immune responses stimulated by CpG oligodeoxynucleotides. Adv. Drug Deliv. Rev. 2009, 61, 226–232. [Google Scholar] [CrossRef]
- Hanagata, N. CpG oligodeoxynucleotide nanomedicines for the prophylaxis or treatment of cancers, infectious diseases, and allergies. Int. J. Nanomed. 2017, 12, 515–531. [Google Scholar] [CrossRef] [Green Version]
- Hammerich, L.; Tacke, F. Emerging roles of myeloid derived suppressor cells in hepatic inflammation and fibrosis. World J. Gastrointest. Pathophysiol. 2015, 6, 43–50. [Google Scholar] [CrossRef]
- Nonnenmann, J.; Stirner, R.; Roider, J.; Jung, M.C.; Schrodl, K.; Bogner, J.R.; Draenert, R. Lack of significant elevation of myeloid-derived suppressor cells in peripheral blood of chronically Hepatitis C virus-infected individuals. J. Virol. 2014, 88, 7678–7682. [Google Scholar] [CrossRef] [Green Version]
- Moreira, L.O.; Smith, A.M.; DeFreitas, A.A.; Qualls, J.E.; El Kasmi, K.C.; Murray, P.J. Modulation of adaptive immunity by different adjuvant-antigen combinations in mice lacking Nod2. Vaccine 2008, 26, 5808–5813. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.K. Freund adjuvant induces TLR2 but not TLR4 expression in the liver of mice. Int. Immunopharmacol. 2003, 3, 115–118. [Google Scholar] [CrossRef]
- Lee, J.M.; Seo, J.H.; Kim, Y.J.; Kim, Y.S.; Ko, H.J.; Kang, C.Y. The restoration of myeloid-derived suppressor cells as functional antigen-presenting cells by NKT cell help and all-trans-retinoic acid treatment. Int. J. Cancer 2012, 131, 741–751. [Google Scholar] [CrossRef]
- Bui, T.M.; Wiesolek, H.L.; Sumagin, R. ICAM-1: A master regulator of cellular responses in inflammation, injury resolution, and tumorigenesis. J. Leukoc. Biol. 2020, 108, 787–799. [Google Scholar] [CrossRef]
- Lampropoulou, V.; Sergushichev, A.; Bambouskova, M.; Nair, S.; Vincent, E.E.; Loginicheva, E.; Cervantes-Barragan, L.; Ma, X.; Huang, S.C.; Griss, T.; et al. Itaconate links inhibition of succinate dehydrogenase with macrophage metabolic remodeling and regulation of inflammation. Cell Metab. 2016, 24, 158–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, C.P.; Maghzal, G.J.; Ayer, A.; Talib, J.; Giltrap, A.M.; Shengule, S.; Wolhuter, K.; Wang, Y.; Chadha, P.; Suarna, C.; et al. Singlet molecular oxygen regulates vascular tone and blood pressure in inflammation. Nature 2019, 566, 548–552. [Google Scholar] [CrossRef] [Green Version]
- Marthandan, S.; Hyland, P.; Pawelec, G.; Barnett, Y. An investigation of the effects of the antioxidants, ebselen or N-acetyl cysteine on human peripheral blood mononuclear cells and T cells. Immun. Ageing 2013, 10, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Flora, S.; Balansky, R.; La Maestra, S. Rationale for the use of N-acetylcysteine in both prevention and adjuvant therapy of COVID-19. FASEB J. 2020, 34, 13185–13193. [Google Scholar] [CrossRef]
- Mao, B.; Wang, Z.; Pi, S.; Long, Q.; Chen, K.; Cui, J.; Huang, A.; Hu, Y. Difluoromethylornithine, a decarboxylase 1 inhibitor, suppresses Hepatitis B virus replication by reducing HBc protein levels. Front. Cell. Infect. Microbiol. 2020, 10, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mounce, B.C.; Cesaro, T.; Moratorio, G.; Hooikaas, P.J.; Yakovleva, A.; Werneke, S.W.; Smith, E.C.; Poirier, E.Z.; Simon-Loriere, E.; Prot, M.; et al. Inhibition of polyamine biosynthesis is a broad-spectrum strategy against RNA viruses. J. Virol. 2016, 90, 9683–9692. [Google Scholar] [CrossRef] [Green Version]
- Smirnova, O.A.; Keinanen, T.A.; Ivanova, O.N.; Hyvonen, M.T.; Khomutov, A.R.; Kochetkov, S.N.; Bartosch, B.; Ivanov, A.V. Hepatitis C virus alters metabolism of biogenic polyamines by affecting expression of key enzymes of their metabolism. Biochem. Biophys. Res. Commun. 2017, 483, 904–909. [Google Scholar] [CrossRef]
- Singh, K.; Coburn, L.A.; Asim, M.; Barry, D.P.; Allaman, M.M.; Shi, C.; Washington, M.K.; Luis, P.B.; Schneider, C.; Delgado, A.G.; et al. Ornithine decarboxylase in macrophages exacerbates colitis and promotes colitis-associated colon carcinogenesis by impairing M1 immune responses. Cancer Res. 2018, 78, 4303–4315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, A.V.; Korovina, A.N.; Tunitskaya, V.L.; Kostyuk, D.A.; Rechinsky, V.O.; Kukhanova, M.K.; Kochetkov, S.N. Development of the system ensuring a high-level expression of Hepatitis C virus nonstructural NS5B and NS5A proteins. Protein Expr. Purif. 2006, 48, 14–23. [Google Scholar] [CrossRef]
- Masalova, O.V.; Lesnova, E.I.; Klimova, R.R.; Momotyuk, E.D.; Kozlov, V.V.; Ivanova, A.M.; Payushina, O.V.; Butorina, N.N.; Zakirova, N.F.; Narovlyansky, A.N.; et al. Genetically modified mouse mesenchymal stem cells expressing non-structural proteins of Hepatitis C virus induce effective immune response. Vaccines 2020, 8, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saydjari, R.; Alexander, R.W.; Upp, J.R., Jr.; Poston, G.J.; Barranco, S.C.; Townsend, C.M., Jr.; Thompson, J.C. The effect of tumor burden on ornithine decarboxylase activity in mice. Cancer Investig. 1991, 9, 415–419. [Google Scholar] [CrossRef]
- Bosgelmez, I.I.; Guvendik, G. Beneficial effects of N-acetyl-L-cysteine or taurine pre- or post-treatments in the heart, spleen, lung, and testis of hexavalent chromium-exposed mice. Biol. Trace Elem. Res. 2019, 190, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Zhang, H.; Dinavahi, R.; Li, F.; Xiang, Y.; Raman, V.; Bhujwalla, Z.M.; Felsher, D.W.; Cheng, L.; Pevsner, J.; et al. HIF-dependent antitumorigenic effect of antioxidants in vivo. Cancer Cell 2007, 12, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Homma, Y.; Kakizoe, T.; Samma, S.; Oyasu, R. Inhibition of N-butyl-N-(4-hydroxybutyl)nitrosamine-induced rat urinary bladder carcinogenesis by alpha-difluoromethylornithine. Cancer Res. 1987, 47, 6176–6179. [Google Scholar]
- Ranocchia, R.P.; Gorlino, C.V.; Crespo, M.I.; Harman, M.F.; Liscovsky, M.V.; Moron, G.; Maletto, B.A.; Pistoresi-Palencia, M.C. Arginase-dependent suppression by CpG-ODN plus IFA-induced splenic myeloid CD11b(+)Gr1(+) cells. Immunol. Cell Biol. 2012, 90, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Harman, M.F.; Ranocchia, R.P.; Gorlino, C.V.; Sanchez Vallecillo, M.F.; Castell, S.D.; Crespo, M.I.; Maletto, B.A.; Moron, G.; Pistoresi-Palencia, M.C. Expansion of myeloid-derived suppressor cells with arginase activity lasts longer in aged than in young mice after CpG-ODN plus IFA treatment. Oncotarget 2015, 6, 13448–13461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masalova, O.V.; Lesnova, E.I.; Shingarova, L.N.; Tunitskaia, V.L.; Ulanova, T.I.; Burkov, A.N.; Kushch, A.A. The combined application of nucleotide and amino acid sequences of NS3 Hepatitis C virus protein, DNA encoding granulocyte macrophage colony-stimulating factor, and inhibitor of regulatory T cells induces effective immune responce against Hepatitis C virus. Mol. Biol. 2012, 46, 473–480. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | Abbreviation | Supplier | Concentration 3 | Target, Functions |
|---|---|---|---|---|---|
| 1 | N-acetylcysteine | NAC | Sigma | 10 mM | Antioxidant |
| 2 | All-trans retinoic acid | ATRA | Sigma | 0.5 µM | Vitamin A metabolite; exerts potent effects on cell growth, differentiation, and apoptosis |
| 3 | Synthetic oligonucleotides bearing unmethylated CpG motifs | CpG ODN 1826, CpG | Invivogen | 2.5 µM | TLR9 agonist, class B |
| 4 | CpG ODN 2395 | Invivogen | 2.5 µM | TLR9 agonist, class C | |
| 5 | 2-D,L-difluoromethylornithine | DFMO | 1 | 10 mM | Irreversible inhibitor of ornithine decarboxylase (ODC) |
| 6 | N1,N11-diethylnorspermine | DenSpm | 2 | 20 µM | Inducer of spermine/spermidine N1-acetyltransferase (SSAT) |
| 7 | N,N1-bis(2,3-butadienyl)-1,4-diaminobutane | MDL 72527, MDL | Sigma | 20 µM | Inhibitor of polyamine oxidases |
| 8 | N1-cyclopropylmethyl-N11-ethylnorspermine | CPENS | 2 | 10 µM | SSAT inducer |
| 9 | N1-cycloheptylmethyl-N11-ethylnorspermine | CHENS | 2 | 10 µM | SSAT inducer that does not affect polyamine levels |
| 10 | Rapamycin | Rapamycin | Cell Signaling Technology | 250 nM | Specific mTOR inhibitor (mTORC1) |
| 11 | Chloroquine | Chloroquine | Sigma | 20 µM | Inhibitor of autophagy, exhibits the anti-inflammatory effect |
| 12 | Nω-hydroxy-nor-arginine | NOHA | Sigma | 0.1 mM | Arginase inhibitor |
| 13 | Antibodies to Tregs | anti-CD152 | BD Biosciences | 1 µg/mL | CTLA4 antigen expressed in Tregs |
| 14 | Antibodies to granulocytes Ly-6G and Ly-6C (Gr-1) | anti-Gr-1 | BD | 1 µg/mL | Gr-1 antigen expressed on monocytes, neutrophils, and granulocytes |
| 15 | Antibodies to CD11b | anti-CD11b | BD | 1 µg/mL | Antigen expressed on monocytes, macrophages, NK cells, and granulocytes |
| 16 | Antibodies to CD3e | anti-CD3 | BD | 1 µg/mL | Antigen expressed on T cells |
| Experiment Series | Groups (Compounds) | Recombinant NS5B, 4 µg/Mouse | Compound Delivery Method | Dose of Compounds |
|---|---|---|---|---|
| 1 | Saline | IP 1 in a mixture with compounds | IP | No |
| NAC | IP | 4 mg/mouse | ||
| DFMO | IP | 4 mg/mouse | ||
| Control | No | No | No | |
| 2 | Saline | SC 2 | No | No |
| NAC | SC | 40-mM solution in drinking water | 20 mg/mouse/day (1 g/kg body weight /day) | |
| DFMO | SC | 0.2% solution in drinking water | 6 mg/mouse/day (0.3 g/kg body weight/day) | |
| CpG | SC in a mixture with CpG | SC | 50 µg/mouse | |
| Control | No | No | No |
| Gene | Primer Sequence (5′–3′) | |
|---|---|---|
| Direct | Reverse | |
| ODC (ornithine decarboxylase) | CCTTGTGAGGAGCTGGTGATA | GGTCCAGAATGTCCTTAGCAGT |
| Arg1 (arginase 1) | GAAGACTAGAGCCATGCGCC | TTTGAGAAAGGCGCTCCGAT |
| Arg2 (arginase 2) | TCTCCTCCACGGGCAAATTC | GCAAGCCAGCTTCTCGAATG |
| iNOS (inducible NO synthase) | CTATGGCCGCTTTGATGTGC | TTGGGATGCTCCATGGTCAC |
| eNOS (endothelial NO synthase) | ATTGGCATGAGGGACCTGTG | GGGATGAGGTTGTCCTGGTG |
| IFN-α (interferon alpha) | CTACTGGCCAACCTGCTCTC | CTGCGGGAATCCAAAGTCCT |
| IFN-γ (interferon gamma) | AGCAAGGCGAAAAAGGATGC | TCATTGAATGCTTGGCGCTG |
| TGF-β1 (transforming growth factor beta 1) | AGCTGCGCTTGCAGAGATTA | GTATCAGTGGGGGTCAGCAG |
| IL-10 (interleukin 10) | CAGAGAAGCATGGCCCAGAA | GCTCCACTGCCTTGCTCTTA |
| IL-12a (interleukin 12a) | CCGAAACCTGCTGAAGACCA | GGTTTGGTCCCGTGTGATGT |
| IDO1 (indolamine-2,3-dioxygenase 1) | TGGTGGAAATCGCAGCTTCT | TGCAGTGCCTTTTCCAATGC |
| IDO2 (indoleamine-2,3-dioxygenase 2) | TGCCATGCTGAGCTTCTTGA | ACTGCTAAGCACCAGGACAC |
| ICAM-1 (intercellular adhesion molecule 1) | CAGCTGCTGCTGCTTTTGAA | AGGCTACAAGTGTGCATCCC |
| IRG1 (immune-responsive gene 1) | TGAGCCAGTTACCCTCCAGA | TCATCCTCTTGCTCCTCCGA |
| GUS (glucuronidase-β) | CCAGAGCGAGTATGGAGCAG | CAGATGAGCTCTCCGACCAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lesnova, E.I.; Masalova, O.V.; Permyakova, K.Y.; Kozlov, V.V.; Nikolaeva, T.N.; Pronin, A.V.; Valuev-Elliston, V.T.; Ivanov, A.V.; Kushch, A.A. Difluoromethylornithine (DFMO), an Inhibitor of Polyamine Biosynthesis, and Antioxidant N-Acetylcysteine Potentiate Immune Response in Mice to the Recombinant Hepatitis C Virus NS5B Protein. Int. J. Mol. Sci. 2021, 22, 6892. https://doi.org/10.3390/ijms22136892
Lesnova EI, Masalova OV, Permyakova KY, Kozlov VV, Nikolaeva TN, Pronin AV, Valuev-Elliston VT, Ivanov AV, Kushch AA. Difluoromethylornithine (DFMO), an Inhibitor of Polyamine Biosynthesis, and Antioxidant N-Acetylcysteine Potentiate Immune Response in Mice to the Recombinant Hepatitis C Virus NS5B Protein. International Journal of Molecular Sciences. 2021; 22(13):6892. https://doi.org/10.3390/ijms22136892
Chicago/Turabian StyleLesnova, Ekaterina I., Olga V. Masalova, Kristina Yu. Permyakova, Vyacheslav V. Kozlov, Tatyana N. Nikolaeva, Alexander V. Pronin, Vladimir T. Valuev-Elliston, Alexander V. Ivanov, and Alla A. Kushch. 2021. "Difluoromethylornithine (DFMO), an Inhibitor of Polyamine Biosynthesis, and Antioxidant N-Acetylcysteine Potentiate Immune Response in Mice to the Recombinant Hepatitis C Virus NS5B Protein" International Journal of Molecular Sciences 22, no. 13: 6892. https://doi.org/10.3390/ijms22136892
APA StyleLesnova, E. I., Masalova, O. V., Permyakova, K. Y., Kozlov, V. V., Nikolaeva, T. N., Pronin, A. V., Valuev-Elliston, V. T., Ivanov, A. V., & Kushch, A. A. (2021). Difluoromethylornithine (DFMO), an Inhibitor of Polyamine Biosynthesis, and Antioxidant N-Acetylcysteine Potentiate Immune Response in Mice to the Recombinant Hepatitis C Virus NS5B Protein. International Journal of Molecular Sciences, 22(13), 6892. https://doi.org/10.3390/ijms22136892