
Curcumin Potentiates α7 Nicotinic Acetylcholine Receptors and Alleviates Autistic-Like Social Deficits and Brain Oxidative Stress Status in Mice

 ,
,  and
and
Abstract
:1. Introduction
2. Results
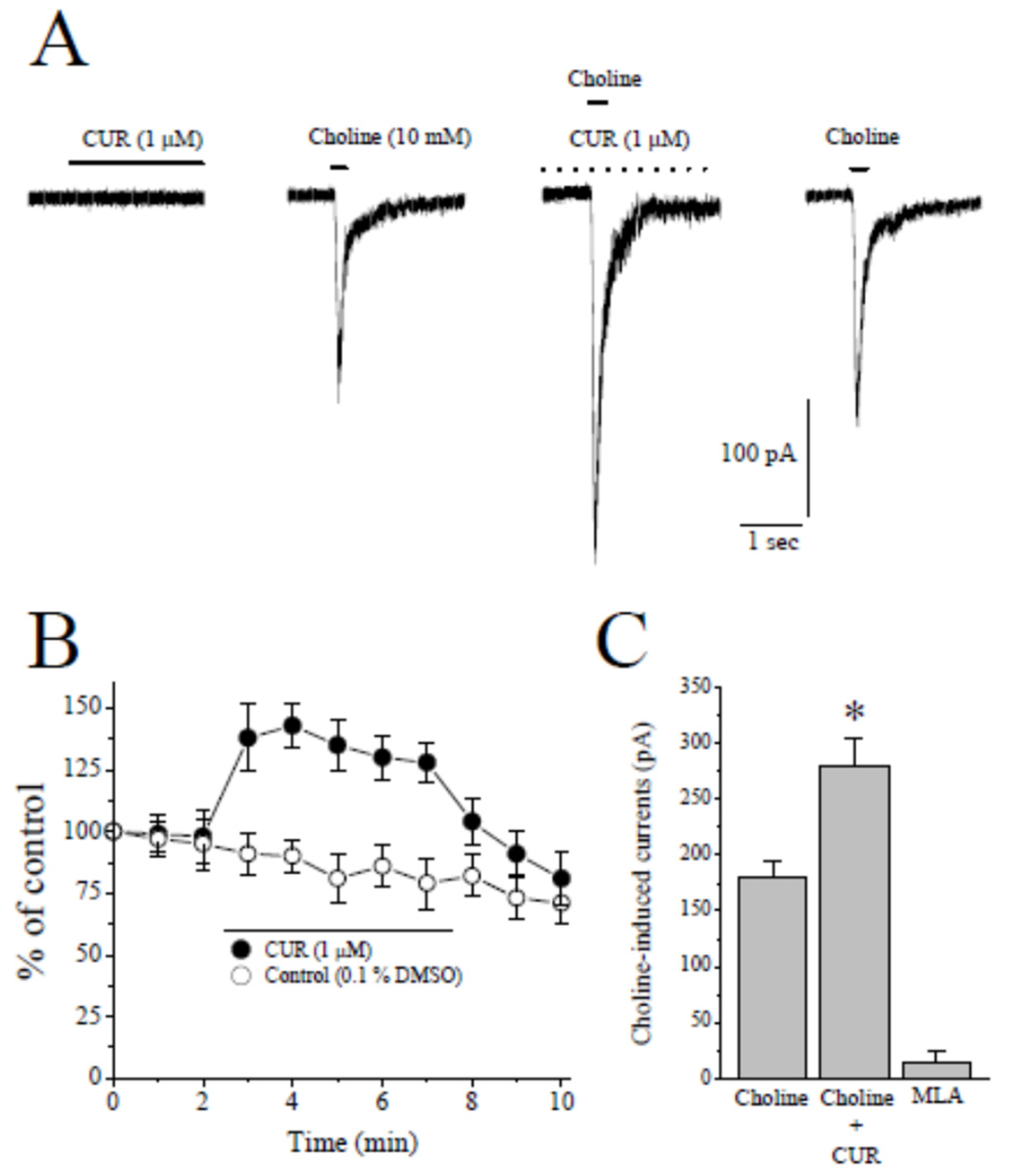
2.1. Effects of CUR on α7-nACh Receptors in the CA1 Region of Stratum Radiatum Interneurons in Rat Hippocampal Brain Slices
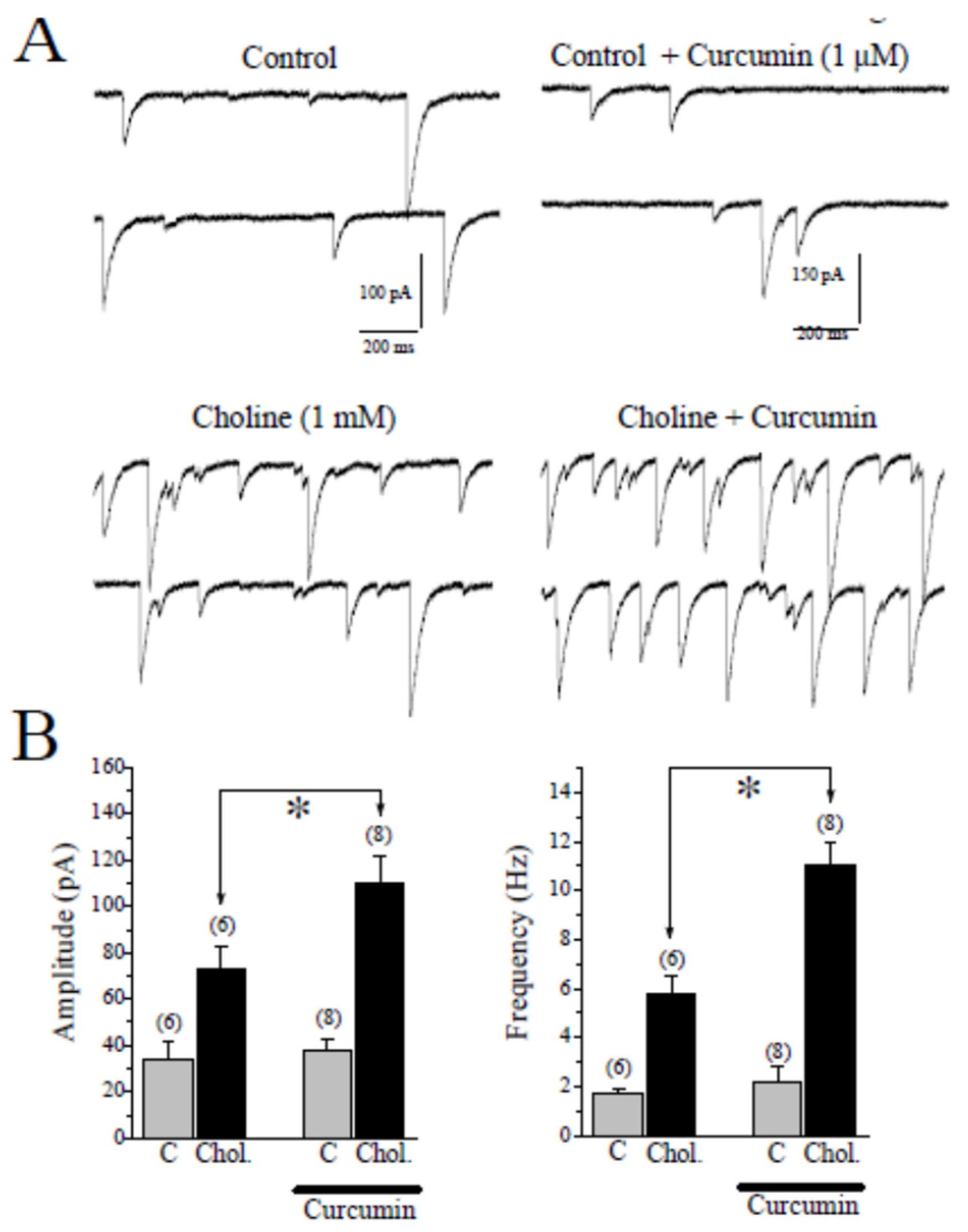
2.2. Effects of CUR on Choline-Induced GABA Responses in CA1 Pyramidal Neurons of Hippocampal Slices
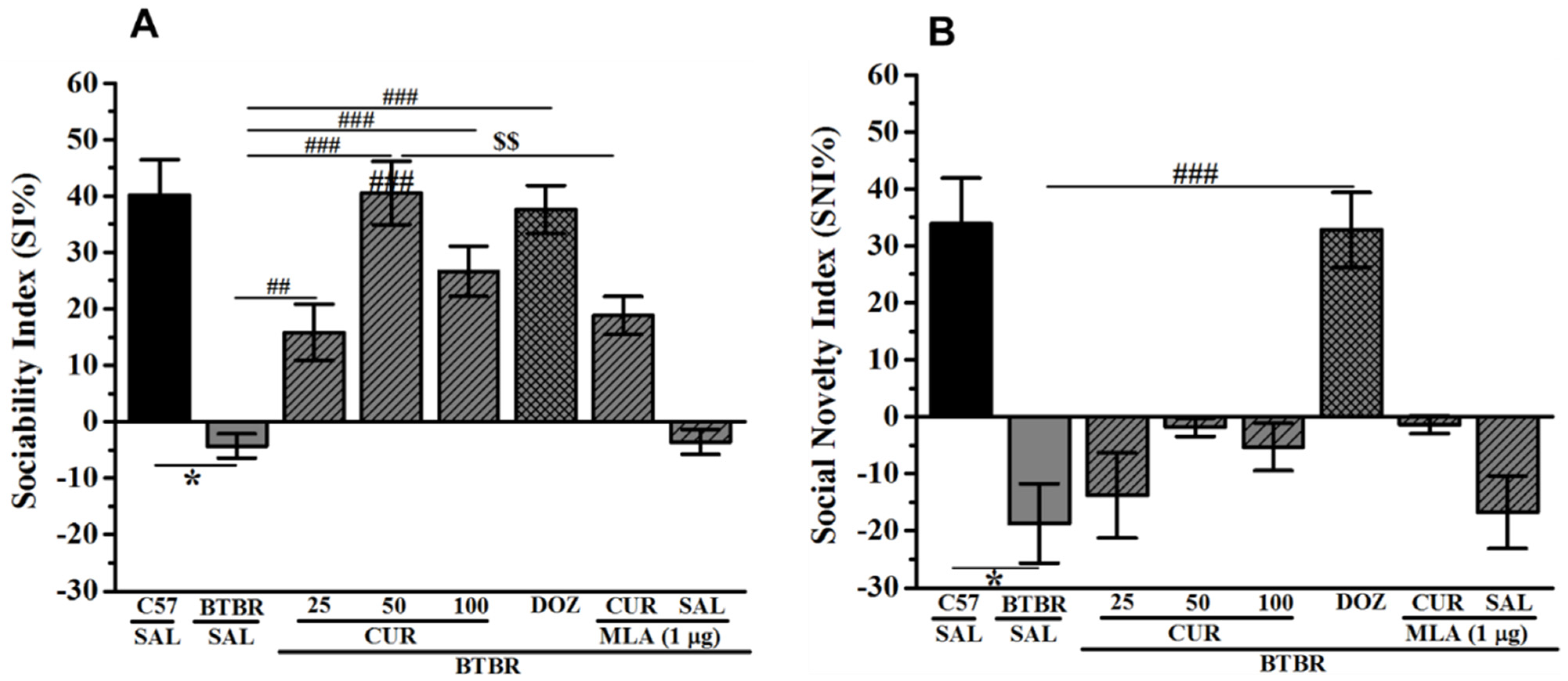
2.3. Effects of CUR and DOZ on Sociability and Social Novelty Deficits of BTBR Mice in TCP
2.4. Effects of CUR and DOZ on Locomotor Activity and Anxiety Levels in OFT
2.5. Effects of CUR and DOZ on Levels of Oxidative Stress Markers in the Hippocampus and the Cerebellum of Treated BTBR Mice
3. Discussion
4. Materials and Methods
4.1. Recordings from Hippocampal Slices
4.2. In-Vivo Behavioral Experiments
4.2.1. Animals
4.2.2. Drugs
4.2.3. Animal Groups and Drug Treatments
4.2.4. Behavioral Tests
Three Chamber Paradigm (TCP)
Open Field Test
4.2.5. Biochemical Assessments
Brain Tissue Collection
Assessment of Antioxidant Enzymes Activity
4.3. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Albuquerque, E.X.; Pereira, E.F.R.; Alkondon, M.; Rogers, S.W. Mammalian Nicotinic Acetylcholine Receptors: From Structure to Function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef] [Green Version]
- Bouzat, C.; Lasala, M.; Nielsen, B.; Corradi, J.; Esandi, M.D.C. Molecular function of α7 nicotinic receptors as drug targets. J. Physiol. 2017, 596, 1847–1861. [Google Scholar] [CrossRef]
- Echeverria, V.; Yarkov, A.; Aliev, G. Positive modulators of the α7 nicotinic receptor against neuroinflammation and cognitive impairment in Alzheimer’s disease. Prog. Neurobiol. 2016, 144, 142–157. [Google Scholar] [CrossRef]
- Oz, M.; El Nebrisi, E.G.; Yang, K.-H.S.; Howarth, F.C.; Al Kury, L.T. Cellular and Molecular Targets of Menthol Actions. Front. Pharmacol. 2017, 8, 472. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Almeida, L.E.F.; Spornick, N.A.; Kenyon, N.; Kamimura, S.; Khaibullina, A.; Nouraie, M.; Quezado, Z.M.N. Modulation of social deficits and repetitive behaviors in a mouse model of autism: The role of the nicotinic cholinergic system. Psychopharmacology 2015, 232, 4303–4316. [Google Scholar] [CrossRef] [PubMed]
- Terry, A.V.; Callahan, P.M. α7 nicotinic acetylcholine receptors as therapeutic targets in schizophrenia: Update on animal and clinical studies and strategies for the future. Neuropharmacology 2020, 170, 108053. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, S.I.; Burket, J.A.; Urbano, M.R.; Benson, A.D. The α7 nicotinic acetylcholine receptor: A mediator of pathogenesis and therapeutic target in autism spectrum disorders and Down syndrome. Biochem. Pharmacol. 2015, 97, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, S.I.; Burket, J.A. An Evolving Therapeutic Rationale for Targeting the α7 Nicotinic Acetylcholine Receptor in Autism Spectrum Disorder. Curr. Top. Behav. Neurosci. 2020, 45, 167–208. [Google Scholar] [CrossRef]
- Eissa, N.; Al-Houqani, M.; Sadeq, A.; Ojha, S.K.; Sasse, A.; Sadek, B. Current Enlightenment about Etiology and Pharmacological Treatment of Autism Spectrum Disorder. Front. Neurosci. 2018, 12, 304. [Google Scholar] [CrossRef] [Green Version]
- Eissa, N.; Sadeq, A.; Sasse, A.; Sadek, B. Role of Neuroinflammation in Autism Spectrum Disorder and the Emergence of Brain Histaminergic System. Lessons Also for BPSD? Front. Pharmacol. 2020, 11, 886. [Google Scholar] [CrossRef]
- Lee, M.; Martin-Ruiz, C.; Graham, A.; Court, J.; Jaros, E.; Perry, R.; Iversen, P.; Bauman, M.; Perry, E. Nicotinic receptor abnormalities in the cerebellar cortex in autism. Brain 2002, 125, 1483–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Ruiz, C.; Lee, M.; Perry, R.; Baumann, M.; Court, J.; Perry, E. Molecular analysis of nicotinic receptor expression in autism. Mol. Brain Res. 2004, 123, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Pardo, M.; Beurel, E.; Jope, R.S. Cotinine administration improves impaired cognition in the mouse model of Fragile X syndrome. Eur. J. Neurosci. 2017, 45, 490–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oginsky, M.F.; Cui, N.; Zhong, W.; Johnson, C.M.; Jiang, C. Alterations in the cholinergic system of brain stem neurons in a mouse model of Rett syndrome. Am. J. Physiol. Physiol. 2014, 307, C508–C520. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.-L.; Adams, C.E.; Stevens, K.E.; Chow, K.-H.; Freedman, R.; Patterson, P.H. The interaction between maternal immune activation and alpha 7 nicotinic acetylcholine receptor in regulating behaviors in the offspring. Brain Behav. Immun. 2015, 46, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Lewis, A.S.; Van Schalkwyk, G.I.; Lopez, M.O.; Volkmar, F.R.; Picciotto, M.R.; Sukhodolsky, D.G. An Exploratory Trial of Transdermal Nicotine for Aggression and Irritability in Adults with Autism Spectrum Disorder. J. Autism Dev. Disord. 2018, 48, 2748–2757. [Google Scholar] [CrossRef]
- Bacchelli, E.; Battaglia, A.; Cameli, C.; Lomartire, S.; Tancredi, R.; Thomson, S.; Sutcliffe, J.S.; Maestrini, E. Analysis of CHRNA7 rare variants in autism spectrum disorder susceptibility. Am. J. Med. Genet. 2015, 167, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, S.I.; Burket, J.A.; Benson, A.D.; Urbano, M.R. The 15q13.3 deletion syndrome: Deficient α7-containing nicotinic acetylcholine receptor-mediated neurotransmission in the pathogenesis of neurodevelopmental disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 64, 109–117. [Google Scholar] [CrossRef]
- Gillentine, M.A.; Schaaf, C.P. The human clinical phenotypes of altered CHRNA7 copy number. Biochem. Pharmacol. 2015, 97, 352–362. [Google Scholar] [CrossRef] [Green Version]
- Gillentine, M.A.; Berry, L.N.; Goin-Kochel, R.P.; Ali, M.A.; Ge, J.; Guffey, D.; Rosenfeld, J.A.; Hannig, V.; Bader, P.; Proud, M.; et al. The Cognitive and Behavioral Phenotypes of Individuals with CHRNA7 Duplications. J. Autism Dev. Disord. 2017, 47, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.T.; Shen, Y.; Weiss, L.A.; Korn, J.; Anselm, I.; Bridgemohan, C.; Cox, G.F.; Dickinson, H.; Gentile, J.; Harris, D.J.; et al. Microdeletion/duplication at 15q13.2q13.3 among individuals with features of autism and other neuropsychiatric disorders. J. Med. Genet. 2008, 46, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasui, D.H.; Scoles, H.A.; Horike, S.-I.; Meguro-Horike, M.; Dunaway, K.W.; Schroeder, D.I.; LaSalle, J.M. 15q11.2–13.3 chromatin analysis reveals epigenetic regulation of CHRNA7 with deficiencies in Rett and autism brain. Hum. Mol. Genet. 2011, 20, 4311–4323. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, H.G.; Kusek, G.K.; Yang, M.; Phoenix, J.L.; Bolivar, V.; Crawley, J.N. Autism-like behavioral phenotypes in BTBR T+tf/J mice. Genes Brain Behav. 2007, 7, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Abrams, D.N.; Zhang, J.Y.; Weber, M.D.; Katz, A.M.; Clarke, A.M.; Silverman, J.L.; Crawley, J.N. Low sociability in BTBR T+tf/J mice is independent of partner strain. Physiol. Behav. 2012, 107, 649–662. [Google Scholar] [CrossRef] [Green Version]
- McTighe, S.M.; Neal, S.J.; Lin, Q.; Hughes, Z.A.; Smith, D.G. The BTBR Mouse Model of Autism Spectrum Disorders Has Learning and Attentional Impairments and Alterations in Acetylcholine and Kynurenic Acid in Prefrontal Cortex. PLoS ONE 2013, 8, e62189. [Google Scholar] [CrossRef]
- Yoshimura, R.F.; Tran, M.B.; Hogenkamp, D.J.; Ayala, N.L.; Johnstone, T.; Dunnigan, A.J.; Gee, T.K.; Gee, K.W. Allosteric modulation of nicotinic and GABAA receptor subtypes differentially modify autism-like behaviors in the BTBR mouse model. Neuropharmacology 2017, 126, 38–47. [Google Scholar] [CrossRef]
- Mahmood, H.M.; Aldhalaan, H.M.; Alshammari, T.K.; Alqasem, M.A.; Alshammari, M.A.; Albekairi, N.A.; AlSharari, S.D. The Role of Nicotinic Receptors in the Attenuation of Autism-Related Behaviors in a Murine BTBR T + tf/J Autistic Model. Autism Res. 2020, 13, 1311–1334. [Google Scholar] [CrossRef]
- Ghaleiha, A.; Ghyasvand, M.; Mohammadi, M.-R.; Farokhnia, M.; Yadegari, N.; Tabrizi, M.; Hajiaghaee, R.; Yekehtaz, H.; Akhondzadeh, S. Galantamine efficacy and tolerability as an augmentative therapy in autistic children: A randomized, double-blind, placebo-controlled trial. J. Psychopharmacol. 2014, 28, 677–685. [Google Scholar] [CrossRef]
- Nicolson, R.; Craven-Thuss, B.; Smith, J. A Prospective, Open-Label Trial of Galantamine in Autistic Disorder. J. Child Adolesc. Psychopharmacol. 2006, 16, 621–629. [Google Scholar] [CrossRef]
- Buckley, A.W.; Sassower, K.; Rodriguez, A.J.; Jennison, K.; Wingert, K.; Buckley, J.; Thurm, A.; Sato, S.; Swedo, S. An Open Label Trial of Donepezil for Enhancement of Rapid Eye Movement Sleep in Young Children with Autism Spectrum Disorders. J. Child Adolesc. Psychopharmacol. 2011, 21, 353–357. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.; Prasad, S.; Aggarwal, B.B. Curcumin, the golden nutraceutical: Multitargeting for multiple chronic diseases. Br. J. Pharmacol. 2017, 174, 1325–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2016, 174, 1290–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molino, S.; Dossena, M.; Buonocore, D.; Ferrari, F.; Venturini, L.; Ricevuti, G.; Verri, M. Polyphenols in dementia: From molecular basis to clinical trials. Life Sci. 2016, 161, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Lopresti, A.L. Curcumin for neuropsychiatric disorders: A review of in vitro, animal and human studies. J. Psychopharmacol. 2017, 31, 287–302. [Google Scholar] [CrossRef] [PubMed]
- El Nebrisi, E.G.; Bagdas, D.; Toma, W.; Al Samri, H.; Brodzik, A.; Alkhlaif, Y.; Yang, K.-H.S.; Howarth, F.C.; Damaj, I.M.; Oz, M. Curcumin Acts as a Positive Allosteric Modulator of α7-Nicotinic Acetylcholine Receptors and Reverses Nociception in Mouse Models of Inflammatory Pain. J. Pharmacol. Exp. Ther. 2018, 365, 190–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ximenis, M.; Mulet, J.; Sala, S.; Sala, F.; Criado, M.; González-Muñiz, R.; de Vega, M.J.P. Natural Polyhydroxy Flavonoids, Curcuminoids, and Synthetic Curcumin Analogs as α7 nAChRs Positive Allosteric Modulators. Int. J. Mol. Sci. 2021, 22, 973. [Google Scholar] [CrossRef]
- El Nebrisi, E.; Al Kury, L.T.; Yang, K.-H.S.; Jayaprakash, P.; Howarth, F.C.; Kabbani, N.; Oz, M. Curcumin potentiates the function of human α7-nicotinic acetylcholine receptors expressed in SH-EP1 cells. Neurochem. Int. 2018, 114, 80–84. [Google Scholar] [CrossRef]
- El Nebrisi, E.; Javed, H.; Ojha, S.K.; Oz, M.; Shehab, S. Neuroprotective Effect of Curcumin on the Nigrostriatal Pathway in a 6-Hydroxydopmine-Induced Rat Model of Parkinson’s Disease is Mediated by α7-Nicotinic Receptors. Int. J. Mol. Sci. 2020, 21, 7329. [Google Scholar] [CrossRef]
- Orr-Urtreger, A.; Seldin, M.F.; Baldini, A.; Beaudet, A.L. Cloning and mapping of the mouse α7-neuronal nicotinic acetylcholine receptor. Genomics 1995, 26, 399–402. [Google Scholar] [CrossRef]
- Riedel, G.; Kang, S.; Choi, D.; Platt, B. Scopolamine-induced deficits in social memory in mice: Reversal by donepezil. Behav. Brain Res. 2009, 204, 217–225. [Google Scholar] [CrossRef]
- Bhandari, R.; Kuhad, A. Neuropsychopharmacotherapeutic efficacy of curcumin in experimental paradigm of autism spectrum disorders. Life Sci. 2015, 141, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Picciotto, M.R. Nicotine as a modulator of behavior: Beyond the inverted U. Trends Pharmacol. Sci. 2003, 24, 493–499. [Google Scholar] [CrossRef]
- Kohut, S.J.; Bergman, J. Reinforcing effectiveness of nicotine in nonhuman primates: Effects of nicotine dose and history of nicotine self-administration. Psychopharmacology 2016, 233, 2451–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, N.E.; Min, W.; Hackett, A.; Lowe, D.; Hanania, T.; Caldarone, B.; Ghavami, A. The high-affinity nAChR partial agonists varenicline and sazetidine-A exhibit reinforcing properties in rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 1455–1464. [Google Scholar] [CrossRef]
- Hopkins, T.J.; Rupprecht, L.E.; Hayes, M.R.; Blendy, J.A.; Schmidt, H.D. Galantamine, an Acetylcholinesterase Inhibitor and Positive Allosteric Modulator of Nicotinic Acetylcholine Receptors, Attenuates Nicotine Taking and Seeking in Rats. Neuropsychopharmacology 2012, 37, 2310–2321. [Google Scholar] [CrossRef] [Green Version]
- Friedman, S.D.; Shaw, D.W.W.; Artru, A.A.; Dawson, G.; Petropoulos, H.; Dager, S.R. Gray and White Matter Brain Chemistry in Young Children with Autism. Arch. Gen. Psychiatry 2006, 63, 786–794. [Google Scholar] [CrossRef]
- Mukaetova-Ladinska, E.B. Silent lives: Why do we fail community-dwelling people with dementia? Age Ageing 2017, 46, 341–343. [Google Scholar] [CrossRef] [Green Version]
- Eissa, N.; Jayaprakash, P.; Stark, H.; Łażewska, D.; Kieć-Kononowicz, K.; Sadek, B. Simultaneous Blockade of Histamine H3 Receptors and Inhibition of Acetylcholine Esterase Alleviate Autistic-Like Behaviors in BTBR T+ tf/J Mouse Model of Autism. Biomolecules 2020, 10, 1251. [Google Scholar] [CrossRef]
- Zhong, H.; Xiao, R.; Ruan, R.; Liu, H.; Li, X.; Cai, Y.; Zhao, J.; Fan, X. Neonatal curcumin treatment restores hippocampal neurogenesis and improves autism-related behaviors in a mouse model of autism. Psychopharmacology 2020, 237, 3539–3552. [Google Scholar] [CrossRef] [PubMed]
- Vecchione, R.; Quagliariello, V.; Calabria, D.; Calcagno, V.; De Luca, E.; Iaffaioli, R.V.; Netti, P.A. Curcumin bioavailability from oil in water nano-emulsions: In Vitro and in vivo study on the dimensional, compositional and interactional dependence. J. Control. Release 2016, 233, 88–100. [Google Scholar] [CrossRef]
- Di Angelantonio, S.; Bernardi, G.; Mercuri, N.B. Donepezil modulates nicotinic receptors of substantia nigra dopaminergic neurones. Br. J. Pharmacol. 2004, 141, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geerts, H.; Guillaumat, P.-O.; Grantham, C.; Bode, W.; Anciaux, K.; Sachak, S. Brain levels and acetylcholinesterase inhibition with galantamine and donepezil in rats, mice, and rabbits. Brain Res. 2005, 1033, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Venkatachalam, K.; Jayaprakash, P.; Falkenstein, M.; Dubiel, M.; Frank, A.; Reiner-Link, D.; Stark, H.; Sadek, B. The Multi-Targeting Ligand ST-2223 with Histamine H3 Receptor and Dopamine D2/D3 Receptor Antagonist Properties Mitigates Autism-Like Repetitive Behaviors and Brain Oxidative Stress in Mice. Int. J. Mol. Sci. 2021, 22, 1947. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.F.; Buxhoeveden, D.; Gomez, J. Disruption in the inhibitory architecture of the cell minicolumn: Implications for autism. Neuroscientist 2003, 9, 496–507. [Google Scholar] [CrossRef]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Łażewska, D.; Kieć-Kononowicz, K.; et al. The dual-active histamine H3 receptor antagonist and acetylcholine esterase inhibitor E100 ameliorates stereotyped repetitive behavior and neuroinflammmation in sodium valproate induced autism in mice. Chem. Biol. Interact. 2019, 312, 108775. [Google Scholar] [CrossRef]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Łażewska, D.; Kieć-Kononowicz, K.; et al. The Dual-Active Histamine H3 Receptor Antagonist and Acetylcholine Esterase Inhibitor E100 Alleviates Autistic-Like Behaviors and Oxidative Stress in Valproic Acid Induced Autism in Mice. Int. J. Mol. Sci. 2020, 21, 3996. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Eissa, N.; Al Awad, M.; Jayaprakash, P.; Zhong, S.; Stölting, F.; Stark, H.; Sadek, B. The histamine H3R and dopamine D2R/D3R antagonist ST-713 ameliorates autism-like behavioral features in BTBR T+tf/J mice by multiple actions. Biomed. Pharmacother. 2021, 138, 111517. [Google Scholar] [CrossRef]
- Al-Askar, M.; Bhat, R.S.; Selim, M.; Al-Ayadhi, L.; El-Ansary, A. Postnatal treatment using curcumin supplements to amend the damage in VPA-induced rodent models of autism. BMC Complement. Altern. Med. 2017, 17, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultan, A.; Yang, K.-H.S.; Isaev, D.; El Nebrisi, E.; Syed, N.; Khan, N.; Howarth, C.F.; Sadek, B.; Oz, M. Thujone inhibits the function of α7-nicotinic acetylcholine receptors and impairs nicotine-induced memory enhancement in one-trial passive avoidance paradigm. Toxicology 2017, 384, 23–32. [Google Scholar] [CrossRef]
- Alzaabi, A.H.; Howarth, L.; El Nebrisi, E.; Syed, N.; Yang, K.-H.S.; Howarth, F.C.; Oz, M. Capsaicin inhibits the function of α7-nicotinic acetylcholine receptors expressed in Xenopus oocytes and rat hippocampal neurons. Eur. J. Pharmacol. 2019, 857, 172411. [Google Scholar] [CrossRef]
- Singhal, S.K.; Zhang, L.; Morales, M.; Oz, M. Antipsychotic clozapine inhibits the function of α7-nicotinic acetylcholine receptors. Neuropharmacology 2007, 52, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Jayaprakash, P.; Azimullah, S.; Ojha, S.K.; Al-Houqani, M.; Jalal, F.Y.; Łażewska, D.; Kiec-Kononowicz, K.; Sadek, B. The histamine H3R antagonist DL77 attenuates autistic behaviors in a prenatal valproic acid-induced mouse model of autism. Sci. Rep. 2018, 8, 13077. [Google Scholar] [CrossRef]
- Eissa, N.; Khan, N.; Ojha, S.K.; Łażewska, D.; Kiec-Kononowicz, K.; Sadek, B. The Histamine H3 Receptor Antagonist DL77 Ameliorates MK801-Induced Memory Deficits in Rats. Front. Neurosci. 2018, 12, 42. [Google Scholar] [CrossRef] [Green Version]
- Babu, A.; Prasanth, K.G.; Balaji, B. Effect of curcumin in mice model of vincristine-induced neuropathy. Pharm. Biol. 2015, 53, 838–848. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.L.; Tolu, S.S.; Barkan, C.L.; Crawley, J.N. Repetitive Self-Grooming Behavior in the BTBR Mouse Model of Autism is Blocked by the mGluR5 Antagonist MPEP. Neuropsychopharmacology 2010, 35, 976–989. [Google Scholar] [CrossRef]
- Khan, N.; Saad, A.; Nurulain, S.M.; Darras, F.H.; Decker, M.; Sadek, B. The dual-acting H3 receptor antagonist and AChE inhibitor UW-MD-71 dose-dependently enhances memory retrieval and reverses dizocilpine-induced memory impairment in rats. Behav. Brain Res. 2016, 297, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Chauhan, V.; Brown, W.T.; Cohen, I. Oxidative stress in autism: Increased lipid peroxidation and reduced serum levels of ceruloplasmin and transferrin—The antioxidant proteins. Life Sci. 2004, 75, 2539–2549. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.B.; Baral, M.; Geis, E.; Mitchell, J.; Ingram, J.; Hensley, A.; Zappia, I.; Newmark, S.; Gehn, E.; Rubin, R.A.; et al. The Severity of Autism Is Associated with Toxic Metal Body Burden and Red Blood Cell Glutathione Levels. J. Toxicol. 2009, 2009, 532640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frances, A.J. Predicting the future of psychiatry. Seishin Shinkeigaku Zasshi 2000, 102, 1203–1208. [Google Scholar]
- Careaga, M.; Schwartzer, J.; Ashwood, P. Inflammatory profiles in the BTBR mouse: How relevant are they to autism spectrum disorders? Brain Behav. Immun. 2015, 43, 11–16. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | Time Spent in Center (s) | Time Spent in Periphery (s) | Total Distance Travelled (cm) |
|---|---|---|---|
| SAL (C57) | 30.64 ± 3.72 | 569.36 ± 5.31 | 2522 ± 187.75 |
| SAL (BTBR) | 62.75 ± 4.18 ** | 537.25 ± 6.19 ** | 4166 ± 168.14 ** |
| CUR (25 mg/kg)/BTBR | 59.04 ± 4.54 | 540.96 ± 10.85 | 3897 ± 128.76 |
| CUR (50 mg/kg)/BTBR | 59.91 ± 5.57 | 536.44 ± 12.82 | 3936 ± 171.15 |
| CUR (100 mg/kg)/BTBR | 58.07 ± 5.79 | 540.87 ± 9.27 | 3787 ± 330.29 |
| DOZ (1 mg/kg)/BTBR | 43.64 ± 3.37 # | 556.36 ± 6.95 | 4066 ± 183.01 |
| CUR(50 mg/kg) + MLA (1µg/kg)/BTBR | 57.61 ± 4.97 | 542.39 ± 7.94 | 3905 ± 152.65 |
| MLA (1µg/kg)/BTBR | 59.00 ± 4.65 | 541.00 ± 8.19 | 4149 ± 147.64 |
| BTBR | ||||||||
|---|---|---|---|---|---|---|---|---|
| CUR | ||||||||
| C57 (SAL) | (SAL) | 25 mg/kg | 50 mg/kg | 100 mg/kg | 50 mg/kg + MLA (1 µg/kg) | MLA (1 µg/kg) | DOZ (1 mg/kg) | |
| SOD | ||||||||
| Cerebellum | 50.95 ± 1.89 | 36.42 ± 2.19 *** | 38.24 ± 2.05 | 36.16 ± 1.73 | 39.41 ± 3.04 | 36.91 ± 1.38 | 37.67 ± 1.26 | 36.52 ± 1.73 |
| Hippo-campus | 49.47 ± 1.52 | 37.70 ± 1.61 *** | 39.55 ± 1.67 | 49.36 ± 0.5 ## | 50.37 ± 1.55 ## | 48.61 ± 3.24 | 38.63 ± 1.45 | 36.95 ± 2.67 |
| CAT | ||||||||
| Cerebellum | 159.43 ± 6.92 | 127.95 ± 3.37 * | 128.30 ± 4.07 | 151.11 ± 5.47 # | 149.82 ± 4.72 # | 148.07 ± 5.10 # | 127.95 ± 3.37 | 133.95 ± 5.21 |
| Hippo-campus | 26.53 ± 4.90 | 106.58 ± 3.01 * | 104.13 ± 7.54 | 105.50 ± 3.77 | 108.11 ± 5.61 | 104.08 ± 3.97 | 103.38 ± 6.23 | 105.58 ± 3.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayaprakash, P.; Isaev, D.; Shabbir, W.; Lorke, D.E.; Sadek, B.; Oz, M. Curcumin Potentiates α7 Nicotinic Acetylcholine Receptors and Alleviates Autistic-Like Social Deficits and Brain Oxidative Stress Status in Mice. Int. J. Mol. Sci. 2021, 22, 7251. https://doi.org/10.3390/ijms22147251
Jayaprakash P, Isaev D, Shabbir W, Lorke DE, Sadek B, Oz M. Curcumin Potentiates α7 Nicotinic Acetylcholine Receptors and Alleviates Autistic-Like Social Deficits and Brain Oxidative Stress Status in Mice. International Journal of Molecular Sciences. 2021; 22(14):7251. https://doi.org/10.3390/ijms22147251
Chicago/Turabian StyleJayaprakash, Petrilla, Dmytro Isaev, Waheed Shabbir, Dietrich E. Lorke, Bassem Sadek, and Murat Oz. 2021. "Curcumin Potentiates α7 Nicotinic Acetylcholine Receptors and Alleviates Autistic-Like Social Deficits and Brain Oxidative Stress Status in Mice" International Journal of Molecular Sciences 22, no. 14: 7251. https://doi.org/10.3390/ijms22147251
APA StyleJayaprakash, P., Isaev, D., Shabbir, W., Lorke, D. E., Sadek, B., & Oz, M. (2021). Curcumin Potentiates α7 Nicotinic Acetylcholine Receptors and Alleviates Autistic-Like Social Deficits and Brain Oxidative Stress Status in Mice. International Journal of Molecular Sciences, 22(14), 7251. https://doi.org/10.3390/ijms22147251