
Increased Endocytosis of Cadmium-Metallothionein through the 24p3 Receptor in an In Vivo Model with Reduced Proximal Tubular Activity

 , ,
, ,  and
and 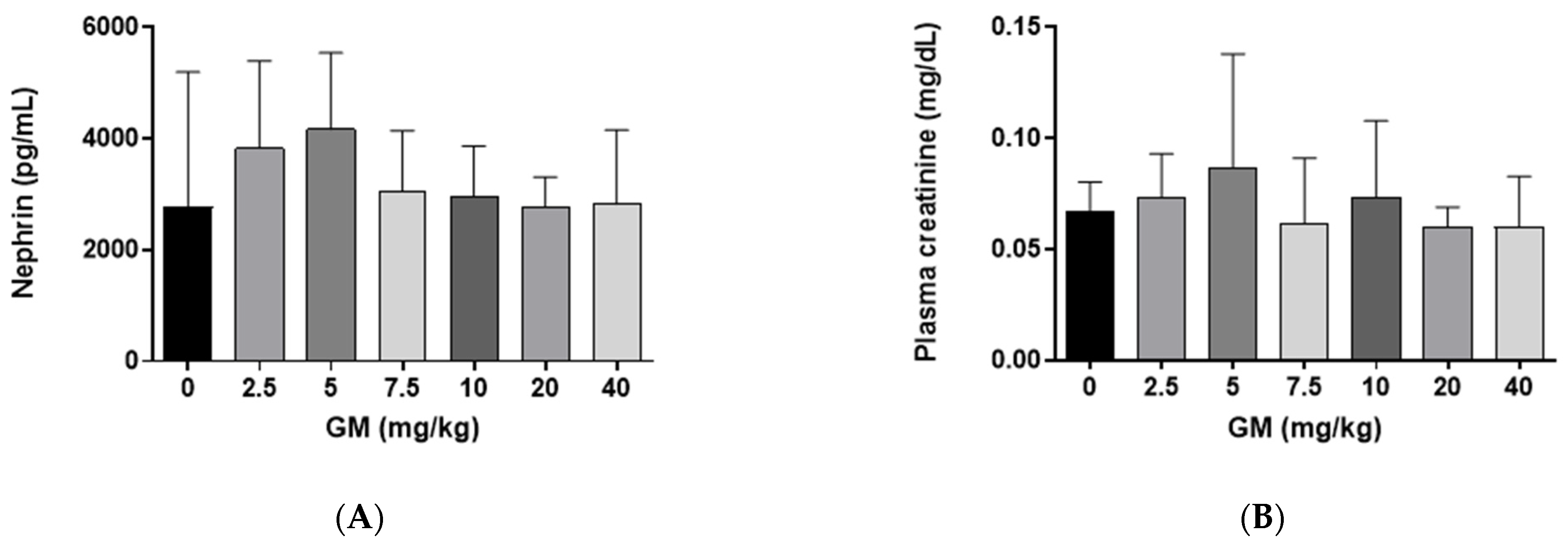{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of GM Administration on Glomerular Function
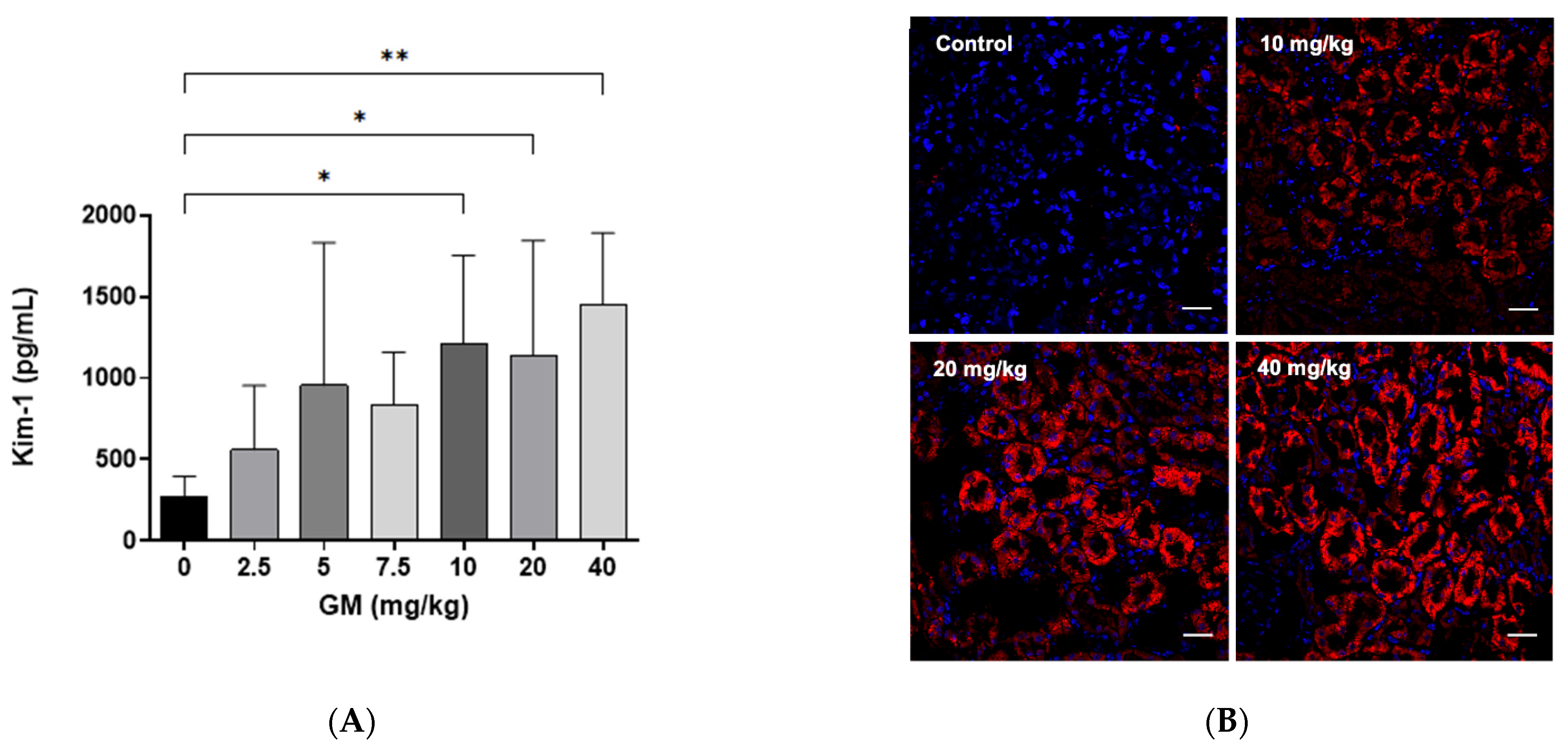
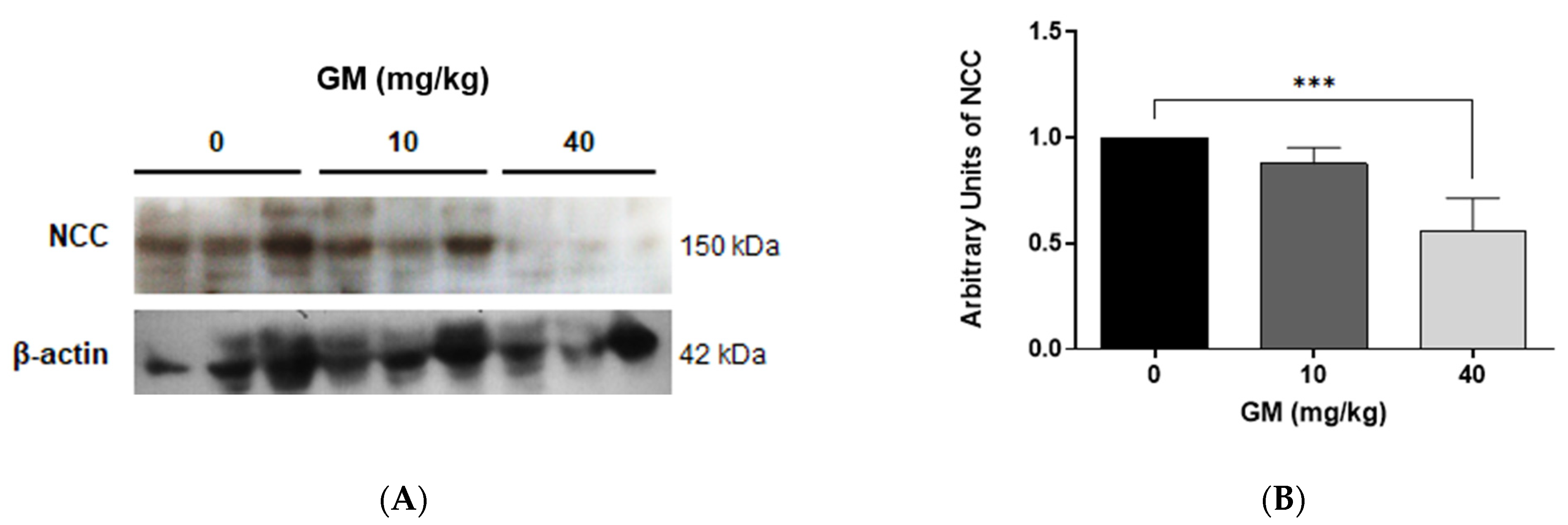
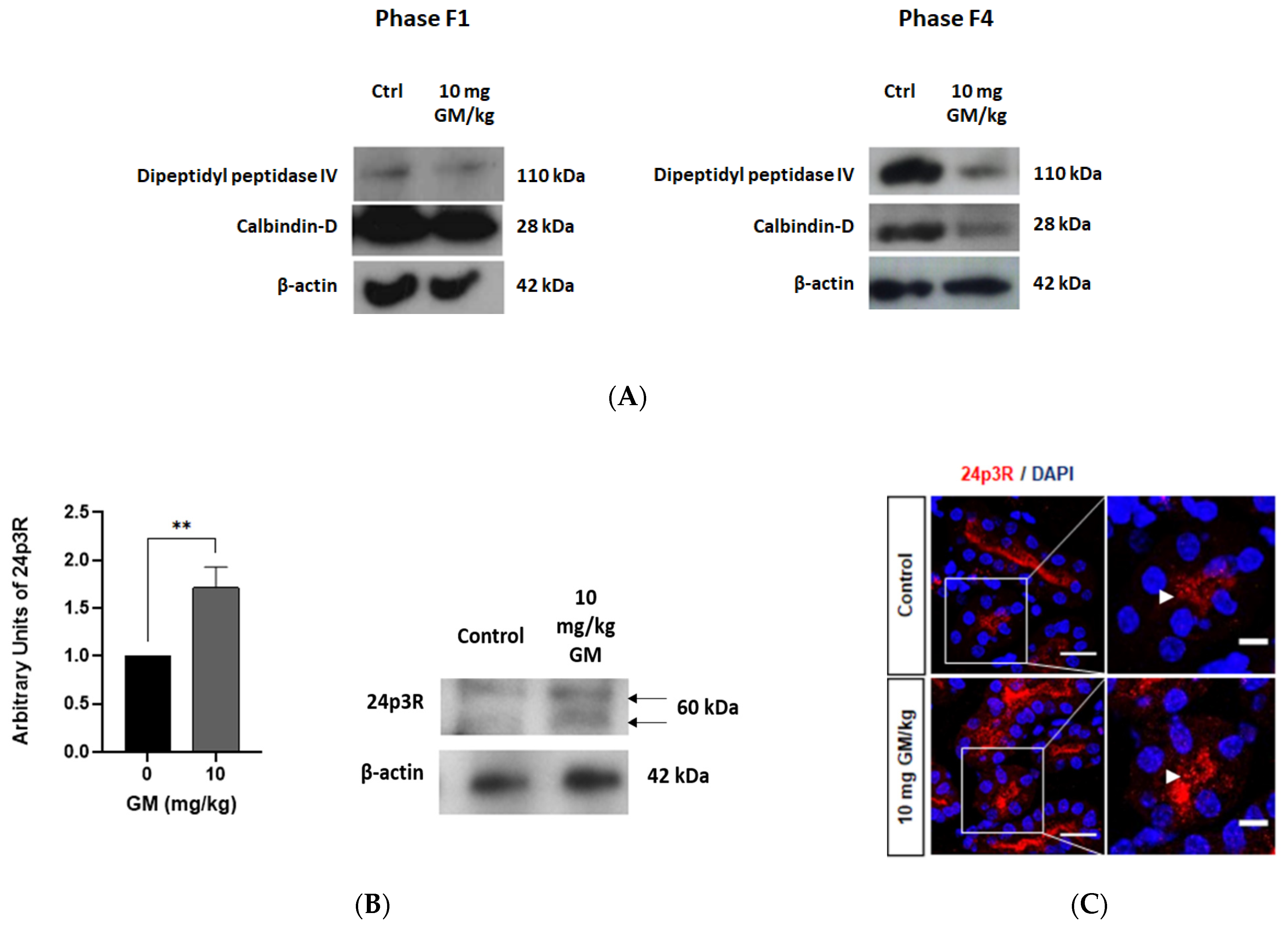
2.2. Effect of GM Administration on Tubular Injury
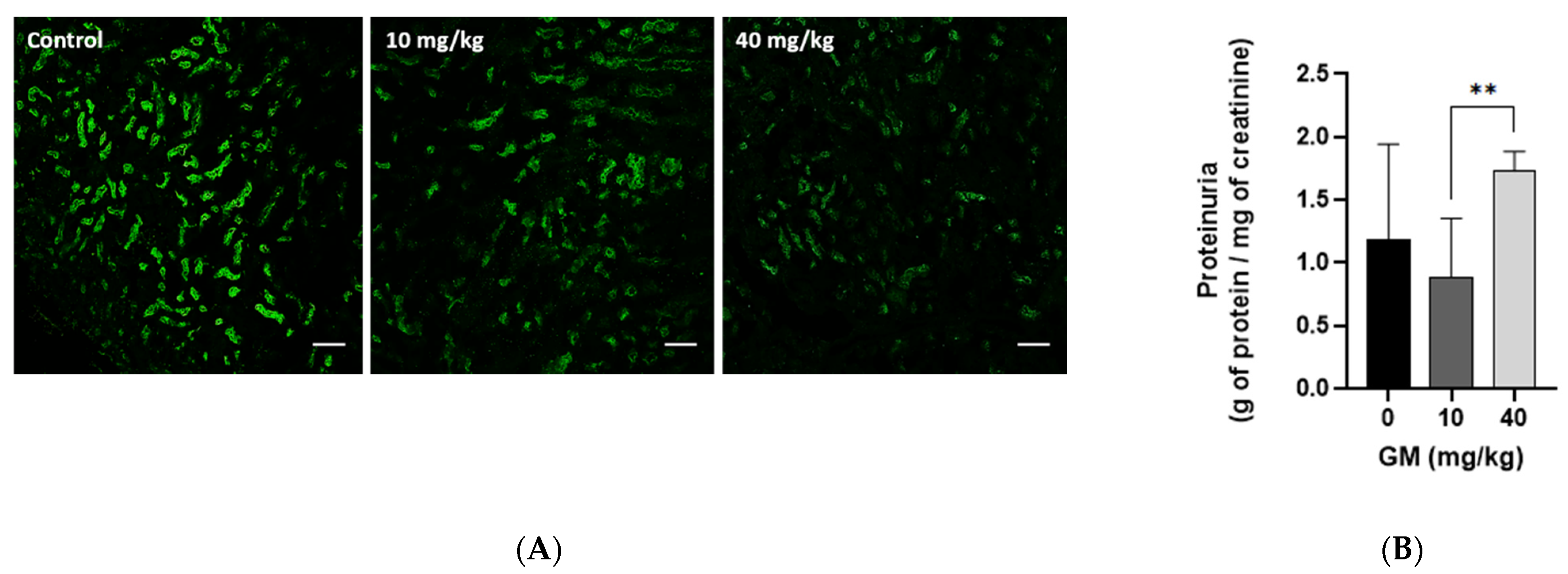
2.3. Effect of GM Administration on Protein Excretion
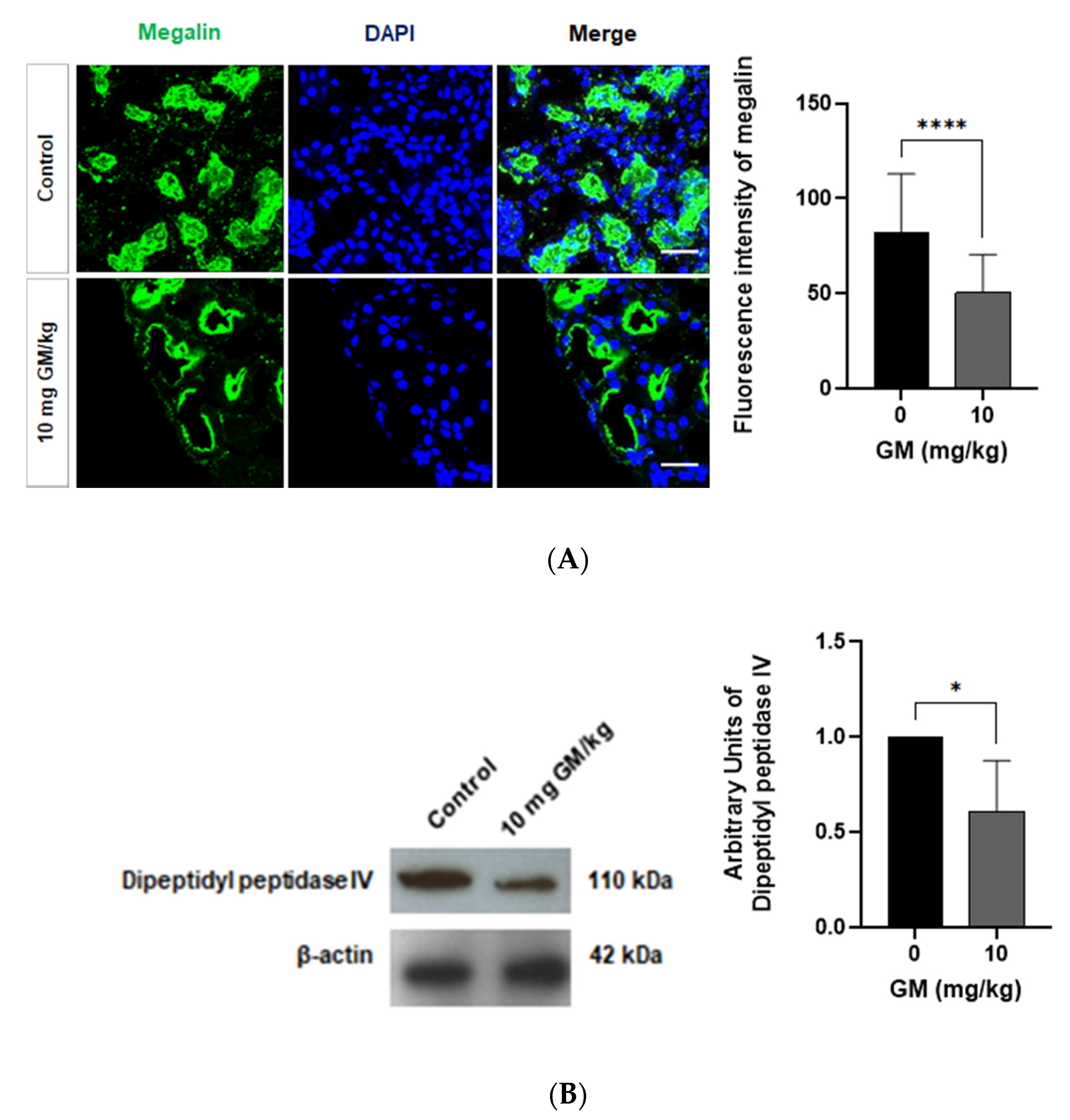
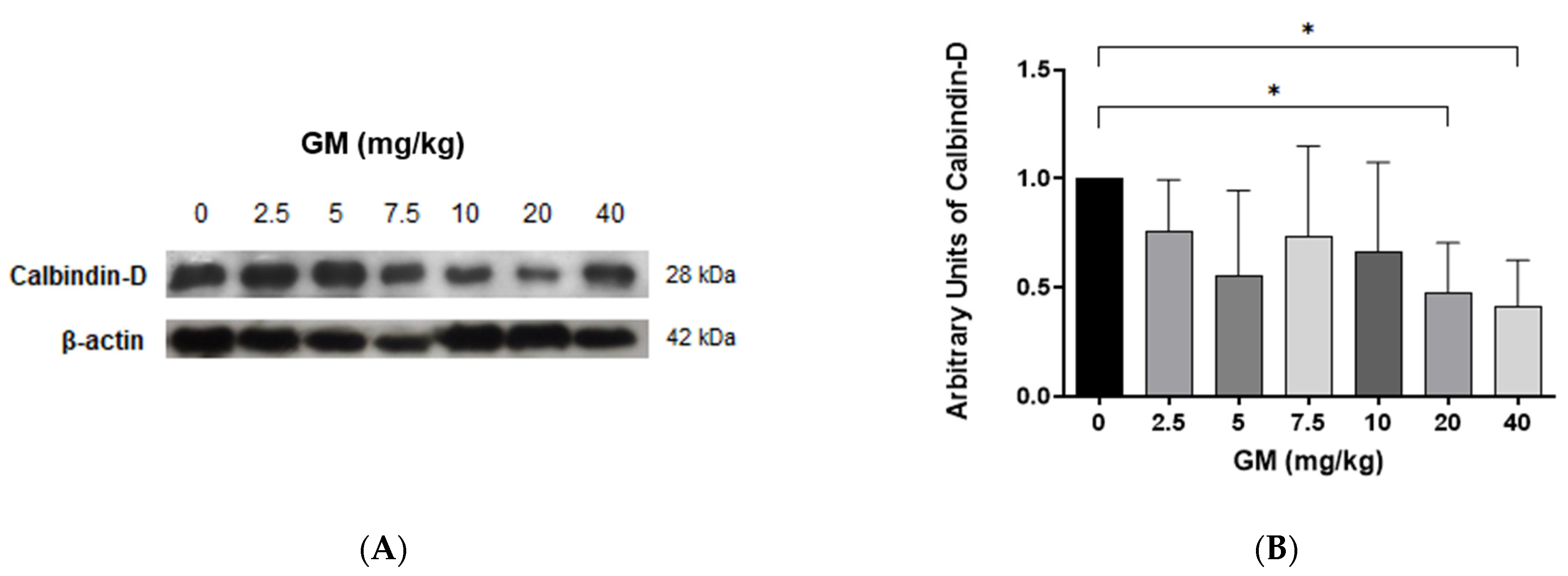
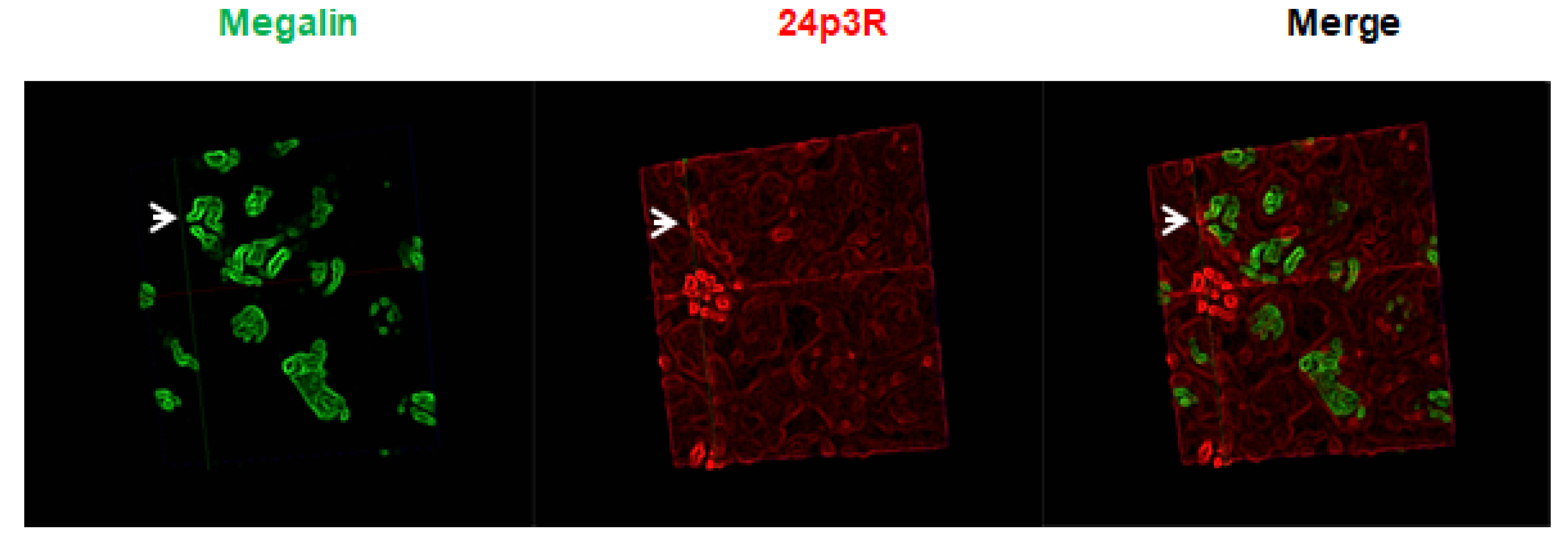
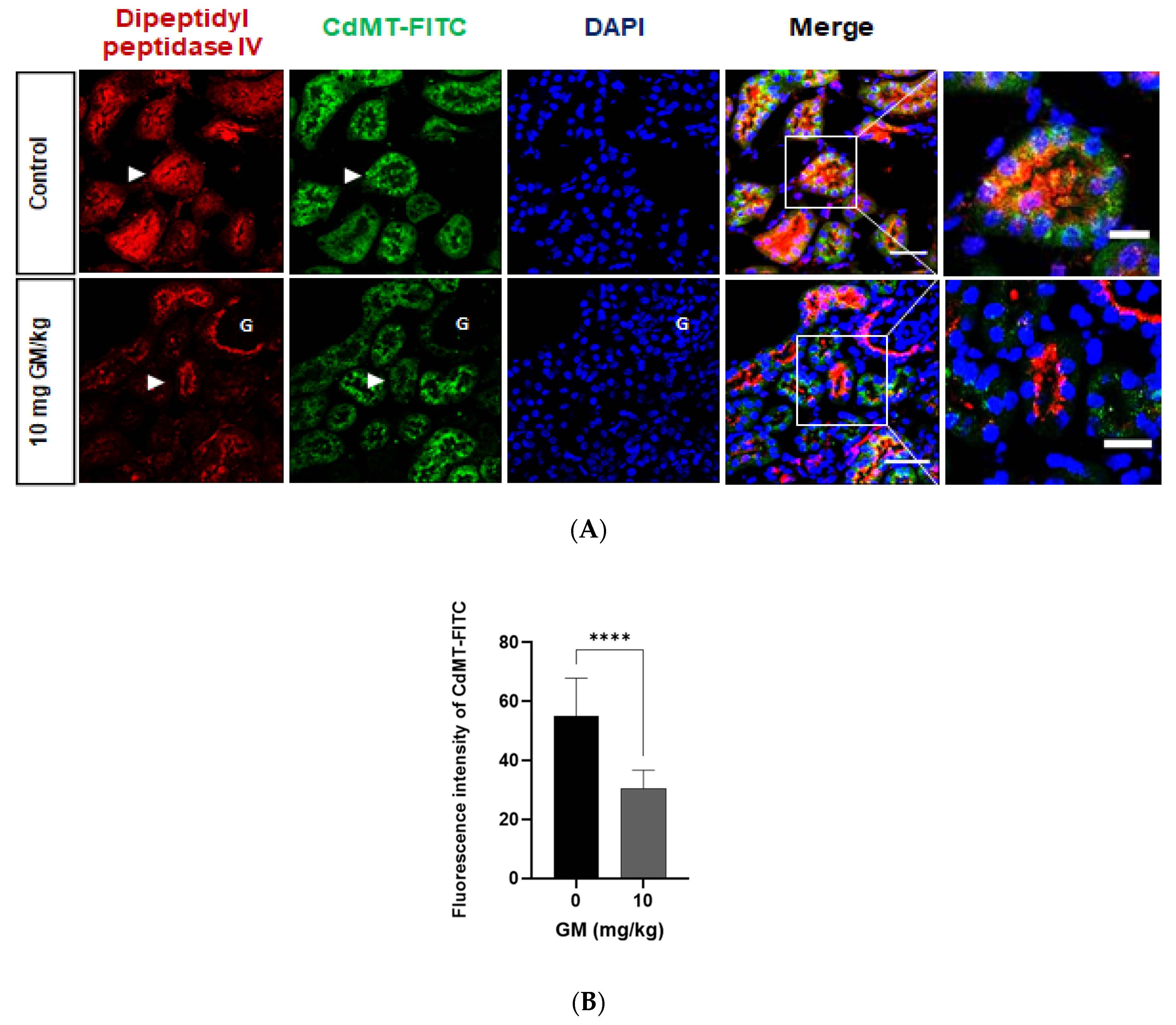
2.4. Expression of Megalin Receptor and 24p3R in the Renal Cortex of Mice Exposed to GM
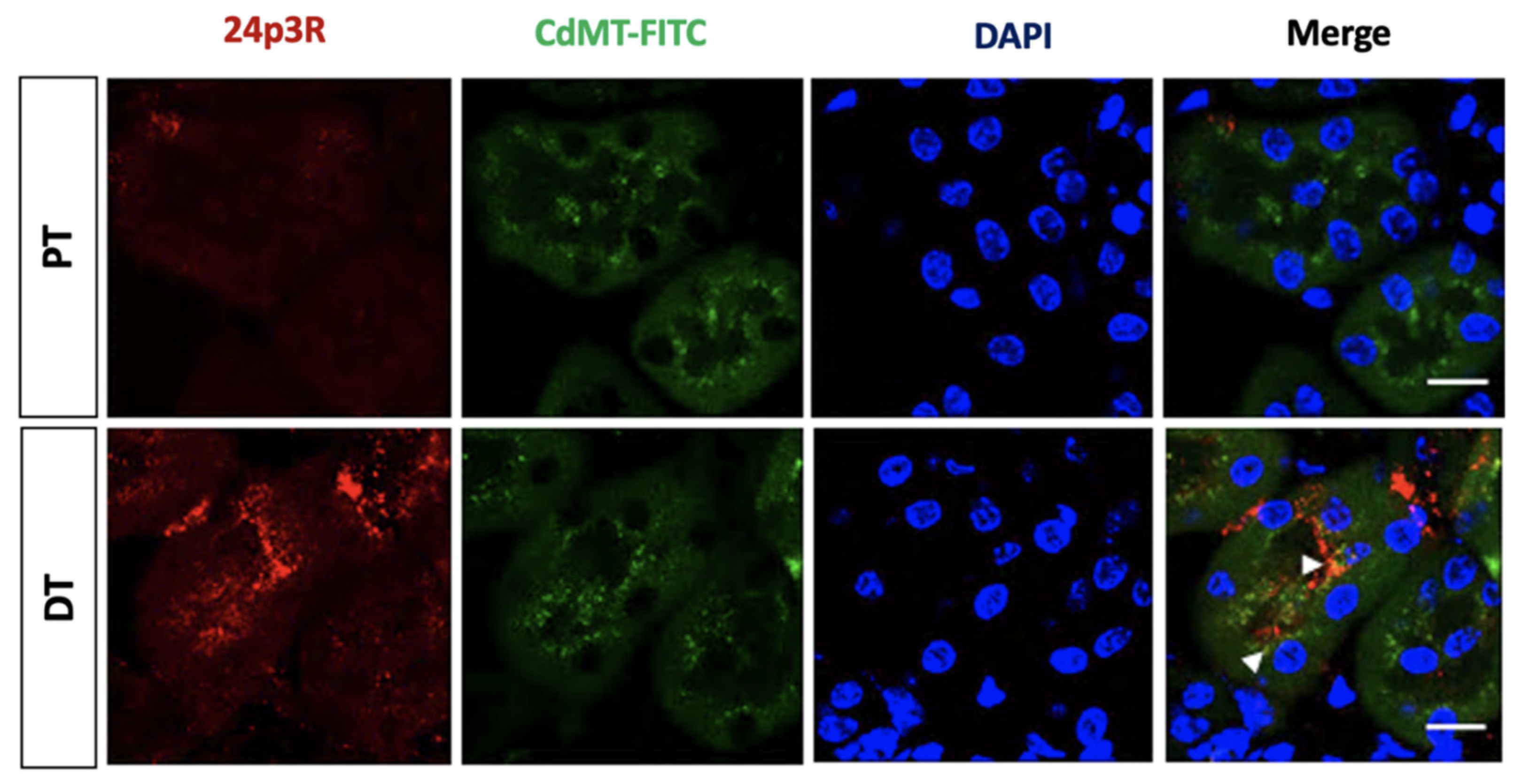
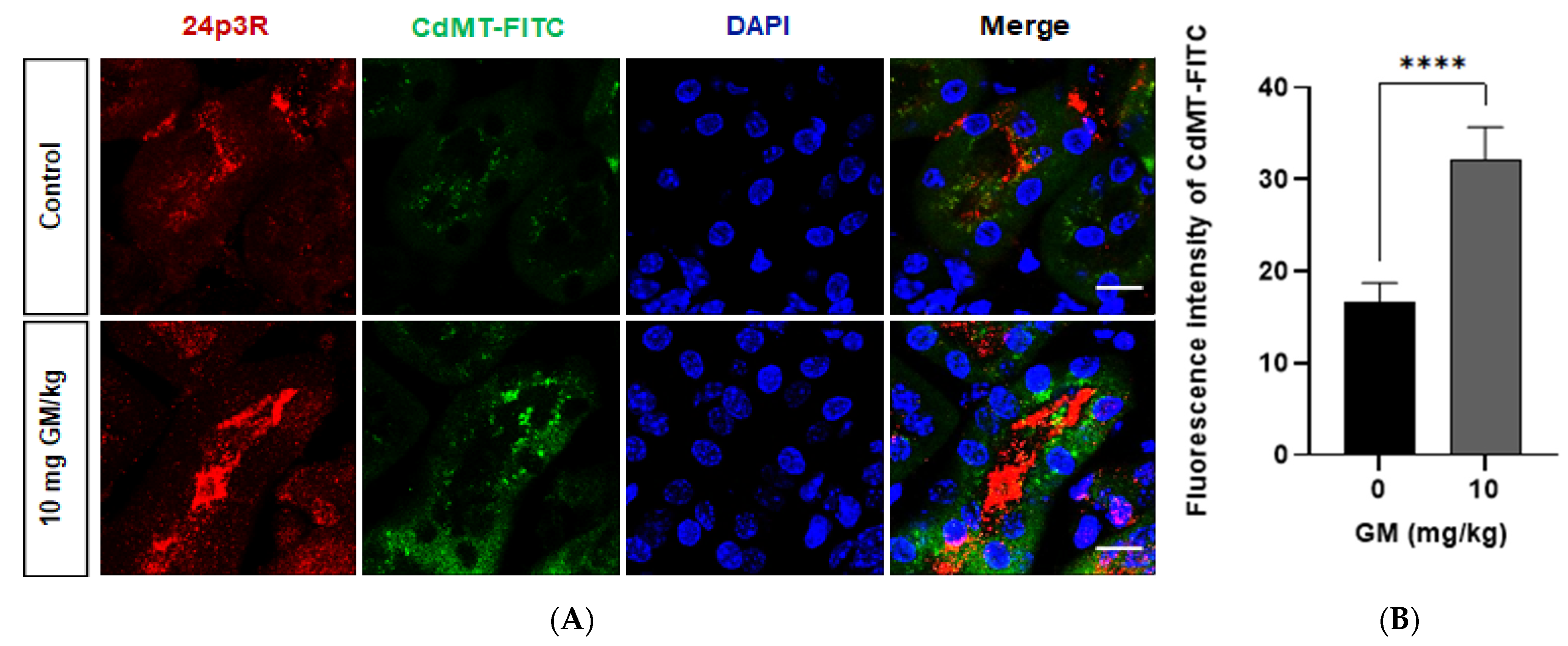
2.5. Endocytosis of Filtered CdMT Complex by PT or DT (and Possibly CD) Expressing 24p3R in Untreated or GM-Treated Mice
3. Discussion
4. Materials and Methods
4.1. Antibodies
4.2. Animals
4.3. Urine and Plasma Measurements
4.4. Magnetic Luminex Assay
4.5. Western-Blot Assay
4.6. Conjugation of CdMT to Fluorescein Isothiocyanate (FITC)
4.7. Injection of the CdMT FITC Complex
4.8. Immunofluorescence
4.9. Staining Quantification
4.10. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thévenod, F.; Lee, W.-K. Toxicology of Cadmium and Its Damage to Mammalian Organs. In Cadmium: From Toxicity to Essentiality; Sigel, A., Sigel, H., Sigel, R.K., Eds.; Metal Ions in Life Sciences; Springer: Dordrecht, The Netherlands, 2013; Volume 11, pp. 415–490. ISBN 978-94-007-5178-1. [Google Scholar]
- Nordberg, M.; Nordberg, G.F. Distribution of Metallothionein-Bound Cadmium and Cadmium Chloride in Mice: Preliminary Studies. Environ. Health Perspect. 1975, 12, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Thévenod, F.; Wolff, N.A. Iron Transport in the Kidney: Implications for Physiology and Cadmium Nephrotoxicity. Metallomics 2016, 8, 17–42. [Google Scholar] [CrossRef]
- Pham, T.N.D.; Ségui, J.A.; Fortin, C.; Campbell, P.G.C.; Denizeau, F.; Jumarie, C. Cadmium Uptake in Rat Hepatocytes in Relation to Speciation and to Complexation with Metallothionein and Albumin: Cd Speciation and Accumulation in Rat Hepatocytes. J. Cell. Physiol. 2004, 201, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Sabolić, I.; Breljak, D.; Škarica, M.; Herak-Kramberger, C.M. Role of Metallothionein in Cadmium Traffic and Toxicity in Kidneys and Other Mammalian Organs. BioMetals 2010, 23, 897–926. [Google Scholar] [CrossRef] [PubMed]
- Freisinger, E.; Vašák, M. Cadmium in Metallothioneins. In Cadmium: From Toxicity to Essentiality; Sigel, A., Sigel, H., Sigel, R.K., Eds.; Metal Ions in Life Sciences; Springer: Dordrecht, The Netherlands, 2013; Volume 11, pp. 339–371. ISBN 978-94-007-5178-1. [Google Scholar]
- Jin, T.; Lu, J.; Nordberg, M. Toxicokinetics and Biochemistry of Cadmium with Special Emphasis on the Role of Metallothionein. Neurotoxicology 1998, 19, 529–535. [Google Scholar] [PubMed]
- Klaassen, C.D.; Liu, J.; Diwan, B.A. Metallothionein Protection of Cadmium Toxicity. Toxicol. Appl. Pharmacol. 2009, 238, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, E.I.; Kristoffersen, I.B.; Grann, B.; Thomsen, J.S.; Andreasen, A.; Nielsen, R. A Well-Developed Endolysosomal System Reflects Protein Reabsorption in Segment 1 and 2 of Rat Proximal Tubules. Kidney Int. 2021, 99, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.I.; Birn, H. Megalin and Cubilin: Synergistic Endocytic Receptors in Renal Proximal Tubule. Am. J. Physiol. Ren. Physiol. 2001, 280, F562–F573. [Google Scholar] [CrossRef]
- Christensen, E.I.; Verroust, P.J.; Nielsen, R. Receptor-Mediated Endocytosis in Renal Proximal Tubule. Pflüg. Arch. Eur. J. Physiol. 2009, 458, 1039–1048. [Google Scholar] [CrossRef]
- Erfurt, C.; Roussa, E.; Thévenod, F. Apoptosis by Cd2+ or CdMT in Proximal Tubule Cells: Different Uptake Routes and Permissive Role of Endo/Lysosomal CdMT Uptake. Am. J. Physiol. Cell Physiol. 2003, 285, C1367–C1376. [Google Scholar] [CrossRef] [Green Version]
- Wolff, N.A.; Abouhamed, M.; Verroust, P.J.; Thévenod, F. Megalin-Dependent Internalization of Cadmium-Metallothionein and Cytotoxicity in Cultured Renal Proximal Tubule Cells. J. Pharmacol. Exp. Ther. 2006, 318, 782–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abouhamed, M.; Gburek, J.; Liu, W.; Torchalski, B.; Wilhelm, A.; Wolff, N.A.; Christensen, E.I.; Thévenod, F.; Smith, C.P. Divalent Metal Transporter 1 in the Kidney Proximal Tubule Is Expressed in Late Endosomes/Lysosomal Membranes: Implications for Renal Handling of Protein-Metal Complexes. Am. J. Physiol. Ren. Physiol. 2006, 290, F1525–F1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abouhamed, M.; Wolff, N.A.; Lee, W.-K.; Smith, C.P.; Thévenod, F. Knockdown of Endosomal/Lysosomal Divalent Metal Transporter 1 by RNA Interference Prevents Cadmium-Metallothionein-1 Cytotoxicity in Renal Proximal Tubule Cells. Am. J. Physiol. Ren. Physiol. 2007, 293, F705–F712. [Google Scholar] [CrossRef] [Green Version]
- Zalups, R.K.; Ahmad, S. Molecular Handling of Cadmium in Transporting Epithelia. Toxicol. Appl. Pharmacol. 2003, 186, 163–188. [Google Scholar] [CrossRef]
- Mitchell, C.J.; Shawki, A.; Ganz, T.; Nemeth, E.; Mackenzie, B. Functional Properties of Human Ferroportin, a Cellular Iron Exporter Reactive Also with Cobalt and Zinc. Am. J. Physiol. Cell Physiol. 2014, 306, C450–C459. [Google Scholar] [CrossRef] [Green Version]
- Wolff, N.A.; Liu, W.; Fenton, R.A.; Lee, W.-K.; Thévenod, F.; Smith, C.P. Ferroportin 1 Is Expressed Basolaterally in Rat Kidney Proximal Tubule Cells and Iron Excess Increases Its Membrane Trafficking. J. Cell. Mol. Med. 2011, 15, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, P.M.; Costanzi, S.; Naticchia, A.; Sturniolo, A.; Gambaro, G. Low Level Exposure to Cadmium Increases the Risk of Chronic Kidney Disease: Analysis of the NHANES 1999–2006. BMC Public Health 2010, 10, 304. [Google Scholar] [CrossRef] [Green Version]
- Johri, N.; Jacquillet, G.; Unwin, R. Heavy Metal Poisoning: The Effects of Cadmium on the Kidney. BioMetals 2010, 23, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.I.; Birn, H. Megalin and Cubilin: Multifunctional Endocytic Receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Tojo, A.; Endou, H. Intrarenal Handling of Proteins in Rats Using Fractional Micropuncture Technique. Am. J. Physiol. Ren. Physiol. 1992, 263, F601–F606. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, R.; Christensen, E.I. Proteinuria and Events beyond the Slit. Pediatr. Nephrol. 2010, 25, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Norden, A.G.W.; Lapsley, M.; Lee, P.J.; Pusey, C.D.; Scheinman, S.J.; Tam, F.W.K.; Thakker, R.V.; Unwin, R.J.; Wrong, O. Glomerular Protein Sieving and Implications for Renal Failure in Fanconi Syndrome. Kidney Int. 2001, 60, 1885–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norden, A.G.W.; Lapsley, M.; Igarashi, T.; Kelleher, C.L.; Lee, P.J.; Matsuyama, T.; Scheinman, S.J.; Shiraga, H.; Sundin, D.P.; Thakker, R.V.; et al. Urinary Megalin Deficiency Implicates Abnormal Tubular Endocytic Function in Fanconi Syndrome. J. Am. Soc. Nephrol. 2002, 13, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Klassen, R.B.; Crenshaw, K.; Kozyraki, R.; Verroust, P.J.; Tio, L.; Atrian, S.; Allen, P.L.; Hammond, T.G. Megalin Mediates Renal Uptake of Heavy Metal Metallothionein Complexes. Am. J. Physiol. Ren. Physiol. 2004, 287, F393–F403. [Google Scholar] [CrossRef]
- Milnerowicz, H.; Bizoń, A. Determination of Metallothionein in Biological Fluids Using Enzyme-Linked Immunoassay with Commercial Antibody. Acta Biochim. Pol. 2010, 57, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Fels, J.; Scharner, B.; Zarbock, R.; Zavala Guevara, I.P.; Lee, W.-K.; Barbier, O.C.; Thévenod, F. Cadmium Complexed with Β2-Microglubulin, Albumin and Lipocalin-2 Rather than Metallothionein Cause Megalin:Cubilin Dependent Toxicity of the Renal Proximal Tubule. Int. J. Mol. Sci. 2019, 20, 2379. [Google Scholar] [CrossRef] [Green Version]
- Mori, K.; Lee, H.T.; Rapoport, D.; Drexler, I.R.; Foster, K.; Yang, J.; Schmidt-Ott, K.M.; Chen, X.; Li, J.Y.; Weiss, S.; et al. Endocytic Delivery of Lipocalin-Siderophore-Iron Complex Rescues the Kidney from Ischemia-Reperfusion Injury. J. Clin. Investig. 2005, 115, 610–621. [Google Scholar] [CrossRef]
- Ashraf, M.I.; Schwelberger, H.G.; Brendel, K.A.; Feurle, J.; Andrassy, J.; Kotsch, K.; Regele, H.; Pratschke, J.; Maier, H.T.; Aigner, F. Exogenous Lipocalin 2 Ameliorates Acute Rejection in a Mouse Model of Renal Transplantation: Role of Lipocalin 2 in Renal Transplantation. Am. J. Transplant. 2016, 16, 808–820. [Google Scholar] [CrossRef] [Green Version]
- Bao, G.-H.; Ho, C.-T.; Barasch, J. The Ligands of Neutrophil Gelatinase-Associated Lipocalin. RSC Adv. 2015, 5, 104363–104374. [Google Scholar] [CrossRef] [Green Version]
- Paragas, N.; Qiu, A.; Zhang, Q.; Samstein, B.; Deng, S.-X.; Schmidt-Ott, K.M.; Viltard, M.; Yu, W.; Forster, C.S.; Gong, G.; et al. The Ngal Reporter Mouse Detects the Response of the Kidney to Injury in Real Time. Nat. Med. 2011, 17, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Devireddy, L.R.; Gazin, C.; Zhu, X.; Green, M.R. A Cell-Surface Receptor for Lipocalin 24p3 Selectively Mediates Apoptosis and Iron Uptake. Cell 2005, 123, 1293–1305. [Google Scholar] [CrossRef] [Green Version]
- Langelueddecke, C.; Roussa, E.; Fenton, R.A.; Wolff, N.A.; Lee, W.-K.; Thévenod, F. Lipocalin-2 (24p3/Neutrophil Gelatinase-Associated Lipocalin (NGAL)) Receptor Is Expressed in Distal Nephron and Mediates Protein Endocytosis. J. Biol. Chem. 2012, 287, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Dizin, E.; Hasler, U.; Nlandu-Khodo, S.; Fila, M.; Roth, I.; Ernandez, T.; Doucet, A.; Martin, P.-Y.; Feraille, E.; de Seigneux, S. Albuminuria Induces a Proinflammatory and Profibrotic Response in Cortical Collecting Ducts via the 24p3 Receptor. Am. J. Physiol. Ren. Physiol. 2013, 305, F1053–F1063. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, Y.; Ushijima, K.; Tsuchiya, H.; Morishige, J.; Mii, A.; Ando, H.; Tsuruoka, S.; Fujimura, A. Influence of Renal Ischaemia-reperfusion Injury on Renal Neutrophil Gelatinase-associated Lipocalin Receptor (24p3R) in Rats. Clin. Exp. Pharmacol. Physiol. 2019, 46, 1166–1173. [Google Scholar] [CrossRef]
- Girolami, J.P.; Bascands, J.L.; Pécher, C.; Cabos, G.; Moatti, J.P.; Mercier, J.F.; Haguenoer, J.M.; Manuel, Y. Renal Kallikrein Excretion as a Distal Nephrotoxicity Marker during Cadmium Exposure in Rats. Toxicology 1989, 55, 117–129. [Google Scholar] [CrossRef]
- Lauwerys, R.; Bernard, A. Preclinical Detection of Nephrotoxicity: Description of the Tests and Appraisal of Their Health Significance. Toxicol. Lett. 1989, 46, 13–29. [Google Scholar] [CrossRef]
- van Swelm, R.P.L.; Vos, M.; Verhoeven, F.; Thévenod, F.; Swinkels, D.W. Endogenous Hepcidin Synthesis Protects the Distal Nephron against Hemin and Hemoglobin Mediated Necroptosis. Cell Death Dis. 2018, 9, 550. [Google Scholar] [CrossRef]
- Nagai, J.; Takano, M. Entry of Aminoglycosides into Renal Tubular Epithelial Cells via Endocytosis-Dependent and Endocytosis-Independent Pathways. Biochem. Pharmacol. 2014, 90, 331–337. [Google Scholar] [CrossRef]
- Hori, Y.; Aoki, N.; Kuwahara, S.; Hosojima, M.; Kaseda, R.; Goto, S.; Iida, T.; De, S.; Kabasawa, H.; Kaneko, R.; et al. Megalin Blockade with Cilastatin Suppresses Drug-Induced Nephrotoxicity. J. Am. Soc. Nephrol. 2017, 28, 1783–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiting, P.H.; Brown, P.A.J. The Relationship Between Enzymuria and Kidney Enzyme Activities in Experimental Gentamicin Nephrotoxicity. Ren. Fail. 1996, 18, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Akankwasa, G.; Jianhua, L.; Guixue, C.; Changjuan, A.; Xiaosong, Q. Urine Markers of Podocyte Dysfunction: A Review of Podocalyxin and Nephrin in Selected Glomerular Diseases. Biomark. Med. 2018, 12, 927–935. [Google Scholar] [CrossRef]
- Mentzel, S.; Dijkman, H.B.; Van Son, J.P.; Koene, R.A.; Assmann, K.J. Organ Distribution of Aminopeptidase A and Dipeptidyl Peptidase IV in Normal Mice. J. Histochem. Cytochem. 1996, 44, 445–461. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Chou, C.-L.; Knepper, M.A. Deep Sequencing in Microdissected Renal Tubules Identifies Nephron Segment–Specific Transcriptomes. J. Am. Soc. Nephrol. 2015, 26, 2669–2677. [Google Scholar] [CrossRef]
- Roth, J.; Brown, D.; Norman, A.W.; Orci, L. Localization of the Vitamin D-Dependent Calcium-Binding Protein in Mammalian Kidney. Am. J. Physiol. Ren. Physiol. 1982, 243, F243–F252. [Google Scholar] [CrossRef]
- Lee, S.M.; Meyer, M.B.; Benkusky, N.A.; O’Brien, C.A.; Pike, J.W. Mechanisms of Enhancer-Mediated Hormonal Control of Vitamin D Receptor Gene Expression in Target Cells. J. Biol. Chem. 2015, 290, 30573–30586. [Google Scholar] [CrossRef] [Green Version]
- Hebert, S.C.; Mount, D.B.; Gamba, G. Molecular Physiology of Cation-Coupled Cl− Cotransport: The SLC12 Family. Pflug. Arch. Eur. J. Physiol. 2004, 447, 580–593. [Google Scholar] [CrossRef] [PubMed]
- Langelueddecke, C.; Roussa, E.; Fenton, R.A.; Thévenod, F. Expression and Function of the Lipocalin-2 (24p3/NGAL) Receptor in Rodent and Human Intestinal Epithelia. PLoS ONE 2013, 8, e71586. [Google Scholar] [CrossRef] [Green Version]
- Nagai, J.; Katsube, T.; Murakami, T.; Takano, M. Effect of Gentamicin on Pharmacokinetics of Lysozyme in Rats: Interaction between Megalin Substrates in the Kidney. J. Pharm. Pharmacol. 2002, 54, 1491–1496. [Google Scholar] [CrossRef] [PubMed]
- Laurent, G.; Kishore, B.K.; Tulkens, P.M. Aminoglycoside-Induced Renal Phospholipidosis and Nephrotoxicity. Biochem. Pharmacol. 1990, 40, 2383–2392. [Google Scholar] [CrossRef]
- Servais, H.; Ortiz, A.; Devuyst, O.; Denamur, S.; Tulkens, P.M.; Mingeot-Leclercq, M.-P. Renal Cell Apoptosis Induced by Nephrotoxic Drugs: Cellular and Molecular Mechanisms and Potential Approaches to Modulation. Apoptosis 2008, 13, 11–32. [Google Scholar] [CrossRef]
- Zhou, Y.; Vaidya, V.S.; Brown, R.P.; Zhang, J.; Rosenzweig, B.A.; Thompson, K.L.; Miller, T.J.; Bonventre, J.V.; Goering, P.L. Comparison of Kidney Injury Molecule-1 and Other Nephrotoxicity Biomarkers in Urine and Kidney Following Acute Exposure to Gentamicin, Mercury, and Chromium. Toxicol. Sci. 2008, 101, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Ichimura, T.; Bonventre, J.V.; Bailly, V.; Wei, H.; Hession, C.A.; Cate, R.L.; Sanicola, M. Kidney Injury Molecule-1 (KIM-1), a Putative Epithelial Cell Adhesion Molecule Containing a Novel Immunoglobulin Domain, Is Up-Regulated in Renal Cells after Injury. J. Biol. Chem. 1998, 273, 4135–4142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichimura, T.; Hung, C.C.; Yang, S.A.; Stevens, J.L.; Bonventre, J.V. Kidney Injury Molecule-1: A Tissue and Urinary Biomarker for Nephrotoxicant-Induced Renal Injury. Am. J. Physiol. Ren. Physiol. 2004, 286, F552–F563. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, V.S.; Ramirez, V.; Ichimura, T.; Bobadilla, N.A.; Bonventre, J.V. Urinary Kidney Injury Molecule-1: A Sensitive Quantitative Biomarker for Early Detection of Kidney Tubular Injury. Am. J. Physiol. Ren. Physiol. 2006, 290, F517–F529. [Google Scholar] [CrossRef]
- Pérez-Rojas, J.; Blanco, J.A.; Cruz, C.; Trujillo, J.; Vaidya, V.S.; Uribe, N.; Bonventre, J.V.; Gamba, G.; Bobadilla, N.A. Mineralocorticoid Receptor Blockade Confers Renoprotection in Preexisting Chronic Cyclosporine Nephrotoxicity. Am. J. Physiol. Ren. Physiol. 2007, 292, F131–F139. [Google Scholar] [CrossRef] [PubMed]
- Prozialeck, W.C.; Vaidya, V.S.; Liu, J.; Waalkes, M.P.; Edwards, J.R.; Lamar, P.C.; Bernard, A.M.; Dumont, X.; Bonventre, J.V. Kidney Injury Molecule-1 Is an Early Biomarker of Cadmium Nephrotoxicity. Kidney Int. 2007, 72, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Brooks, C.R.; Xiao, S.; Sabbisetti, V.; Yeung, M.Y.; Hsiao, L.-L.; Ichimura, T.; Kuchroo, V.; Bonventre, J.V. KIM-1–Mediated Phagocytosis Reduces Acute Injury to the Kidney. J. Clin. Investig. 2015, 125, 1620–1636. [Google Scholar] [CrossRef]
- Humphreys, B.D.; Xu, F.; Sabbisetti, V.; Grgic, I.; Naini, S.M.; Wang, N.; Chen, G.; Xiao, S.; Patel, D.; Henderson, J.M.; et al. Chronic Epithelial Kidney Injury Molecule-1 Expression Causes Murine Kidney Fibrosis. J. Clin. Investig. 2013, 123, 4023–4035. [Google Scholar] [CrossRef] [Green Version]
- Myrdal, S.E.; Steyger, P.S. TRPV1 Regulators Mediate Gentamicin Penetration of Cultured Kidney Cells. Hear. Res. 2005, 204, 170–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karasawa, T.; Wang, Q.; Fu, Y.; Cohen, D.M.; Steyger, P.S. TRPV4 Enhances the Cellular Uptake of Aminoglycoside Antibiotics. J. Cell Sci. 2008, 121, 2871–2879. [Google Scholar] [CrossRef] [Green Version]
- Hemmingsen, C. Regulation of Renal Calbindin-D28K. Pharmacol. Toxicol. 2000, 87 (Suppl. 3), 5–30. [Google Scholar]
- Aicher, L.; Meier, G.; Norcross, A.J.; Jakubowski, J.; Del Carmen Varela, M.; Cordier, A.; Steiner, S. Decrease in Kidney Calbindin-d 28kda as a Possible Mechanism Mediating Cyclosporine A- and FK-506-Induced Calciuria and Tubular Mineralization. Biochem. Pharmacol. 1997, 53, 723–731. [Google Scholar] [CrossRef]
- Sassen, M.C.; Kim, S.W.; Kwon, T.-H.; Knepper, M.A.; Miller, R.T.; Frøkiær, J.; Nielsen, S. Dysregulation of Renal Sodium Transporters in Gentamicin-Treated Rats. Kidney Int. 2006, 70, 1026–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabedo, M.A.-I.; Weinhäupl, K.; Lee, W.-K.; Wolff, N.A.; Storch, B.; Żerko, S.; Konrat, R.; Koźmiński, W.; Breuker, K.; Thévenod, F.; et al. Biochemical and Structural Characterization of the Interaction between the Siderocalin NGAL/LCN2 (Neutrophil Gelatinase-Associated Lipocalin/Lipocalin 2) and the N-Terminal Domain of Its Endocytic Receptor SLC22A. J. Biol. Chem. 2016, 291, 2917–2930. [Google Scholar] [CrossRef] [Green Version]
- Barasch, J.; Hollmen, M.; Deng, R.; Hod, E.A.; Rupert, P.B.; Abergel, R.J.; Allred, B.E.; Xu, K.; Darrah, S.F.; Tekabe, Y.; et al. Disposal of Iron by a Mutant Form of Lipocalin. Nat. Commun. 2016, 7, 12973. [Google Scholar] [CrossRef]
- Axelsson, L.; Bergenfeldt, M.; Ohlsson, K. Studies of the Release and Turnover of a Human Neutrophil Lipocalin. Scand. J. Clin. Lab. Investig. 1995, 55, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Betten, R.; Scharner, B.; Probst, S.; Edemir, B.; Wolff, N.A.; Langelueddecke, C.; Lee, W.-K.; Thévenod, F. Tonicity Inversely Modulates Lipocalin-2 (Lcn2/24p3/NGAL) Receptor (SLC22A17) and Lcn2 Expression via Wnt/β-Catenin Signaling in Renal Inner Medullary Collecting Duct Cells: Implications for Cell Fate and Bacterial Infection. Cell Commun. Signal. 2018, 16, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Probst, S.; Scharner, B.; McErlean, R.; Lee, W.-K.; Thévenod, F. Inverse Regulation of Lipocalin-2/24p3 Receptor/SLC22A17 and Lipocalin-2 Expression by Tonicity, NFAT5/TonEBP and Arginine Vasopressin in Mouse Cortical Collecting Duct Cells MCCD(Cl.1): Implications for Osmotolerance. Int. J. Mol. Sci. 2019, 20, 5398. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Z.; Wang, S.-Z.; Green, M.R. Transcription and Signalling Pathways Involved in BCR–ABL-Mediated Misregulation of 24p3 and 24p3R. EMBO J. 2009, 28, 866–876. [Google Scholar] [CrossRef] [Green Version]
- Weyer, K.; Storm, T.; Shan, J.; Vainio, S.; Kozyraki, R.; Verroust, P.J.; Christensen, E.I.; Nielsen, R. Mouse Model of Proximal Tubule Endocytic Dysfunction. Nephrol. Dial. Transplant. 2011, 26, 3446–3451. [Google Scholar] [CrossRef] [Green Version]
- Vinay, P.; Gougoux, A.; Lemieux, G. Isolation of a Pure Suspension of Rat Proximal Tubules. Am. J. Physiol. Ren. Physiol. 1981, 241, F403–F411. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, S.; Kurasaki, M.; Kojima, Y.; Kawanishi, S. Oxidative and Nonoxidative Mechanisms of Site-Specific DNA Cleavage Induced by Copper-Containing Metallothioneins. Biochemistry 1995, 34, 8763–8770. [Google Scholar] [CrossRef] [PubMed]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zavala-Guevara, I.P.; Ortega-Romero, M.S.; Narváez-Morales, J.; Jacobo-Estrada, T.L.; Lee, W.-K.; Arreola-Mendoza, L.; Thévenod, F.; Barbier, O.C. Increased Endocytosis of Cadmium-Metallothionein through the 24p3 Receptor in an In Vivo Model with Reduced Proximal Tubular Activity. Int. J. Mol. Sci. 2021, 22, 7262. https://doi.org/10.3390/ijms22147262
Zavala-Guevara IP, Ortega-Romero MS, Narváez-Morales J, Jacobo-Estrada TL, Lee W-K, Arreola-Mendoza L, Thévenod F, Barbier OC. Increased Endocytosis of Cadmium-Metallothionein through the 24p3 Receptor in an In Vivo Model with Reduced Proximal Tubular Activity. International Journal of Molecular Sciences. 2021; 22(14):7262. https://doi.org/10.3390/ijms22147262
Chicago/Turabian StyleZavala-Guevara, Itzel Pamela, Manolo Sibael Ortega-Romero, Juana Narváez-Morales, Tania Libertad Jacobo-Estrada, Wing-Kee Lee, Laura Arreola-Mendoza, Frank Thévenod, and Olivier Christophe Barbier. 2021. "Increased Endocytosis of Cadmium-Metallothionein through the 24p3 Receptor in an In Vivo Model with Reduced Proximal Tubular Activity" International Journal of Molecular Sciences 22, no. 14: 7262. https://doi.org/10.3390/ijms22147262
APA StyleZavala-Guevara, I. P., Ortega-Romero, M. S., Narváez-Morales, J., Jacobo-Estrada, T. L., Lee, W. -K., Arreola-Mendoza, L., Thévenod, F., & Barbier, O. C. (2021). Increased Endocytosis of Cadmium-Metallothionein through the 24p3 Receptor in an In Vivo Model with Reduced Proximal Tubular Activity. International Journal of Molecular Sciences, 22(14), 7262. https://doi.org/10.3390/ijms22147262