
CD73 Overexpression in Podocytes: A Novel Marker of Podocyte Injury in Human Kidney Disease

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
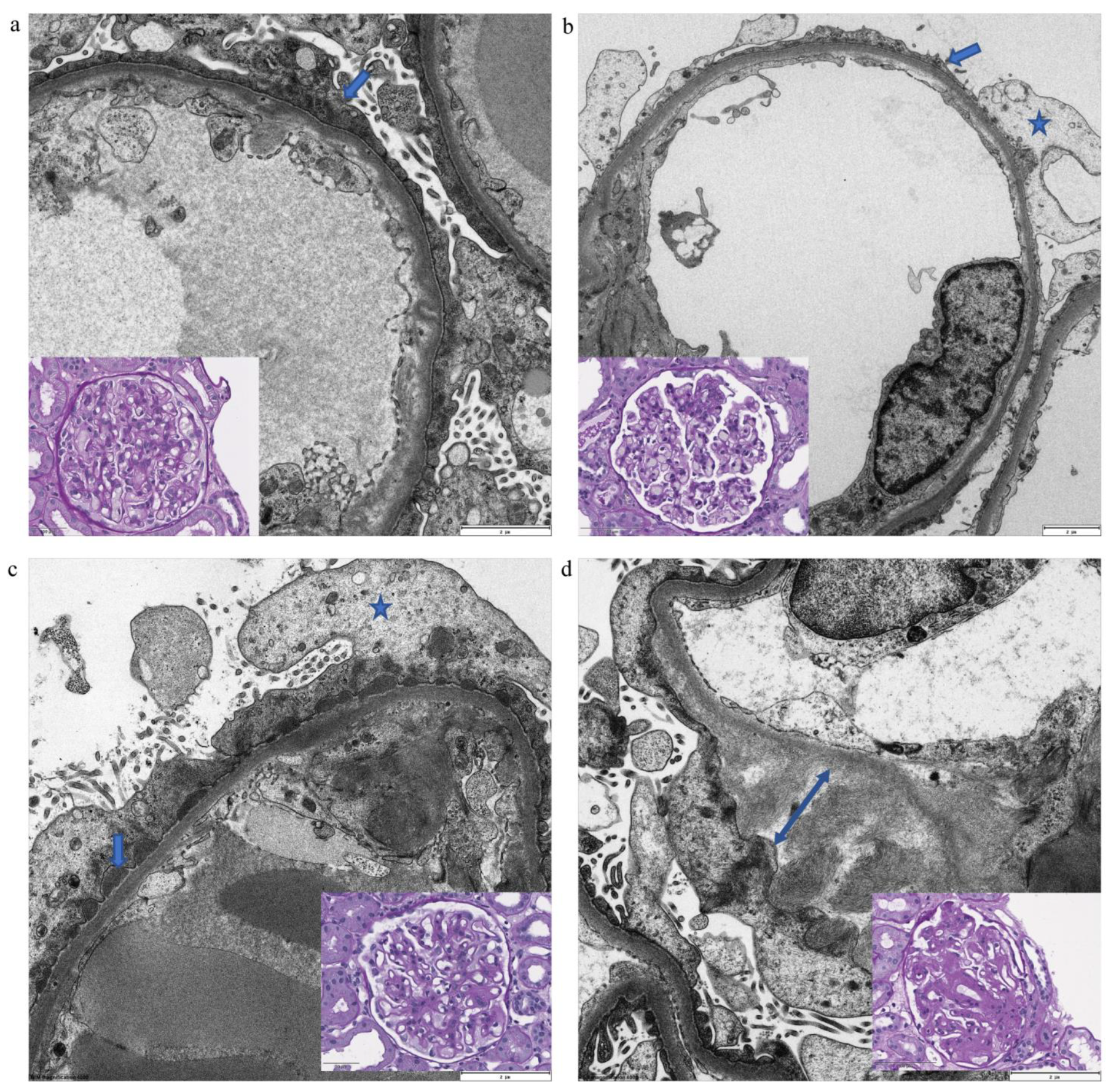
2.1. Clinical Data and Ultrastructural Imaging of Podocyte Damage
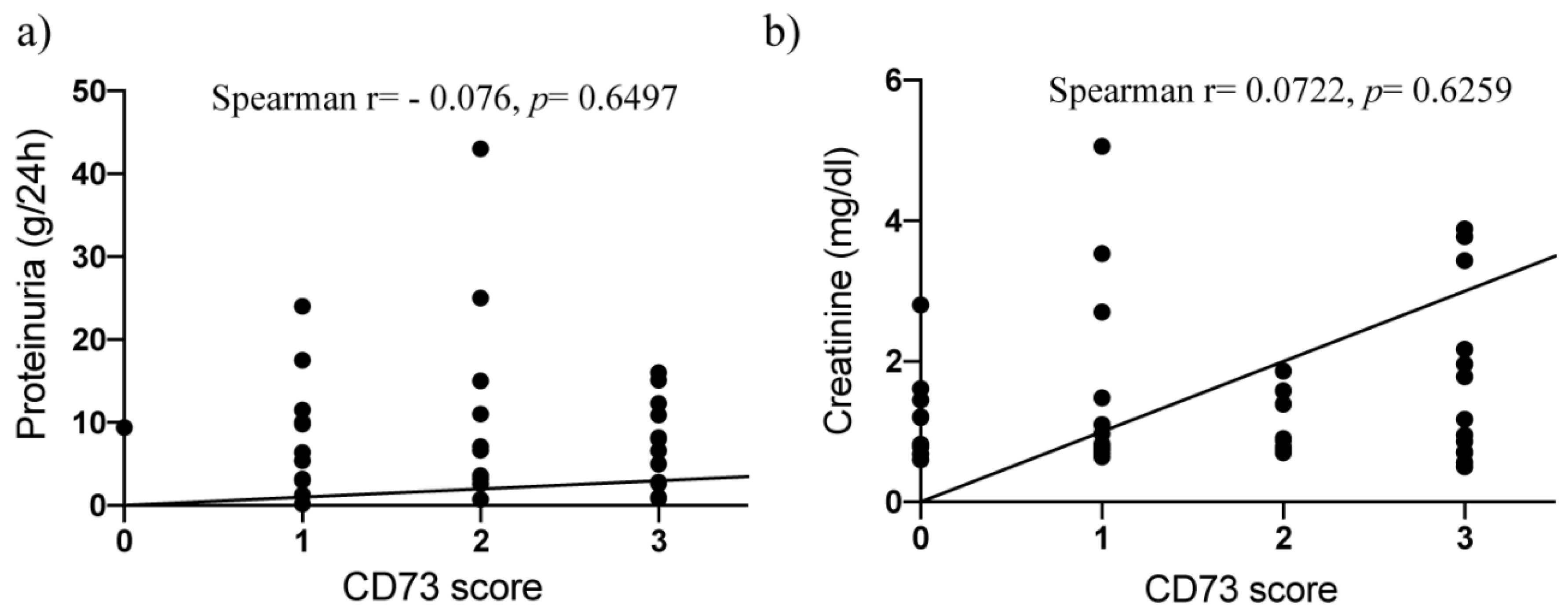
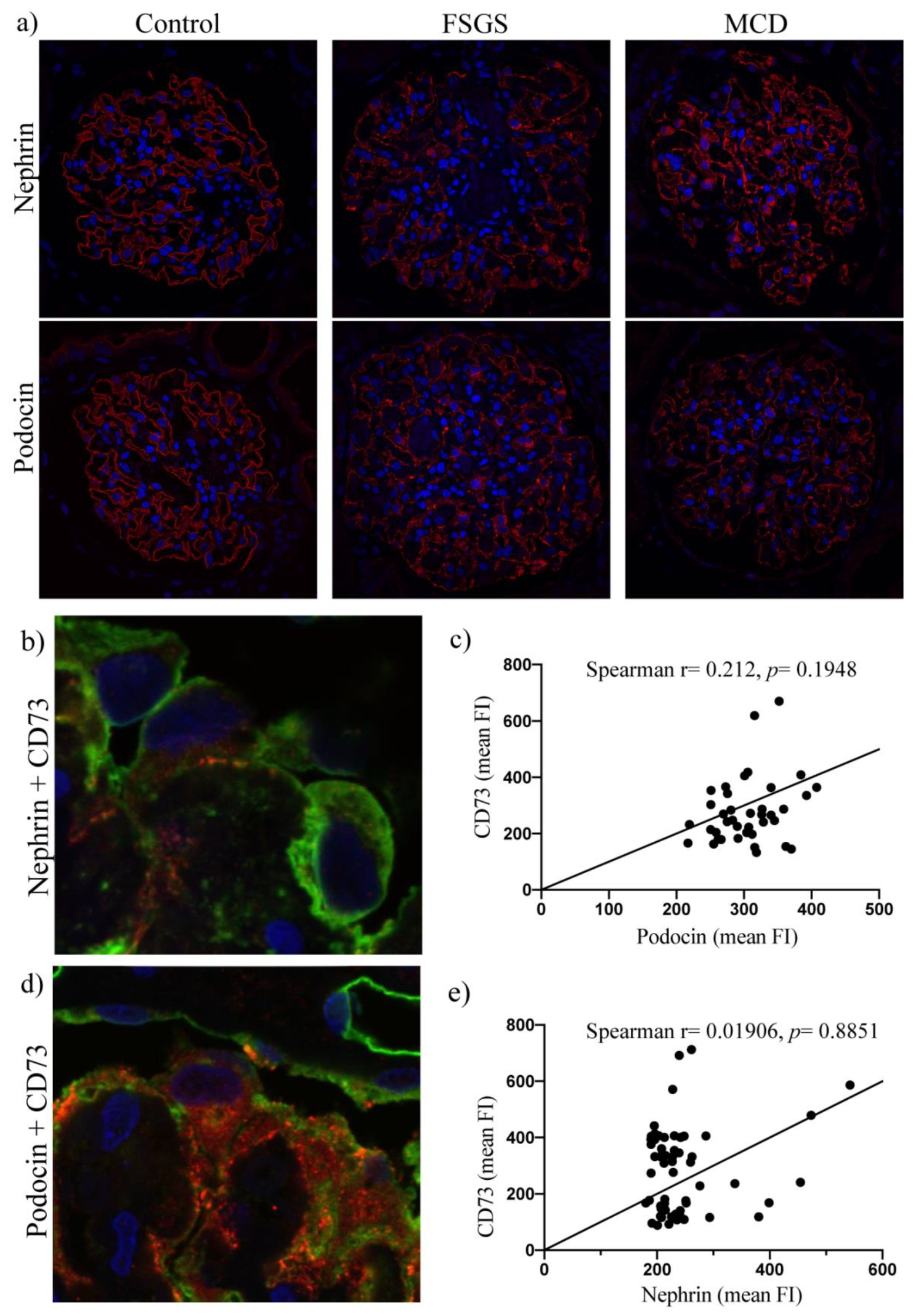
2.2. Surface Expression of CD73 Is Associated with Podocyte Injury in Human Kidney Disease
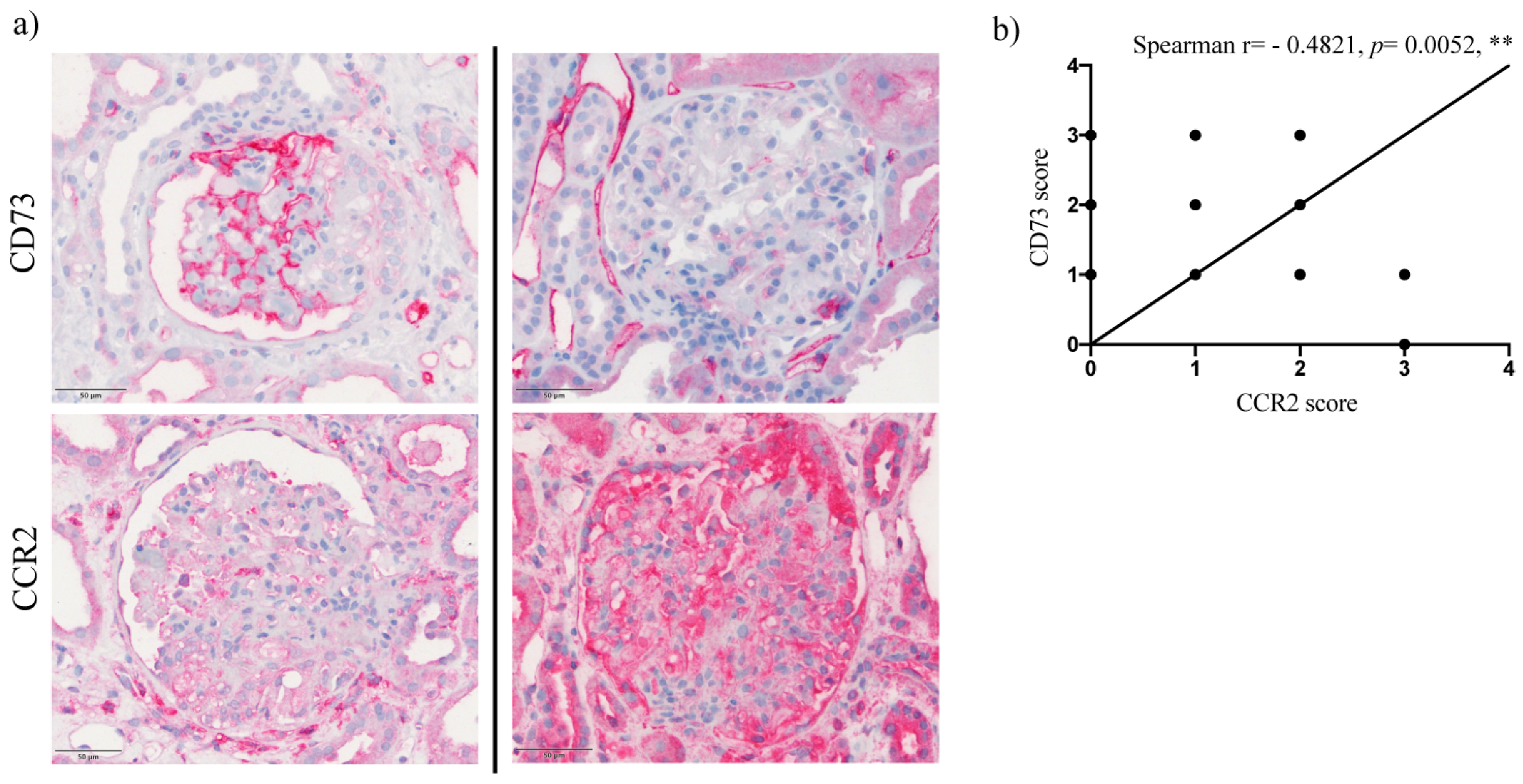
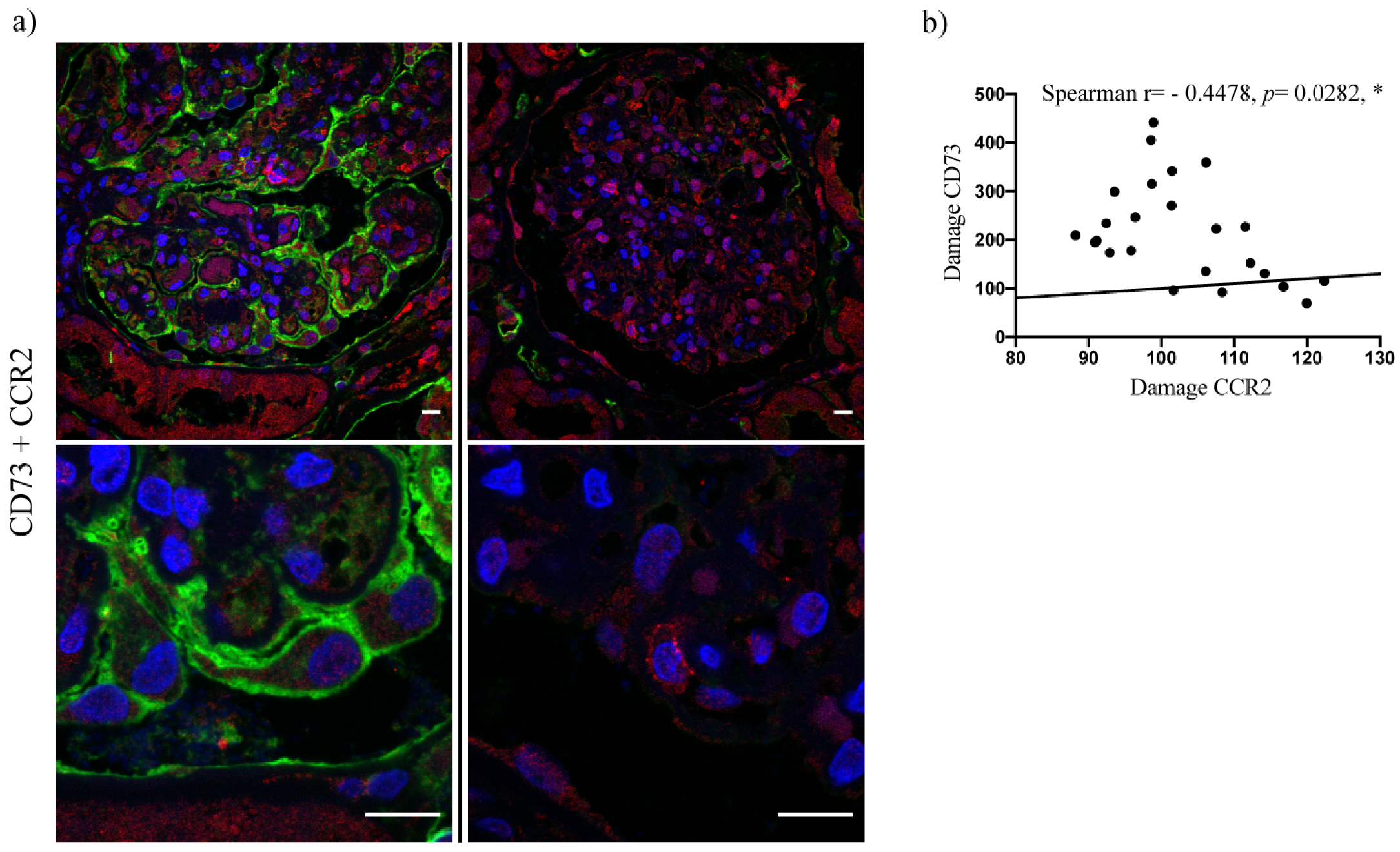
2.3. Upregulation of Podocyte-Derived CD73 in Human Glomerular Disease Negatively Correlates with Expression of the CCR2 Receptor and Does Not Correlate with Nephrin and Podocin Expression
3. Discussion
4. Material and Methods
4.1. Study Cohort
4.2. Histology, Immunohistochemistry, Immunofluorescence and Electron Microscopy
4.3. Light Microscopy and Morphometric Analyses
4.4. Confocal Microscopy
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burnstock, G. Endothelium-derived vasoconstriction by purines and pyrimidines. Circ. Res. 2008, 103, 1056–1057. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G. Unresolved issues and controversies in purinergic signalling. J. Physiol. 2008, 586, 3307–3312. [Google Scholar] [CrossRef]
- Antonioli, L.; Colucci, R.; Pellegrini, C.; Giustarini, G.; Tuccori, M.; Blandizzi, C.; Fornai, M. The role of purinergic pathways in the pathophysiology of gut diseases: Pharmacological modulation and potential therapeutic applications. Pharmacol. Ther. 2013, 139, 157–188. [Google Scholar] [CrossRef]
- Hasko, G.; Csoka, B.; Koscso, B.; Chandra, R.; Pacher, P.; Thompson, L.F.; Deitch, E.A.; Spolarics, Z.; Virag, L.; Gergely, P.; et al. Ecto-5′-nucleotidase (CD73) decreases mortality and organ injury in sepsis. J. Immunol. 2011, 187, 4256–4267. [Google Scholar] [CrossRef] [PubMed]
- Hasko, G.; Linden, J.; Cronstein, B.; Pacher, P. Adenosine receptors: Therapeutic aspects for inflammatory and immune diseases. Nat. Rev. Drug Discov. 2008, 7, 759–770. [Google Scholar] [CrossRef]
- Junger, W.G. Immune cell regulation by autocrine purinergic signalling. Nat. Rev. Immunol. 2011, 11, 201–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trautmann, A. Extracellular ATP in the immune system: More than just a “danger signal”. Sci. Signal. 2009, 2, pe6. [Google Scholar] [CrossRef]
- Kukulski, F.; Bahrami, F.; Ben Yebdri, F.; Lecka, J.; Martin-Satue, M.; Levesque, S.A.; Sevigny, J. NTPDase1 controls IL-8 production by human neutrophils. J. Immunol. 2011, 187, 644–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strater, N. Ecto-5′-nucleotidase: Structure function relationships. Purinergic Signal. 2006, 2, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonioli, L.; Pacher, P.; Vizi, E.S.; Hasko, G. CD39 and CD73 in immunity and inflammation. Trends Mol. Med. 2013, 19, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Knapp, K.; Zebisch, M.; Pippel, J.; El-Tayeb, A.; Muller, C.E.; Strater, N. Crystal structure of the human ecto-5′-nucleotidase (CD73): Insights into the regulation of purinergic signaling. Structure 2012, 20, 2161–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennon, P.F.; Taylor, C.T.; Stahl, G.L.; Colgan, S.P. Neutrophil-derived 5′-adenosine monophosphate promotes endothelial barrier function via CD73-mediated conversion to adenosine and endothelial A2B receptor activation. J. Exp. Med. 1998, 188, 1433–1443. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; Zhu, H.; Zhang, J.; Ouyang, W.; Tang, J.; Zhang, Y.; Qiu, L.; Liu, X.; Ding, Z.; Deng, X. CD73 Expression on Mesenchymal Stem Cells Dictates the Reparative Properties via Its Anti-Inflammatory Activity. Stem Cells Int. 2019, 2019, 8717694. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.F.; Eltzschig, H.K.; Ibla, J.C.; Van De Wiele, C.J.; Resta, R.; Morote-Garcia, J.C.; Colgan, S.P. Crucial role for ecto-5′-nucleotidase (CD73) in vascular leakage during hypoxia. J. Exp. Med. 2004, 200, 1395–1405. [Google Scholar] [CrossRef]
- Peralta, C.; Hotter, G.; Closa, D.; Prats, N.; Xaus, C.; Gelpi, E.; Rosello-Catafau, J. The protective role of adenosine in inducing nitric oxide synthesis in rat liver ischemia preconditioning is mediated by activation of adenosine A2 receptors. Hepatology 1999, 29, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Eckle, T.; Fullbier, L.; Wehrmann, M.; Khoury, J.; Mittelbronn, M.; Ibla, J.; Rosenberger, P.; Eltzschig, H.K. Identification of ectonucleotidases CD39 and CD73 in innate protection during acute lung injury. J. Immunol. 2007, 178, 8127–8137. [Google Scholar] [CrossRef]
- Eckle, T.; Krahn, T.; Grenz, A.; Kohler, D.; Mittelbronn, M.; Ledent, C.; Jacobson, M.A.; Osswald, H.; Thompson, L.F.; Unertl, K.; et al. Cardioprotection by ecto-5′-nucleotidase (CD73) and A2B adenosine receptors. Circulation 2007, 115, 1581–1590. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, T.; Changelian, P.S.; Bouis, D.; Noon, K.; Harada, H.; Lama, V.N.; Pinsky, D.J. Ecto-5′-nucleotidase (CD73) attenuates allograft airway rejection through adenosine 2A receptor stimulation. J. Immunol. 2010, 185, 1321–1329. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Chernogorova, P.; Ayata, K.; Gerlach, U.V.; Rughani, A.; Ritchey, J.W.; Ganesan, J.; Follo, M.; Zeiser, R.; Thompson, L.F.; et al. Deficiency of CD73/ecto-5′-nucleotidase in mice enhances acute graft-versus-host disease. Blood 2012, 119, 4554–4564. [Google Scholar] [CrossRef] [Green Version]
- Sung, S.J.; Li, L.; Huang, L.; Lawler, J.; Ye, H.; Rosin, D.L.; Vincent, I.S.; Le, T.H.; Yu, J.; Gorldt, N.; et al. Proximal Tubule CD73 Is Critical in Renal Ischemia-Reperfusion Injury Protection. J. Am. Soc. Nephrol. 2017, 28, 888–902. [Google Scholar] [CrossRef]
- Blume, C.; Felix, A.; Shushakova, N.; Gueler, F.; Falk, C.S.; Haller, H.; Schrader, J. Autoimmunity in CD73/Ecto-5′-nucleotidase deficient mice induces renal injury. PLoS ONE 2012, 7, e37100. [Google Scholar] [CrossRef] [Green Version]
- Nagata, M. Podocyte injury and its consequences. Kidney Int. 2016, 89, 1221–1230. [Google Scholar] [CrossRef]
- You, H.; Gao, T.; Raup-Konsavage, W.M.; Cooper, T.K.; Bronson, S.K.; Reeves, W.B.; Awad, A.S. Podocyte-specific chemokine (C-C motif) receptor 2 overexpression mediates diabetic renal injury in mice. Kidney Int. 2017, 91, 671–682. [Google Scholar] [CrossRef] [Green Version]
- Miao, Z.; Ertl, L.S.; Newland, D.; Zhao, B.; Wang, Y.; Zang, X.; Campbell, J.J.; Liu, X.; Dang, T.; Miao, S.; et al. CCR2 antagonism leads to marked reduction in proteinuria and glomerular injury in murine models of focal segmental glomerulosclerosis (FSGS). PLoS ONE 2018, 13, e0192405. [Google Scholar] [CrossRef]
- Lee, S.J.; Kang, J.S.; Kim, H.M.; Lee, E.S.; Lee, J.H.; Chung, C.H.; Lee, E.Y. CCR2 knockout ameliorates obesity-induced kidney injury through inhibiting oxidative stress and ER stress. PLoS ONE 2019, 14, e0222352. [Google Scholar] [CrossRef]
- Serbina, N.V.; Pamer, E.G. Monocyte emigration from bone marrow during bacterial infection requires signals mediated by chemokine receptor CCR2. Nat. Immunol. 2006, 7, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Tsou, C.L.; Peters, W.; Si, Y.; Slaymaker, S.; Aslanian, A.M.; Weisberg, S.P.; Mack, M.; Charo, I.F. Critical roles for CCR2 and MCP-3 in monocyte mobilization from bone marrow and recruitment to inflammatory sites. J. Clin. Investiga. 2007, 117, 902–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimoto, K.; Wada, T.; Furuichi, K.; Sakai, N.; Iwata, Y.; Yokoyama, H. CD68 and MCP-1/CCR2 expression of initial biopsies reflect the outcomes of membranous nephropathy. Nephron Clin. Pract. 2004, 98, c25–c34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eltzschig, H.K.; Eckle, T. Ischemia and reperfusion--from mechanism to translation. Nat. Med. 2011, 17, 1391–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eltzschig, H.K.; Eckle, T.; Mager, A.; Kuper, N.; Karcher, C.; Weissmuller, T.; Boengler, K.; Schulz, R.; Robson, S.C.; Colgan, S.P. ATP release from activated neutrophils occurs via connexin 43 and modulates adenosine-dependent endothelial cell function. Circ. Res. 2006, 99, 1100–1108. [Google Scholar] [CrossRef] [Green Version]
- Rajakumar, S.V.; Lu, B.; Crikis, S.; Robson, S.C.; d’Apice, A.J.; Cowan, P.J.; Dwyer, K.M. Deficiency or inhibition of CD73 protects in mild kidney ischemia-reperfusion injury. Transplantation 2010, 90, 1260–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tak, E.; Ridyard, D.; Kim, J.H.; Zimmerman, M.; Werner, T.; Wang, X.X.; Shabeka, U.; Seo, S.W.; Christians, U.; Klawitter, J.; et al. CD73-dependent generation of adenosine and endothelial Adora2b signaling attenuate diabetic nephropathy. J. Am. Soc. Nephrol. 2014, 25, 547–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddy, A.A.; Symons, J.M. Nephrotic syndrome in childhood. Lancet 2003, 362, 629–639. [Google Scholar] [CrossRef]
- Floege, J.; Amann, K. Primary glomerulonephritides. Lancet 2016, 387, 2036–2048. [Google Scholar] [CrossRef]
- Bertelli, R.; Bonanni, A.; Di Donato, A.; Cioni, M.; Ravani, P.; Ghiggeri, G.M. Regulatory T cells and minimal change nephropathy: In the midst of a complex network. Clin. Exp. Immunol. 2016, 183, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Lai, K.W.; Wei, C.L.; Tan, L.K.; Tan, P.H.; Chiang, G.S.; Lee, C.G.; Jordan, S.C.; Yap, H.K. Overexpression of interleukin-13 induces minimal-change-like nephropathy in rats. J. Am. Soc. Nephrol. 2007, 18, 1476–1485. [Google Scholar] [CrossRef] [Green Version]
- Le Berre, L.; Herve, C.; Buzelin, F.; Usal, C.; Soulillou, J.P.; Dantal, J. Renal macrophage activation and Th2 polarization precedes the development of nephrotic syndrome in Buffalo/Mna rats. Kidney Int. 2005, 68, 2079–2090. [Google Scholar] [CrossRef] [Green Version]
- Vivarelli, M.; Massella, L.; Ruggiero, B.; Emma, F. Minimal Change Disease. Clin. J. Am. Soc. Nephrol. 2017, 12, 332–345. [Google Scholar] [CrossRef] [Green Version]
- Kawachi, H.; Fukusumi, Y. New insight into podocyte slit diaphragm, a therapeutic target of proteinuria. Clin. Exp. Nephrol. 2020, 24, 193–204. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | MCD | FSGS | MGN | Amyloidosis | |
|---|---|---|---|---|---|
| n | 10 | 8 | 7 | 12 | 12 |
| Age (range) | 61.5 * (19–74) | 46.5 (20–65) | 44 (24–80) | 55.5 (24–80) | 60 (52–75) |
| Gender (n = females, %) | 7 (70) | 5 (62.5) | 5 (71.4) | 3 (25) | 1 (8.3) |
| Hypertension f (n, %) | 4 (40) | 5 (62.5) | 1 (14.3) | 4 (33.3) | 2 (16.67) |
| Diabetes mellitus (n, %) | 3 (30) | 0 (0) | 1 (14.3) | 2 (16.67) | 0 (0) |
| Creatinine, mg/dL (range) | 1.01 * (0.68–2.8) | 0.85 (0.64–1.48) | 0.88 (0.71–2.7) | 0.76 (0.5–3.88) | 1.88 (0.76–3.77) |
| Proteinuria, g/24 h (range) | 0.36 * (0.1–0.86) | 10.65 (3.2–24) | 9.4 (5–43) | 3.12 (0.2–15) | 6.75 (2.63–16) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popovic, Z.V.; Bestvater, F.; Krunic, D.; Krämer, B.K.; Bergner, R.; Löffler, C.; Hocher, B.; Marx, A.; Porubsky, S. CD73 Overexpression in Podocytes: A Novel Marker of Podocyte Injury in Human Kidney Disease. Int. J. Mol. Sci. 2021, 22, 7642. https://doi.org/10.3390/ijms22147642
Popovic ZV, Bestvater F, Krunic D, Krämer BK, Bergner R, Löffler C, Hocher B, Marx A, Porubsky S. CD73 Overexpression in Podocytes: A Novel Marker of Podocyte Injury in Human Kidney Disease. International Journal of Molecular Sciences. 2021; 22(14):7642. https://doi.org/10.3390/ijms22147642
Chicago/Turabian StylePopovic, Zoran V., Felix Bestvater, Damir Krunic, Bernhard K. Krämer, Raoul Bergner, Christian Löffler, Berthold Hocher, Alexander Marx, and Stefan Porubsky. 2021. "CD73 Overexpression in Podocytes: A Novel Marker of Podocyte Injury in Human Kidney Disease" International Journal of Molecular Sciences 22, no. 14: 7642. https://doi.org/10.3390/ijms22147642
APA StylePopovic, Z. V., Bestvater, F., Krunic, D., Krämer, B. K., Bergner, R., Löffler, C., Hocher, B., Marx, A., & Porubsky, S. (2021). CD73 Overexpression in Podocytes: A Novel Marker of Podocyte Injury in Human Kidney Disease. International Journal of Molecular Sciences, 22(14), 7642. https://doi.org/10.3390/ijms22147642