1. Introduction
Duchenne muscular dystrophy (DMD) is an X-linked recessive disorder caused by mutations in the gene encoding the dystrophin protein. The lack of a fully functional dystrophin protein results in severe and progressive weakness of both the skeletal and cardiac muscles. Although it is considered a rare disease, DMD is the most common form of muscular dystrophy affecting one in 3500 to 5000 boys worldwide [
1] with these boys generally diagnosed between the age of 3 to 5 years. The disease primarily affects the pelvic-girdle muscles at the initial stages, therefore as adolescents the boys are dependent on assisted mobility devices such as crutches or a wheelchair. As the disease progresses, in young adults affected with DMD, muscle dysfunction starts to affect cardiac and respiratory function, eventually requiring artificial ventilation to aid respiration. Most patients do not survive beyond 25–30 years of age, with cardiac failure being the major cause of death. In order to improve the quality and longevity of life, currently a comprehensive multi-disciplinary approach is used to manage DMD, involving pharmacological and non-pharmacological therapies that mostly improve symptoms, along with regular assessments of orthopedics, cardiology, pulmonology, osteology as well as mental health [
2,
3,
4].
In the recent years, DMD has been an attractive indication for gene-based therapies. There has been growing interest especially with therapies based on ‘exon skipping’ approaches, including Exondys51, Vyondys53 and Viltolarsen approved in the last year [
5]. While there are several studies currently in clinical trials aiming to restore dystrophin production, thereby treating the underlying cause of DMD, currently, most therapies aid by treating the symptoms [
5]. To date, the only symptom-modifying therapy approved for DMD are glucocorticoids, including prednisone and deflazacort, which improve skeletal muscle strength and function, whereas other therapies are used off-label to manage respiratory and cardiovascular symptoms. While gene therapies restoring expression of functional dystrophin in skeletal muscle and heart would ideally provide a cure for DMD, available therapies are effective only in a selected subpopulation of DMD patients, and novel improved gene therapies still need to prove efficacy. Moreover, while future therapies might be able to greatly reduce mortality, residual loss of muscle cells is likely to translate into chronic and progressive morbidity requiring further medical treatment.
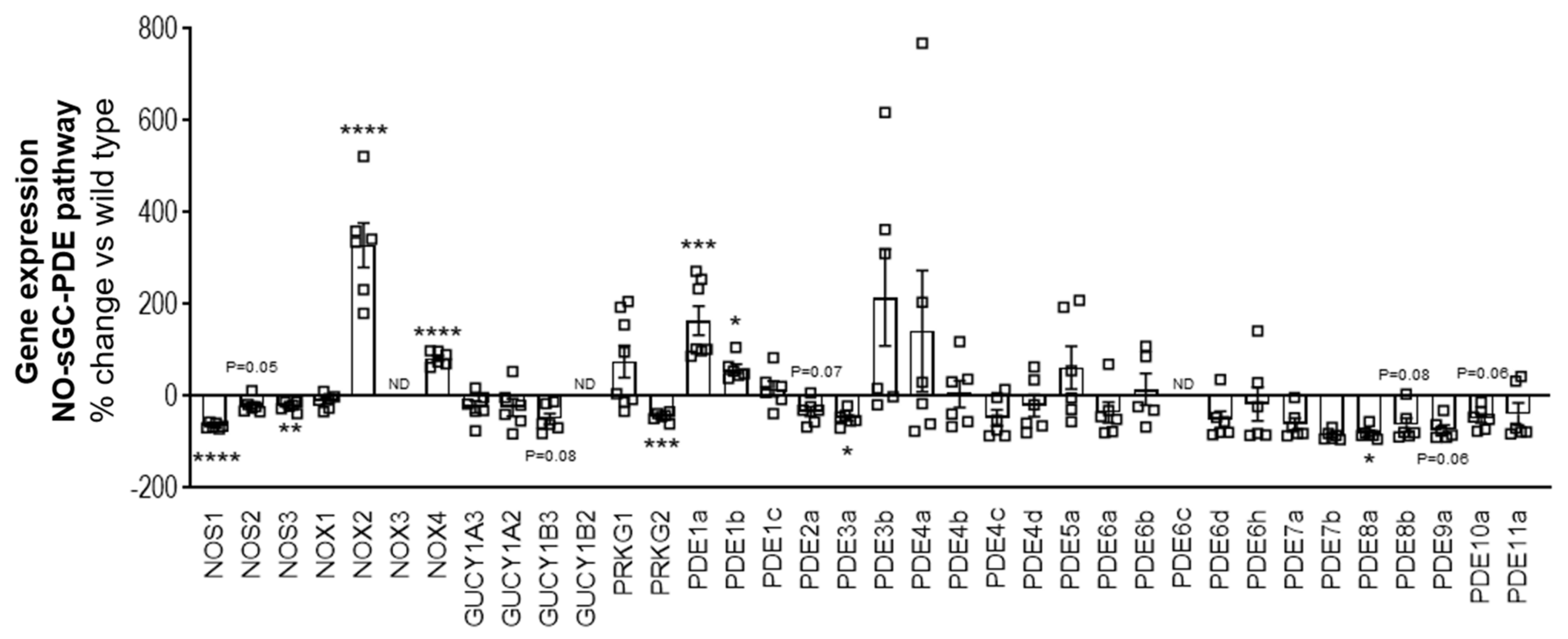
The dystrophin glycoprotein complex (DGC), present in the skeletal and cardiac muscle forms a link between the intracellular cytoskeleton and extracellular matrix and plays a key role in membrane sarcolemma stability, protecting the muscle during repeated contractions. DGC also serves as a scaffold to assemble a macromolecular complex with numerous, yet ill-defined functions. One of the subcomplexes linked to the DGC via the α-syntrophin protein is neuronal NO synthase (nNOS) [
6,
7]. The absence of functional dystrophin protein also displaces nNOS from the membrane, resulting in impaired skeletal muscle perfusion and exercise efficiency in mdx mice as well as in DMD boys [
8]. This occurs directly as a consequence of impaired NO synthesis and cGMP production in the skeletal muscle. Therefore, one treatment approach to improve exercise efficiency could target the low endogenous cGMP production.
The nitric oxide-soluble guanylyl cyclase–cyclic guanosine monophosphate–phosphodiesterase (NO-sGC-cGMP-PDE) pathway is known to play an important role in skeletal muscle function and exercise. During exercise, in order to meet the metabolic demands of the skeletal muscle, there is increased dilatation and perfusion of the blood vessels in the skeletal muscles, described as functional sympatholysis. NO improves functional sympatholysis by stimulating the sGC enzyme, triggering cGMP production and causing vasodilatation and perfusion of the skeletal muscles, which in turn increases oxygen delivery to the working skeletal muscles and improves exercise efficiency [
7,
9]. In addition to perfusion matching via functional sympatholysis, the NO-sGC-cGMP axis has been suggested to modulate muscle microtubule organization and mitochondrial oxidative phosphorylation ultimately regulating muscle fatigability [
7].
Therefore, cGMP increasing pharmacological mechanisms came into focus for treating muscular dystrophies. Previously, PDE5 inhibitors, which elevate cGMP levels by preventing its breakdown, have been assessed as a potential therapeutic for the treatment of DMD. Preclinical studies showed that administration of PDE5 inhibitors such as sildenafil or tadalafil reduced muscle damage [
10], reversed cardiac dysfunction and delayed the onset of cardiomyopathy [
11,
12], improved diaphragm contractility and reduced fibrosis [
13] in mdx mice. In a small-scale clinical trial with 10 DMD patients who displayed functional ischemia despite being on corticosteroids, treatment with sildenafil or tadalafil enhanced blood flow and improved exercise-induced functional sympatholysis in a dose-dependent manner in the skeletal muscle [
14]. In addition, tadalafil was able to alleviate muscle ischemia in eight out of nine patients with Becker muscular dystrophy (BMD), a milder form of muscular dystrophy which is also characterized by dystrophin mutations [
15]. Despite these positive outcomes in smaller pilot trials, a large-scale Phase III trial assessing the use of tadalafil for the treatment of DMD did not reach its primary endpoint of improving six-minute walking distance after 48 weeks of treatment [
16]. While tadalafil also showed no signals in a comprehensive ambulatory assessment used as secondary endpoint, it was interesting to note that in a subgroup analysis performed on a prespecified subgroup of boys over 10 years of age, the total and shoulder-level upper limb scores showed lesser decline following treatment with tadalafil compared to placebo treatment [
16]. This suggests that PDE5 inhibitors could show less decline in ambulation in younger and less disabled DMD boys [
6], and in addition, it also suggests that enhancement of cGMP with PDE5 inhibitors was beneficial but still under the critical threshold to show clinical benefit.
The failure or low efficacy of PDE5 inhibitors in DMD patients could have been due to the reason that the PDE5 inhibitors rely on NO and the downstream cGMP production in order to effectively work by inhibiting the breakdown of cGMP. In the early 2000s, a new class of cGMP enhancing compounds, known as sGC stimulators were discovered [
17,
18]. These compounds target the sGC enzyme and can actively cause cGMP production without requiring endogenous NO but also synergistically potentiate the effects of NO, even if the levels are very low [
17]. Therefore, in this study we examined the therapeutic potential of the sGC stimulator, BAY-747, in vivo in a mouse model of DMD (mdx/mTR
G2 mice). BAY-747 (
N-(2-amino-2-methylbutyl)-8-[(2,6-difluorobenzyl)oxy]-2,6-dimethylimidazo [1,2-a]pyridine-3-carboxamide) belongs to a new chemical class of long-acting sGC stimulators and shares its molecular mode of action with riociguat and vericiguat [
19,
20]. Treatment with BAY-747 improved skeletal muscle function, reduced serum creatine kinase levels, and biomarkers of inflammation and fibrosis in the skeletal muscle with no effect on myeloid cell infiltration and collagen accumulation. Therefore, findings from this study suggest that sGC stimulators might have treatment potential in muscular dystrophies.
4. Discussion
DMD is still a rare but devastating disease with a substantially reduced life expectancy. While the efforts for gene therapies for the treatment of DMD are ongoing, the high technical risk associated with high costs involved in the research and development of gene therapeutics for rare diseases could be a limiting factor [
25]. Therefore, researchers are still investigating the use of compounds with a known and disease-relevant mode of action, that are already in late-stage development or in the market, which provide the advantages of shorter development times and a well described safety profile. The compound class of sGC stimulators can stimulate the sGC enzyme independent of NO and can thereby increase cGMP levels in muscle cells, which are thought to be beneficial in DMD [
7,
13]. We therefore investigated in a preclinical “humanized mouse model of Duchenne muscular dystrophy” (mdx/mTR
G2 mice) [
26,
27], the effects of the novel sGC stimulator, BAY-747.
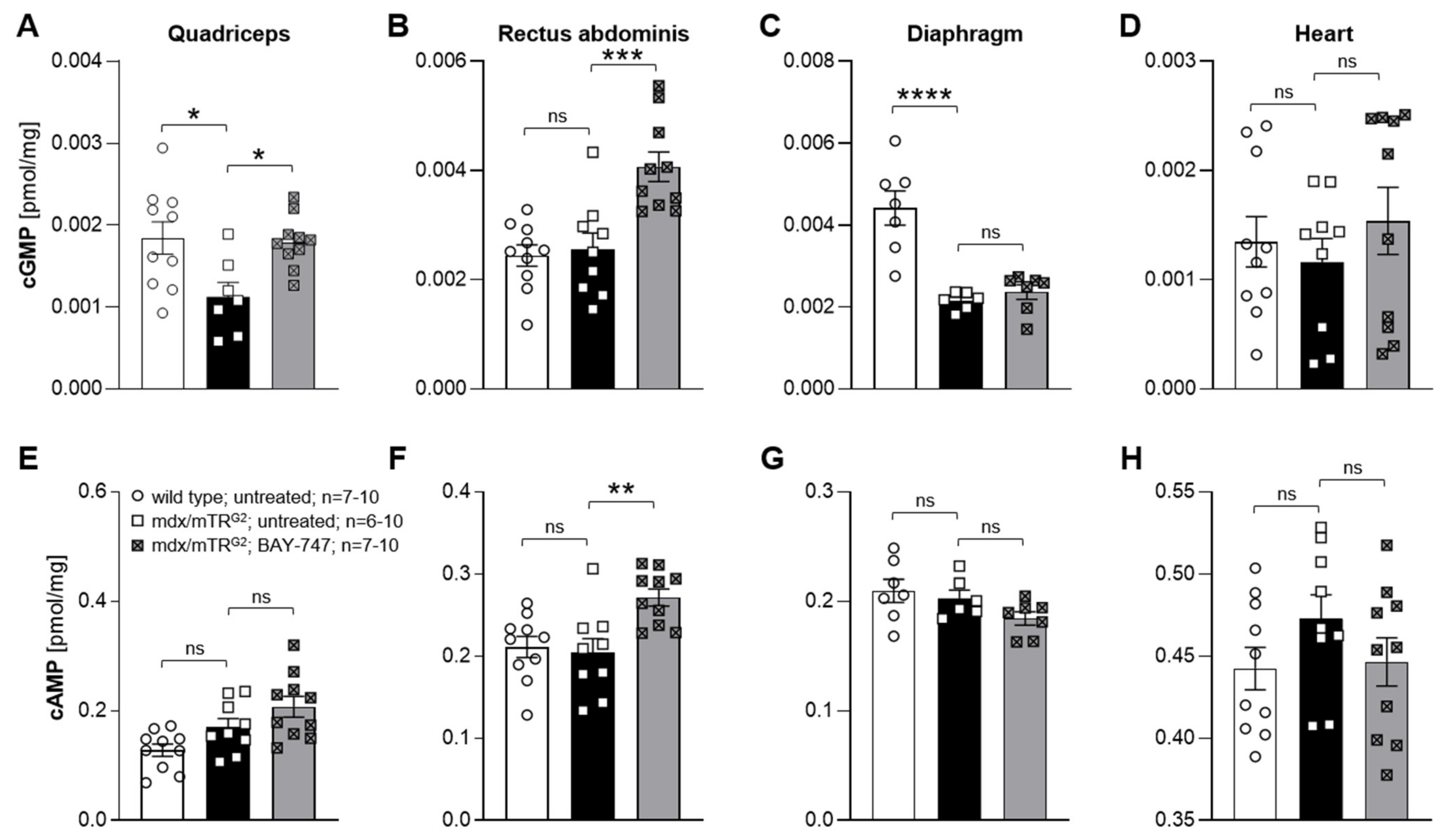
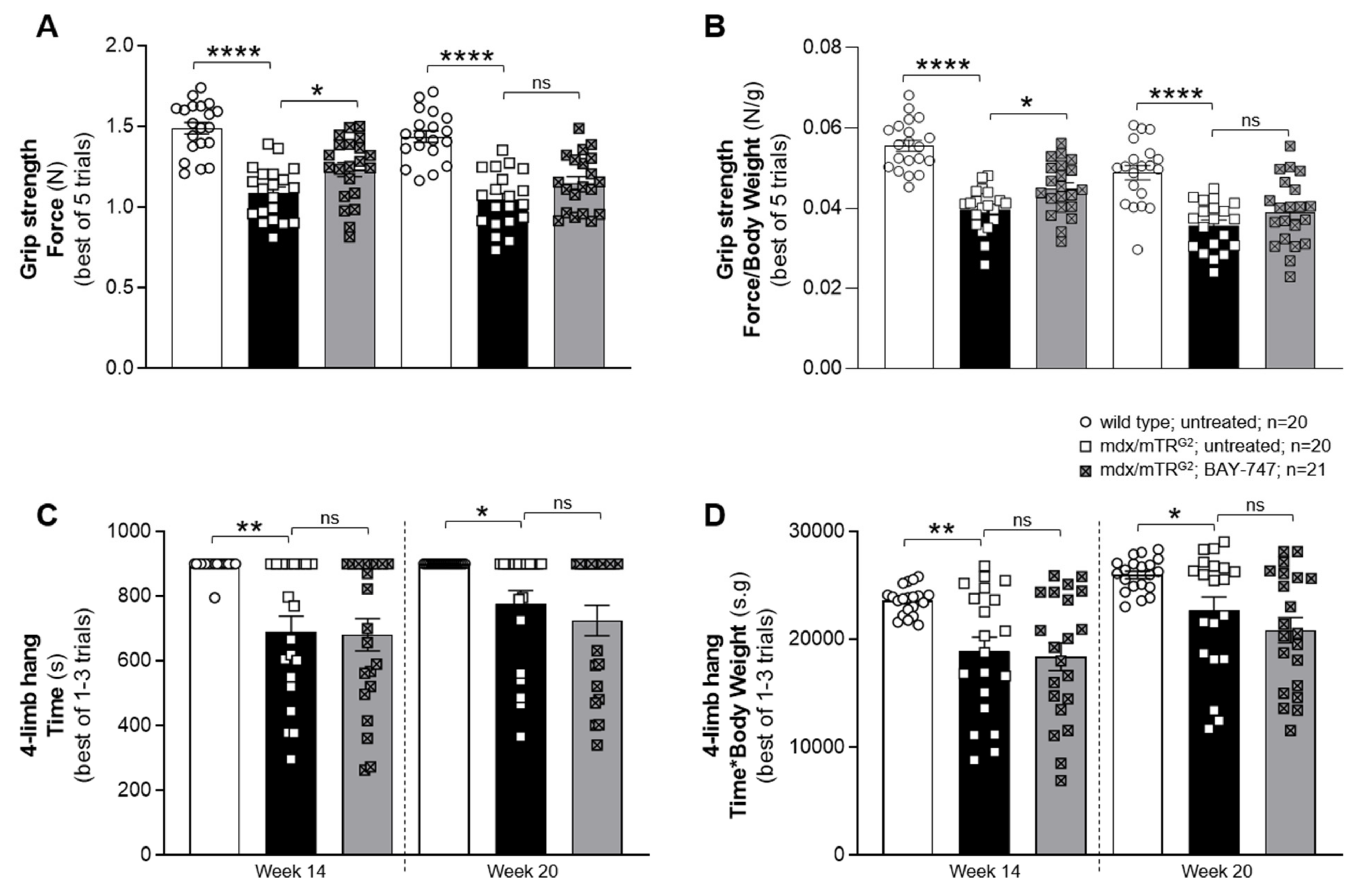
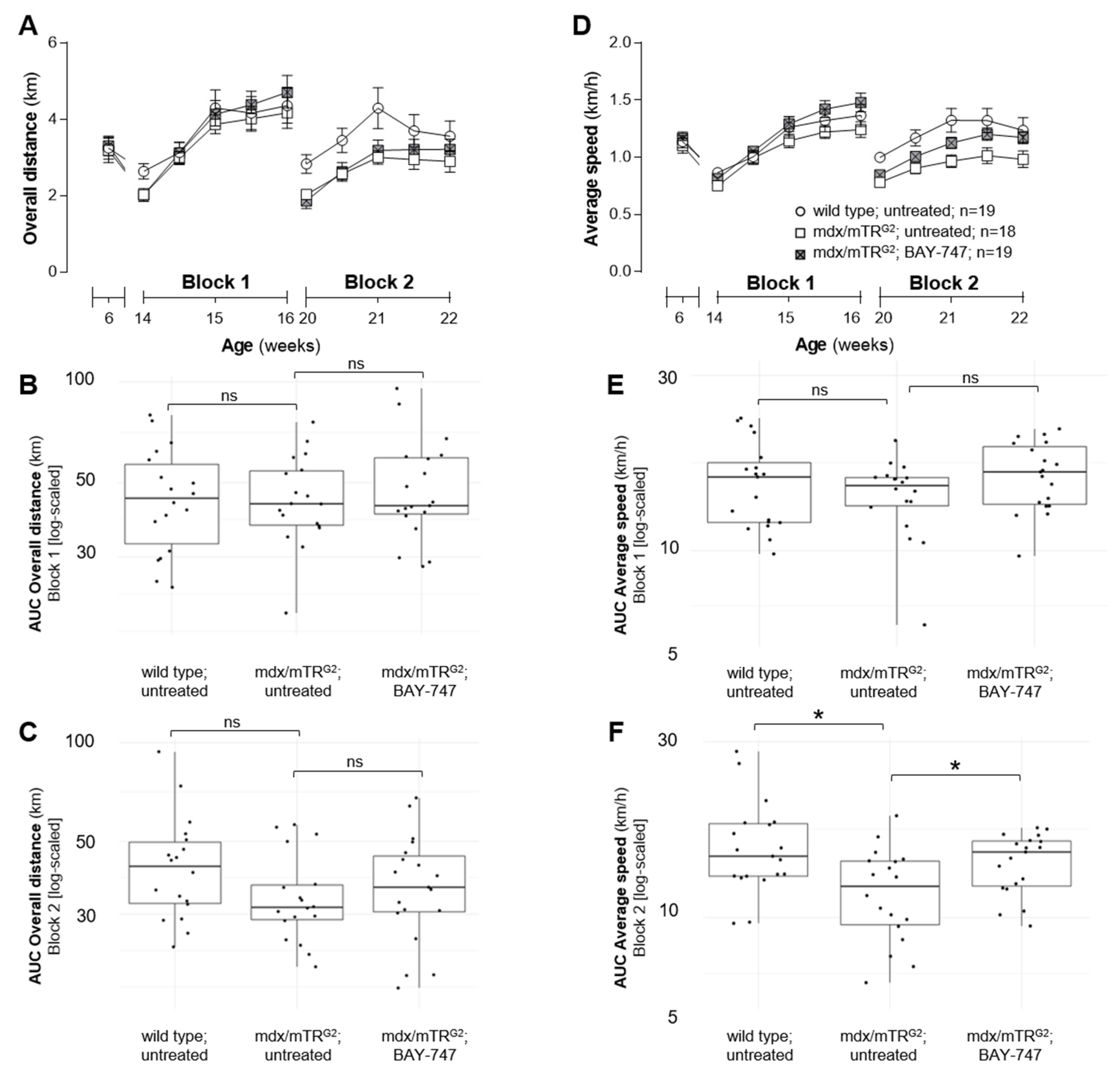
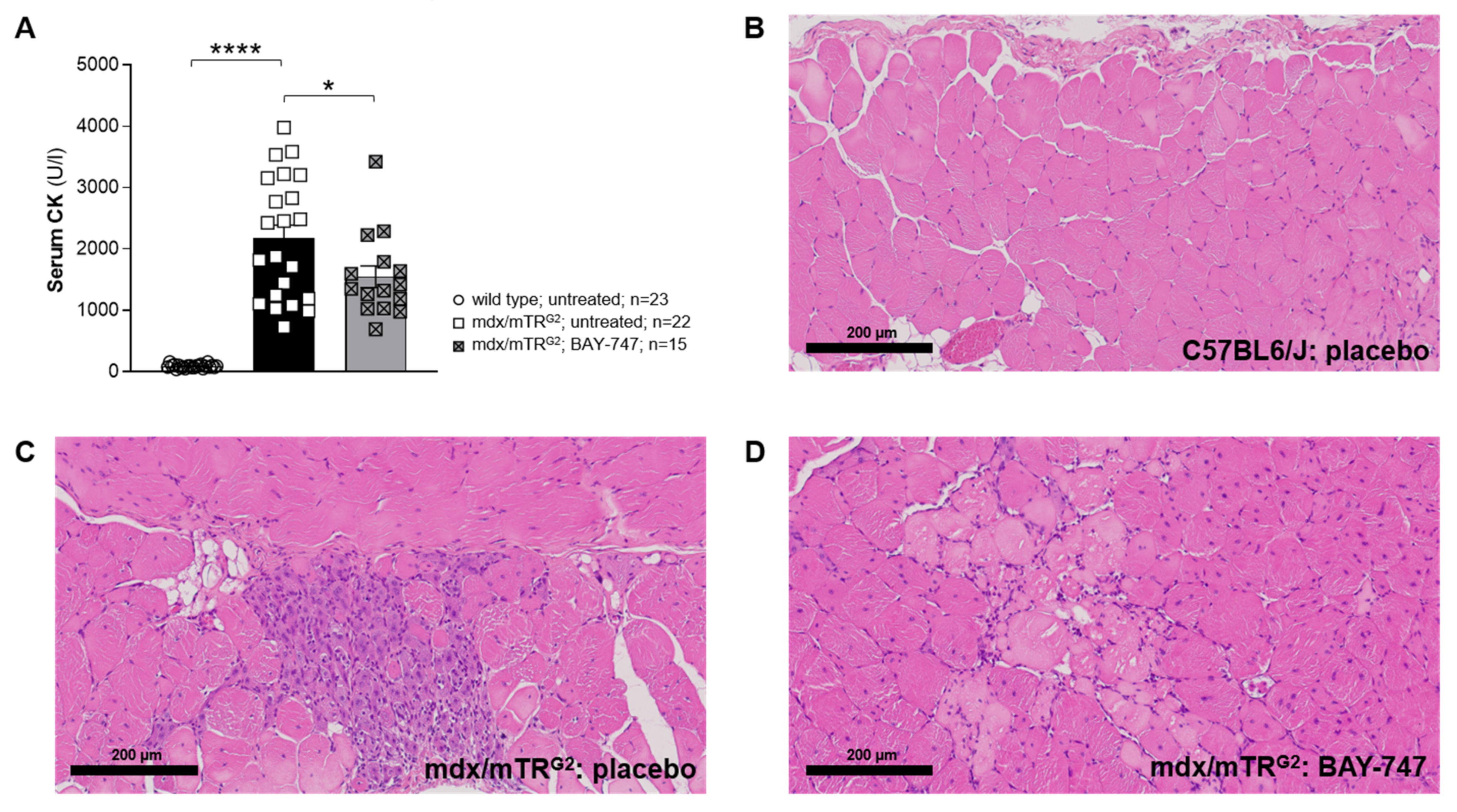
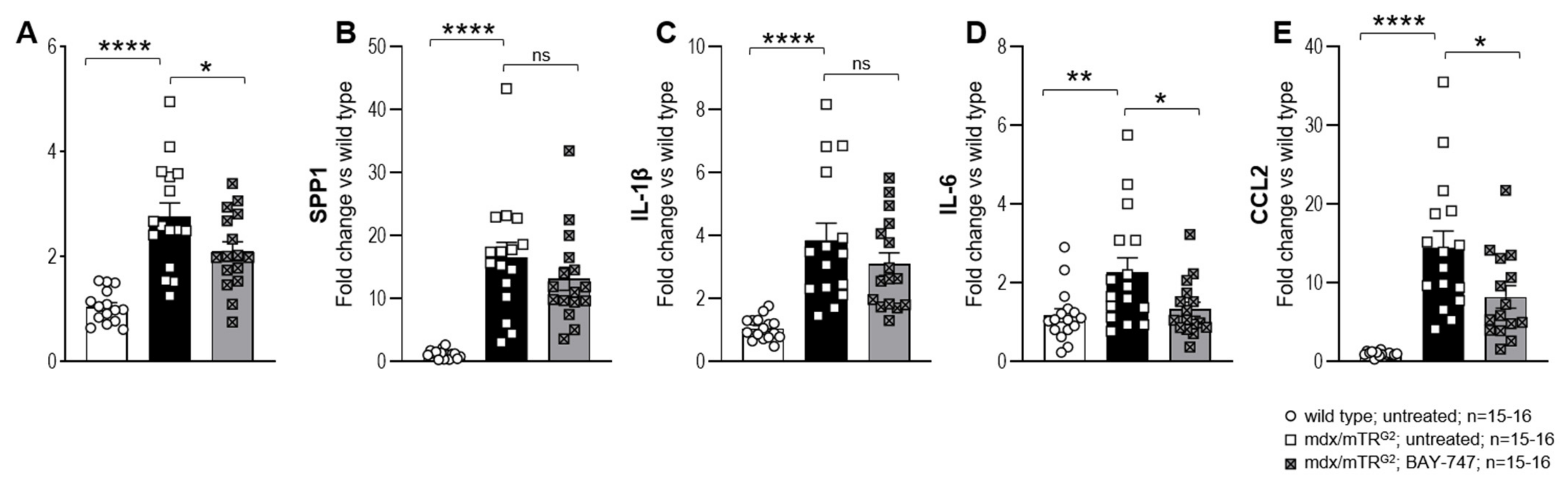
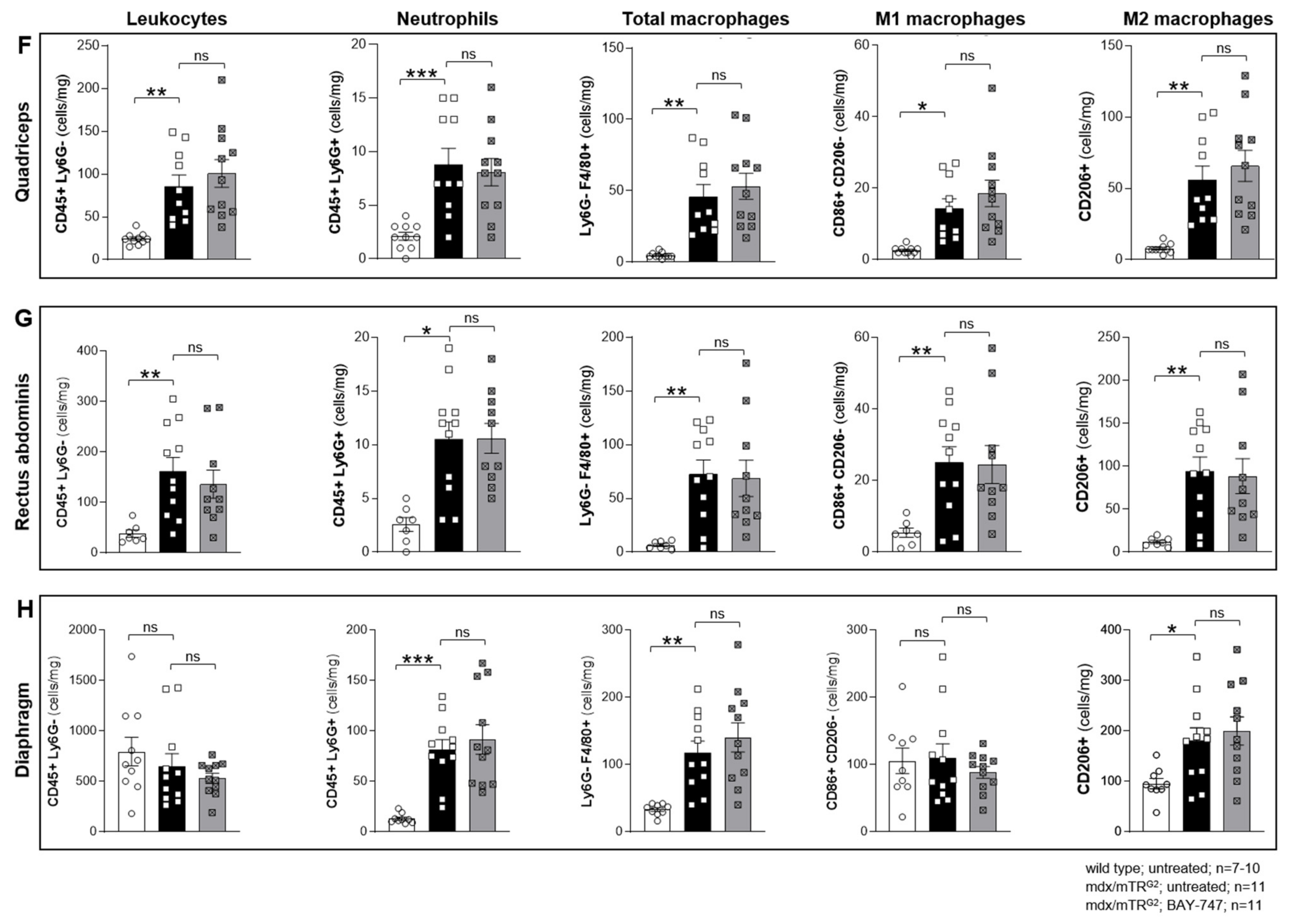
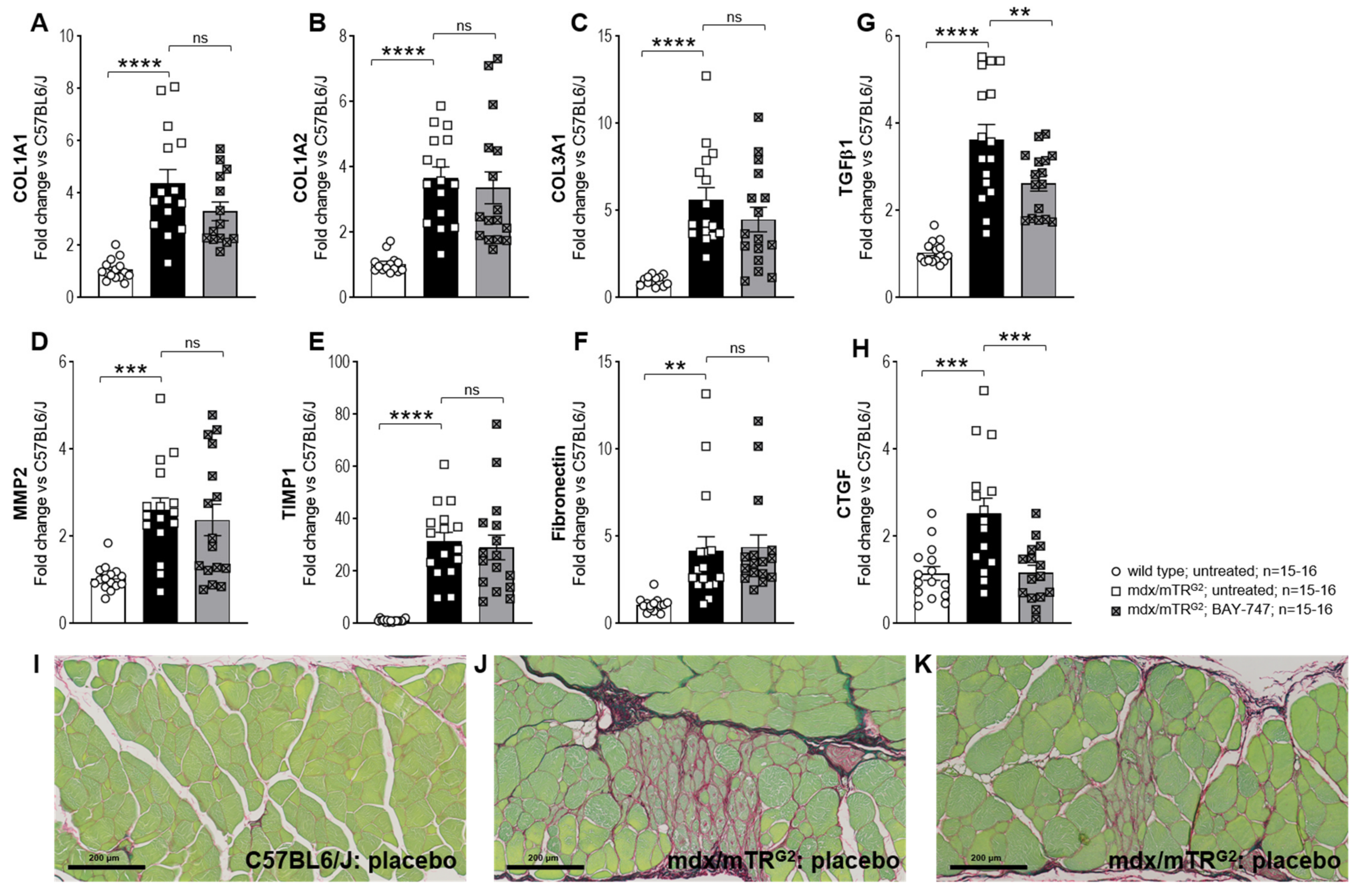
The major findings from this study highlight that, a chronic, 16-week treatment with BAY-747 results in an improvement of skeletal muscle function and damage in mdx/mTRG2 mice. Although no improvements were observed in the four-limb hang and overall voluntary running distance, mdx/mTRG2 mice that consumed food supplemented with BAY-747 had improved forelimb grip strength and running speed in the voluntary running wheel as well as reduced levels of a key biomarker of skeletal muscle damage, serum creatine kinase. Furthermore, BAY-747 treatment also showed reduced expression of markers of inflammation and fibrosis at the gene expression level but not at the histopathological level. The observed improvements are likely due to the stimulation of the NO/sGC/cGMP pathway by the sGC stimulator since cGMP production in the quadriceps and rectus abdominis muscles of mdx/mTRG2 mice were found to be increased after BAY-747 treatment.
In our study, we found that the beneficial effects of treatment of mdx/mTR
G2 mice with BAY-747 were moderate. This is at least in part in contrast to earlier results in preclinical models with cGMP-increasing drugs like PDE5 inhibitors [
10,
13]. Sildenafil and tadalafil have shown various therapeutic benefits including reversing cardiac dysfunction [
11] and improved respiratory muscle strength and reduced fibrosis of the diaphragm [
13]. This enhanced efficacy in these previous studies might be due to the different dosage regimens. Before our treatment study, we performed a pharmacokinetic and pharmacodynamic dose-finding study (Supplementary Results Section) with 50 and 150 mg/kg of BAY-747. The 150-mg/kg dose of BAY-747 had a moderate blood pressure-reducing effect, which indicated target engagement in the resistance vessels of the mice. We decided not to increase the dose further since a significant blood pressure drop could have a potential impact on the functional muscle read outs. To address whether the chosen dose allowed for proper exposure to striated muscle tissues, we quantified the concentration of cGMP in different muscles and found an increase of cGMP content in the quadriceps and the rectus abdominus, suggesting target engagement in these muscle types as well. In contrast, in the diaphragm, the concentration of cGMP remained unchanged after BAY-747 treatment. Since we used whole muscle homogenates for the quantification of cyclic nucleotides, we could not exclude higher local concentrations in the diaphragm due to the high intracellular compartmentalization of the cGMP signaling cascade. Overall, there is only little information on cGMP concentrations in muscles and it therefore would be very interesting to confirm and extend these finding on cNMPs in the future, which will also help to better understand the role of cGMP in striated muscles. Concomitantly, the levels of cAMP were also increased in the rectus abdominus, which could be due to cGMP to cAMP crosstalk via cGMP-induced inhibition of PDE3.
Additionally, the treatment effects in our study might be also impacted by the time of initiation of the BAY-747 treatment. Early diagnosis and pharmacological intervention are crucial for the treatment and management of symptoms associated with DMD. Boys with DMD are usually diagnosed between the ages of three to five years and treatments and interventions start immediately post-assessment. In our study, treatment was initiated at six weeks of age in mdx/mTR
G2 mice, and this is perhaps not early enough to show the full potential benefits of BAY-747. In fact, in a recent study that also used the mdx/mTR mouse model of DMD, ifetroban, an antagonist of the thromboxane-prostanoid receptor, treatment was initiated as early as three weeks of age and continued for an additional three months [
28].
From a mechanistical perspective, the failure of PDE5 inhibitor drugs might be due to the low NO production in DMD. In addition, we could also not exclude that oxidative stress could at least in part blunt NO as well as sGC stimulator effects. According to previously published data it is evident that DMD is also a condition associated with oxidative stress in the skeletal muscles (Tidball 2018, Grounds 2020). Oxidative stress does not only impair NO binding to the sGC enzyme thereby interrupting NO/sGC/cGMP signaling, but also limits the effects of sGC stimulators like BAY-747. Therefore, another class of sGC modulating compounds known as sGC activators, which activate oxidized and heme free sGC and restore sGC signaling under oxidative stress conditions [
29] might be a more effective alternative to sGC stimulators. In the future, head-to-head comparisons of sGC stimulators and sGC activators are needed to fully exploit the treatment opportunities of cGMP increase by sGC agonists in DMD.
Interestingly, in our study, we observed different degrees of improvement and efficacy in different skeletal muscles such as the quadriceps, rectus abdominis, and diaphragm. Skeletal muscles can be classified according to various categories, for example based on the predominant fiber type, i.e., slow (Type I) or fast (Type II) twitch, type of energy used i.e., aerobic (oxidative) or anaerobic (glycolytic) etc. [
30]. The primary mechanism of action of NO-sGC-cGMP modulators in improving exercise efficiency is by causing vasodilatation and thereby improving the amount of oxygen delivered to the muscle fibers, as well as reducing the cost of ATP and oxygen used for muscle contraction and ATP resynthesis [
9]. Our running wheel data demonstrated that mdx/mTR
G2 mice treated with BAY-747 displayed good sprint but not endurance efficiency, as they were able to run the same distance as untreated mdx/mTR
G2 mice but were able to run at a much faster pace. Therefore, it appears that chronic treatment with BAY-747 plays a more effective role in muscles with fast glycolytic fibers which are able to generate strong contractions but only for a short while as they fatigue much faster when compared to slow oxidative fibers [
31].
Overall, chronic sGC stimulation with BAY-747 improved various parameters in mdx/mTR
G2 mice. However, the treatment effects were moderate and dose escalation might not be appropriate due to blood pressure lowering. Therefore, sGC stimulators might provide an adjunctive treatment modality for DMD patients rather than a stand-alone option, similar to current treatment regimens which include combinations like MR-antagonists with ACE-inhibitors [
32]. Additional preclinical studies are necessary to substantiate the treatment effects of sGC stimulators in DMD especially when combined with MR-antagonists or ACE-inhibitors. In addition, since DMD is associated with inflammation and increased oxidative stress it might be very interesting to investigate the efficacy of sGC activators as a stand-alone option in mdx mice for evaluation of the treatment potential of this drug class in DMD.
5. Materials and Methods
5.1. Animal Model and Experimental Design
All experiments were performed according to the guidelines approved by the local animal welfare authorities for the German state of North-Rhine Westphalia (Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen; 400a152).
The mdx
4cv/mTR
G2 (hereon referred to as mdx/mTR
G2) mouse model has previously been described as the “humanized mdx mouse model of DMD” by Blau and colleagues [
26,
33]. The DMD
mdx4cv mice have a nonsense mutation in Exon 53 of the gene encoding for dystrophin, coding for a truncated dystrophin protein, whereas the mTR
−/− mice are homozygous for the telomerase RNA component null allele and lack telomerase activity. Male mdx/mTR
G2 mice approximately 6–7 weeks of age were used for the study reported in this paper. Age-matched C57BL6/JRj (Janvier Labs, Le Genest-Saint-Isle, France) mice were used as wild type controls. The license for the mdx/mTR
G2 mice was obtained from Stanford University and breeding pairs were obtained from Jackson Laboratories, Bar Harbour, ME, USA (Stock No.: 023535), and the breeding of both the mdx/mTR
G2 and C57BL6/JRj mice were performed by Janvier Labs. The mice were housed in individual cages under standard conditions including 12-h light/dark cycle, approximately 22 °C temperature and 55% humidity. Standard mouse food was commercially purchased and was provided
ad libitum as is (untreated) or supplemented with 150 mg of the sGC stimulator BAY-747 (
N-(2-amino-2-methylbutyl)-8-[(2,6-difluorobenzyl)oxy]-2,6-dimethylimidazo[1,2-a]pyridine-3-carboxamide) per kg of food (Ssniff Spezial-diäten GmbH, Soest, Germany) for a total treatment duration of 16 weeks. Body weight and food consumption was monitored every week. At the end of the study, animals were killed under deep anesthesia and tissue and organ samples were collected for further analysis and histopathological scoring.
5.2. Radiotelemetry for Short-Term Monitoring of Blood Pressure and Heart Rate
To examine the effect of BAY-747 on blood pressure and heart rate, radiotelemetry was used. The 20-week-old adult C57BL6/J and mdx/mTRG2 mice were implanted with a telemeter probe (Model TA11PA-C10, Data Sciences International, St. Paul, MN, USA) for monitoring blood pressure in conscious mice. After a 10-day recovery period, baseline recordings were obtained over 4 days. Following this, all mice were given food either untreated or supplemented with BAY-747 50 mg/kg or 150 mg/kg for 1 week, and BP was measured in a cross-over study manner with a wash-out period of 1 week (untreated food) in between.
5.3. Measuring Plasma Concentrations of BAY-747
Wild type and mdx/mTRG2 mice were fed food supplemented with BAY-747 (150 mg/kg) for 5 days, following which plasma was collected at various time points starting at the end of the dark cycle (active/feeding period), for analysis of plasma concentration. BAY-747 concentration was analyzed in the plasma using an LC-system for mass separation (Kinetex 5 µm C18 100 A LC Column 150 × 4.6 mm) coupled to a 4500 Triple Quad Sciex mass analyzer (positive mode; Framingham, MA, USA). A generic internal standard was added to the samples. A 5-point calibration curve and quality control samples were used for relative quantification.
5.4. Voluntary Running Wheel
Prior to treatment, mice were placed in individual running wheel cages (built in-house) for one week. Each running wheel was calibrated at least once every week to ensure accurate recording of wheel revolutions. Since the mice are more active in the night period, the data were recorded daily between 3 pm and 9 am the following day. Following one week of voluntary running, mice were randomized into treatment groups based on the overall distance run. In order to observe treatment effects that are not biased by chronic exercise, we used a staggered approach in assessing the effects of the running wheel. To do this, the mice were housed in their regular cages (sedentary) and only moved to the running wheel cages when required. In our study, mice were placed in the running wheel cages during weeks 8 to 10 and weeks 14 to 16 of treatment, and overall distance and average speed run every 2–3 days was calculated during these periods.
5.5. Forelimb Grip Strength
Forelimb grip strength was assessed using a force transducer (Sauter GmbH, Balingen, Germany) as described in the Treat-NMD SOP (use of grip strength meter to assess the limb strength of mdx mice—DMD_M.2.2.001). The assessment was performed in an unblinded manner, by the same experienced operator, and always in the same room, with temperature and humidity controlled at the same time. In brief, a grid-like mesh was attached horizontally to the force transducer, which was set to 0 prior to each measurement. The mouse was lifted by its tail and brought close to the mesh allowing the mouse to grab the mesh with its fore paws. The mouse was slowly pulled back until it lets go off the mesh, and the maximum force built up in resistance was recorded by the force transducer. This was repeated 5 times, with a gap of 1 min between each measurement. The average force measured was normalized to body weight.
5.6. Four-Limb Hang
The four-limb hang assessment was performed as described in the Treat-NMD SOP (The use of four limb hanging tests to monitor muscle strength and condition over time—DMD_M.2.1.005). Since we observed that most wild type mice were able to hang for prolonged periods, for this assessment all the mice were allowed to hang for a maximum of 900 s. If they did not succeed in doing so the first time, they were made to repeat the trial a second or third time with a 10-to-15-min resting period between each trial.
5.7. Measurement of Serum Creatine Kinase
Whole blood was collected through the inferior vena cava and left to clot at room temperature for 30 min. The samples were then centrifuged at 1000 rcf and 4 °C for 10 min, following which the serum was transferred to a fresh tube and stored at −80 °C, until ready for analysis. Creatine kinase levels in the serum were measured in the automated clinical chemistry analyzer, ADVIA® Chemistry XPT System (Siemens, Munich, Germany) according to the kit’s instruction for the creatine kinase assay (Siemens, Munich, Germany).
5.8. Gene Expression Analysis by RT/qPCR
Total RNA was extracted from the snap-frozen quadricep tissue using the miRNeasy Mini Kit (Qiagen, Hilden, Germany) according to kit instructions. RNA was quantified and controlled for purity (260/280 absorbance ratio ~2.0) using the NanoDrop. A total of 1µg of RNA was reverse transcribed to cDNA using the ImProm-IITM Reverse Transcription System (Promega, Madison, WI, USA) for use as a template for real-time qPCR analysis. The TaqMan primer/probe sequences for the consecutive qPCR are listed in
Supplementary Table S1 (sequences were designed by Primer Express 1.5 Software, Applied Biosystems). qPCR was performed with the qPCR Master Mix Plus (Eurogentech; Seraing, Belgium) on a 7900 HT Fast Real-Time PCR System (Applied Biosystems, ThermoFischer Scientific, Waltham, MA, USA). Ct values were determined by Applied Biosystems’s SDS Software (version 2.4), normalized to the housekeeping gene Rpl32. To calculate the fold-change in mRNA expression relative to wild type control samples, the comparative Ct method was used [
34].
5.9. Flow Cytometry
Single-cell suspensions of various skeletal muscles (quadriceps, rectus abdominis and diaphragm) were obtained by digesting the tissues using the Multi Tissue Dissociation Kit 1 (Miltenyi Biotec, Auburn, CA, USA) according to kit instructions. The cells were passed through a 70 µM cell strainer, erythrocytes were lysed using 1× BD Pharm Lyse™ (BD Biosciences, Franklin Lakes, NJ, USA) and washed with autoMACS® Running Buffer (Miltenyi Biotec, USA). Fc receptors were blocked with CD16/CD32 (BD Biosciences, USA) for 10 min. Extracellular surface markers were stained with an antibody cocktail of the Live/Dead stain, APC-Cy™7 anti-mouse CD45 (clone 30-F11, BD Biosciences, USA), APC anti-mouse Ly-6G (clone 1A8, BD Biosciences, USA), PE anti-mouse F4/80 (clone T45-2342, BD Biosciences, USA), FITC anti-mouse CD86 (clone GL-1, BioLegend, San Diego, CA, USA) each at a 1:100 dilution for 30 min on ice. For intracellular staining, the cells were fixed with Leucoperm™ Reagent A 10 min, washed and following the cells were permeabilized with Leucoperm™ reagent B (Bio-Rad, Hercules, CA, USA) containing BV421™ anti-mouse CD206 (clone C068C2, BioLegend, USA) at a 1:100 dilution. The cells were washed and then analyzed on the BD FACSVerse™ Flow Cytometer (BD Biosciences, USA). Data analysis was performed using FlowJo 7.6 software (FlowJo, Ashland, OR, USA) and normalized to tissue weight.
5.10. Histopathology
Skeletal muscle tissues (quadriceps, rectus abdominis and diaphragm) were fixed in 10% neutral buffered formalin and processed for histopathology. One cross- and/or longitudinal-section of each the muscle samples were embedded in Paraplast® (Sigma-Aldrich, USA). Paraffin sections, approximately 4 µm thick were prepared and stained with hematoxylin and eosin (H&E) for identification of muscle degeneration/regeneration and fiber necrosis. Sirius Red/Fast Green (SRFG) stain was applied for the detection of collagen deposition. Per mouse, three different muscle samples and two stains per sample were evaluated using the same localization, in total 6 slides/per mouse. The slides were analyzed in an unblinded fashion. Histopathological lesions were graded by a certified veterinary pathologist semi-quantitatively. The overall grading of a certain lesion is based on the evaluation of the entire area of the standardized muscle sample. Thus, the grading takes into account the severity of a single lesions as well as the multiplicity and distribution pattern of this change (i.e., number of affected fibers, affected area, amount of collagen deposition). Two major terms describe a certain muscle lesion: degeneration/necrosis (H&E) and fibrosis (SR_FG). The grading system applied is part of the Pathdata™system which is used for the collection and storage of microscopic data from toxicological and pharmacological studies. For the semiquantitative grading of a certain microscopic change the following scores were applied: GRADE 0 = no finding present; GRADE 1 = minimal/very few/very small; GRADE 2 = slight/few/small; and GRADE 3 = moderate/moderate number/moderate size.
5.11. Measurement of cNMPs in Skeletal Muscle Tissues
sGC stimulators are known to increase cGMP levels by binding to the reduced, heme-containing sGC enzyme. To assess whether BAY-747 increased 3′,5′-cyclic guanosine monophosphate (cGMP) levels and had any impact on 3′,5′-cyclic adenosine monophosphate (cAMP) levels, we measured the cyclic nucleotides levels in snap frozen skeletal muscle tissues (quadriceps, rectus abdominis and diaphragm) and heart using liquid chromatography coupled with tandem mass spectrometry (HPLC-MS/MS) using the UFLC HPLC system (Shimadzu, Kyoto, Japan) and the QTRAP5500™ triple quadrupole mass spectrometer (ABSciex, Framingham, MA, USA). A detailed description of the materials and methods used were previously reported [
35,
36].
5.12. Statistics
Running wheel data analysis: To summarize the overall distance and average speed data per block across time, the area under the curve (AUC) is calculated for each animal. AUC is log transformed to better fulfil the assumption of normality for the subsequent Dunnett’s test, comparing mdx/mTRG2 mice on BAY-747 with untreated wild type and mdx/mTRG2 mice, respectively. The family-wise error rate is set to 0.05. All results are transformed back to their original scale. The difference between group averages from the log-scaled data then corresponds to ratios on the original scale. The analysis was carried out using SAS 9.4 and R version 4.0.2.
All other data analysis: Data are expressed as mean ± standard error of mean (SEM). All analyses were perform using GraphPad Prism software v8 (GraphPad Software, San Diego, CA, USA). To identify outliers a ROUT test with Q = 1% was used. To identify statistical differences between groups, a one-way ANOVA was first performed, and if significant this was followed by a post-hoc Dunnett’s multiple comparisons test to identify which groups were different from untreated mdx/mTRG2 mice. Similarly, when measurements were repeated over time in the same mouse, a two-way repeated measures ANOVA was performed, followed by a Dunnett’s multiple comparison test. p < 0.05 was considered significant.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







