
Platelet-Derived Extracellular Vesicles for Regenerative Medicine

 ,
,  and
and
Abstract
:1. Introduction
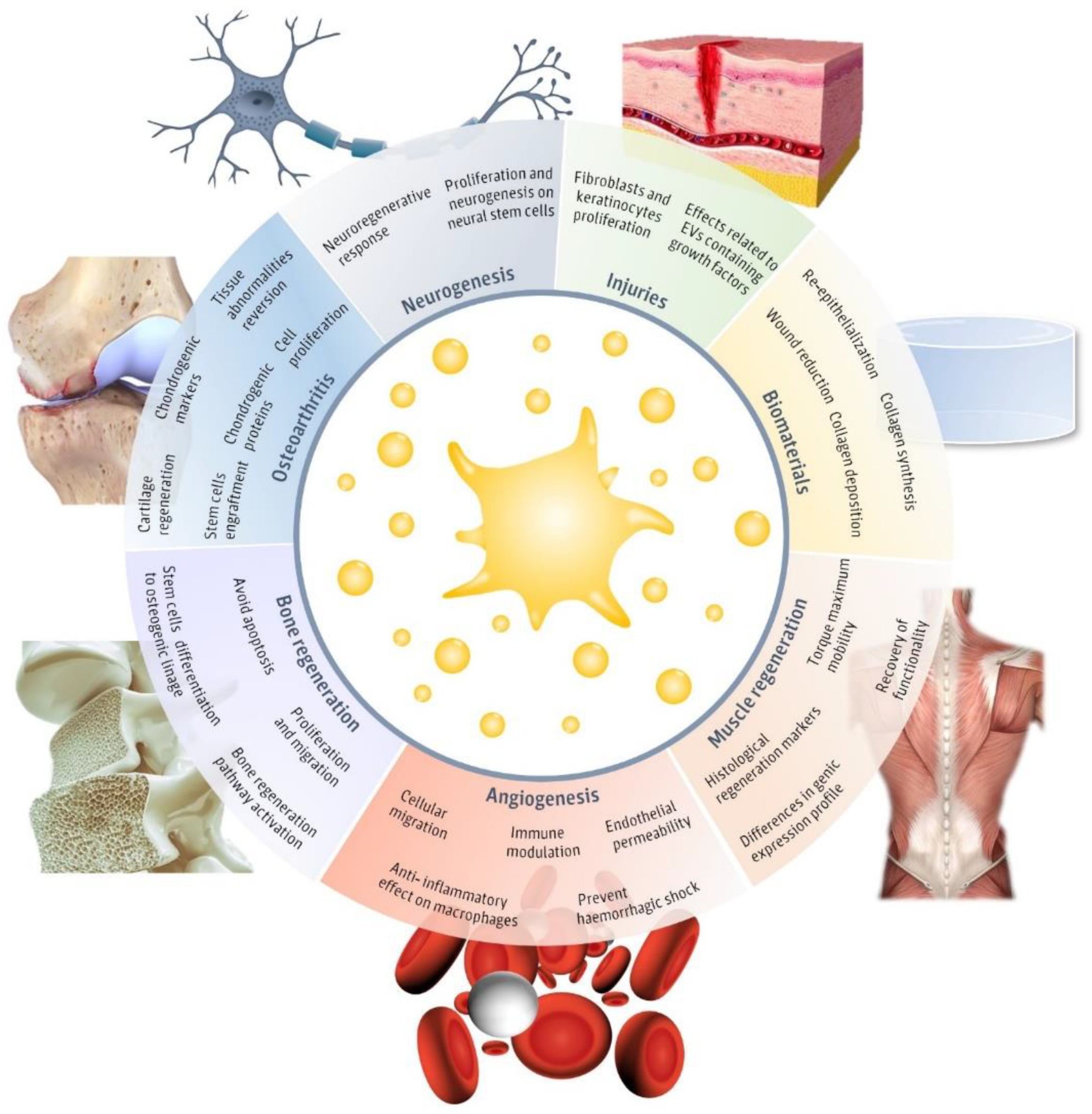
2. Regenerative Effects of pEVs
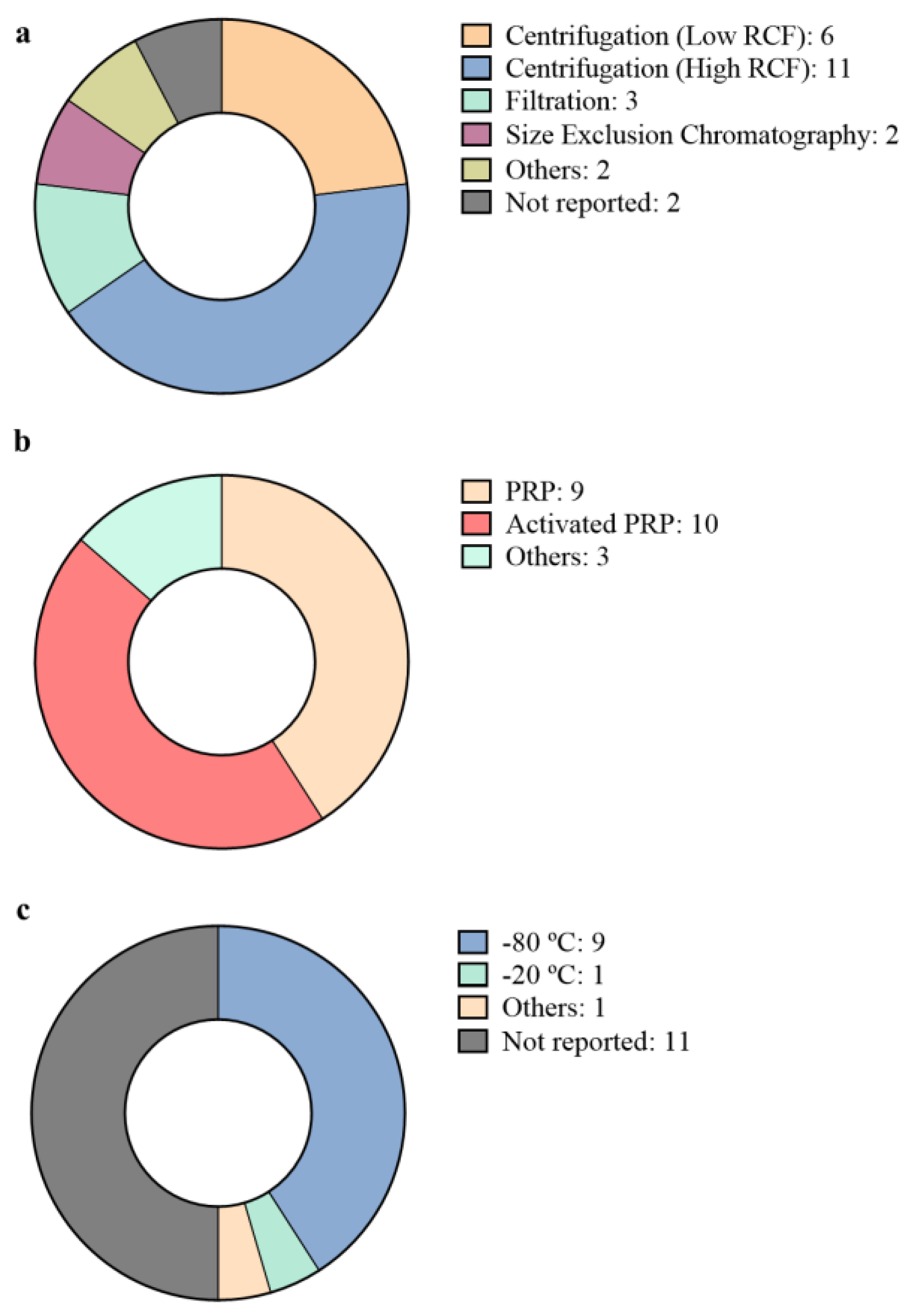
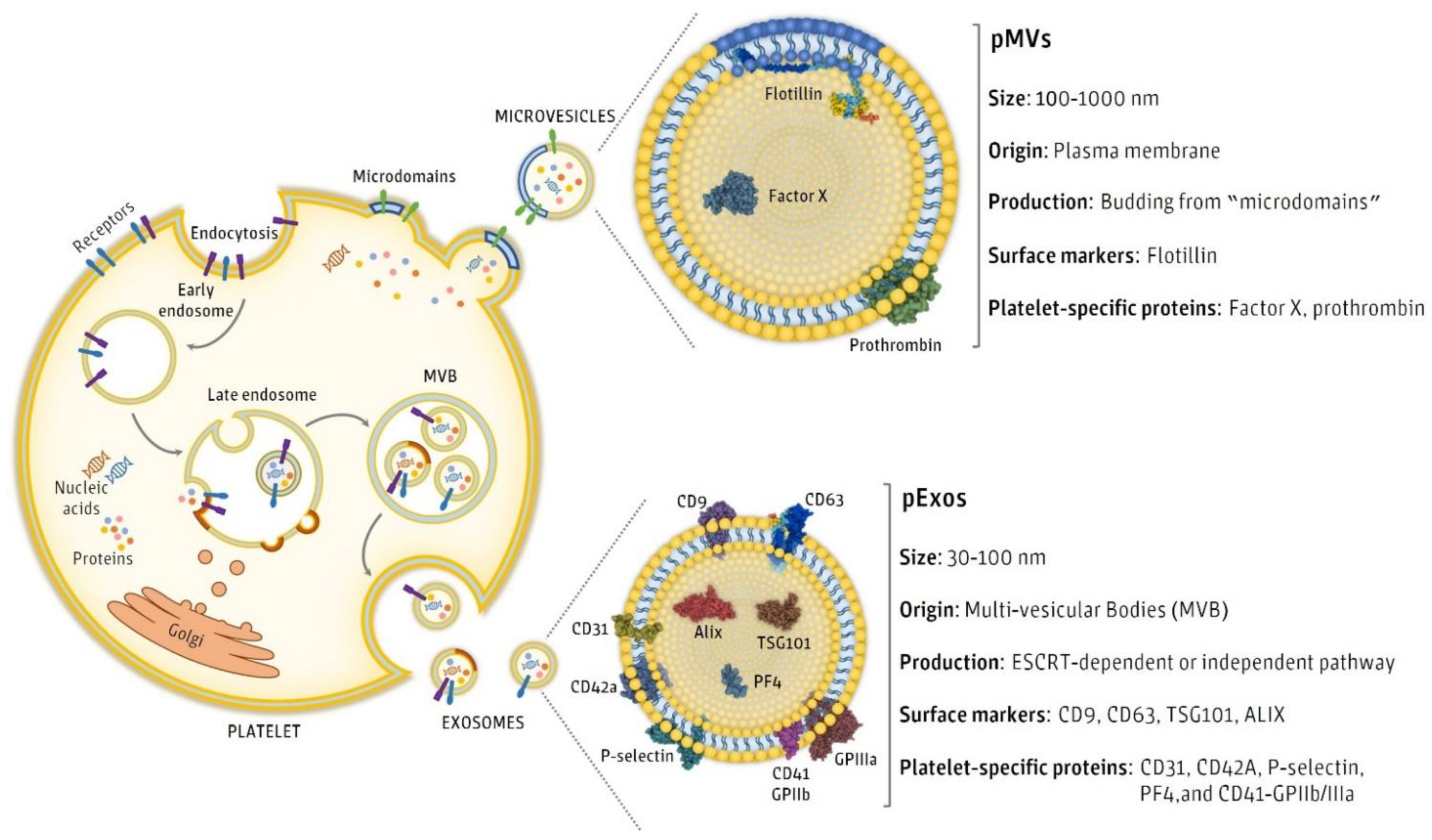
3. Isolation and Characterization of pEVs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Wiklander, O.P.B.; Brennan, M.Á.; Lötvall, J.; Breakefield, X.O.; El Andaloussi, S. Advances in therapeutic applications of extracellular vesicles. Sci. Transl. Med. 2019, 11, eaav8521. [Google Scholar] [CrossRef]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular vesicles for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156. [Google Scholar] [CrossRef]
- Rani, S.; Ryan, A.E.; Griffin, M.D.; Ritter, T. Mesenchymal Stem Cell-derived Extracellular Vesicles: Toward Cell-free Therapeutic Applications. Mol. Ther. 2015, 23, 812–823. [Google Scholar] [CrossRef] [Green Version]
- Veerman, R.E.; Akpinar, G.G.; Eldh, M.; Gabrielsson, S. Immune Cell-Derived Extracellular Vesicles—Functions and Therapeutic Applications. Trends Mol. Med. 2019, 25, 382–394. [Google Scholar] [CrossRef]
- Johnson, J.; Wu, Y.-W.; Blyth, C.; Lichtfuss, G.; Goubran, H.; Burnouf, T. Prospective Therapeutic Applications of Platelet Extracellular Vesicles. Trends Biotechnol. 2021, 39, 598–612. [Google Scholar] [CrossRef]
- Everts, P.; Onishi, K.; Jayaram, P.; Lana, J.; Mautner, K. Platelet-Rich Plasma: New Performance Understandings and Therapeutic Considerations in 2020. Int. J. Mol. Sci. 2020, 21, 7794. [Google Scholar] [CrossRef] [PubMed]
- Samadi, P.; Sheykhhasan, M.; Khoshinani, H.M. The Use of Platelet-Rich Plasma in Aesthetic and Regenerative Medicine: A Comprehensive Review. Aesthetic Plast. Surg. 2019, 43, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.I.-K.; Diaz, R.; Borg-Stein, J. Platelet-Rich Plasma. Phys. Med. Rehabil. Clin. N. Am. 2016, 27, 825–853. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, R.S.; Schwarz, E.M.; Maloney, M.D. Platelet-rich plasma therapy—Future or trend? Arthritis Res. Ther. 2012, 14, 219. [Google Scholar] [CrossRef] [Green Version]
- Etulain, J. Platelets in wound healing and regenerative medicine. Platelets 2018, 29, 556–568. [Google Scholar] [CrossRef]
- Tao, S.-C.; Guo, S.-C.; Zhang, C.-Q. Platelet-derived Extracellular Vesicles: An Emerging Therapeutic Approach. Int. J. Biol. Sci. 2017, 13, 828–834. [Google Scholar] [CrossRef] [Green Version]
- Puhm, F.; Boilard, E.; Machlus, K.R. Platelet Extracellular Vesicles. Arter. Thromb. Vasc. Biol. 2020, 2020, 87–96. [Google Scholar] [CrossRef]
- Wolf, P. The Nature and Significance of Platelet Products in Human Plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef]
- Crawford, N. The Presence of Contractile Proteins in Platelet Microparticles Isolated from Human and Animal Platelet-free Plasma. Br. J. Haematol. 1971, 21, 53–69. [Google Scholar] [CrossRef]
- Fadadu, P.P.; Mazzola, A.J.; Hunter, C.W.; Davis, T.T. Review of concentration yields in commercially available platelet-rich plasma (PRP) systems: A call for PRP standardization. Reg. Anesthesia Pain Med. 2019, 44, 652–659. [Google Scholar] [CrossRef]
- Fioravanti, C.; Frustaci, I.; Armellin, E.; Condò, R.; Arcuri, C.; Cerroni, L. Autologous blood preparations rich in platelets, fibrin and growth factors. Oral Implant. 2016, 8, 96–113. [Google Scholar] [CrossRef]
- Lippross, S.; Alini, M. Platelet-rich plasma for bone healing—To use or not to use? AO Dialogue 2007, 25–29. [Google Scholar]
- Marques, L.F.; Stessuk, T.; Camargo, I.C.C.; Junior, N.S.; Dos Santos, L.; Ribeiro-Paes, J.T. Platelet-rich plasma (PRP): Methodological aspects and clinical applications. Platelets 2014, 26, 101–113. [Google Scholar] [CrossRef]
- Harmon, K.; Hanson, R.; Bowen, J.; Greenberg, S.; Magaziner, E.; Vandenbosch, J.; Harshfield, D.; Shiple, B.; Audley, D. Guidelines for the use of Platelet Rich Plasma—Draft. Int. Cell Med. Soc. 2010, 1–11. Available online: http://www.cellmedicinesociety.org/attachments/206_ICMS%20-%20Guidelines%20for%20the%20use%20of%20Platelet%20Rich%20Plasma%20-%20Draft.pdf (accessed on 31 December 2011).
- Antich-Rosselló, M.; Forteza-Genestra, M.A.; Calvo, J.; Gayà, A.; Monjo, M.; Ramis, J.M. Platelet-derived extracellular vesicles promote osteoinduction of mesenchymal stromal cells. Bone Jt. Res. 2020, 9, 667–674. [Google Scholar] [CrossRef]
- Torreggiani, E.; Perut, F.; Roncuzzi, L.; Zini, N.; Baglio, S.R.; Baldini, N. Exosomes: Novel effectors of human platelet lysate activity. Eur. Cells Mater. 2014, 28, 137–151. [Google Scholar] [CrossRef]
- Sinauridze, E.I.; Kireev, D.A.; Popenko, N.Y.; Pichugin, A.V.; Panteleev, M.A.; Krymskaya, O.V.; Ataullakhanov, F.I. Platelet microparticle membranes have 50- to 100-fold higher specific procoagulant activity than activated platelets. Thromb. Haemost. 2007, 97, 425–434. [Google Scholar] [CrossRef]
- Melki, I.; Tessandier, N.; Zufferey, A.; Boilard, E. Platelet microvesicles in health and disease. Platelets 2017, 28, 214–221. [Google Scholar] [CrossRef]
- Kerris, E.W.J.; Hoptay, C.; Calderon, T.; Freishtat, R. Platelets and platelet extracellular vesicles in hemostasis and sepsis. J. Investig. Med. 2019, 68, 813–820. [Google Scholar] [CrossRef]
- Guo, S.-C.; Tao, S.-C.; Yin, W.-J.; Qi, X.; Yuan, T.; Zhang, C.-Q. Exosomes derived from platelet-rich plasma promote the re-epithelization of chronic cutaneous wounds via activation of YAP in a diabetic rat model. Theranostics 2017, 7, 81–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, N.; Wang, L.; Guan, J.; Tang, C.; He, N.; Zhang, W.; Fu, S. Wound healing effects of a Curcuma zedoaria polysaccharide with platelet-rich plasma exosomes assembled on chitosan/silk hydrogel sponge in a diabetic rat model. Int. J. Biol. Macromol. 2018, 117, 102–107. [Google Scholar] [CrossRef]
- Lovisolo, F.; Carton, F.; Gino, S.; Migliario, M.; Renò, F. Platelet rich plasma-derived microvesicles increased in vitro wound healing. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 9658–9664. [Google Scholar] [PubMed]
- Lopez, E.; Srivastava, A.; Burchfield, J.; Wang, Y.-W.; Cardenas, J.C.; Togarrati, P.P.; Miyazawa, B.; Gonzalez, E.; Holcomb, J.B.; Pati, S.; et al. Platelet-derived-Extracellular Vesicles Promote Hemostasis and Prevent the Development of Hemorrhagic Shock. Sci. Rep. 2019, 9, 17676. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, B.; Trivedi, A.; Togarrati, P.P.; Potter, D.; Baimukanova, G.; Vivona, L.; Lin, M.; Lopez, E.; Callcut, R.; Srivastava, A.; et al. Regulation of endothelial cell permeability by platelet-derived extracellular vesicles. J. Trauma Acute Care Surg. 2019, 86, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jung, H.; Song, J.; Choi, E.S.; You, G.; Mok, H. Activated Platelet-Derived Vesicles for Efficient Hemostatic Activity. Macromol. Biosci. 2020, 20, e1900338. [Google Scholar] [CrossRef]
- Sadallah, S.; Eken, C.; Martin, P.; Schifferli, J.A. Microparticles (Ectosomes) Shed by Stored Human Platelets Downregulate Macrophages and Modify the Development of Dendritic Cells. J. Immunol. 2011, 186, 6543–6552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayon, Y.; Dashevsky, O.; Shai, E.; Varon, D.; Leker, R.R. Platelet Microparticles Promote Neural Stem Cell Proliferation, Survival and Differentiation. J. Mol. Neurosci. 2012, 47, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Hayon, Y.; Dashevsky, O.; Shai, E.; Brill, A.; Varon, D.; Leker, R.R. Platelet Microparticles Induce Angiogenesis and Neurogenesis after Cerebral Ischemia. Curr. Neurovasc. Res. 2012, 9, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.R.; Scheiber, A.L.; Yarowsky, P.; Henn, I.R.F.; Otsuru, S.; Lovering, R.M. Exosomes Isolated From Platelet-Rich Plasma and Mesenchymal Stem Cells Promote Recovery of Function After Muscle Injury. Am. J. Sports Med. 2020, 48, 2277–2286. [Google Scholar] [CrossRef]
- Mause, S.F.; Ritzel, E.; Liehn, E.A.; Hristov, M.; Bidzhekov, K.; Müller-Newen, G.; Soehnlein, O.; Weber, C. Platelet Microparticles Enhance the Vasoregenerative Potential of Angiogenic Early Outgrowth Cells After Vascular Injury. Circulation 2010, 122, 495–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.K.; Song, K.S.; Chung, J.-H.; Lee, K.R.; Lee, S.-N. Platelet microparticles induce angiogenesisin vitro. Br. J. Haematol. 2004, 124, 376–384. [Google Scholar] [CrossRef]
- Brill, A.; Dashevsky, O.; Rivo, J.; Gozal, Y.; Varon, D. Platelet-derived microparticles induce angiogenesis and stimulate post-ischemic revascularization. Cardiovasc. Res. 2005, 67, 30–38. [Google Scholar] [CrossRef]
- Ferreira, M.R.; Zambuzzi, W.F. Platelet microparticles load a repertory of miRNAs programmed to drive osteogenic phenotype. J. Biomed. Mater. Res. Part A 2021, 109, 1502–1511. [Google Scholar] [CrossRef]
- Tao, S.-C.; Yuan, T.; Rui, B.-Y.; Zhu, Z.-Z.; Guo, S.-C.; Zhang, C.-Q. Exosomes derived from human platelet-rich plasma prevent apoptosis induced by glucocorticoid-associated endoplasmic reticulum stress in rat osteonecrosis of the femoral head via the Akt/Bad/Bcl-2 signal pathway. Theranostics 2017, 7, 733–750. [Google Scholar] [CrossRef]
- Moest, T.; Koehler, F.; Prechtl, C.; Schmitt, C.; Watzek, G.; Schlegel, K.A. Bone formation in peri-implant defects grafted with microparticles: A pilot animal experimental study. J. Clin. Periodontol. 2014, 41, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Otahal, A.; Kuten-Pella, O.; Kramer, K.; Neubauer, M.; Lacza, Z.; Nehrer, S.; De Luna, A. Functional repertoire of EV-associated miRNA profiles after lipoprotein depletion via ultracentrifugation and size exclusion chromatography from autologous blood products. Sci. Rep. 2021, 11, 5823. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, L.; Ma, C.; Wang, G.; Zhang, Y.; Sun, S. Exosomes derived from platelet-rich plasma present a novel potential in alleviating knee osteoarthritis by promoting proliferation and inhibiting apoptosis of chondrocyte via Wnt/β-catenin signaling pathway. J. Orthop. Surg. Res. 2019, 14, 470. [Google Scholar] [CrossRef] [Green Version]
- Otahal, A.; Kramer, K.; Kuten-Pella, O.; Weiss, R.; Stotter, C.; Lacza, Z.; Weber, V.; Nehrer, S.; De Luna, A. Characterization and Chondroprotective Effects of Extracellular Vesicles from Plasma- and Serum-Based Autologous Blood-Derived Products for Osteoarthritis Therapy. Front. Bioeng. Biotechnol. 2020, 8, 584050. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Huang, J.; Luo, P.; Wang, Z.; He, J.; Wu, S.; Peng, C.; Cao, X. Platelet-Derived Microparticles Mediate the Intra-Articular Homing of Mesenchymal Stem Cells in Early-Stage Cartilage Lesions. Stem Cells Dev. 2020, 29, 414–424. [Google Scholar] [CrossRef]
- Hess, J.R.; Lelkens, C.C.; Holcomb, J.B.; Scalea, T.M. Advances in military, field, and austere transfusion medicine in the last decade. Transfus. Apher. Sci. 2013, 49, 380–386. [Google Scholar] [CrossRef]
- Lener, T.; Gimona, M.; Aigner, L.; Börger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; Del Portillo, H.A.; et al. Applying extracellular vesicles based therapeutics in clinical trials—an ISEV position paper. J. Extracell. Vesicles 2015, 4, 30087. [Google Scholar] [CrossRef]
- Milioli, M.; Ibáñez-Vea, M.; Sidoli, S.; Palmisano, G.; Careri, M.; Larsen, M.R. Quantitative proteomics analysis of platelet-derived microparticles reveals distinct protein signatures when stimulated by different physiological agonists. J. Proteom. 2015, 121, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, K.B.; Gudbergsson, J.M.; Andresen, T.L.; Simonsen, J.B. What is the blood concentration of extracellular vesicles? Implications for the use of extracellular vesicles as blood-borne biomarkers of cancer. Biochim. Biophys. Acta (BBA) Bioenergy 2019, 1871, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Greening, D.; Zhu, H.-J.; Takahashi, N.; Simpson, R.J. Extracellular vesicle isolation and characterization: Toward clinical application. J. Clin. Investig. 2016, 126, 1152–1162. [Google Scholar] [CrossRef] [Green Version]
- Gandham, S.; Su, X.; Wood, J.; Nocera, A.L.; Alli, S.C.; Milane, L.; Zimmerman, A.; Amiji, M.; Ivanov, A.R. Technologies and Standardization in Research on Extracellular Vesicles. Trends Biotechnol. 2020, 38, 1066–1098. [Google Scholar] [CrossRef]
- Sidhom, K.; Obi, P.; Saleem, A. A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, Y.; Ishikawa, Y. Remnant lipoproteins as strong key particles to atherogenesis. J. Atheroscler. Thromb. 2009, 16, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Antwi-Baffour, S.; Adjei, J.; Aryeh, C.; Kyeremeh, R.; Kyei, F.; Seidu, M.A. Understanding the biosynthesis of platelets-derived extracellular vesicles. Immun. Inflamm. Dis. 2015, 3, 133–140. [Google Scholar] [CrossRef]
- Posma, J.J.N.; Posthuma, J.J.; Spronk, H.M.H. Coagulation and non-coagulation effects of thrombin. J. Thromb. Haemost. 2016, 14, 1908–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hougie, C. The Activation of Platelets by Plasma. Br. J. Haematol. 1955, 1, 213–222. [Google Scholar] [CrossRef]
- Tissot, J.-D.; Canellini, G.; Rubin, O.; Angelillo-Scherrer, A.; Delobel, J.; Prudent, M.; Lion, N. Blood microvesicles: From proteomics to physiology. Transl. Proteom. 2013, 1, 38–52. [Google Scholar] [CrossRef] [Green Version]
- Qin, B.; Zhang, Q.; Hu, X.; Mi, T.; Yu, H.; Liu, S.; Zhang, B.; Tang, M.; Huang, J.; Xiong, K. How does temperature play a role in the storage of extracellular vesicles? J. Cell. Physiol. 2020, 235, 7663–7680. [Google Scholar] [CrossRef] [PubMed]
- Maroto, R.; Zhao, Y.; Jamaluddin, M.; Popov, V.L.; Wang, H.; Kalubowilage, M.; Zhang, Y.; Luisi, J.; Sun, H.; Culbertson, C.T.; et al. Effects of storage temperature on airway exosome integrity for diagnostic and functional analyses. J. Extracell. Vesicles 2017, 6, 1359478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lőrincz, Á.; Timar, C.; Marosvári, K.A.; Veres, D.S.; Otrokocsi, L.; Kittel, Á.; Ligeti, E. Effect of storage on physical and functional properties of extracellular vesicles derived from neutrophilic granulocytes. J. Extracell. Vesicles 2014, 3, 25465. [Google Scholar] [CrossRef]
- Royo, F.; Théry, C.; Falcón-Pérez, J.M.; Nieuwland, R.; Witwer, K.W. Methods for Separation and Characterization of Extracellular Vesicles: Results of a Worldwide Survey Performed by the ISEV Rigor and Standardization Subcommittee. Cells 2020, 9, 1955. [Google Scholar] [CrossRef] [PubMed]
- De Luna, A.; Otahal, A.; Nehrer, S. Mesenchymal Stromal Cell-Derived Extracellular Vesicles—Silver Linings for Cartilage Regeneration? Front. Cell Dev. Biol. 2020, 8, 1548. [Google Scholar] [CrossRef] [PubMed]
- French, S.L.; Butov, K.R.; Allaeys, I.; Canas, J.; Morad, G.; Davenport, P.; Laroche, A.; Trubina, N.M.; Italiano, J.E.; Moses, M.A.; et al. Platelet-derived extracellular vesicles infiltrate and modify the bone marrow during inflammation. Blood Adv. 2020, 4, 3011–3023. [Google Scholar] [CrossRef]
- Penolazzi, L.; Vecchiatini, R.; Bignardi, S.; Lambertini, E.; Torreggiani, E.; Canella, A.; Franceschetti, T.; Calura, G.; Vesce, F.; Piva, R. Influence of obstetric factors on osteogenic potential of umbilical cord-derived mesenchymal stem cells. Reprod. Biol. Endocrinol. 2009, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Soleymani, S.; Yari, F.; Bolhassani, A.; Bakhshandeh, H. Platelet microparticles: An effective delivery system for anti-viral drugs. J. Drug Deliv. Sci. Technol. 2019, 51, 290–296. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regenerative Medicine Field | Platelet Source | Isolation Method | pEVs Storage Conditions | Characterization | Study Model | Reference |
|---|---|---|---|---|---|---|
| Injuries and wounds Biomaterials Angiogenesis | PRP | High RCF centrifugation | Frozen at −80 °C | Physical characterization and pEV marker detection | In vitro cell culture In vivo diabetic rat model | [30] |
| PRP | Not specified | Not specified | Not specified | In vivo diabetic rat model | [31] | |
| Injuries and wounds | Activated PRP | Low RCF centrifugation | Not specified | Physical characterization | In vitro cell culture | [32] |
| PRP | Filtration | Frozen at −20 °C | Physical characterization and pEV marker detection | In vitro blood samples In vivo bleeding rat model | [33] | |
| Activated PRP | High RCF centrifugation | Frozen at −80 °C | Physical characterization and pEV marker detection | In vitro cell culture In vitro blood samples In vivo mice model | [34] | |
| 3 days stored activated platelets | Sonication | Not specified | Physical characterization | In vitro cell culture In vivo mice model. | [35] | |
| 5 days stored PRP | High RCF centrifugation | Stored at −80 °C until final centrifugation. | Physical characterization and pEV marker detection | In vitro cell culture | [36] | |
| Angiogenesis | Activated platelets | Low RCF centrifugation | Not specified | Not specified | In vitro cell culture | [40] |
| Activated PRP | Low RCF centrifugation | Not specified | Not specified | In vitro cell culture | [41] | |
| Activated PRP | High RCF centrifugation | Not specified | Physical characterization and pEV marker detection | In vitro cell culture In vivo ischemic heart rat model | [42] | |
| Angiogenesis Neural regeneration | Activated PRP | High RCF centrifugation | Not specified | Physical characterization and pEV marker detection | In vitro cell culture | [37] |
| Activated PRP | High RCF centrifugation | Not specified | pEV marker detection | In vivo focal ischemia rat model | [38] | |
| Osteoarthritis | PRP | High RCF centrifugation Filtration Size exclusion chromatography A combination of different techniques | Frozen at −80 °C | Physical characterization and pEV marker detection | miRNA profiling | [46] |
| PRP | Spin column based commercial kit | Frozen at −80 °C | Physical characterization and pEV marker detection | In vitro cell culture In vivo osteoarthritic rabbit model | [47] | |
| PRP | High RCF centrifugation | Frozen at −80 °C | Physical characterization and pEV marker detection | In vitro cell culture | [48] | |
| Activated PRP | Low RCF centrifugation | Not specified | Not specified | In vitro cell culture In vivo rat model | [49] | |
| Musculoskeletal regeneration | Not appliable | Not appliable | Not appliable | Not appliable | In silico miRNA profiling | [43] |
| PL | High RCF centrifugation Size Exclusion Chromatography | Frozen at −80 °C | Physical characterization and pEV marker detection | In vitro cell culture | [25] | |
| PL | High RCF centrifugation | Frozen at −80 °C | Physical characterization and pEV marker detection | In vitro cell culture | [26] | |
| PRP | High RCF centrifugation | Frozen at −80 °C | Physical characterization and pEV marker detection | In vitro cell culture In vivo rat model | [44] | |
| PRP | Sonication | Not specified | Not specified | In vivo pig model | [45] | |
| Activated PRP | High RCF centrifugation | Frozen at −80 °C | Physical characterization and pEV marker detection | In vivo rat model | [39] |
| Kind of Proteins Commonly Reported | pEV Markers | References |
|---|---|---|
| EV membrane markers | CD9 | [25,30,33,34,39,44,46,47,48] |
| CD61 | [33,36] | |
| CD63 | [25,26,30,33,34,39,44,47,48,66] | |
| D81 | [30,33,34,39,44,47] | |
| Platelet source markers | CD31 | [34] |
| CD41 | [33,34,37,38,42,44,48,67,68,69] | |
| CD42 | [40] | |
| EV cytosolic markers | ALIX | [46,48] |
| HSP90 | [33] | |
| HPS101 | [47] | |
| TSG101 | [44] | |
| Non-EVs structures | APOA1 | [46,48] |
| APOB100 | [46,48] | |
| Calnexin | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antich-Rosselló, M.; Forteza-Genestra, M.A.; Monjo, M.; Ramis, J.M. Platelet-Derived Extracellular Vesicles for Regenerative Medicine. Int. J. Mol. Sci. 2021, 22, 8580. https://doi.org/10.3390/ijms22168580
Antich-Rosselló M, Forteza-Genestra MA, Monjo M, Ramis JM. Platelet-Derived Extracellular Vesicles for Regenerative Medicine. International Journal of Molecular Sciences. 2021; 22(16):8580. https://doi.org/10.3390/ijms22168580
Chicago/Turabian StyleAntich-Rosselló, Miquel, Maria Antònia Forteza-Genestra, Marta Monjo, and Joana M. Ramis. 2021. "Platelet-Derived Extracellular Vesicles for Regenerative Medicine" International Journal of Molecular Sciences 22, no. 16: 8580. https://doi.org/10.3390/ijms22168580
APA StyleAntich-Rosselló, M., Forteza-Genestra, M. A., Monjo, M., & Ramis, J. M. (2021). Platelet-Derived Extracellular Vesicles for Regenerative Medicine. International Journal of Molecular Sciences, 22(16), 8580. https://doi.org/10.3390/ijms22168580