
A Time-Course Study of the Expression Level of Synaptic Plasticity-Associated Genes in Un-Lesioned Spinal Cord and Brain Areas in a Rat Model of Spinal Cord Injury: A Bioinformatic Approach

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
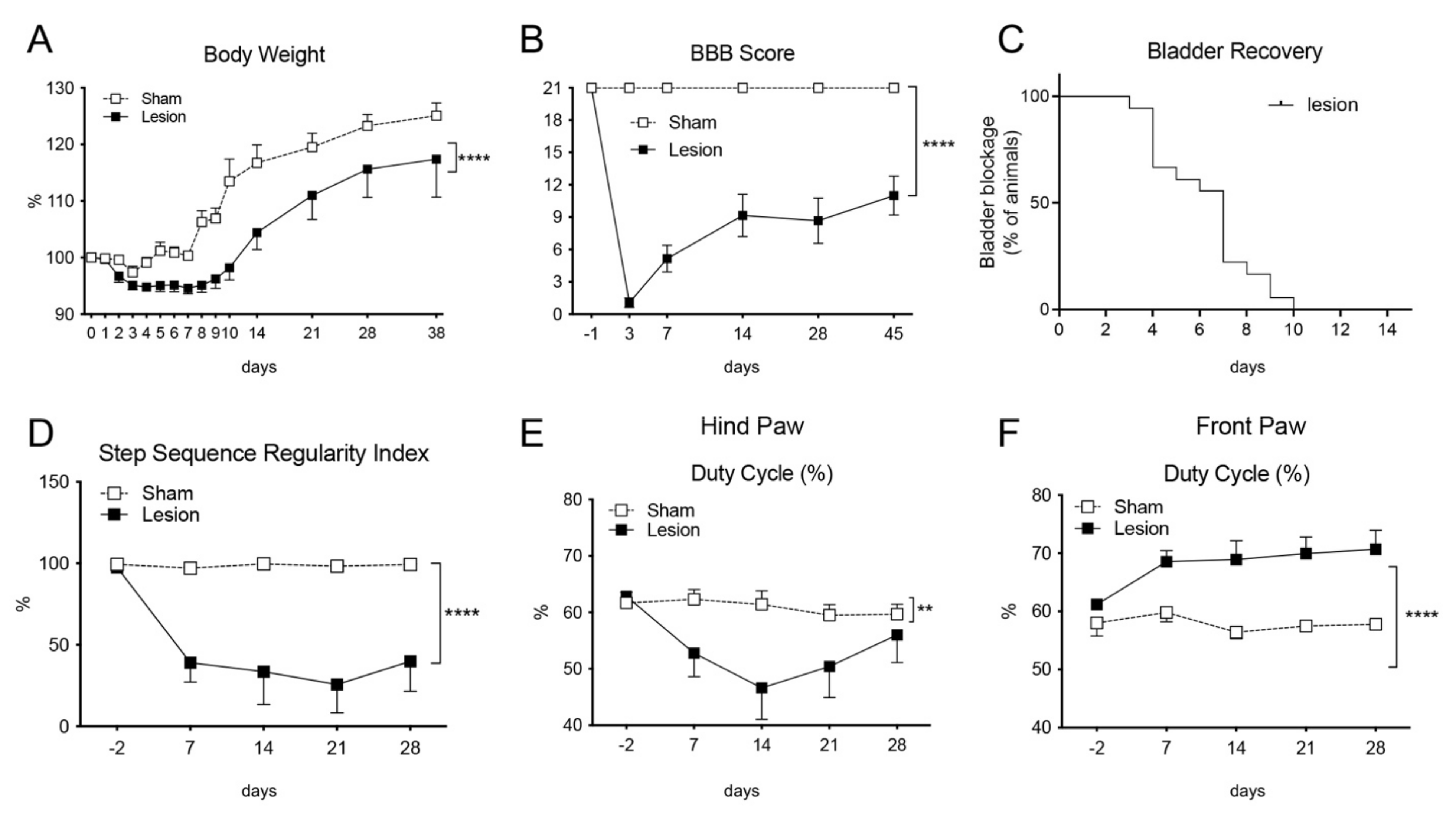
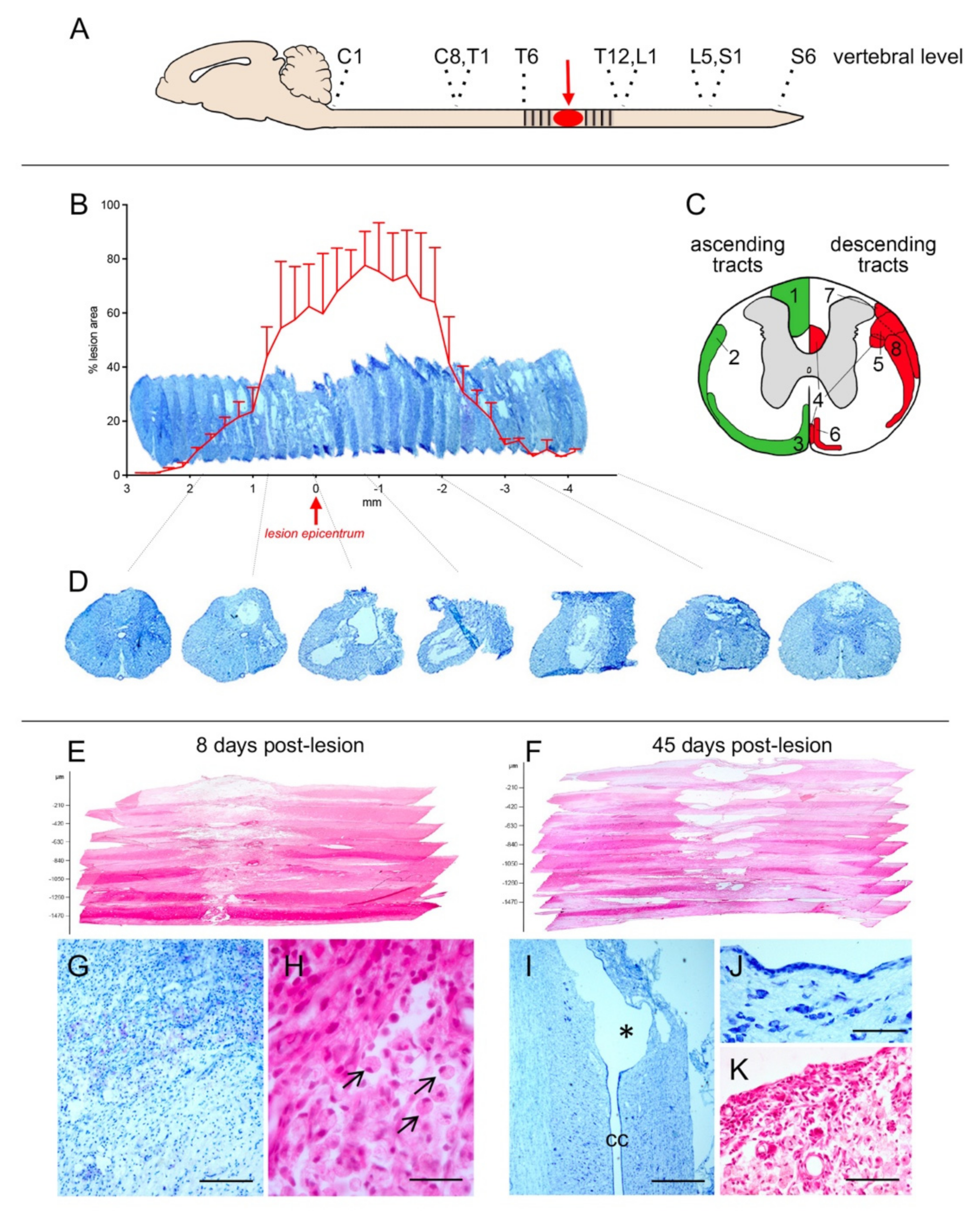
2.1. Lesion Characterization
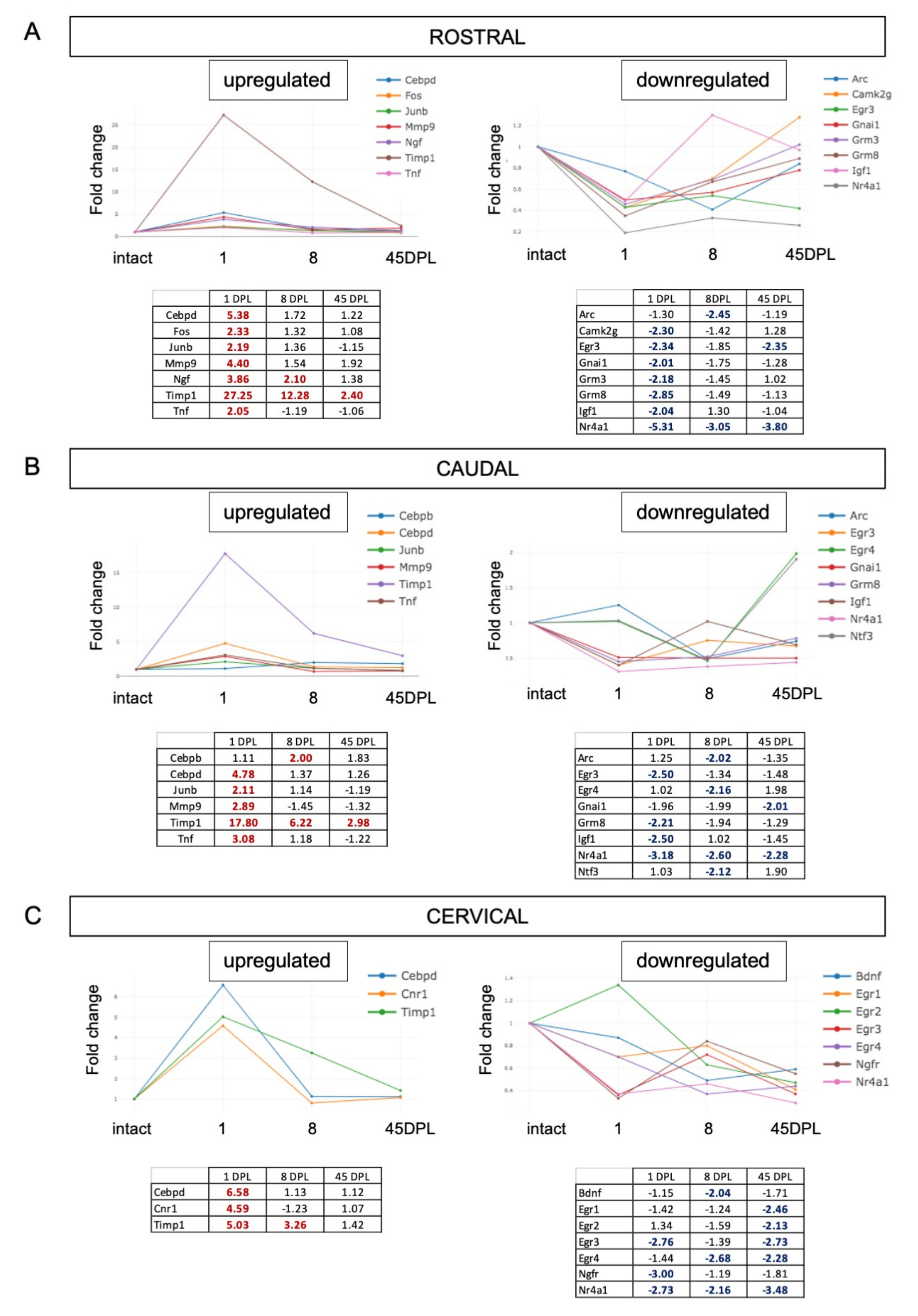
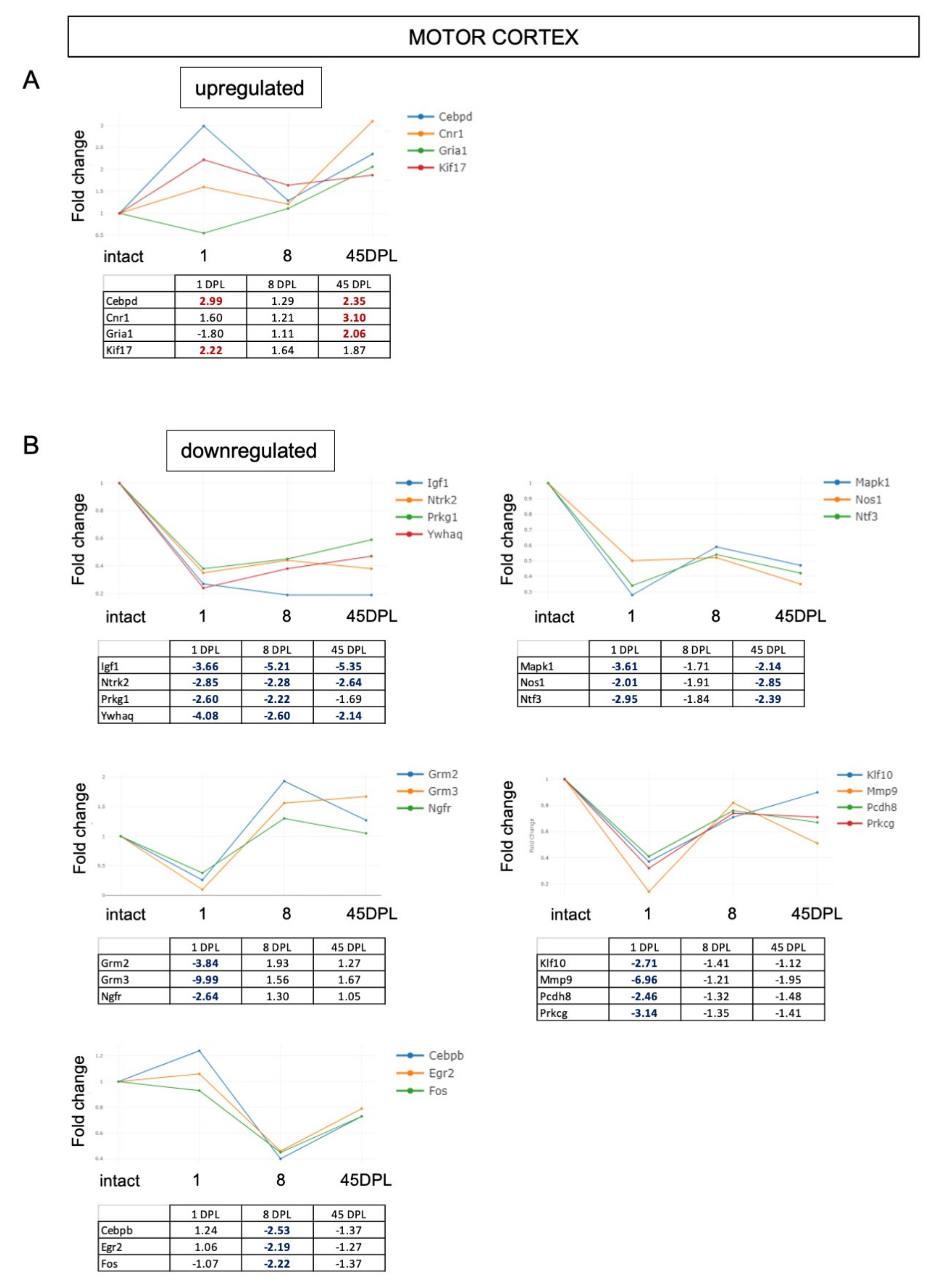
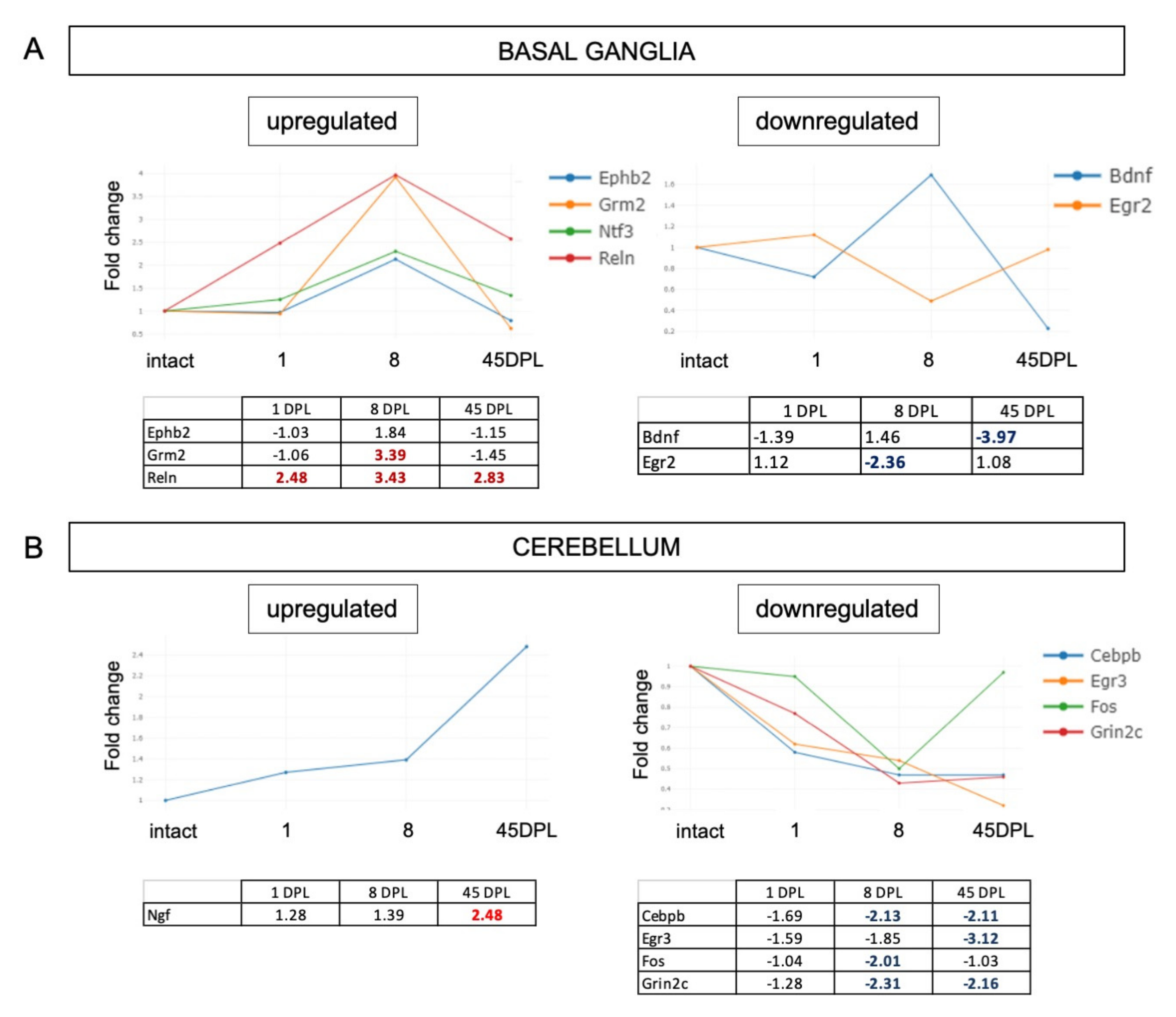
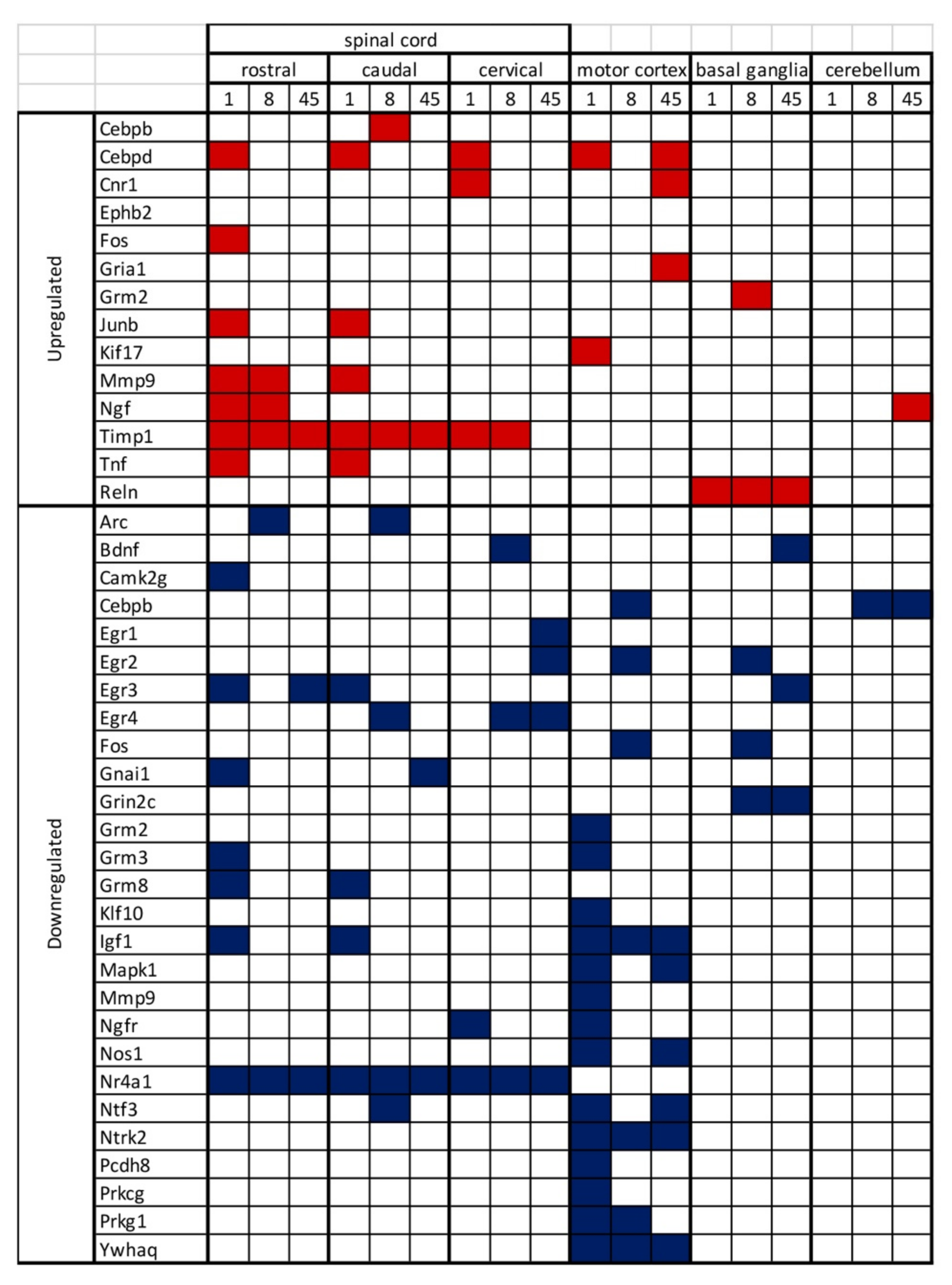
2.2. mRNA Expression Level of Synaptic Plasticity Associated Genes
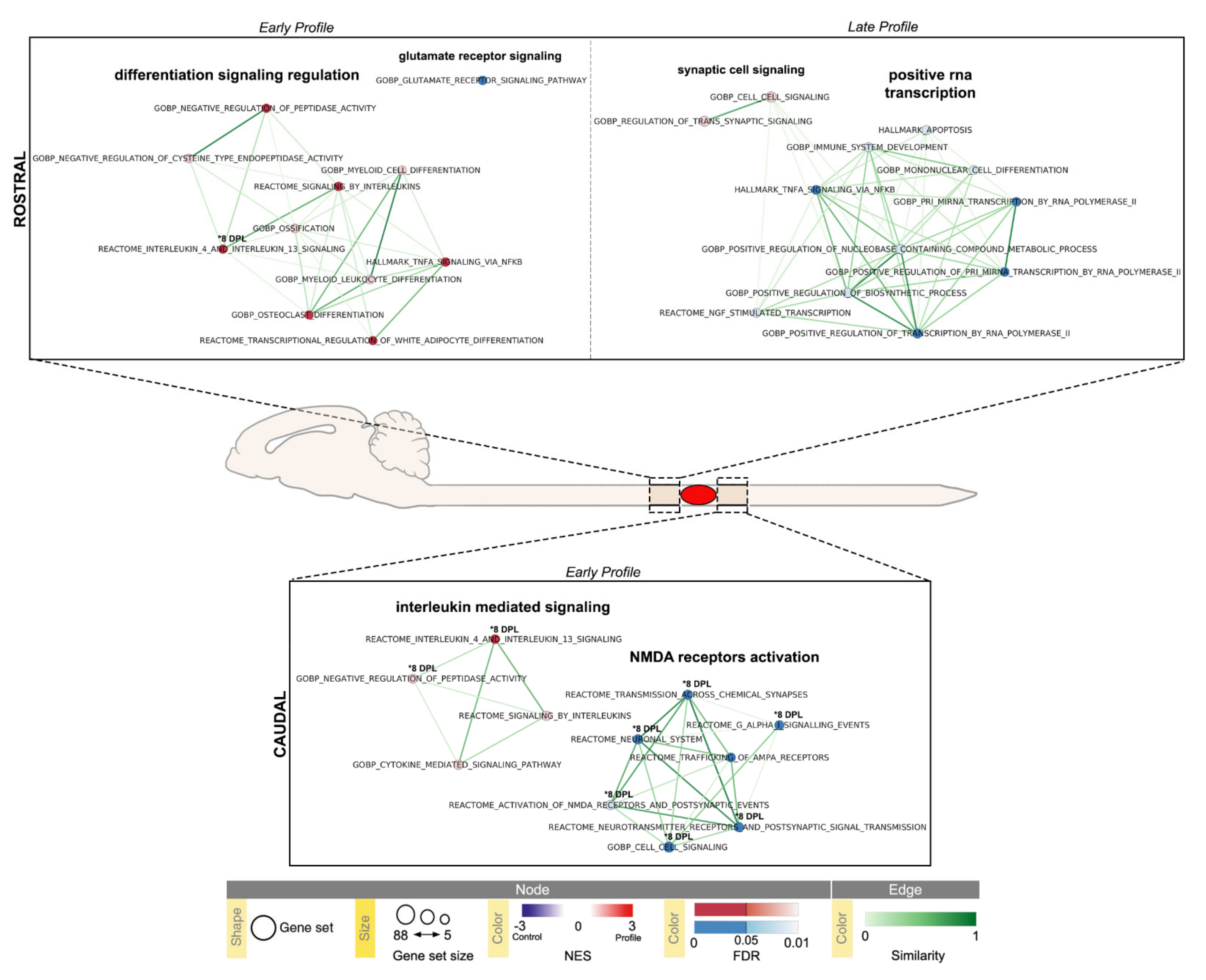
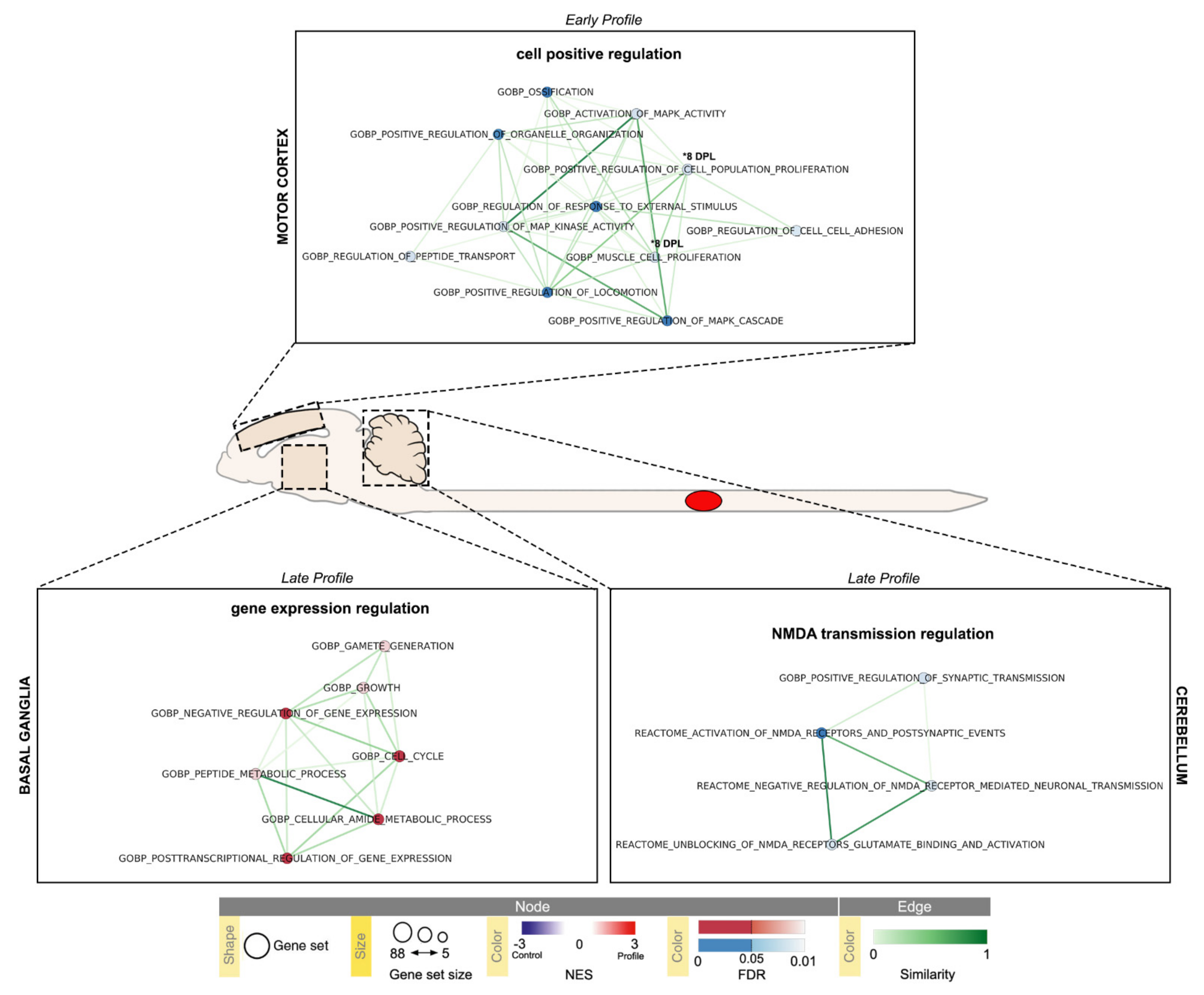
2.3. Pathway Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Animals, Surgery and Care
4.2. BBB Score, Locomotion and Gait Analysis
4.3. Histology
4.4. Total RNA Isolation, Reverse Transcription and PCR-Arrays
4.5. Functional Pathway and Gene Set Enrichment Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beauparlant, J.; Van Den Brand, R.; Barraud, Q.; Friedli, L.; Musienko, P.; Dietz, V.; Courtine, G. Undirected compensatory plasticity contributes to neuronal dysfunction after severe spinal cord injury. Brain 2013, 136, 3347–3361. [Google Scholar] [CrossRef] [Green Version]
- Dietz, V. Behavior of spinal neurons deprived of supraspinal input. Nat. Rev. Neurol. 2010, 6, 167–174. [Google Scholar] [CrossRef]
- Dietz, V.; Müller, R. Degradation of neuronal function following a spinal cord injury: Mechanics and countermeasures. Brain 2004, 127, 2221–2231. [Google Scholar] [CrossRef] [Green Version]
- Grau, J.W.; Baine, R.E.; Bean, P.A.; Davis, J.A.; Fauss, G.N.; Henwood, M.K.; Hudson, K.E.; Johnston, D.T.; Tarbet, M.M.; Strain, M.M. Learning to promote recovery after spinal cord injury. Exp. Neurol. 2020, 330, 113334. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.S.; Park, J.W.; Jin, S.U.; Jang, K.E.; Nam, H.U.; Lee, Y.S.; Jung, T.-D.; Chang, Y. Alteration of resting-state brain sensorimotor connectivity following spinal cord injury: A resting-state functional magnetic resonance imaging study. J. Neurotrauma 2015, 32, 1422–1427. [Google Scholar] [CrossRef] [PubMed]
- Hutson, T.H.; Di Giovanni, S. The translational landscape in spinal cord injury: Focus on neuroplasticity and regeneration. Nat. Rev. Neurol. 2019, 15, 732–745. [Google Scholar] [CrossRef] [PubMed]
- Krucoff, M.O.; Miller, J.P.; Saxena, T.; Bellamkonda, R.; Rahimpour, S.; Harward, S.C.; Lad, S.P.; Turner, D.A. Toward functional restoration of the central nervous system: A review of translational neuroscience principles. Clin. Neurosurg. 2019, 84, 30–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Espín, A.; Beaudry, E.; Fenrich, K.; Fouad, K. Rehabilitative training in animal models of spinal cord injury. J. Neurotrauma 2018, 35, 1970–1985. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Dong, X. Preparation and characterization of novel HDL-mimicking nanoparticles for nerve growth factor encapsulation. J. Vis. Exp. 2017, 2017, 55584. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.; He, X.; Wang, F.; Tan, J.M.; Wang, J.X.; Xing, S.M.; Shen, L.B.; Fang, L.Q.; Yang, P.; Tan, J.M. Gene expression signature analysis and protein-protein interaction network construction of spinal cord injury. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2941–2948. [Google Scholar]
- Ni, Y.; Zhang, K. Clustering analysis to identify key genes associated with motor neuron excitability following spinal cord injury. Int. J. Neurosci. 2019, 129, 856–863. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Song, J.; Wang, L.G.; Shao, J. Identification of key genes and pathways associated with spinal cord injury. Mol. Med. Rep. 2017, 15, 1577–1584. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Lv, Y.; Li, S.; Feng, T.; Zhou, Y.; Sun, Y.; Mi, D. Changes in transcriptome profiling during the acute/subacute phases of contusional spinal cord injury in rats. Ann. Transl. Med. 2020, 8, 1682. [Google Scholar] [CrossRef]
- Shin, H.Y.; Kim, H.; Kwon, M.J.; Hwang, D.H.; Lee, K.Y.; Kim, B.G. Molecular and cellular changes in the lumbar spinal cord following thoracic injury: Regulation by treadmill locomotor training. PLoS ONE 2014, 9, e88215. [Google Scholar] [CrossRef]
- Chen, X.J.; Mou, X.Q.; Zou, Y.; Peng, Z.Y.; Yang, J.X. Screening of key genes associated with contused rat spinal cord with DNA microarray. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2949–2955. [Google Scholar]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A sensitive and reliable locomotor rating scale for open field testing in rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chabas, D.; Baranzini, S.E.; Mitchell, D.; Bernard, C.C.A.; Rittling, S.R.; Denhardt, D.T.; Sobel, R.A.; Lock, C.; Karpuj, M.; Pedotti, R.; et al. The influence of the proinflammatory cytokine, osteopontin, on autoimmue demyelinating desease. Science 2001, 294, 1731–1735. [Google Scholar] [CrossRef] [PubMed]
- Kendziorski, C.M.; Zhang, Y.; Lan, H.; Attie, A.D. The efficiency of pooling mRNA in microarray experiments. Biostatistics 2003, 4, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Kendziorski, C.; Irizarry, R.A.; Chen, K.S.; Haag, J.D.; Gould, M.N. On the utility of pooling biological samples in microarray experiments. Proc. Natl. Acad. Sci. USA 2005, 102, 4252–4257. [Google Scholar] [CrossRef] [Green Version]
- Van Den Brand, R.; Heutschi, J.; Barraud, Q.; DiGiovanna, J.; Bartholdi, K.; Huerlimann, M.; Friedli, L.; Vollenweider, I.; Moraud, E.M.; Duis, S.; et al. Restoring voluntary control of locomotion after paralyzing spinal cord injury. Science 2012, 336, 1182–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadoya, K.; Lu, P.; Nguyen, K.; Lee-Kubli, C.; Kumamaru, H.; Yao, L.; Knackert, J.; Poplawski, G.; Dulin, J.N.; Strobl, H.; et al. Spinal cord reconstitution with homologous neural grafts enables robust corticospinal regeneration. Nat. Med. 2016, 22, 479–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyinbo, C.A. Secondary injury mechanisms in traumatic spinal cord injury: A nugget of this multiply cascade. Acta Neurobiol. Exp. 2011, 71, 281–299. [Google Scholar]
- Tran, A.P.; Warren, P.M.; Silver, J. The biology of regeneration failure and success after spinal cord injury. Physiol. Rev. 2018, 98, 881–917. [Google Scholar] [CrossRef]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic spinal cord injury: An overview of pathophysiology, models and acute injury mechanisms. Front. Neurol. 2019, 10, 282. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Myelin damage and repair in pathologic CNS: Challenges and prospects. Front. Mol. Neurosci. 2015, 8, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, P.K.; Song, J.; Kim, S.; Zhong, H.; Roy, R.R.; Edgerton, V.R. Rodent estrous cycle response to incomplete spinal cord injury, surgical interventions, and locomotor training. Behav. Neurosci. 2011, 125, 996–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjell, J.; Olson, L. Rat models of spinal cord injury: From pathology to potential therapies. DMM Dis. Model. Mech. 2016, 9, 1125–1137. [Google Scholar] [CrossRef] [Green Version]
- Watson, C.; Kayalioglu, G. The organization of the spinal cord. In the Spinal Cord; Elsevier Ltd.: Amsterdam, The Netherlands, 2009; pp. 1–7. ISBN 9780123742476. [Google Scholar]
- Bighinati, A.; Khalajzeyqami, Z.; Baldassarro, V.A.; Lorenzini, L.; Cescatti, M.; Moretti, M.; Giardino, L.; Calzà, L. Time-course changes of extracellular matrix encoding genes expression level in the spinal cord following contusion injury—A data-driven approach. Int. J. Mol. Sci. 2021, 22, 1744. [Google Scholar] [CrossRef]
- Sammons, R.P.; Keck, T. Adult plasticity and cortical reorganization after peripheral lesions. Curr. Opin. Neurobiol. 2015, 35, 136–141. [Google Scholar] [CrossRef]
- Jacobi, A.; Schmalz, A.; Bareyre, F.M. Abundant expression of guidance and synaptogenic molecules in the injured spinal cord. PLoS ONE 2014, 9, e88449. [Google Scholar] [CrossRef]
- Lim, A.F.Y.; Lim, W.L.; Ch′ng, T.H. Activity-dependent synapse to nucleus signaling. Neurobiol. Learn. Mem. 2017, 138, 78–84. [Google Scholar] [CrossRef]
- Poirier, R.; Cheval, H.; Mailhes, C.; Garel, S.; Charnay, P.; Davis, S.; Laroche, S. Distinct functions of egr gene family members in cognitive processes. Front. Neurosci. 2008, 2, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Zhong, L.; Wang, A.-Q.; Zhang, H.; Yin, Z.S. Identification of regeneration and hub genes and pathways at different time points after spinal cord injury. Mol. Neurobiol. 2021, 58, 2643–2662. [Google Scholar] [CrossRef] [PubMed]
- Azzarito, M.; Seif, M.; Kyathanahally, S.; Curt, A.; Freund, P. Tracking the neurodegenerative gradient after spinal cord injury. NeuroImage Clin. 2020, 26, 102221. [Google Scholar] [CrossRef]
- Yokota, K.; Kubota, K.; Kobayakawa, K.; Saito, T.; Hara, M.; Kijima, K.; Maeda, T.; Katoh, H.; Ohkawa, Y.; Nakashima, Y.; et al. Pathological changes of distal motor neurons after complete spinal cord injury. Mol. Brain 2019, 12, 4. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, Y.; Yamamoto, M.; Sugiura, Y.; Setoyama, D.; Kishima, H. Rostro-caudal different energy metabolism leading to differences in degeneration in spinal cord injury. Brain Commun. 2021, 3, fcab058. [Google Scholar] [CrossRef]
- David, G.; Mohammadi, S.; Martin, A.R.; Cohen-Adad, J.; Weiskopf, N.; Thompson, A.; Freund, P. Traumatic and nontraumatic spinal cord injury: Pathological insights from neuroimaging. Nat. Rev. Neurol. 2019, 15, 718–731. [Google Scholar] [CrossRef] [PubMed]
- Hassannejad, Z.; Shakouri-Motlagh, A.; Mokhatab, M.; Zadegan, S.A.; Sharif-Alhoseini, M.; Shokraneh, F.; Rahimi-Movaghar, V. Oligodendrogliogenesis and axon remyelination after traumatic spinal cord injuries in animal studies: A systematic review. Neuroscience 2019, 402, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Hilton, B.J.; Anenberg, E.; Harrison, T.C.; Boyd, J.D.; Murphy, T.H.; Tetzlaff, W. Re-establishment of cortical motor output maps and spontaneous functional recovery via spared dorsolaterally projecting corticospinal neurons after dorsal column spinal cord injury in adult mice. J. Neurosci. 2016, 36, 4080–4092. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Wu, W.; Xiong, W.; Chai, Z.; Xu, X.M.; Jin, X. Longitudinal optogenetic motor mapping revealed structural and functional impairments and enhanced corticorubral projection after contusive spinal cord injury in Mice. J. Neurotrauma 2019, 36, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Baek, A.; Cho, S.R.; Kim, S.H. Elucidation of gene expression patterns in the brain after spinal cord injury. Cell Transplant. 2017, 26, 1286–1300. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.W.; Wu, G.Y.; Min, L.X.; Feng, Z.; Chen, H.; Tan, M.L.; Sui, J.F.; Liu, H.L.; Hou, J.M. Optogenetic neuronal stimulation promotes functional recovery after spinal cord injury. Front. Neurosci. 2021, 15, 371. [Google Scholar] [CrossRef] [PubMed]
- Durieux, L.J.A.; Gilissen, S.R.J.; Arckens, L. Endocannabinoids and cortical plasticity: CB1R as a possible regulator of the excitation/inhibition balance in health and disease. Eur. J. Neurosci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Mysoet, J.; Dupont, E.; Bastide, B.; Canu, M.H. Role of IGF-1 in cortical plasticity and functional deficit induced by sensorimotor restriction. Behav. Brain Res. 2015, 290, 117–123. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, H.S.; Park, M.H. Transcriptome analysis of deafness: Intracellular signal transduction signaling pathways regulate neuroplastic changes in the auditory cortex. Otol. Neurotol. 2020, 41, 986–996. [Google Scholar] [CrossRef]
- Cowansage, K.; LeDoux, J.; Monfils, M.-H. Brain-derived neurotrophic factor: A dynamic gatekeeper of neural plasticity. Curr. Mol. Pharmacol. 2012, 3, 12–29. [Google Scholar] [CrossRef]
- Henderson, L.A.; Gustin, S.M.; Macey, P.M.; Wrigley, P.J.; Siddall, P.J. Functional reorganization of the brain in humans following spinal cord injury: Evidence for underlying changes in cortical anatomy. J. Neurosci. 2011, 31, 2630–2637. [Google Scholar] [CrossRef]
- Freund, P.; Curt, A.; Friston, K.; Thompson, A. Tracking changes following spinal cord injury: Insights from neuroimaging. Neuroscientist 2013, 19, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Serradj, N.; Agger, S.F.; Hollis, E.R. Corticospinal circuit plasticity in motor rehabilitation from spinal cord injury. Neurosci. Lett. 2017, 652, 94–104. [Google Scholar] [CrossRef]
- Christiansen, L.; Perez, M.A. Targeted-plasticity in the corticospinal tract after human spinal cord injury. Neurotherapeutics 2018, 15, 618–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazley, F.A.; Pourmorteza, A.; Gupta, S.; Pashai, N.; Kerr, C.; All, A.H. DTI for assessing axonal integrity after contusive spinal cord injury and transplantation of oligodendrocyte progenitor cells. In Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology Society, San Diego, CA, USA, 28 August–1 September 2012; pp. 82–85. [Google Scholar]
- Vipin, A.; Thow, X.Y.; Mir, H.; Kortelainen, J.; Manivannan, J.; Al-Nashash, H.; All, A.H. Natural progression of spinal cord transection injury and reorganization of neural pathways. J. Neurotrauma 2016, 33, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Baydyuk, M.; Xu, B. BDNF signaling and survival of striatal neurons. Front. Cell. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Huber, E.; Patel, R.; Hupp, M.; Weiskopf, N.; Chakravarty, M.M.; Freund, P. Extrapyramidal plasticity predicts recovery after spinal cord injury. Sci. Rep. 2020, 10, 14102. [Google Scholar] [CrossRef]
- Gelfo, F.; Cutuli, D.; Foti, F.; Laricchiuta, D.; De-Bartolo, P.; Caltagirone, C.; Petrosini, L.; Angelucci, F. Enriched environment improves motor function and increases neurotrophins in hemicerebellar lesioned rats. Neurorehabil. Neural Repair 2011, 25, 243–252. [Google Scholar] [CrossRef]
- Meis, S.; Endres, T.; Munsch, T.; Lessmann, V. The relation between long-term synaptic plasticity at glutamatergic synapses in the amygdala and fear learning in adult heterozygous BDNF-knockout mice. Cereb. Cortex 2018, 28, 1195–1208. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.-L.; Zhang, N.; Xie, X.-M.; Chen, Y.-J.; Wang, R.; Shen, L.; Zhou, J.-S.; Hu, J.-G.; Lü, H.-Z. Transcriptome profile of rat genes in injured spinal cord at different stages by RNA-sequencing. BMC Genom. 2017, 18, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrnes, K.R.; Fricke, S.T.; Faden, A.I. Neuropathological differences between rats and mice after spinal cord injury. J. Magn. Reson. Imaging 2010, 32, 836–846. [Google Scholar] [CrossRef] [Green Version]
- Milich, L.M.; Choi, J.S.; Ryan, C.; Cerqueira, S.R.; Benavides, S.; Yahn, S.L.; Tsoulfas, P.; Lee, J.K. Single-cell analysis of the cellular heterogeneity and interactions in the injured mouse spinal cord. J. Exp. Med. 2021, 218, e20210040. [Google Scholar] [CrossRef]
- Matson, K.J.E.; Russ, D.E.; Kathe, C.; Maric, D.; Hua, I.; Krynitsky, J.; Pursley, R.; Sathyamurthy, A.; Squair, J.W.; Courtine, G.; et al. A single cell atlas of spared tissue below a spinal cord injury reveals cellular mechanisms of repair. bioRxiv 2021, 1–24. [Google Scholar] [CrossRef]
- Ramsey, J.B.G.; Ramer, L.M.; Inskip, J.A.; Alan, N.; Ramer, M.S.; Krassioukov, A.V. Care of rats with complete high-thoracic spinal cord injury. J Neurotrauma. 2010, 27, 1709–1722. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The molecular signatures database hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Carlson, M.; Falcon, S.; Pages, H.; Li, N. A set of annotation maps describing the entire Gene Ontology. R Package Version 2007, 3. [Google Scholar] [CrossRef]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g: Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NAME | ID | SIZE | ES | NES | NOM.p.val | FDR.q.val | FWER.p.val |
|---|---|---|---|---|---|---|---|
| 1 DPL | |||||||
| REACTOME_INTERLEUKIN_4_AND_INTERLEUKIN_13_SIGNALING | R-RNO-6785807 | 8 | 0.922751 | 2.960325 | 0 | 0.001343 ** | 0.001 |
| REACTOME_SIGNALING_BY_INTERLEUKINS | R-RNO-449147 | 15 | 0.59045 | 2.48295 | 0 | 0.02287 * | 0.036 |
| HALLMARK_TNFA_SIGNALING_VIA_NFKB | M5890 | 14 | 0.591473 | 2.428519 | 0 | 0.020713 * | 0.048 |
| GOBP_NEGATIVE_REGULATION_OF_PEPTIDASE_ACTIVITY | GO:0010466 | 6 | 0.83201 | 2.357697 | 0 | 0.026939 * | 0.082 |
| GOBP_OSTEOCLAST_DIFFERENTIATION | GO:0030316 | 6 | 0.807556 | 2.354578 | 0 | 0.021834 * | 0.082 |
| REACTOME_TRANSCRIPTIONAL_REGULATION_OF_WHITE_ADIPOCYTE_DIFFERENTIATION | R-RNO-381340 | 6 | 0.807932 | 2.300164 | 0 | 0.027823 * | 0.125 |
| GOBP_OSSIFICATION | GO:0001503 | 9 | 0.624718 | 2.155755 | 0.003165 | 0.069799 | 0.32 |
| GOBP_NEGATIVE_REGULATION_OF_CYSTEINE_TYPE_ENDOPEPTIDASE_ACTIVITY | GO:2000117 | 5 | 0.793688 | 2.100962 | 0.00274 | 0.09092 | 0.439 |
| GOBP_MYELOID_LEUKOCYTE_DIFFERENTIATION | GO:0002573 | 11 | 0.594451 | 2.088907 | 0.003676 | 0.086556 | 0.455 |
| GOBP_MYELOID_CELL_DIFFERENTIATION | GO:0030099 | 13 | 0.502879 | 2.049668 | 0 | 0.097413 | 0.537 |
| GOBP_GLUTAMATE_RECEPTOR_SIGNALING_PATHWAY | GO:0007215 | 17 | −0.64627 | −2.23902 | 0 | 0.031969 * | 0.036 |
| 8 DPL | |||||||
| REACTOME_INTERLEUKIN_4_AND_INTERLEUKIN_13_SIGNALING | R-RNO-6785807 | 8 | 0.784993 | 2.51836 | 0 | 0.021501 * | 0.015 |
| 45 DPL | |||||||
| GOBP_CELL_CELL_SIGNALING | GO:0007267 | 56 | 0.573257 | 2.007903 | 0 | 0.06429 | 0.117 |
| GOBP_REGULATION_OF_TRANS_SYNAPTIC_SIGNALING | GO:0099177 | 40 | 0.549315 | 1.942583 | 0 | 0.081764 | 0.277 |
| GOBP_POSITIVE_REGULATION_OF_NUCLEOBASE_CONTAINING_COMPOUND_METABOLIC_PROCESS | GO:0045935 | 28 | −0.43293 | −1.97778 | 0 | 0.082897 | 0.595 |
| REACTOME_NGF_STIMULATED_TRANSCRIPTION | R-RNO-9031628 | 9 | −0.66178 | −2.01156 | 0.014235 | 0.069736 | 0.493 |
| GOBP_IMMUNE_SYSTEM_DEVELOPMENT | GO:0002520 | 17 | −0.50455 | −2.01332 | 0 | 0.077424 | 0.489 |
| HALLMARK_APOPTOSIS | M5902 | 6 | −0.79285 | −2.06413 | 0.003185 | 0.057833 | 0.346 |
| GOBP_POSITIVE_REGULATION_OF_BIOSYNTHETIC_PROCESS | GO:0009891 | 27 | −0.47244 | −2.09493 | 0 | 0.05279 | 0.282 |
| GOBP_MONONUCLEAR_CELL_DIFFERENTIATION | GO:1903131 | 7 | −0.80153 | −2.10801 | 0.003155 | 0.057605 | 0.261 |
| GOBP_POSITIVE_REGULATION_OF_TRANSCRIPTION_BY_RNA_POLYMERASE_II | GO:0045944 | 24 | −0.51624 | −2.26319 | 0 | 0.013703 * | 0.056 |
| GOBP_POSITIVE_REGULATION_OF_PRI_MIRNA_TRANSCRIPTION_BY_RNA_POLYMERASE_II | GO:1902895 | 7 | −0.80546 | −2.26659 | 0 | 0.017004 * | 0.052 |
| GOBP_PRI_MIRNA_TRANSCRIPTION_BY_RNA_POLYMERASE_II | GO:0061614 | 7 | −0.80546 | −2.27329 | 0 | 0.022732 * | 0.046 |
| HALLMARK_TNFA_SIGNALING_VIA_NFKB | M5890 | 14 | −0.76035 | −2.75168 | 0 | 0 ** | 0 |
| NAME | ID | SIZE | ES | NES | NOM.p.val | FDR.q.val | FWER.p.val |
|---|---|---|---|---|---|---|---|
| 1 DPL | |||||||
| REACTOME_INTERLEUKIN_4_AND_INTERLEUKIN_13_SIGNALING | R-RNO-6785807 | 8 | 0.9 | 2.823968 | 0 | 0 ** | 0 |
| REACTOME_SIGNALING_BY_INTERLEUKINS | R-RNO-449147 | 15 | 0.571289 | 2.286192 | 0.003788 | 0.09028 | 0.118 |
| GOBP_CYTOKINE_MEDIATED_SIGNALING_PATHWAY | GO:0019221 | 16 | 0.55529 | 2.273639 | 0 | 0.066719 | 0.129 |
| GOBP_NEGATIVE_REGULATION_OF_PEPTIDASE_ACTIVITY | GO:0010466 | 6 | 0.816037 | 2.221286 | 0 | 0.0732 | 0.185 |
| REACTOME_ACTIVATION_OF_NMDA_RECEPTORS_AND_POSTSYNAPTIC_EVENTS | R-RNO-442755 | 16 | −0.61111 | −1.96957 | 0 | 0.06652 | 0.439 |
| REACTOME_TRAFFICKING_OF_AMPA_RECEPTORS | R-RNO-399719 | 11 | −0.69351 | −2.03043 | 0 | 0.03897 * | 0.245 |
| REACTOME_G_ALPHA_I_SIGNALLING_EVENTS | R-RNO-418594 | 18 | −0.61807 | −2.10975 | 0 | 0.01606 * | 0.092 |
| GOBP_CELL_CELL_SIGNALING | GO:0007267 | 56 | −0.56237 | −2.17293 | 0 | 0.00946 * | 0.044 |
| REACTOME_NEUROTRANSMITTER_RECEPTORS_AND_POSTSYNAPTIC_SIGNAL_TRANSMISSION | R-RNO-112314 | 22 | −0.63534 | −2.23874 | 0 | 0.006321 * | 0.023 |
| REACTOME_TRANSMISSION_ACROSS_CHEMICAL_SYNAPSES | R-RNO-112315 | 23 | −0.65278 | −2.33446 | 0 | 0.002479 ** | 0.006 |
| REACTOME_NEURONAL_SYSTEM | R-RNO-112316 | 26 | −0.6346 | −2.35683 | 0 | 0.003253 ** | 0.004 |
| 8 DPL | |||||||
| REACTOME_INTERLEUKIN_4_AND_INTERLEUKIN_13_SIGNALING | R-RNO-6785807 | 8 | 0.85 | 2.650963 | 0 | 0.004133 ** | 0.003 |
| GOBP_NEGATIVE_REGULATION_OF_PEPTIDASE_ACTIVITY | GO:0010466 | 6 | 0.829268 | 2.291362 | 0 | 0.058247 | 0.082 |
| REACTOME_SIGNALING_BY_GPCR | R-RNO-372790 | 22 | −0.5614 | −1.85086 | 0.003501 | 0.098777 | 0.748 |
| REACTOME_ACTIVATION_OF_NMDA_RECEPTORS_AND_POSTSYNAPTIC_EVENTS | R-RNO-442755 | 16 | −0.60603 | −1.90244 | 0.002331 | 0.075281 | 0.55 |
| GOBP_REGULATION_OF_SYNAPTIC_PLASTICITY | GO:0048167 | 21 | −0.58122 | −1.92569 | 0.001145 | 0.063086 | 0.455 |
| REACTOME_G_ALPHA_I_SIGNALLING_EVENTS | R-RNO-418594 | 18 | −0.61145 | −1.95418 | 0 | 0.051105 | 0.361 |
| GOBP_CELL_CELL_SIGNALING | GO:0007267 | 56 | −0.54694 | −1.97041 | 0 | 0.047453 * | 0.304 |
| GOBP_SENSORY_PERCEPTION | GO:0007600 | 9 | −0.74845 | −1.97867 | 0.001295 | 0.050751 | 0.278 |
| GOBP_REGULATION_OF_TRANS_SYNAPTIC_SIGNALING | GO:0099177 | 40 | −0.55088 | −2.01597 | 0 | 0.036091 * | 0.183 |
| GOBP_RESPONSE_TO_ETHANOL | GO:0045471 | 5 | −0.92069 | −2.05396 | 0.001368 | 0.029496 * | 0.128 |
| REACTOME_NEUROTRANSMITTER_RECEPTORS_AND_POSTSYNAPTIC_SIGNAL_TRANSMISSION | R-RNO-112314 | 22 | −0.6351 | −2.10719 | 0 | 0.019659 * | 0.066 |
| REACTOME_TRANSMISSION_ACROSS_CHEMICAL_SYNAPSES | R-RNO-112315 | 23 | −0.6471 | −2.22689 | 0 | 0.004021 ** | 0.009 |
| REACTOME_NEURONAL_SYSTEM | R-RNO-112316 | 26 | −0.6586 | −2.30701 | 0 | 0.003537 ** | 0.004 |
| NAME | ID | SIZE | ES | NES | NOM.p.val | FDR.q.val | FWER.p.val |
|---|---|---|---|---|---|---|---|
| 1 DPL | |||||||
| GOBP_POSITIVE_REGULATION_OF_CELL_POPULATION_PROLIFERATION | GO:0008284 | 21 | −0.55747 | −2.01551 | 0 | 0.074913 | 0.441 |
| GOBP_ACTIVATION_OF_MAPK_ACTIVITY | GO:0000187 | 6 | −0.8384 | −2.01837 | 0 | 0.080351 | 0.431 |
| GOBP_REGULATION_OF_CELL_CELL_ADHESION | GO:0022407 | 9 | −0.75613 | −2.06508 | 0.004202 | 0.055305 | 0.294 |
| GOBP_POSITIVE_REGULATION_OF_LOCOMOTION | GO:0040017 | 11 | −0.70714 | −2.08796 | 0 | 0.049878 * | 0.249 |
| GOBP_POSITIVE_REGULATION_OF_MAP_KINASE_ACTIVITY | GO:0043406 | 6 | −0.8384 | −2.08928 | 0 | 0.056779 | 0.248 |
| GOBP_OSSIFICATION | GO:0001503 | 9 | −0.77164 | −2.11155 | 0 | 0.0504 * | 0.192 |
| GOBP_POSITIVE_REGULATION_OF_MAPK_CASCADE | GO:0043410 | 9 | −0.76861 | −2.15364 | 0 | 0.039986 * | 0.13 |
| GOBP_POSITIVE_REGULATION_OF_ORGANELLE_ORGANIZATION | GO:0010638 | 10 | −0.74741 | −2.15712 | 0.002299 | 0.047046 * | 0.122 |
| GOBP_REGULATION_OF_PEPTIDE_TRANSPORT | GO:0090087 | 10 | −0.73899 | −2.15933 | 0 | 0.060828 | 0.118 |
| GOBP_MUSCLE_CELL_PROLIFERATION | GO:0033002 | 8 | −0.79286 | −2.16342 | 0 | 0.089101 | 0.115 |
| GOBP_REGULATION_OF_RESPONSE_TO_EXTERNAL_STIMULUS | GO:0032101 | 15 | −0.68666 | −2.28367 | 0 | 0.037286 * | 0.025 |
| 8 DPL | |||||||
| REACTOME_NGF_STIMULATED_TRANSCRIPTION | R-RNO-9031628 | 9 | −0.70493 | −1.96836 | 0.006316 | 0.096106 | 0.478 |
| GOBP_MUSCLE_TISSUE_DEVELOPMENT | GO:0060537 | 11 | −0.67464 | −2.0182 | 0.002304 | 0.091098 | 0.334 |
| GOBP_POSITIVE_REGULATION_OF_CELL_POPULATION_PROLIFERATION | GO:0008284 | 21 | −0.57177 | −2.03369 | 0 | 0.09632 | 0.289 |
| GOBP_POSITIVE_REGULATION_OF_BIOSYNTHETIC_PROCESS | GO:0009891 | 27 | −0.53552 | −2.05973 | 0.002088 | 0.099952 | 0.235 |
| NAME | ID | SIZE | ES | NES | NOM.p.val | FDR.q.val | FWER.p.val |
|---|---|---|---|---|---|---|---|
| GOBP_CELL_CYCLE | GO:0007049 | 21 | 0.688508 | 2.187956 | 0 | 0.008712 * | 0.007 |
| GOBP_NEGATIVE_REGULATION_OF_GENE_EXPRESSION | GO:0010629 | 12 | 0.747671 | 2.080675 | 0 | 0.027282 * | 0.043 |
| GOBP_POSTTRANSCRIPTIONAL_REGULATION_OF_GENE_EXPRESSION | GO:0010608 | 8 | 0.837801 | 2.059416 | 0 | 0.022322 * | 0.052 |
| GOBP_CELLULAR_AMIDE_METABOLIC_PROCESS | GO:0043603 | 13 | 0.704689 | 2.000565 | 0 | 0.042499 * | 0.13 |
| GOBP_GROWTH | GO:0040007 | 14 | 0.678726 | 1.941892 | 0 | 0.075044 | 0.264 |
| GOBP_GAMETE_GENERATION | GO:0007276 | 9 | 0.749268 | 1.908591 | 0 | 0.091444 | 0.364 |
| GOBP_PEPTIDE_METABOLIC_PROCESS | GO:0006518 | 12 | 0.689573 | 1.903534 | 0.001361 | 0.082782 | 0.383 |
| NAME | ID | SIZE | ES | NES | NOM.p.val | FDR.q.val | FWER.p.val |
|---|---|---|---|---|---|---|---|
| 8 DPL | |||||||
| GOBP_RESPONSE_TO_TEMPERATURE_STIMULUS | GO:0009266 | 9 | −0.83582 | −1.94008 | 0 | 0.059114 | 0.056 |
| 45 DPL | |||||||
| REACTOME_UNBLOCKING_OF_NMDA_RECEPTORS_GLUTAMATE_BINDING_AND_ACTIVATION | R-RNO-438066 | 12 | −0.65789 | −1.94966 | 0.004431 | 0.091273 | 0.444 |
| REACTOME_NEGATIVE_REGULATION_OF_NMDA_RECEPTOR_MEDIATED_NEURONAL_TRANSMISSION | R-HSA-9617324 | 8 | −0.7625 | −1.95528 | 0.003063 | 0.097942 | 0.429 |
| GOBP_POSITIVE_REGULATION_OF_SYNAPTIC_TRANSMISSION | GO:0050806 | 15 | −0.66669 | −2.06219 | 0 | 0.088657 | 0.138 |
| REACTOME_ACTIVATION_OF_NMDA_RECEPTORS_AND_POSTSYNAPTIC_EVENTS | R-RNO-442755 | 16 | −0.67115 | −2.15044 | 0 | 0.04458 * | 0.035 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldassarro, V.A.; Sanna, M.; Bighinati, A.; Sannia, M.; Gusciglio, M.; Giardino, L.; Lorenzini, L.; Calzà, L. A Time-Course Study of the Expression Level of Synaptic Plasticity-Associated Genes in Un-Lesioned Spinal Cord and Brain Areas in a Rat Model of Spinal Cord Injury: A Bioinformatic Approach. Int. J. Mol. Sci. 2021, 22, 8606. https://doi.org/10.3390/ijms22168606
Baldassarro VA, Sanna M, Bighinati A, Sannia M, Gusciglio M, Giardino L, Lorenzini L, Calzà L. A Time-Course Study of the Expression Level of Synaptic Plasticity-Associated Genes in Un-Lesioned Spinal Cord and Brain Areas in a Rat Model of Spinal Cord Injury: A Bioinformatic Approach. International Journal of Molecular Sciences. 2021; 22(16):8606. https://doi.org/10.3390/ijms22168606
Chicago/Turabian StyleBaldassarro, Vito Antonio, Marco Sanna, Andrea Bighinati, Michele Sannia, Marco Gusciglio, Luciana Giardino, Luca Lorenzini, and Laura Calzà. 2021. "A Time-Course Study of the Expression Level of Synaptic Plasticity-Associated Genes in Un-Lesioned Spinal Cord and Brain Areas in a Rat Model of Spinal Cord Injury: A Bioinformatic Approach" International Journal of Molecular Sciences 22, no. 16: 8606. https://doi.org/10.3390/ijms22168606
APA StyleBaldassarro, V. A., Sanna, M., Bighinati, A., Sannia, M., Gusciglio, M., Giardino, L., Lorenzini, L., & Calzà, L. (2021). A Time-Course Study of the Expression Level of Synaptic Plasticity-Associated Genes in Un-Lesioned Spinal Cord and Brain Areas in a Rat Model of Spinal Cord Injury: A Bioinformatic Approach. International Journal of Molecular Sciences, 22(16), 8606. https://doi.org/10.3390/ijms22168606