
Presence, Mode of Action, and Application of Pathway Specific Transcription Factors in Aspergillus Biosynthetic Gene Clusters




Abstract
:1. Introduction
2. Occurrence and Types of Pathway Specific Transcription Factors
3. Role and Evolution of Pathway Specific Transcription Factor (PSTF)
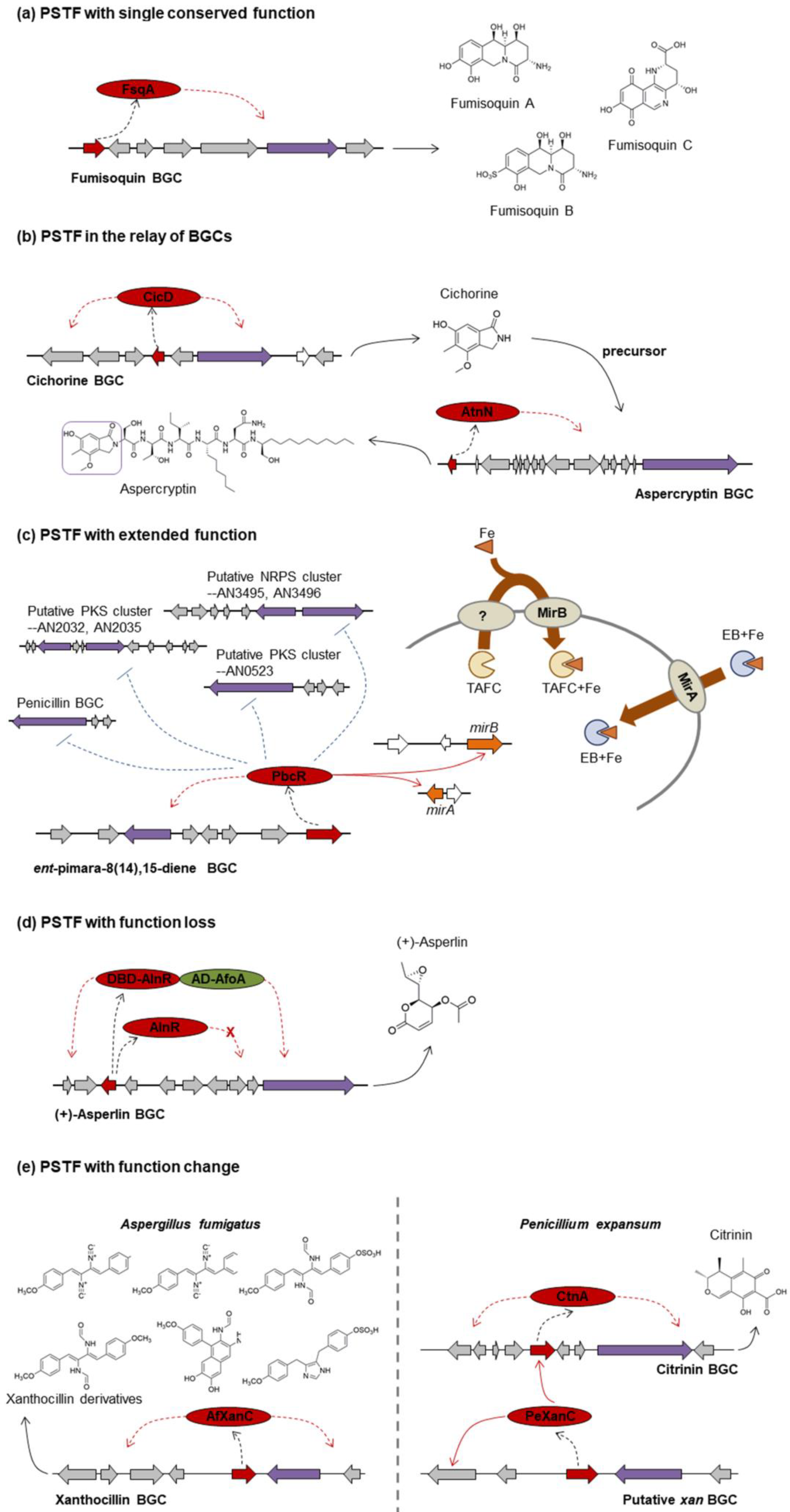
3.1. One PSTF per BGC
3.1.1. PSTF with Single Conserved Function
3.1.2. PSTF in the Relay of BGCs: cicD, atnN
3.1.3. PSTF with Extended Function: pbcR
3.1.4. PSTF with Function Loss: alnR
3.1.5. PSTF with Function Change: xanC
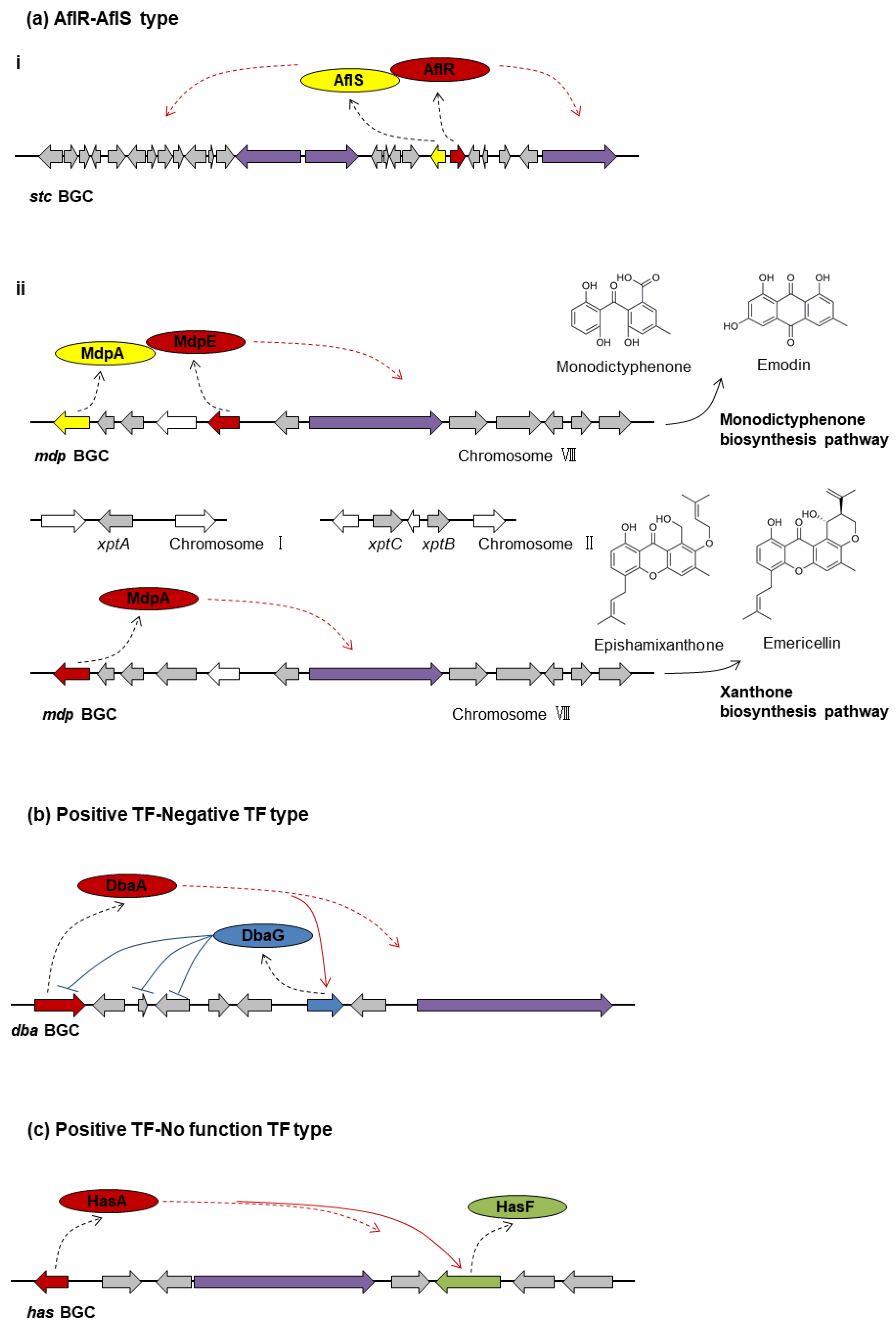
3.2. Two PSTFs per BGC
3.2.1. AflR-AflS Type
- aflR-aflS
- mdpE-mdpA
- tpcE-tpcD
3.2.2. Positive TF-Negative TF Type: dbaA-dbaG
3.2.3. Positive TF-No Function TF Type: hasA-hasF
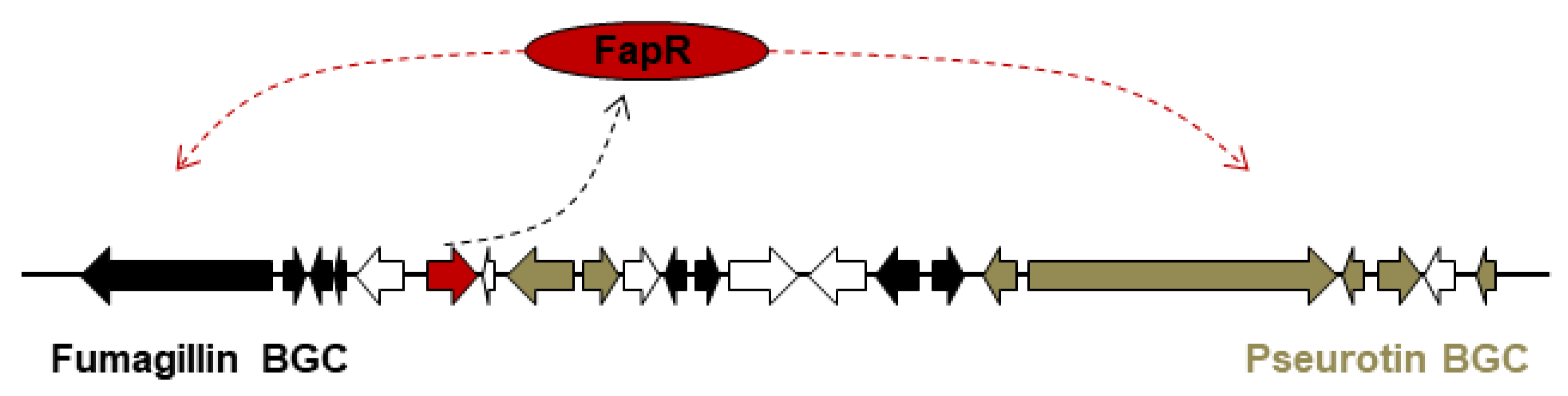
3.3. One PSTF for Two BGCs: fapR
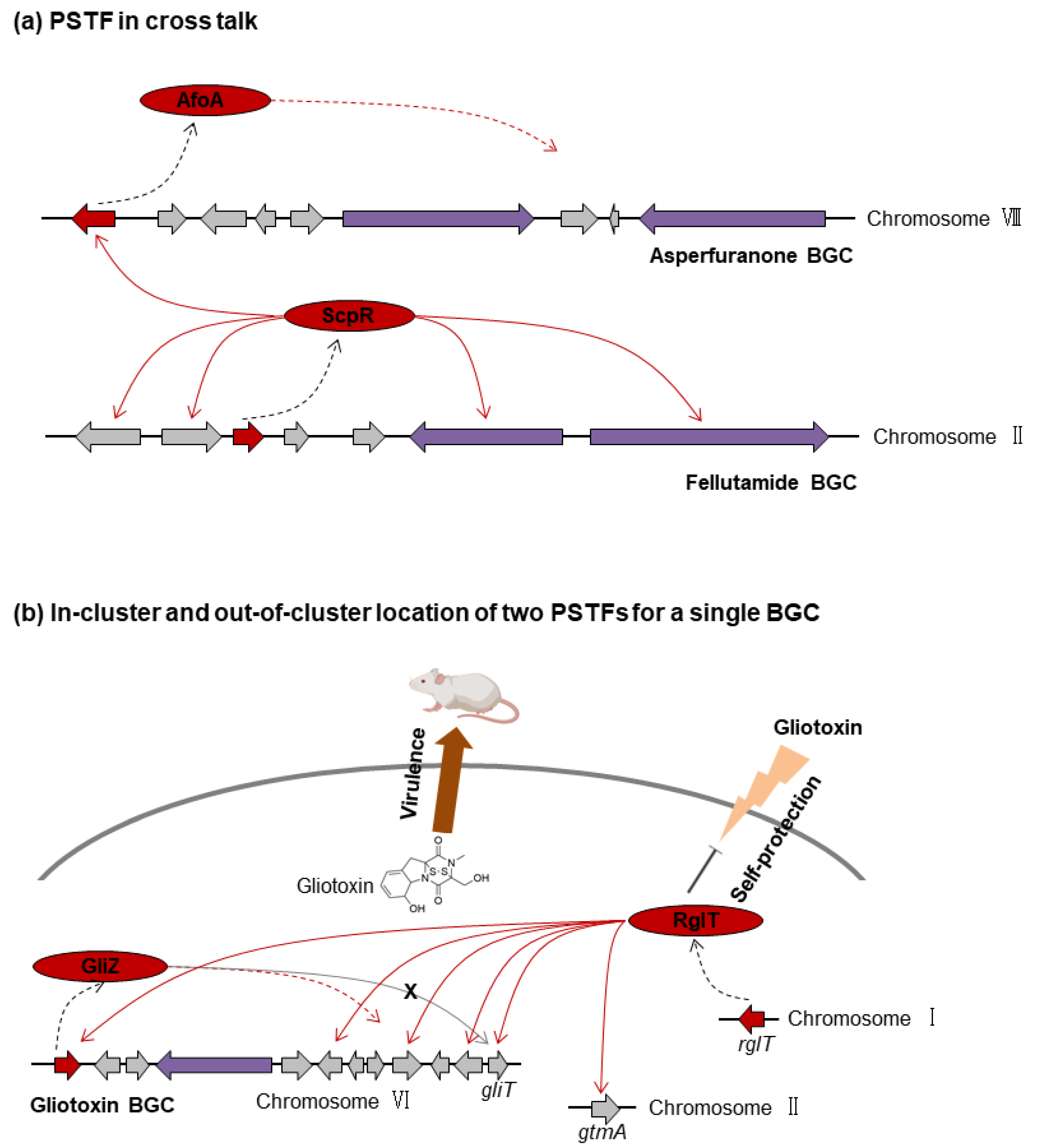
3.4. Additional Complexities in PSTF Regulation
3.4.1. PSTF in Cross Talk: scpR-afoA
3.4.2. In-Cluster and Out-of-Cluster Locations of Two PSTFs for a Single BGC: gliZ and rglT
4. Pathway Specific Approaches to Explore Biosynthetic Gene Clusters
4.1. Overexpression/Deletion of PSTF
4.2. Synthetic PSTF
4.2.1. Hybrid PSTF
4.2.2. CRISPR-Mediated Activation System (Artificial PSTF)
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Robey, M.T.; Caesar, L.K.; Drott, M.T.; Keller, N.P.; Kelleher, N.L. An interpreted atlas of biosynthetic gene clusters from 1000 fungal genomes. Proc. Natl. Acad. Sci. USA 2021, 118, e2020230118. [Google Scholar] [CrossRef]
- Keller, N.P. Translating biosynthetic gene clusters into fungal armor and weaponry. Nat. Chem. Biol. 2015, 11, 671–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macheleidt, J.; Mattern, D.J.; Fischer, J.; Netzker, T.; Weber, J.; Schroeckh, V.; Valiante, V.; Brakhage, A.A. Regulation and role of fungal secondary metabolites. Annu. Rev. Genet. 2016, 50, 371–392. [Google Scholar] [CrossRef]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Sbaraini, N.; Hu, J.; Roux, I.; Phan, C.S.; Motta, H.; Rezaee, H.; Schrank, A.; Chooi, Y.H.; Staats, C.C. Polyketides produced by the entomopathogenic fungus Metarhizium anisopliae induce Candida albicans growth. Fungal Genet. Biol. 2021, 152, 103568. [Google Scholar] [CrossRef]
- Lim, S.; Bijlani, S.; Blachowicz, A.; Chiang, Y.-M.; Lee, M.-S.; Torok, T.; Venkateswaran, K.; Wang, C.C.C. Identification of the pigment and its role in UV resistance in Paecilomyces variotii, a Chernobyl isolate, using genetic manipulation strategies. Fungal Genet. Biol. 2021, 152, 103567. [Google Scholar] [CrossRef] [PubMed]
- Boruta, T. Uncovering the repertoire of fungal secondary metabolites: From Fleming’s laboratory to the International Space Station. Bioengineered 2018, 9, 12–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.A.; Bacha, N.; Ahmad, B.; Lutfullah, G.; Farooq, U.; Cox, R.J. Fungi as chemical industries and genetic engineering for the production of biologically active secondary metabolites. Asian Pac. J. Trop. Biomed. 2014, 4, 859–870. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Ahuja, S.; Garg, S. Fungal secondary metabolites: Biological activity and potential applications. In Recent Trends in Mycological Research: Volume 1: Agricultural and Medical Perspective; Yadav, A.N., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 159–188. [Google Scholar]
- Florey, H.W. Penicillin: A survey. Br. Med. J. 1944, 2, 169–171. [Google Scholar] [CrossRef]
- Anjum, T.; Azam, A.; Irum, W. Production of cyclosporine A by submerged fermentation from a local isolate of Penicillium fellutanum. Indian J. Pharm. Sci. 2012, 74, 372–374. [Google Scholar] [CrossRef] [Green Version]
- Endo, A. A historical perspective on the discovery of statins. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Tsunematsu, Y. Genomics-directed activation of cryptic natural product pathways deciphers codes for biosynthesis and molecular function. J. Nat. Med. 2021, 75, 261–274. [Google Scholar] [CrossRef]
- Greco, C.; Keller, N.P.; Rokas, A. Unearthing fungal chemodiversity and prospects for drug discovery. Curr. Opin. Microbiol. 2019, 51, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Kjaerbolling, I.; Mortensen, U.H.; Vesth, T.; Andersen, M.R. Strategies to establish the link between biosynthetic gene clusters and secondary metabolites. Fungal Genet. Biol. 2019, 130, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Lim, F.Y.; Won, T.H.; Raffa, N.; Baccile, J.A.; Wisecaver, J.; Rokas, A.; Schroeder, F.C.; Keller, N.P. Fungal isocyanide synthases and xanthocillin biosynthesis in Aspergillus fumigatus. mBio 2018, 9, e00785-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Estrada, C.; Dominguez-Santos, R.; Kosalkova, K.; Martin, J.F. Transcription factors controlling primary and secondary metabolism in filamentous fungi: The beta-Lactam paradigm. Fermentation 2018, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Keller, N.P. Transcriptional regulatory elements in fungal secondary metabolism. J. Microbiol. 2011, 49, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Shelest, E. Transcription factors in fungi. FEMS Microbiol. Lett. 2008, 286, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Park, H.S.; Bayram, O.; Braus, G.H.; Kim, S.C.; Yu, J.H. Characterization of the velvet regulators in Aspergillus fumigatus. Mol. Microbiol. 2012, 86, 937–953. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Y.L.; Gerke, J.; Park, H.-S.; Bayram, Ö.; Neumann, P.; Ni, M.; Dickmanns, A.; Kim, S.C.; Yu, J.-H.; Braus, G.H.; et al. The velvet family of fungal regulators contains a DNA-binding domain structurally similar to NF-κB. PLoS Biol. 2013, 11, e1001750. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Chen, Y.; Tian, S. Function of pH-dependent transcription factor PacC in regulating development, pathogenicity, and mycotoxin biosynthesis of phytopathogenic fungi. FEBS J. 2021, 15808. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Chiang, Y.M.; Szewczyk, E.; Reyes-Dominguez, Y.; Davidson, A.D.; Sanchez, J.F.; Lo, H.C.; Watanabe, K.; Strauss, J.; Oakley, B.R.; et al. Chromatin-level regulation of biosynthetic gene clusters. Nat. Chem. Biol. 2009, 5, 462–464. [Google Scholar] [CrossRef] [Green Version]
- Pfannenstiel, B.T.; Keller, N.P. On top of biosynthetic gene clusters: How epigenetic machinery influences secondary metabolism in fungi. Biotechnol. Adv. 2019, 37, 107345. [Google Scholar] [CrossRef]
- Roux, I.; Woodcraft, C.; Hu, J.; Wolters, R.; Gilchrist, C.L.M.; Chooi, Y.H. CRISPR-mediated activation of biosynthetic gene clusters for bioactive molecule discovery in filamentous fungi. ACS Synth. Biol. 2020, 9, 1843–1854. [Google Scholar] [CrossRef]
- Caesar, L.K.; Kelleher, N.L.; Keller, N.P. In the fungus where it happens: History and future propelling Aspergillus nidulans as the archetype of natural products research. Fungal Genet. Biol. 2020, 144, 103477. [Google Scholar] [CrossRef] [PubMed]
- Romsdahl, J.; Wang, C.C.C. Recent advances in the genome mining of Aspergillus secondary metabolites (covering 2012–2018). Medchemcomm 2019, 10, 840–866. [Google Scholar] [CrossRef]
- Stroe, M.C.; Netzker, T.; Scherlach, K.; Kruger, T.; Hertweck, C.; Valiante, V.; Brakhage, A.A. Targeted induction of a silent fungal gene cluster encoding the bacteria-specific germination inhibitor fumigermin. Elife 2020, 9, e52541. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Zhang, P.; Tao, Q.; Keller, N.P.; Yang, Y.; Yin, W.B.; Liu, H. Characterization and biosynthesis of a rare fungal hopane-type triterpenoid glycoside involved in the antistress property of Aspergillus fumigatus. Org. Lett. 2019, 21, 3252–3256. [Google Scholar] [CrossRef] [PubMed]
- Schmaler-Ripcke, J.; Sugareva, V.; Gebhardt, P.; Winkler, R.; Kniemeyer, O.; Heinekamp, T.; Brakhage, A.A. Production of pyomelanin, a second type of melanin, via the tyrosine degradation pathway in Aspergillus fumigatus. Appl. Environ. Microbiol. 2009, 75, 493–503. [Google Scholar] [CrossRef] [Green Version]
- Yeh, H.H.; Ahuja, M.; Chiang, Y.M.; Oakley, C.E.; Moore, S.; Yoon, O.; Hajovsky, H.; Bok, J.W.; Keller, N.P.; Wang, C.C.; et al. Resistance gene-guided genome mining: Serial promoter exchanges in Aspergillus nidulans reveal the biosynthetic pathway for fellutamide B, a proteasome inhibitor. ACS Chem. Biol. 2016, 11, 2275–2284. [Google Scholar] [CrossRef]
- Li, S.M.; Unsold, I.A. Post-genome research on the biosynthesis of ergot alkaloids. Planta Med. 2006, 72, 1117–1120. [Google Scholar] [CrossRef]
- O’Hanlon, K.A.; Gallagher, L.; Schrettl, M.; Jochl, C.; Kavanagh, K.; Larsen, T.O.; Doyle, S. Nonribosomal peptide synthetase genes pesL and pes1 are essential for Fumigaclavine C production in Aspergillus fumigatus. Appl. Environ. Microbiol. 2012, 78, 3166–3176. [Google Scholar] [CrossRef] [Green Version]
- Westphal, K.R.; Muurmann, A.T.; Paulsen, I.E.; Nørgaard, K.T.H.; Overgaard, M.L.; Dall, S.M.; Aalborg, T.; Wimmer, R.; Sørensen, J.L.; Sondergaard, T.E. Who needs neighbors? PKS8 Is a stand-alone gene in Fusarium graminearum responsible for production of gibepyrones and prolipyrone B. Molecules 2018, 23, 2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.A. Fungal gene clusters. Nat. Biotechnol. 2007, 25, 67. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, M.; Chiang, Y.M.; Chang, S.L.; Praseuth, M.B.; Entwistle, R.; Sanchez, J.F.; Lo, H.C.; Yeh, H.H.; Oakley, B.R.; Wang, C.C. Illuminating the diversity of aromatic polyketide synthases in Aspergillus nidulans. J. Am. Chem. Soc. 2012, 134, 8212–8221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cary, J.W.; Ehrlich, K.C. Aflatoxigenicity in Aspergillus: Molecular genetics, phylogenetic relationships and evolutionary implications. Mycopathologia 2006, 162, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Dhingra, S. Conserved regulatory mechanisms controlling aflatoxin and sterigmatocystin biosynthesis. In Aflatoxins: Biochemistry and Molecular Biology; Intech Open: Rijeka, Croatia, 2011; pp. 67–88. [Google Scholar]
- Oakley, C.E.; Ahuja, M.; Sun, W.W.; Entwistle, R.; Akashi, T.; Yaegashi, J.; Guo, C.J.; Cerqueira, G.C.; Russo Wortman, J.; Wang, C.C.; et al. Discovery of McrA, a master regulator of Aspergillus secondary metabolism. Mol. Microbiol. 2017, 103, 347–365. [Google Scholar] [CrossRef] [Green Version]
- Gerke, J.; Bayram, O.; Feussner, K.; Landesfeind, M.; Shelest, E.; Feussner, I.; Braus, G.H. Breaking the silence: Protein stabilization uncovers silenced biosynthetic gene clusters in the fungus Aspergillus nidulans. Appl. Environ. Microbiol. 2012, 78, 8234–8244. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.M.; Ahuja, M.; Oakley, C.E.; Entwistle, R.; Asokan, A.; Zutz, C.; Wang, C.C.; Oakley, B.R. Development of genetic dereplication strains in Aspergillus nidulans results in the discovery of aspercryptin. Angew. Chem. Int. Ed. Engl. 2016, 55, 1662–1665. [Google Scholar] [CrossRef] [Green Version]
- Henke, M.T.; Soukup, A.A.; Goering, A.W.; McClure, R.A.; Thomson, R.J.; Keller, N.P.; Kelleher, N.L. New aspercryptins, lipopeptide natural products, revealed by HDAC inhibition in Aspergillus nidulans. ACS Chem. Biol. 2016, 11, 2117–2123. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.M.; Szewczyk, E.; Davidson, A.D.; Keller, N.; Oakley, B.R.; Wang, C.C. A gene cluster containing two fungal polyketide synthases encodes the biosynthetic pathway for a polyketide, asperfuranone, in Aspergillus nidulans. J. Am. Chem. Soc. 2009, 131, 2965–2970. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, S.; Funk, A.N.; Scherlach, K.; Schroeckh, V.; Shelest, E.; Horn, U.; Hertweck, C.; Brakhage, A.A. Activation of a silent fungal polyketide biosynthesis pathway through regulatory cross talk with a cryptic nonribosomal peptide synthetase gene cluster. Appl. Environ. Microbiol. 2010, 76, 8143–8149. [Google Scholar] [CrossRef] [Green Version]
- Grau, M.F.; Entwistle, R.; Chiang, Y.M.; Ahuja, M.; Oakley, C.E.; Akashi, T.; Wang, C.C.C.; Todd, R.B.; Oakley, B.R. Hybrid transcription factor engineering activates the silent secondary metabolite gene cluster for (+)-asperlin in Aspergillus nidulans. ACS Chem. Biol. 2018, 3, 3193–3205. [Google Scholar] [CrossRef]
- Yaegashi, J.; Praseuth, M.B.; Tyan, S.W.; Sanchez, J.F.; Entwistle, R.; Chiang, Y.M.; Oakley, B.R.; Wang, C.C. Molecular genetic characterization of the biosynthesis cluster of a prenylated isoindolinone alkaloid aspernidine A in Aspergillus nidulans. Org. Lett. 2013, 15, 2862–2865. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.S.; Chen, B.; Chiang, Y.M.; Wang, C.C.C. Discovery and elucidation of the biosynthesis of aspernidgulenes: Novel polyenes from Aspergillus nidulans by using serial promoter replacement. Chembiochem 2019, 20, 329–334. [Google Scholar] [CrossRef]
- Szewczyk, E.; Chiang, Y.M.; Oakley, C.E.; Davidson, A.D.; Wang, C.C.; Oakley, B.R. Identification and characterization of the asperthecin gene cluster of Aspergillus nidulans. Appl. Environ. Microbiol. 2008, 74, 7607–7612. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, S.; Schumann, J.; Scherlach, K.; Lange, C.; Brakhage, A.A.; Hertweck, C. Genomics-driven discovery of PKS-NRPS hybrid metabolites from Aspergillus nidulans. Nat. Chem. Biol. 2007, 3, 213–217. [Google Scholar] [CrossRef]
- Lo, H.C.; Entwistle, R.; Guo, C.J.; Ahuja, M.; Szewczyk, E.; Hung, J.H.; Chiang, Y.M.; Oakley, B.R.; Wang, C.C. Two separate gene clusters encode the biosynthetic pathway for the meroterpenoids austinol and dehydroaustinol in Aspergillus nidulans. J. Am. Chem. Soc. 2012, 134, 4709–4720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, J.F.; Entwistle, R.; Corcoran, D.; Oakley, B.R.; Wang, C.C. Identification and molecular genetic analysis of the cichorine gene cluster in Aspergillus nidulans. Medchemcomm 2012, 3, 997–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttel, W.; Youssar, L.; Gruning, B.A.; Gunther, S.; Hugentobler, K.G. Echinocandin B biosynthesis: A biosynthetic cluster from Aspergillus nidulans NRRL 8112 and reassembly of the subclusters Ecd and Hty from Aspergillus pachycristatus NRRL 11440 reveals a single coherent gene cluster. BMC Genom. 2016, 17, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, Y.M.; Szewczyk, E.; Nayak, T.; Davidson, A.D.; Sanchez, J.F.; Lo, H.C.; Ho, W.Y.; Simityan, H.; Kuo, E.; Praseuth, A.; et al. Molecular genetic mining of the Aspergillus secondary metabolome: Discovery of the emericellamide biosynthetic pathway. Chem. Biol. 2008, 15, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Bromann, K.; Toivari, M.; Viljanen, K.; Vuoristo, A.; Ruohonen, L.; Nakari-Setala, T. Identification and characterization of a novel diterpene gene cluster in Aspergillus nidulans. PLoS ONE 2012, 7, e35450. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.F.; Chiang, Y.M.; Szewczyk, E.; Davidson, A.D.; Ahuja, M.; Elizabeth Oakley, C.; Woo Bok, J.; Keller, N.; Oakley, B.R.; Wang, C.C. Molecular genetic analysis of the orsellinic acid/F9775 gene cluster of Aspergillus nidulans. Mol. Biosyst. 2010, 6, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.L.; Nielsen, J.B.; Rank, C.; Klejnstrup, M.L.; Holm, D.K.; Brogaard, K.H.; Hansen, B.G.; Frisvad, J.C.; Larsen, T.O.; Mortensen, U.H. A genome-wide polyketide synthase deletion library uncovers novel genetic links to polyketides and meroterpenoids in Aspergillus nidulans. FEMS Microbiol. Lett. 2011, 321, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Eisendle, M.; Oberegger, H.; Zadra, I.; Haas, H. The siderophore system is essential for viability of Aspergillus nidulans: Functional analysis of two genes encoding L-ornithine N5-monooxygenase (sidA) and a non-ribosomal peptide synthetase (sidC). Mol. Microbiol. 2003, 49, 359–375. [Google Scholar] [CrossRef] [PubMed]
- Sung, C.T.; Chang, S.L.; Entwistle, R.; Ahn, G.; Lin, T.S.; Petrova, V.; Yeh, H.H.; Praseuth, M.B.; Chiang, Y.M.; Oakley, B.R.; et al. Overexpression of a three-gene conidial pigment biosynthetic pathway in Aspergillus nidulans reveals the first NRPS known to acetylate tryptophan. Fungal Genet. Biol. 2017, 101, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, N.; Tanaka, H.; Koyama, F.; Noguchi, H.; Wang, C.C.; Hotta, K.; Watanabe, K. Non-heme dioxygenase catalyzes atypical oxidations of 6,7-bicyclic systems to form the 6,6-quinolone core of viridicatin-type fungal alkaloids. Angew. Chem. Int. Ed. Engl. 2014, 53, 12880–12884. [Google Scholar] [CrossRef]
- Yeh, H.H.; Chiang, Y.M.; Entwistle, R.; Ahuja, M.; Lee, K.H.; Bruno, K.S.; Wu, T.K.; Oakley, B.R.; Wang, C.C. Molecular genetic analysis reveals that a nonribosomal peptide synthetase-like (NRPS-like) gene in Aspergillus nidulans is responsible for microperfuranone biosynthesis. Appl. Microbiol. Biotechnol. 2012, 96, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.F.; Entwistle, R.; Hung, J.H.; Yaegashi, J.; Jain, S.; Chiang, Y.M.; Wang, C.C.; Oakley, B.R. Genome-based deletion analysis reveals the prenyl xanthone biosynthesis pathway in Aspergillus nidulans. J. Am. Chem. Soc. 2011, 133, 4010–4017. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.M.; Szewczyk, E.; Davidson, A.D.; Entwistle, R.; Keller, N.P.; Wang, C.C.; Oakley, B.R. Characterization of the Aspergillus nidulans monodictyphenone gene cluster. Appl. Environ. Microbiol. 2010, 76, 2067–2074. [Google Scholar] [CrossRef] [Green Version]
- Scherlach, K.; Sarkar, A.; Schroeckh, V.; Dahse, H.M.; Roth, M.; Brakhage, A.A.; Horn, U.; Hertweck, C. Two induced fungal polyketide pathways converge into antiproliferative spiroanthrones. Chembiochem 2011, 12, 1836–1839. [Google Scholar] [CrossRef]
- Andersen, M.R.; Nielsen, J.B.; Klitgaard, A.; Petersen, L.M.; Zachariasen, M.; Hansen, T.J.; Blicher, L.H.; Gotfredsen, C.H.; Larsen, T.O.; Nielsen, K.F.; et al. Accurate prediction of secondary metabolite gene clusters in filamentous fungi. Proc. Natl. Acad. Sci. USA 2013, 110, E99–E107. [Google Scholar] [CrossRef] [Green Version]
- Bergh, K.T.; Brakhage, A.A. Regulation of the Aspergillus nidulans penicillin biosynthesis gene acvA (pcbAB) by amino acids: Implication for involvement of transcription factor PACC. Appl. Environ. Microbiol. 1998, 64, 843–849. [Google Scholar] [CrossRef] [Green Version]
- Bouhired, S.; Weber, M.; Kempf-Sontag, A.; Keller, N.P.; Hoffmeister, D. Accurate prediction of the Aspergillus nidulans terrequinone gene cluster boundaries using the transcriptional regulator LaeA. Fungal. Genet. Biol. 2007, 44, 1134–1145. [Google Scholar] [CrossRef]
- Chooi, Y.H.; Cacho, R.; Tang, Y. Identification of the viridicatumtoxin and griseofulvin gene clusters from Penicillium aethiopicum. Chem. Biol. 2010, 17, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Drott, M.T.; Bastos, R.W.; Rokas, A.; Ries, L.N.A.; Gabaldon, T.; Goldman, G.H.; Keller, N.P.; Greco, C. Diversity of secondary metabolism in Aspergillus nidulans clinical isolates. mSphere 2020, 5, e00156-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayorga, M.E.; Timberlake, W.E. Isolation and molecular characterization of the Aspergillus nidulans wA gene. Genetics 1990, 126, 73–79. [Google Scholar] [CrossRef]
- Tsai, H.F.; Wheeler, M.H.; Chang, Y.C.; Kwon-Chung, K.J. A developmentally regulated gene cluster involved in conidial pigment biosynthesis in Aspergillus fumigatus. J. Bacteriol. 1999, 181, 6469–6477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayry, J.; Beaussart, A.; Dufrene, Y.F.; Sharma, M.; Bansal, K.; Kniemeyer, O.; Aimanianda, V.; Brakhage, A.A.; Kaveri, S.V.; Kwon-Chung, K.J.; et al. Surface structure characterization of Aspergillus fumigatus conidia mutated in the melanin synthesis pathway and their human cellular immune response. Infect. Immun. 2014, 82, 3141–3153. [Google Scholar] [CrossRef] [Green Version]
- Throckmorton, K.; Lim, F.Y.; Kontoyiannis, D.P.; Zheng, W.; Keller, N.P. Redundant synthesis of a conidial polyketide by two distinct secondary metabolite clusters in Aspergillus fumigatus. Environ. Microbiol. 2016, 18, 246–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blatzer, M.; Schrettl, M.; Sarg, B.; Lindner, H.H.; Pfaller, K.; Haas, H. SidL, an Aspergillus fumigatus transacetylase involved in biosynthesis of the siderophores ferricrocin and hydroxyferricrocin. Appl. Environ. Microb. 2011, 77, 4959–4966. [Google Scholar] [CrossRef] [Green Version]
- Schrettl, M.; Kim, H.S.; Eisendle, M.; Kragl, C.; Nierman, W.C.; Heinekamp, T.; Werner, E.R.; Jacobsen, I.; Illmer, P.; Yi, H.; et al. SreA-mediated iron regulation in Aspergillus fumigatus. Mol. Microbiol. 2008, 70, 27–43. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.C.; Chooi, Y.H.; Dhingra, S.; Xu, W.; Calvo, A.M.; Tang, Y. The fumagillin biosynthetic gene cluster in Aspergillus fumigatus encodes a cryptic terpene cyclase involved in the formation of β-trans-bergamotene. J. Am. Chem. Soc. 2013, 135, 4616–4619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhingra, S.; Lind, A.L.; Lin, H.C.; Tang, Y.; Rokas, A.; Calvo, A.M. The fumagillin gene cluster, an example of hundreds of genes under veA control in Aspergillus fumigatus. PLoS ONE 2013, 8, e77147. [Google Scholar] [CrossRef] [Green Version]
- Wiemann, P.; Guo, C.J.; Palmer, J.M.; Sekonyela, R.; Wang, C.C.; Keller, N.P. Prototype of an intertwined secondary-metabolite supercluster. Proc. Natl. Acad. Sci. USA 2013, 110, 17065–17070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ames, B.D.; Haynes, S.W.; Gao, X.; Evans, B.S.; Kelleher, N.L.; Tang, Y.; Walsh, C.T. Complexity generation in fungal peptidyl alkaloid biosynthesis: Oxidation of fumiquinazoline A to the heptacyclic hemiaminal fumiquinazoline C by the flavoenzyme Af12070 from Aspergillus fumigatus. Biochemistry 2011, 50, 8756–8769. [Google Scholar] [CrossRef] [Green Version]
- Macheleidt, J.; Scherlach, K.; Neuwirth, T.; Schmidt-Heck, W.; Straßburger, M.; Spraker, J.; Baccile, J.A.; Schroeder, F.C.; Keller, N.P.; Hertweck, C.; et al. Transcriptome analysis of cyclic AMP-dependent protein kinase A–regulated genes reveals the production of the novel natural compound fumipyrrole by Aspergillus fumigatus. Mol. Microbiol. 2015, 96, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Baccile, J.A.; Spraker, J.E.; Le, H.H.; Brandenburger, E.; Gomez, C.; Bok, J.W.; Macheleidt, J.; Brakhage, A.A.; Hoffmeister, D.; Keller, N.P.; et al. Plant-like biosynthesis of isoquinoline alkaloids in Aspergillus fumigatus. Nat. Chem. Biol. 2016, 12, 419–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiya, S.; Grundmann, A.; Li, S.M.; Turner, G. The fumitremorgin gene cluster of Aspergillus fumigatus: Identification of a gene encoding brevianamide F synthetase. Chembiochem 2006, 7, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Suzuki, H.; Takagi, H.; Asami, Y.; Kakeya, H.; Uramoto, M.; Usui, T.; Takahashi, S.; Sugimoto, Y.; Osada, H. Identification of cytochrome P450s required for fumitremorgin biosynthesis in Aspergillus fumigatus. Chembiochem 2009, 10, 920–928. [Google Scholar] [CrossRef]
- Gardiner, D.M.; Howlett, B.J. Bioinformatic and expression analysis of the putative gliotoxin biosynthetic gene cluster of Aspergillus fumigatus. FEMS Microbiol. Lett. 2005, 248, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, D.M.; Waring, P.; Howlett, B.J. The epipolythiodioxopiperazine (ETP) class of fungal toxins: Distribution, mode of action, functions and biosynthesis. Microbiology 2005, 151, 1021–1032. [Google Scholar] [CrossRef] [Green Version]
- Forseth, R.R.; Fox, E.M.; Chung, D.; Howlett, B.J.; Keller, N.P.; Schroeder, F.C. Identification of cryptic products of the gliotoxin gene cluster using NMR-based comparative metabolomics and a model for gliotoxin biosynthesis. J. Am. Chem. Soc. 2011, 133, 9678–9681. [Google Scholar] [CrossRef] [Green Version]
- Ries, L.N.A.; Pardeshi, L.; Dong, Z.L.; Tan, K.; Steenwyk, J.L.; Colabardini, A.C.; Ferreira, J.A.; de Castro, P.A.; Silva, L.P.; Preite, N.W.; et al. The Aspergillus fumigatus transcription factor RglT is important for gliotoxin biosynthesis and self-protection, and virulence. PLoS Pathog. 2020, 16, e1008645. [Google Scholar] [CrossRef]
- Lv, J.M.; Hu, D.; Gao, H.; Kushiro, T.; Awakawa, T.; Chen, G.D.; Wang, C.X.; Abe, I.; Yao, X.S. Biosynthesis of helvolic acid and identification of an unusual C-4-demethylation process distinct from sterol biosynthesis. Nat. Commun. 2017, 8, 1644. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.B.; Baccile, J.A.; Bok, J.W.; Chen, Y.; Keller, N.P.; Schroeder, F.C. A nonribosomal peptide synthetase-derived iron(III) complex from the pathogenic fungus Aspergillus fumigatus. J. Am. Chem. Soc. 2013, 135, 2064–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chooi, Y.H.; Wang, P.; Fang, J.; Li, Y.; Wu, K.; Wang, P.; Tang, Y. Discovery and characterization of a group of fungal polycyclic polyketide prenyltransferases. J. Am. Chem. Soc. 2012, 134, 9428–9437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konig, C.C.; Scherlach, K.; Schroeckh, V.; Horn, F.; Nietzsche, S.; Brakhage, A.A.; Hertweck, C. Bacterium induces cryptic meroterpenoid pathway in the pathogenic fungus Aspergillus fumigatus. Chembiochem 2013, 14, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.H.; Fang, J.; Liu, H.; Filler, S.G.; Wang, P.; Tang, Y. Genome mining of a prenylated and immunosuppressive polyketide from pathogenic fungi. Org. Lett. 2013, 15, 780–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vodisch, M.; Scherlach, K.; Winkler, R.; Hertweck, C.; Braun, H.P.; Roth, M.; Haas, H.; Werner, E.R.; Brakhage, A.A.; Kniemeyer, O. Analysis of the Aspergillus fumigatus proteome reveals metabolic changes and the activation of the pseurotin A biosynthesis gene cluster in response to hypoxia. J. Proteome. Res. 2011, 10, 2508–2524. [Google Scholar] [CrossRef]
- Keller, S.; Macheleidt, J.; Scherlach, K.; Schmaler-Ripcke, J.; Jacobsen, I.D.; Heinekamp, T.; Brakhage, A.A. Pyomelanin formation in Aspergillus fumigatus requires HmgX and the transcriptional activator HmgR but is dispensable for virulence. PLoS ONE 2011, 6, e26604. [Google Scholar] [CrossRef]
- Itoh, T.; Tokunaga, K.; Matsuda, Y.; Fujii, I.; Abe, I.; Ebizuka, Y.; Kushiro, T. Reconstitution of a fungal meroterpenoid biosynthesis reveals the involvement of a novel family of terpene cyclases. Nat. Chem. 2010, 2, 858–864. [Google Scholar] [CrossRef]
- Wang, W.; Drott, M.; Greco, C.; Luciano-Rosario, D.; Wang, P.; Keller, N.P. Transcription factor repurposing offers insights into evolution of biosynthetic gene cluster regulation. mBio 2021, e01399-21. [Google Scholar] [CrossRef]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered pathway genes in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.H.; Butchko, R.A.; Fernandes, M.; Keller, N.P.; Leonard, T.J.; Adams, T.H. Conservation of structure and function of the aflatoxin regulatory gene aflR from Aspergillus nidulans and A. flavus. Curr. Genet. 1996, 29, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K. Lack of interaction between AFLR and AFLJ contributes to nonaflatoxigenicity of Aspergillus sojae. J. Biotechnol. 2004, 107, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Perrin, R.M.; Fedorova, N.D.; Bok, J.W.; Cramer, R.A.; Wortman, J.R.; Kim, H.S.; Nierman, W.C.; Keller, N.P. Transcriptional regulation of chemical diversity in Aspergillus fumigatus by LaeA. PLoS Pathog. 2007, 3, e50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowles, S.L.; Mead, M.E.; Silva, L.P.; Raja, H.A.; Steenwyk, J.L.; Goldman, G.H.; Oberlies, N.H.; Rokas, A. Gliotoxin, a known virulence factor in the major human pathogen Aspergillus fumigatus, is also biosynthesized by its nonpathogenic relative Aspergillus fischeri. mBio 2020, 11, e03361-19. [Google Scholar] [CrossRef] [Green Version]
- Bok, J.W.; Chung, D.; Balajee, S.A.; Marr, K.A.; Andes, D.; Nielsen, K.F.; Frisvad, J.C.; Kirby, K.A.; Keller, N.P. GliZ, a transcriptional regulator of gliotoxin biosynthesis, contributes to Aspergillus fumigatus virulence. Infect. Immun. 2006, 74, 6761–6768. [Google Scholar] [CrossRef] [Green Version]
- Schrettl, M.; Carberry, S.; Kavanagh, K.; Haas, H.; Jones, G.W.; O’Brien, J.; Nolan, A.; Stephens, J.; Fenelon, O.; Doyle, S. Self-protection against gliotoxin-A component of the gliotoxin biosynthetic cluster, GliT, completely protects Aspergillus fumigatus against exogenous gliotoxin. PLoS Pathog. 2010, 6, e1000952. [Google Scholar] [CrossRef]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Hicks, J.K.; Huang, T.P.; Keller, N.P. Pka, Ras and RGS protein interactions regulate activity of AflR, a Zn(II)2Cys6 transcription factor in Aspergillus nidulans. Genetics 2003, 165, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- McCarty, N.S.; Graham, A.E.; Studena, L.; Ledesma-Amaro, R. Multiplexed CRISPR technologies for gene editing and transcriptional regulation. Nat. Commun. 2020, 11, 1281. [Google Scholar] [CrossRef] [PubMed]
- Tak, Y.E.; Kleinstiver, B.P.; Nunez, J.K.; Hsu, J.Y.; Horng, J.E.; Gong, J.Y.; Weissman, J.S.; Joung, J.K. Inducible and multiplex gene regulation using CRISPR-Cpf1-based transcription factors. Nat. Methods 2017, 14, 1163. [Google Scholar] [CrossRef] [Green Version]
- Castellanos, M.; Mothi, N.; Munoz, V. Eukaryotic transcription factors can track and control their target genes using DNA antennas. Nat. Commun. 2020, 11, 540. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SM | Backbone Gene | Backbone Gene Type | Gene Number | PSTF Number | PSTF | PSTF Type | Reference |
|---|---|---|---|---|---|---|---|
| 2,4-dihydroxy-3-methyl-6-(2-oxopropyl)benzaldehyde (DHMBA)/felinone A | dbaI/pkeA (AN7903) | NR-PKS | 9 | 2 | dbaA, dbaG | Zn(II)2Cys6, no conserved domain 1 | [39,40] |
| 6-hydroxy-7-methyl-3-nonylisoquinoline-5,8-dione | pkiA (AN3386) | NR-PKS | 3 | 0 | / | / | [36] |
| Alternariol/isocoumarins | pkgA (AN7071) | NR-PKS | 2 | 0 | / | / | [36] |
| Aspercryptins | atnA (AN7884), pkbA (AN6448) | NRPS, NR-PKS | 14 + 7 2 | 1 + 1 3 | atnN, cicD | Zn(II)2Cys6, Myb-like DNA-binding domain | [41,42] |
| Asperfuranone | afoE (AN1034), afoG (AN1036) | NR-PKS, HR-PKS | 7 | 1 + 1 3 | afoA, scpR | Zn(II)2Cys6, C2H2 type zinc finger | [43,44] |
| (+)-Asperlin | alnA (AN11191) | HR-PKS | 10 | 1 | alnR | Zn(II)2Cys6 | [45] |
| Aspernidine A | pkfA (AN3230) | NR-PKS | 6 | 0 | / | / | [46] |
| Asperniduglene A1 and A2 | sdgA/pkjA (AN1784) | HR-PKS | 4 | 0 | / | / | [47] |
| Asperthecin | aptA (AN6000) | NR-PKS | 3 | 0 | / | / | [48] |
| Aspyridone A and B | apdA (AN8412) | PKS/NRPS hybrid | 8 | 1 | apdR | Zn(II)2Cys6 | [49] |
| Austinol/dehydroaustinol | ausA (AN8383) | NR-PKS | 4 + 10 2 | 0 | / | / | [50] |
| Cichorine | pkbA (AN6448) | NR-PKS | 7 | 1 | cicD | Myb-like DNA-binding domain | [51] |
| Echinocandin B | aniA | NRPS | 12 | 0 | / | / | [52] |
| Emericellamides | easA (AN2545), easB (AN2547) | NRPS, HR-PKS | 4 | 0 | / | / | [53] |
| ent-pimara-8(14),15-diene | AN1594 | TC | 7 | 1 | pbcR | Zn(II)2Cys6 | [54] |
| F-9775 A and B/violaceol I and II/orsellinic acid | orsA (AN7909) | NR-PKS | 3 | 0 | / | / | [55,56] |
| Fellutamide B | inpA (AN3495), inpB (AN3496) | NRPS, NRPS | 6 | 1 | scpR | C2H2 type zinc finger | [31] |
| Ferricrocin | sidC (AN0607) | NRPS | 3 | 0 | / | / | [57] |
| Grey-brown conidiophore pigment | ivoA (AN10576) | NRPS | 2 + 1 2 | 0 | / | / | [58] |
| 4′-Methoxyviridicatin | asqK (AN9226) | NRPS | 14 | 1 | asqA | Fungal specific transcription factor domain 4 | [59] |
| Microperfuranone/dehydromicroperfuranone | micA (AN3396) | NRPS-like | 3 | 0 | / | / | [60] |
| Monodictyphenone, emodin, Xanthones, Arugosin A and H, Sanghaspirodins A and B | mdpG (AN0150) | NR-PKS | 1 + 2 + 10 2 | 2 | mdpE, mdpA | Zn(II)2Cys6, no conserved domain 1 | [23,61,62,63] |
| Nidulanin A | nlsA (AN1242) | NRPS | 1 + 1 2 | 0 | / | / | [64] |
| Penicillin | acvA (AN2621) | NRPS | 3 | 0 | / | / | [65] |
| Sterigmatocystin | stcA/pksST (AN7825) | NR-PKS | 25 | 2 | aflR, aflS/aflJ | Zn(II)2Cys6, no conserved domain 1 | [37,64] |
| Terrequinone A | tdiA (AN8513) | NRPS-like | 5 | 0 | / | / | [66] |
| Viridicatumtoxin 5 | vrtA | NR-PKS | 13 | 2 | vrtR1, vrtR2 | Fungal specific transcription factor domain 4, Zn(II)2Cys6 | [67,68] |
| YWA1 | wA (AN8209) | NR-PKS | 2 | 0 | / | / | [69] |
| SM | Backbone Gene | Backbone Gene Type | Gene Number | PSTF Number | PSTF | PSTF Type | Reference |
|---|---|---|---|---|---|---|---|
| DHN-melanin 1 | pksP/alb1 (Afu2g17600) | NR-PKS | 6 | 0 | / | / | [70,71] |
| Endocrocin/trypacidin | encA (Afu4g00210), tpcC (Afu4g14560) | NR-PKS, NR-PKS | 4 + 13 2 | 2 | tpcE, tpcD | Zn(II)2Cys6, no conserved domain 3 | [72] |
| Ferricrocin/TAFC 4 | sidC (Afu1g17200), sidD (Afu3g03420) | NRPS, NRPS | 1 + 1 + 3 2 | 0 | / | / | [73,74] |
| Fumagillin | fmaB/fma-PKS (Afu8g00370) | HR-PKS | 10 | 1 | fapR/fumR | Zn(II)2Cys6 | [75,76,77] |
| Fumigaclavine C | fgaPT1 (Afu2g17990), pes1/pesB (Afu1g10380), pesL/fqzC (Afu6g12050) | DMATS, NRPS, NRPS | 11 + 1 + 1 2 | 0 | / | / | [32,33] |
| Fumigermin | fgnA (Afu1g01010) | PR-PKS | 5 | 0 | / | / | [28] |
| Fumihopaside A and B | afumA (AFUB_071550) | TC | 4 | 1 | afumD | Zn(II)2Cys6 | [29] |
| Fumiquinazolines | pesM (Afu6g12080) | NRPS | 4 | 0 | / | / | [33,78] |
| Fumisoquins/fumipyrrole | fsqF/fmpE (Afu6g03480) | NRPS-like | 7 | 1 | fsqA/fmpR | Zn(II)2Cys6 | [79,80] |
| Fumitremorgin/brevianamide F | ftmA (Afu8g00170) | NRPS | 9 | 0 | / | / | [81,82] |
| Gliotoxin | gliP (Afu6g09660) | NRPS | 12 + 1 + 1 2 | 1 + 1 5 | gliZ, rglT | Zn(II)2Cys6, Zn(II)2Cys6 | [83,84,85,86] |
| Helvolic acid | helA (Afu4g14770) | TC | 9 | 0 | / | / | [87] |
| Hexadehydroastechrome | hasD/pesF (Afu3g12920), hasE (Afu3g12930) | NRPS, DMATS | 8 | 2 | hasA, hasF | Zn(II)2Cys6, Zn(II)2Cys6 | [88] |
| Neosartoricin/fumicyclines | nscA/fccA (Afu7g00160) | NRPS | 6 | 1 | nscR | Zn(II)2Cys6 | [89,90,91] |
| Psecurotin A | posA (Afu8g00540) | PKS/NRPS hybrid | 6 | 1 | fapR/fumR | Zn(II)2Cys6 | [77,92] |
| Pyomelanin | hppD (Afu2g04200) 6 | / | 6 | 1 | hmgR | Zn(II)2Cys6 | [30,93] |
| Pyripyropene A | pyr2 (Afu6g13930) | HR-PKS | 8 | 0 | / | / | [94] |
| Xanthocillin | xanB (Afu5g02660) | ICS | 6 | 1 | xanC | bZIP | [16] |
| BGCs | PSTF-Containing BGCs | |
|---|---|---|
| Aspergillus nidulans | ||
| In total | 28 | 12 |
| Genes ≤ 5 | 13 | 0 |
| Genes > 5 | 15 | 12 |
| Aspergillus fumigatus | ||
| In total | 18 | 10 |
| Genes ≤ 5 | 4 | 1 |
| Genes > 5 | 14 | 9 |
| Case Type | Aspergillus nidulans | Aspergillus fumigatus | Case in Total |
|---|---|---|---|
| One PSTF per BGC | 6 (6 PSTFs + 6 BGCs) 1 | 5 (5 PSTFs + 5 BGCs) 1 | 11 |
| Two PSTFs per BGC | 4 (8 PSTFs + 4 BGCs) 1 | 2 (4 PSTFs + 2 BGCs) 1 | 6 |
| One PSTF for two BGCs | / | 1 (1 PSTF + 2 BGCs) 1 | 1 |
| Additional case 2 | 1 (2 PSTFs + 2 BGCs) 1 | 1 (2 PSTFs + 1 BGC) 1 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Yu, Y.; Keller, N.P.; Wang, P. Presence, Mode of Action, and Application of Pathway Specific Transcription Factors in Aspergillus Biosynthetic Gene Clusters. Int. J. Mol. Sci. 2021, 22, 8709. https://doi.org/10.3390/ijms22168709
Wang W, Yu Y, Keller NP, Wang P. Presence, Mode of Action, and Application of Pathway Specific Transcription Factors in Aspergillus Biosynthetic Gene Clusters. International Journal of Molecular Sciences. 2021; 22(16):8709. https://doi.org/10.3390/ijms22168709
Chicago/Turabian StyleWang, Wenjie, Yuchao Yu, Nancy P. Keller, and Pinmei Wang. 2021. "Presence, Mode of Action, and Application of Pathway Specific Transcription Factors in Aspergillus Biosynthetic Gene Clusters" International Journal of Molecular Sciences 22, no. 16: 8709. https://doi.org/10.3390/ijms22168709
APA StyleWang, W., Yu, Y., Keller, N. P., & Wang, P. (2021). Presence, Mode of Action, and Application of Pathway Specific Transcription Factors in Aspergillus Biosynthetic Gene Clusters. International Journal of Molecular Sciences, 22(16), 8709. https://doi.org/10.3390/ijms22168709