
Pivotal Role of Corneal Fibroblasts in Progression to Corneal Ulcer in Bacterial Keratitis


Abstract
:1. Introduction
2. Onset and Clinical Course of Bacterial Keratitis
3. Culture Model for Measurement of Collagen Degradation by Corneal Fibroblasts
4. Molecular Mechanisms of Corneal Ulceration
4.1. Direct Collagen Destruction by Factors Released from Bacteria
4.2. Cell-Dependent Collagen Destruction
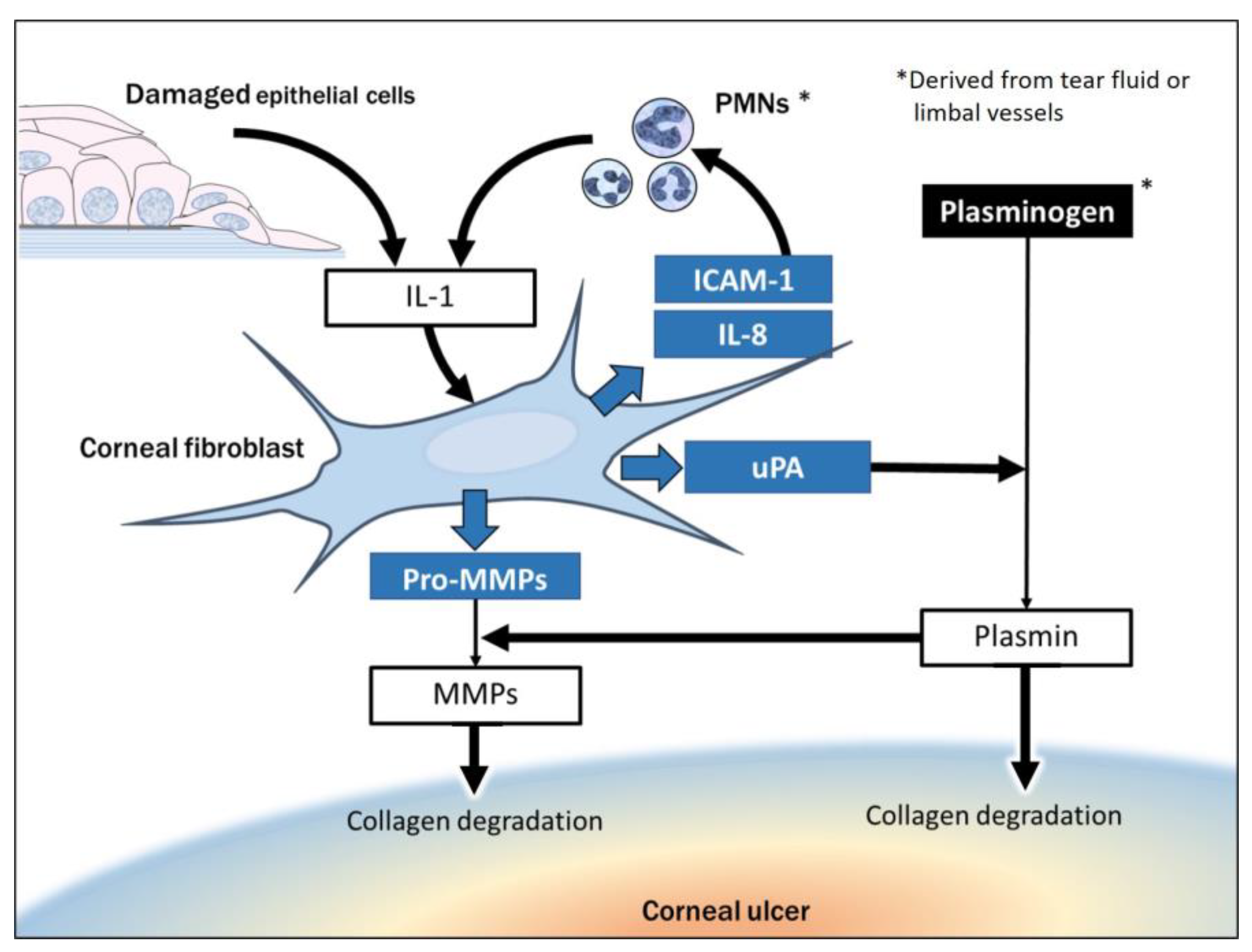
4.2.1. Collagen Degradation Resulting from Exposure of Corneal Fibroblasts to Alarmins
4.2.2. Collagen Degradation Due to Interaction of Corneal Fibroblasts with Infiltrated PMNs
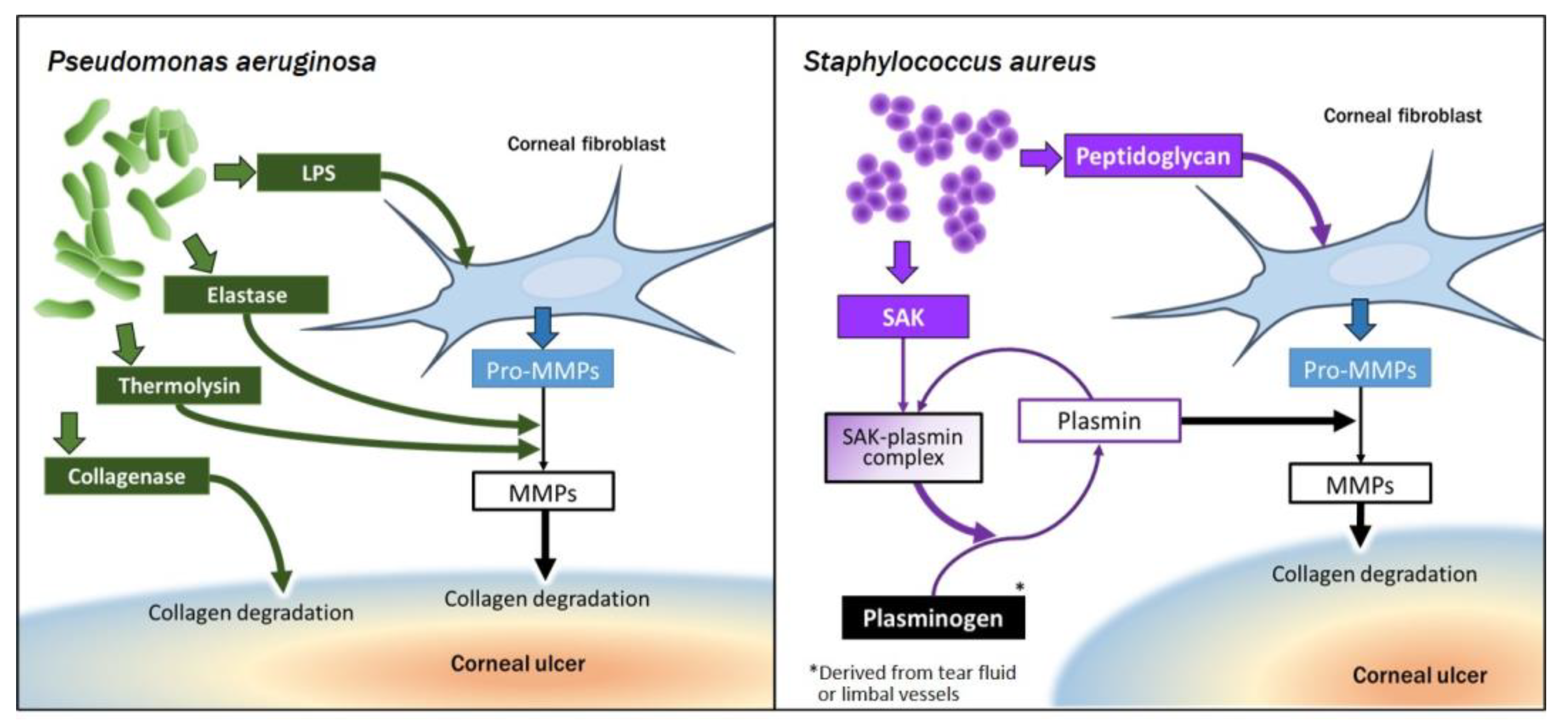
4.2.3. Collagen Degradation Resulting from Interaction of Corneal Fibroblasts with Bacteria
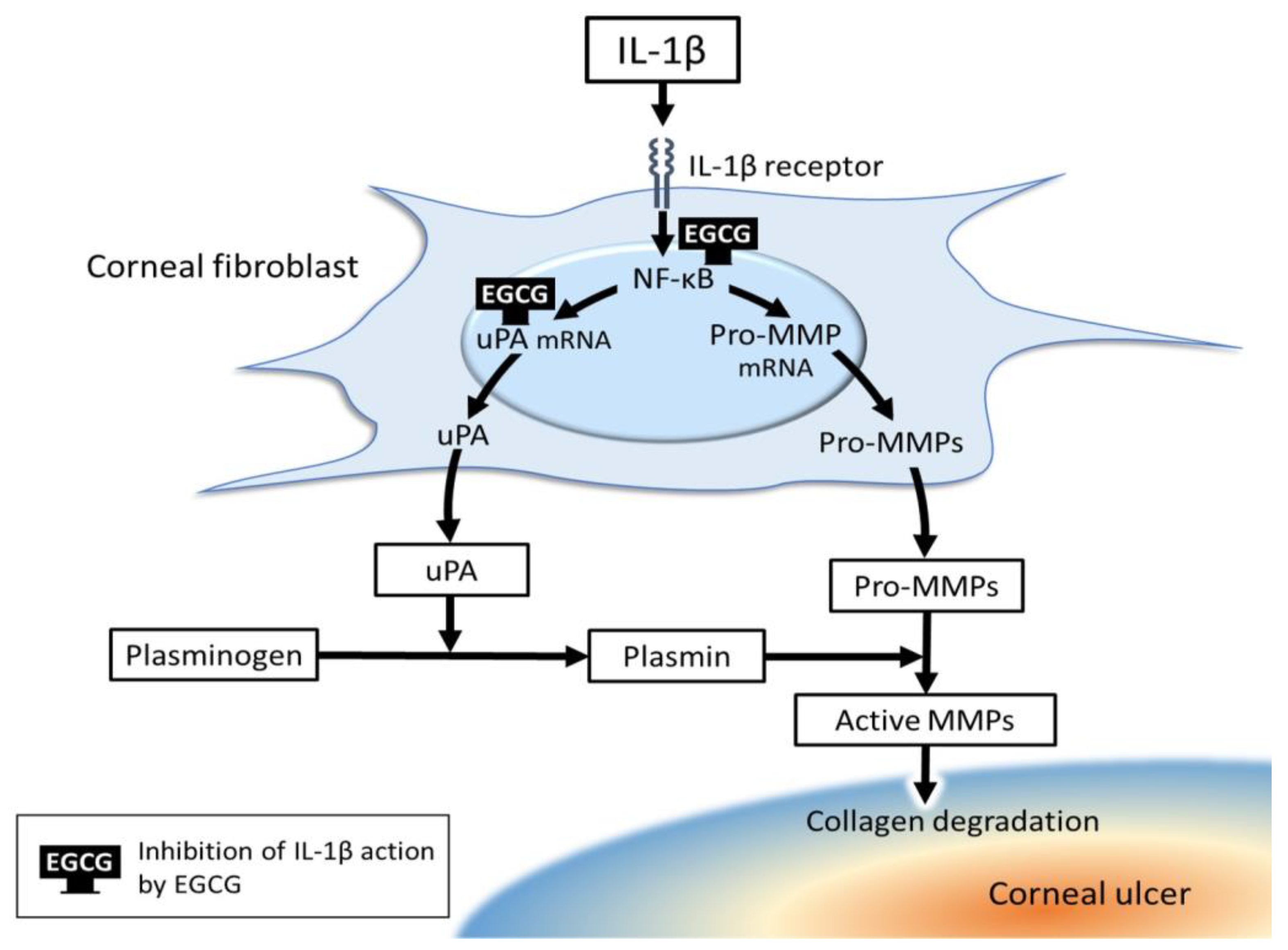
5. Exploration of New Medical Treatment Modalities for Corneal Ulcer
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barabino, S.; Chen, Y.; Chauhan, S.; Dana, R. Ocular surface immunity: Homeostatic mechanisms and their disruption in dry eye disease. Prog. Retin. Eye Res. 2012, 31, 271–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, Y.; Dogru, M.; Tsubota, K. Laboratory findings in tear fluid analysis. Clin. Chim. Acta 2006, 369, 17–28. [Google Scholar] [CrossRef]
- Seal, D.V.; McGill, J.I.; Mackie, I.A.; Liakos, G.M.; Jacobs, P.; Goulding, N.J. Bacteriology and tear protein profiles of the dry eye. Br. J. Ophthalmol. 1986, 70, 122–125. [Google Scholar] [CrossRef] [Green Version]
- Nishida, T. Future trends in the treatment of dry eye disease. In Dry Eye Disease: The Clinician’s Guide to Diagnosis and Treatment; Asbell, P., Lemp, M., Eds.; Thieme Medical Publishers: New York, NY, USA, 2006; pp. 192–197. [Google Scholar]
- Flanagan, J.L.; Willcox, M.D. Role of lactoferrin in the tear film. Biochimie 2009, 91, 35–43. [Google Scholar] [CrossRef]
- Augustin, D.K.; Heimer, S.R.; Tam, C.; Li, W.Y.; Le Due, J.M.; Evans, D.J.; Fleiszig, S.M. Role of defensins in corneal epithelial barrier function against Pseudomonas aeruginosa traversal. Infect. Immun. 2011, 79, 595–605. [Google Scholar] [CrossRef] [Green Version]
- McDermott, A.M. Antimicrobial compounds in tears. Exp. Eye Res. 2013, 117, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Di Zazzo, A.; Micera, A.; Coassin, M.; Varacalli, G.; Foulsham, W.; De Piano, M.; Bonini, S. InflammAging at ocular surface: Clinical and biomolecular analyses in healthy volunteers. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1769–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulsham, W.; Coco, G.; Amouzegar, A.; Chauhan, S.K.; Dana, R. When Clarity Is Crucial: Regulating Ocular Surface Immunity. Trends Immunol. 2018, 39, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, I.; Said, D.G.; Dua, H.S. Human antimicrobial peptides in ocular surface defense. Prog. Retin. Eye Res. 2017, 61, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Lavker, R.M.; Kaplan, N.; Wang, J.; Peng, H. Corneal epithelial biology: Lessons stemming from old to new. Exp. Eye Res. 2020, 198, 108094. [Google Scholar] [CrossRef]
- Sosnova-Netukova, M.; Kuchynka, P.; Forrester, J.V. The suprabasal layer of corneal epithelial cells represents the major barrier site to the passive movement of small molecules and trafficking leukocytes. Br. J. Ophthalmol. 2007, 91, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Harhaj, N.S.; Antonetti, D.A. Regulation of tight junctions and loss of barrier function in pathophysiology. Int. J. Biochem. Cell Biol. 2004, 36, 1206–1237. [Google Scholar] [CrossRef]
- Suzuki, K.; Saito, J.; Yanai, R.; Yamada, N.; Chikama, T.; Seki, K.; Nishida, T. Cell-matrix and cell-cell interactions during corneal epithelial wound healing. Prog. Retin. Eye Res. 2003, 22, 113–133. [Google Scholar] [CrossRef]
- Kalinin, A.E.; Kajava, A.V.; Steinert, P.M. Epithelial barrier function: Assembly and structural features of the cornified cell envelope. Bioessays 2002, 24, 789–800. [Google Scholar] [CrossRef]
- Kinoshita, S.; Adachi, W.; Sotozono, C.; Nishida, K.; Yokoi, N.; Quantock, A.J.; Okubo, K. Characteristics of the human ocular surface epithelium. Prog. Retin. Eye Res. 2001, 20, 639–673. [Google Scholar] [CrossRef]
- Hilliam, Y.; Kaye, S.; Winstanley, C. Pseudomonas aeruginosa and microbial keratitis. J. Med. Microbiol. 2020, 69, 3–13. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, R.; Caballero, A.; Tang, A.; Bierdeman, M. Pseudomonas aeruginosa Keratitis: Protease IV and PASP as Corneal Virulence Mediators. Microorganisms 2019, 7, 281. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Somerville, T.; Kaye, S.B.; Romano, V. Staphylococcus aureus Keratitis: Incidence, Pathophysiology, Risk Factors and Novel Strategies for Treatment. J. Clin. Med. 2021, 10, 758. [Google Scholar] [CrossRef]
- Ting, D.S.J.; Ho, C.S.; Deshmukh, R.; Said, D.G.; Dua, H.S. Infectious keratitis: An update on epidemiology, causative microorganisms, risk factors, and antimicrobial resistance. Eye 2021, 35, 1084–1101. [Google Scholar] [CrossRef]
- Jeng, B.H.; Gritz, D.C.; Kumar, A.B.; Holsclaw, D.S.; Porco, T.C.; Smith, S.D.; Whitcher, J.P.; Margolis, T.P.; Wong, I.G. Epidemiology of ulcerative keratitis in Northern California. Arch. Ophthalmol. 2010, 128, 1022–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.J.; Huang, Y.C.; Tan, H.Y.; Ma, D.H.; Lin, H.C.; Yeh, L.K.; Chen, P.Y.; Chen, H.C.; Chuang, C.C.; Chang, C.J.; et al. Staphylococcus aureus keratitis: A review of hospital cases. PLoS ONE 2013, 8, e80119. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, L.D. Bacterial infections of the cornea (Pseudomonas aeruginosa). Chem. Immunol. Allergy 2007, 92, 185–194. [Google Scholar] [CrossRef]
- Bourcier, T.; Thomas, F.; Borderie, V.; Chaumeil, C.; Laroche, L. Bacterial keratitis: Predisposing factors, clinical and microbiological review of 300 cases. Br. J. Ophthalmol. 2003, 87, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Pachigolla, G.; Blomquist, P.; Cavanagh, H.D. Microbial keratitis pathogens and antibiotic susceptibilities: A 5-year review of cases at an urban county hospital in north Texas. Eye Contact Lens 2007, 33, 45–49. [Google Scholar] [CrossRef]
- Green, M.; Apel, A.; Stapleton, F. Risk factors and causative organisms in microbial keratitis. Cornea 2008, 27, 22–27. [Google Scholar] [CrossRef]
- Hong, A.R.; Zao, H.; Huang, A.J.W. Bacterial keratitis. In Cornea: Fundamentals, Diagnosis and Management, 5th ed.; Mannis, M.J., Holland, E.J., Eds.; Elsevier: London, UK, 2018; Volume 1, pp. 802–824. [Google Scholar]
- Ung, L.; Chodosh, J. Foundational concepts in the biology of bacterial keratitis. Exp. Eye Res. 2021, 209, 108647. [Google Scholar] [CrossRef]
- Shrestha, G.S.; Vijay, A.K.; Stapleton, F.; Henriquez, F.L.; Carnt, N. Understanding clinical and immunological features associated with Pseudomonas and Staphylococcus keratitis. Cont. Lens Anterior Eye 2021, 44, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Fleiszig, S.M.J.; Kroken, A.R.; Nieto, V.; Grosser, M.R.; Wan, S.J.; Metruccio, M.M.E.; Evans, D.J. Contact lens-related corneal infection: Intrinsic resistance and its compromise. Prog. Retin. Eye Res. 2020, 76, 100804. [Google Scholar] [CrossRef]
- O’Callaghan, R.J. The Pathogenesis of Staphylococcus aureus Eye Infections. Pathogens 2018, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Hazlett, L.D. Corneal response to Pseudomonas aeruginosa infection. Prog. Retin. Eye Res. 2004, 23, 1–30. [Google Scholar] [CrossRef]
- Nishida, T. Commanding roles of keratocytes in health and disease. Cornea 2010, 29, S3–S6. [Google Scholar] [CrossRef]
- Fukuda, K.; Ishida, W.; Fukushima, A.; Nishida, T. Corneal Fibroblasts as Sentinel Cells and Local Immune Modulators in Infectious Keratitis. Int. J. Mol. Sci. 2017, 18, 1831. [Google Scholar] [CrossRef]
- Fukuda, K. Corneal fibroblasts: Function and markers. Exp. Eye Res. 2020, 200, 108229. [Google Scholar] [CrossRef]
- Nishida, T. Inflammation of the conjunctiva. In Encyclopedia of the Eye; Dartt, D.A., Ed.; Academic Press: Oxford, UK, 2010; pp. 324–329. [Google Scholar]
- Fukuda, K.; Ishida, W.; Miura, Y.; Kishimoto, T.; Fukushima, A. Cytokine expression and barrier disruption in human corneal epithelial cells induced by alarmin released from necrotic cells. Jpn. J. Ophthalmol. 2017, 61, 415–422. [Google Scholar] [CrossRef]
- Kimura, K.; Teranishi, S.; Nishida, T. Interleukin-1beta-induced disruption of barrier function in cultured human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2009, 50, 597–603. [Google Scholar] [CrossRef]
- Kimura, K.; Teranishi, S.; Fukuda, K.; Kawamoto, K.; Nishida, T. Delayed Disruption of Barrier Function in Cultured Human Corneal Epithelial Cells Induced by Tumor Necrosis Factor-α in a Manner Dependent on NF-κB. Investig. Ophthalmol. Vis. Sci. 2008, 49, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Yu, F.S. Toll-like receptors and corneal innate immunity. Curr. Mol. Med. 2006, 6, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Kumar, H.; Kawai, T.; Akira, S. Toll-like receptors and innate immunity. Biochem. Biophys. Res. Commun. 2009, 388, 621–625. [Google Scholar] [CrossRef]
- Lambiase, A.; Micera, A.; Sacchetti, M.; Mantelli, F.; Bonini, S. Toll-like receptors in ocular surface diseases: Overview and new findings. Clin. Sci. 2011, 120, 441–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, K.; Kumagai, N.; Nishida, T. Levels of soluble CD14 and lipopolysaccharide-binding protein in human basal tears. Jpn. J. Ophthalmol. 2010, 54, 241–242. [Google Scholar] [CrossRef]
- Fukuda, K.; Kumagai, N.; Yamamoto, K.; Fujitsu, Y.; Chikamoto, N.; Nishida, T. Potentiation of lipopolysaccharide-induced chemokine and adhesion molecule expression in corneal fibroblasts by soluble CD14 or LPS-binding protein. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3095–3101. [Google Scholar] [CrossRef]
- Kumagai, N.; Fukuda, K.; Fujitsu, Y.; Lu, Y.; Chikamoto, N.; Nishida, T. Lipopolysaccharide-induced expression of intercellular adhesion molecule-1 and chemokines in cultured human corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2005, 46, 114–120. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, Y.; Fukuda, K.; Nakamura, Y.; Kumagai, N.; Nishida, T. Inhibition by triptolide of chemokine, proinflammatory cytokine, and adhesion molecule expression induced by lipopolysaccharide in corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3796–3800. [Google Scholar] [CrossRef] [Green Version]
- Livingston, E.T.; Mursalin, M.H.; Callegan, M.C. A Pyrrhic Victory: The PMN Response to Ocular Bacterial Infections. Microorganisms 2019, 7, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, M.B. Regulation of corneal fibroblast MMP-1 collagenase secretion by plasmin. Cornea 1993, 12, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Fini, M.E.; Cook, J.R.; Mohan, R. Proteolytic mechanisms in corneal ulceration and repair. Arch. Dermatol. Res. 1998, 290, S12–S23. [Google Scholar] [CrossRef] [PubMed]
- Sugioka, K.; Mishima, H.; Kodama, A.; Itahashi, M.; Fukuda, M.; Shimomura, Y. Regulatory Mechanism of Collagen Degradation by Keratocytes and Corneal Inflammation: The Role of Urokinase-Type Plasminogen Activator. Cornea 2016, 35 (Suppl. 1), S59–S64. [Google Scholar] [CrossRef]
- Li, Q.; Fukuda, K.; Lu, Y.; Nakamura, Y.; Chikama, T.; Kumagai, N.; Nishida, T. Enhancement by neutrophils of collagen degradation by corneal fibroblasts. J. Leukoc. Biol. 2003, 74, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Nishida, T.; Yasumoto, K.; Otori, T.; Desaki, J. The network structure of corneal fibroblasts in the rat as revealed by scanning electron microscopy. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1887–1890. [Google Scholar]
- Ueda, A.; Nishida, T.; Otori, T.; Fujita, H. Electron-microscopic studies on the presence of gap junctions between corneal fibroblasts in rabbits. Cell Tissue Res. 1987, 249, 473–475. [Google Scholar] [CrossRef] [PubMed]
- Nishida, T.; Ueda, A.; Fukuda, M.; Mishima, H.; Yasumoto, K.; Otori, T. Interactions of extracellular collagen and corneal fibroblasts: Morphologic and biochemical changes of rabbit corneal cells cultured in a collagen matrix. In Vitro Cell. Dev. Biol. 1988, 24, 1009–1014. [Google Scholar] [CrossRef]
- Sugioka, K.; Kodama-Takahashi, A.; Yoshida, K.; Aomatsu, K.; Okada, K.; Nishida, T.; Shimomura, Y. Extracellular Collagen Promotes Interleukin-1beta-Induced Urokinase-Type Plasminogen Activator Production by Human Corneal Fibroblasts. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1487–1498. [Google Scholar] [CrossRef] [PubMed]
- Quan, W.Y.; Ko, J.A.; Yanai, R.; Nakamura, Y.; Nishida, T. Integrin-mediated inhibition of interleukin-8 secretion from human neutrophils by collagen type I. J. Leukoc. Biol. 2010, 87, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Mishima, H.; Okamoto, J.; Nakamura, M.; Wada, Y.; Otori, T. Collagenolytic activity of keratocytes cultured in a collagen matrix. Jpn. J. Ophthalmol. 1998, 42, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.L.; Nagano, T.; Nakamura, M.; Kumagai, N.; Mishima, H.; Nishida, T. Effect of galardin on collagen degradation by Pseudomonas aeruginosa. Exp. Eye Res. 1999, 69, 595–601. [Google Scholar] [CrossRef]
- Mishima, H.; Ueda, A.; Nishida, T.; Otori, T. Increased lysosomal enzyme activity of keratocytes after endocytosis of foreign particles. Jpn. J. Ophthalmol. 1992, 36, 84–90. [Google Scholar]
- Madsen, D.H.; Jurgensen, H.J.; Siersbaek, M.S.; Kuczek, D.E.; Grey Cloud, L.; Liu, S.; Behrendt, N.; Grontved, L.; Weigert, R.; Bugge, T.H. Tumor-Associated Macrophages Derived from Circulating Inflammatory Monocytes Degrade Collagen through Cellular Uptake. Cell Rep. 2017, 21, 3662–3671. [Google Scholar] [CrossRef] [Green Version]
- Liesegang, T.J. Bacterial and fungal keratitis. In The Cornea; Kaufman, H.E., Barron, B.A., McDonald, M.B., Waltman, S.R., Eds.; Churchill Livingstone: New York, NY, USA, 1988; pp. 217–270. [Google Scholar]
- Zhang, Y.Z.; Ran, L.Y.; Li, C.Y.; Chen, X.L. Diversity, Structures, and Collagen-Degrading Mechanisms of Bacterial Collagenolytic Proteases. Appl. Environ. Microbiol. 2015, 81, 6098–6107. [Google Scholar] [CrossRef] [Green Version]
- Duarte, A.S.; Correia, A.; Esteves, A.C. Bacterial collagenases—A review. Crit. Rev. Microbiol. 2016, 42, 106–126. [Google Scholar] [CrossRef]
- Liu, P.V. Extracellular toxins of Pseudomonas aeruginosa. J. Infect. Dis. 1974, 130, S94–S99. [Google Scholar] [CrossRef]
- Twining, S.S.; Lohr, K.M.; Moulder, J.E. The immune system in experimental Pseudomonas keratitis. Model and early effects. Investig. Ophthalmol. Vis. Sci. 1986, 27, 507–515. [Google Scholar]
- Ohbayashi, T.; Irie, A.; Murakami, Y.; Nowak, M.; Potempa, J.; Nishimura, Y.; Shinohara, M.; Imamura, T. Degradation of fibrinogen and collagen by staphopains, cysteine proteases released from Staphylococcus aureus. Microbiology 2011, 157, 786–792. [Google Scholar] [CrossRef] [Green Version]
- Sugioka, K.; Kodama-Takahshi, A.; Sato, T.; Okada, K.; Murakami, J.; Park, A.M.; Mishima, H.; Shimomura, Y.; Kusaka, S.; Nishida, T. Plasminogen-Dependent Collagenolytic Properties of Staphylococcus aureus in Collagen Gel Cultures of Human Corneal Fibroblasts. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5098–5107. [Google Scholar] [CrossRef]
- Loffek, S.; Schilling, O.; Franzke, C.W. Series “matrix metalloproteinases in lung health and disease”: Biological role of matrix metalloproteinases: A critical balance. Eur. Respir. J. 2011, 38, 191–208. [Google Scholar] [CrossRef] [Green Version]
- Jablonska-Trypuc, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzym. Inhib. Med. Chem. 2016, 31, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilgus, T.A. Alerting the body to tissue injury: The role of alarmins and DAMPs in cutaneous wound healing. Curr. Pathobiol. Rep. 2018, 6, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Iwatake, A.; Murakami, A.; Ebihara, N. The expression of matrix metalloproteinases and their inhibitors in corneal fibroblasts by alarmins from necrotic corneal epithelial cells. Jpn. J. Ophthalmol. 2018, 62, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Takahata, Y.; Hata, K.; Nishimura, R. Role of interleukin-1 and inflammasomes in oral disease. J. Oral Biosci. 2020, 62, 242–248. [Google Scholar] [CrossRef]
- Kitanaka, N.; Nakano, R.; Sakai, M.; Kitanaka, T.; Namba, S.; Konno, T.; Nakayama, T.; Sugiya, H. ERK1/ATF-2 signaling axis contributes to interleukin-1beta-induced MMP-3 expression in dermal fibroblasts. PLoS ONE 2019, 14, e0222869. [Google Scholar] [CrossRef]
- Xie, Z.; Singh, M.; Singh, K. Differential regulation of matrix metalloproteinase-2 and -9 expression and activity in adult rat cardiac fibroblasts in response to interleukin-1beta. J. Biol. Chem. 2004, 279, 39513–39519. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, F.; Fan, C.; Wang, C.; Ruan, H. Effects and relationship of ERK1 and ERK2 in interleukin-1beta-induced alterations in MMP3, MMP13, type II collagen and aggrecan expression in human chondrocytes. Int. J. Mol. Med. 2011, 27, 583–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.S.; Lin, C.Y.; Chiu, Y.H.; Chen, C.P.; Tsai, P.J.; Wang, H.S. IL-1beta-Induced Matrix Metalloprotease-1 Promotes Mesenchymal Stem Cell Migration via PAR1 and G-Protein-Coupled Signaling Pathway. Stem Cells Int. 2018, 2018, 3524759. [Google Scholar] [CrossRef] [Green Version]
- Fini, M.E.; Strissel, K.J.; Girard, M.T.; Mays, J.W.; Rinehart, W.B. Interleukin 1 alpha mediates collagenase synthesis stimulated by phorbol 12-myristate 13-acetate. J. Biol. Chem. 1994, 269, 11291–11298. [Google Scholar] [CrossRef]
- Hao, J.L.; Nagano, T.; Nakamura, M.; Kumagai, N.; Mishima, H.; Nishida, T. Galardin inhibits collagen degradation by rabbit keratocytes by inhibiting the activation of pro-matrix metalloproteinases. Exp. Eye Res. 1999, 68, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Mountain, D.J.; Singh, M.; Menon, B.; Singh, K. Interleukin-1beta increases expression and activity of matrix metalloproteinase-2 in cardiac microvascular endothelial cells: Role of PKCalpha/beta1 and MAPKs. Am. J. Physiol. Cell Physiol. 2007, 292, C867–C875. [Google Scholar] [CrossRef] [Green Version]
- Liang, K.C.; Lee, C.W.; Lin, W.N.; Lin, C.C.; Wu, C.B.; Luo, S.F.; Yang, C.M. Interleukin-1beta induces MMP-9 expression via p42/p44 MAPK, p38 MAPK, JNK, and nuclear factor-kappaB signaling pathways in human tracheal smooth muscle cells. J. Cell. Physiol. 2007, 211, 759–770. [Google Scholar] [CrossRef]
- Lu, Y.; Fukuda, K.; Seki, K.; Nakamura, Y.; Kumagai, N.; Nishida, T. Inhibition by triptolide of IL-1-induced collagen degradation by corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2003, 44, 5082–5088. [Google Scholar] [CrossRef] [Green Version]
- Rudner, X.L.; Kernacki, K.A.; Barrett, R.P.; Hazlett, L.D. Prolonged elevation of IL-1 in Pseudomonas aeruginosa ocular infection regulates macrophage-inflammatory protein-2 production, polymorphonuclear neutrophil persistence, and corneal perforation. J. Immunol. 2000, 164, 6576–6582. [Google Scholar] [CrossRef] [Green Version]
- Hobden, J.A.; Masinick, S.A.; Barrett, R.P.; Hazlett, L.D. Aged mice fail to upregulate ICAM-1 after Pseudomonas aeruginosa corneal infection. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1107–1114. [Google Scholar]
- Gagen, D.; Laubinger, S.; Li, Z.; Petrescu, M.S.; Brown, E.S.; Smith, C.W.; Burns, A.R. ICAM-1 mediates surface contact between neutrophils and keratocytes following corneal epithelial abrasion in the mouse. Exp. Eye Res. 2010, 91, 676–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, G.; Zhao, X.; Zhang, L.; Zhang, J.; L’Huillier, A.; Ling, W.; Roberts, A.I.; Le, A.D.; Shi, S.; Shao, C.; et al. Inflammatory cytokine-induced intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 in mesenchymal stem cells are critical for immunosuppression. J. Immunol. 2010, 184, 2321–2328. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.; Sethu, C.; Louafi, F.; Hossain, P. Lipopolysaccharide regulation of toll-like receptor-4 and matrix metalloprotease-9 in human primary corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2796–2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Du, W.; McClellan, S.A.; Barrett, R.P.; Hazlett, L.D. TLR4 is required for host resistance in Pseudomonas aeruginosa keratitis. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4910–4916. [Google Scholar] [CrossRef] [Green Version]
- Schultz, C.L.; Morck, D.W.; McKay, S.G.; Olson, M.E.; Buret, A. Lipopolysaccharide induced acute red eye and corneal ulcers. Exp. Eye Res. 1997, 64, 3–9. [Google Scholar] [CrossRef]
- Hurley, J.C. Antibiotic-induced release of endotoxin: A reappraisal. Clin. Infect. Dis. 1992, 15, 840–854. [Google Scholar] [CrossRef]
- Pfister, R.R.; Haddox, J.L.; Lank, K.M. Citrate or ascorbate/citrate treatment of established corneal ulcers in the alkali-injured rabbit eye. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1110–1115. [Google Scholar]
- Burns, F.R.; Paterson, C.A.; Gray, R.D.; Wells, J.T. Inhibition of Pseudomonas aeruginosa elastase and Pseudomonas keratitis using a thiol-based peptide. Antimicrob. Agents Chemother. 1990, 34, 2065–2069. [Google Scholar] [CrossRef] [Green Version]
- Burns, F.R.; Gray, R.D.; Paterson, C.A. Inhibition of alkali-induced corneal ulceration and perforation by a thiol peptide. Investig. Ophthalmol. Vis. Sci. 1990, 31, 107–114. [Google Scholar]
- Brown, S.I.; Weller, C.A. Collagenase inhibitors in prevention of ulcers of alkali-burned cornea. Arch. Ophthalmol. 1970, 83, 352–353. [Google Scholar] [CrossRef]
- Wentworth, J.S.; Paterson, C.A.; Gray, R.D. Effect of a metalloproteinase inhibitor on established corneal ulcers after an alkali burn. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2174–2179. [Google Scholar]
- Brooks, D.E.; Ollivier, F.J. Matrix metalloproteinase inhibition in corneal ulceration. Vet. Clin. N. Am. Small Anim. Pract. 2004, 34, 611–622. [Google Scholar] [CrossRef]
- Li, D.Q.; Chen, Z.; Song, X.J.; Luo, L.; Pflugfelder, S.C. Stimulation of matrix metalloproteinases by hyperosmolarity via a JNK pathway in human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4302–4311. [Google Scholar] [CrossRef]
- Li, D.Q.; Shang, T.Y.; Kim, H.S.; Solomon, A.; Lokeshwar, B.L.; Pflugfelder, S.C. Regulated expression of collagenases MMP-1, -8, and -13 and stromelysins MMP-3, -10, and -11 by human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2928–2936. [Google Scholar] [CrossRef] [Green Version]
- Seedor, J.A.; Perry, H.D.; McNamara, T.F.; Golub, L.M.; Buxton, D.F.; Guthrie, D.S. Systemic tetracycline treatment of alkali-induced corneal ulceration in rabbits. Arch. Ophthalmol. 1987, 105, 268–271. [Google Scholar] [CrossRef]
- Dursun, D.; Kim, M.C.; Solomon, A.; Pflugfelder, S.C. Treatment of recalcitrant recurrent corneal erosions with inhibitors of matrix metalloproteinase-9, doxycycline and corticosteroids. Am. J. Ophthalmol. 2001, 132, 8–13. [Google Scholar] [CrossRef]
- McElvanney, A.M. Doxycycline in the management of pseudomonas corneal melting: Two case reports and a review of the literature. Eye Contact Lens 2003, 29, 258–261. [Google Scholar] [CrossRef]
- Bian, F.; Pelegrino, F.S.; Henriksson, J.T.; Pflugfelder, S.C.; Volpe, E.A.; Li, D.Q.; de Paiva, C.S. Differential Effects of Dexamethasone and Doxycycline on Inflammation and MMP Production in Murine Alkali-Burned Corneas Associated with Dry Eye. Ocul. Surf. 2016, 14, 242–254. [Google Scholar] [CrossRef] [Green Version]
- Hindman, H.B.; Patel, S.B.; Jun, A.S. Rationale for adjunctive topical corticosteroids in bacterial keratitis. Arch. Ophthalmol. 2009, 127, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Kimura, K.; Orita, T.; Nishida, T.; Sonoda, K.H. Inhibition by female sex hormones of collagen degradation by corneal fibroblasts. Mol. Vis. 2011, 17, 3415–3422. [Google Scholar]
- Zhou, H.; Kimura, K.; Orita, T.; Nishida, T.; Sonoda, K.H. Inhibition by medroxyprogesterone acetate of interleukin-1beta-induced collagen degradation by corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4213–4219. [Google Scholar] [CrossRef] [Green Version]
- Acharya, N.R.; Srinivasan, M.; Mascarenhas, J.; Ravindran, M.; Rajaraman, R.; Zegans, M.; McLeod, S.; Lietman, T.M. The steroid controversy in bacterial keratitis. Arch. Ophthalmol. 2009, 127, 1231. [Google Scholar] [CrossRef] [Green Version]
- Tallab, R.T.; Stone, D.U. Corticosteroids as a therapy for bacterial keratitis: An evidence-based review of ‘who, when and why’. Br. J. Ophthalmol. 2016, 100, 731–735. [Google Scholar] [CrossRef]
- Srinivasan, M.; Mascarenhas, J.; Rajaraman, R.; Ravindran, M.; Lalitha, P.; Glidden, D.V.; Ray, K.J.; Hong, K.C.; Oldenburg, C.E.; Lee, S.M.; et al. Corticosteroids for bacterial keratitis: The Steroids for Corneal Ulcers Trial (SCUT). Arch. Ophthalmol. 2012, 130, 143–150. [Google Scholar] [CrossRef]
- Sugioka, K.; Yoshida, K.; Murakami, J.; Itahashi, M.; Mishima, H.; Nishida, T.; Kusaka, S. Inhibition by Epigallocatechin Gallate of IL-1-Induced Urokinase-Type Plasminogen Activator Expression and Collagen Degradation by Corneal Fibroblasts. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2895–2903. [Google Scholar] [CrossRef] [Green Version]
- Brocks, D.; Mead, O.G.; Tighe, S.; Tseng, S.C.G. Self-Retained Cryopreserved Amniotic Membrane for the Management of Corneal Ulcers. Clin. Ophthalmol. 2020, 14, 1437–1443. [Google Scholar] [CrossRef]
- Sheha, H.; Liang, L.; Li, J.; Tseng, S.C. Sutureless amniotic membrane transplantation for severe bacterial keratitis. Cornea 2009, 28, 1118–1123. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Phase | Clinical Manifestation | Principal Players | Pathobiology | Treatment |
|---|---|---|---|---|
| Preceding event | Disruption of epithelial barrier and adhesion of bacteria | Bacteria | Trauma, CL wear, inflammation | Hygiene, proper CL handling, prophylactic administration of antimicrobials |
| Phase 1 | Onset of keratitis | Bacteria (PAMPs), corneal fibroblasts, PMNs | Innate immune responses by corneal fibroblasts | Elimination of bacteria by antimicrobials |
| Phase 2 | Onset of corneal ulceration | Bacteria | Collagen degradation by bacterial proteinases | Elimination of bacteria by antimicrobials. No drugs except steroids available to halt excessive stromal melting |
| Corneal fibroblasts | Cell-mediated collagen degradation by uPA-plasmin-MMP cascade | |||
| Phase 3 | Prolonged corneal melting and perforation of the cornea | Corneal fibroblasts and PMNs | Vicious cycle between PMNs and corneal fibroblasts | Surgery including corneal transplantation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishida, T.; Sugioka, K.; Fukuda, K.; Murakami, J. Pivotal Role of Corneal Fibroblasts in Progression to Corneal Ulcer in Bacterial Keratitis. Int. J. Mol. Sci. 2021, 22, 8979. https://doi.org/10.3390/ijms22168979
Nishida T, Sugioka K, Fukuda K, Murakami J. Pivotal Role of Corneal Fibroblasts in Progression to Corneal Ulcer in Bacterial Keratitis. International Journal of Molecular Sciences. 2021; 22(16):8979. https://doi.org/10.3390/ijms22168979
Chicago/Turabian StyleNishida, Teruo, Koji Sugioka, Ken Fukuda, and Junko Murakami. 2021. "Pivotal Role of Corneal Fibroblasts in Progression to Corneal Ulcer in Bacterial Keratitis" International Journal of Molecular Sciences 22, no. 16: 8979. https://doi.org/10.3390/ijms22168979
APA StyleNishida, T., Sugioka, K., Fukuda, K., & Murakami, J. (2021). Pivotal Role of Corneal Fibroblasts in Progression to Corneal Ulcer in Bacterial Keratitis. International Journal of Molecular Sciences, 22(16), 8979. https://doi.org/10.3390/ijms22168979