
Peroxiredoxin 1 Controls Ovulation and Ovulated Cumulus–Oocyte Complex Activity through TLR4-Derived ERK1/2 Signaling in Mice



Abstract
:1. Introduction
2. Results
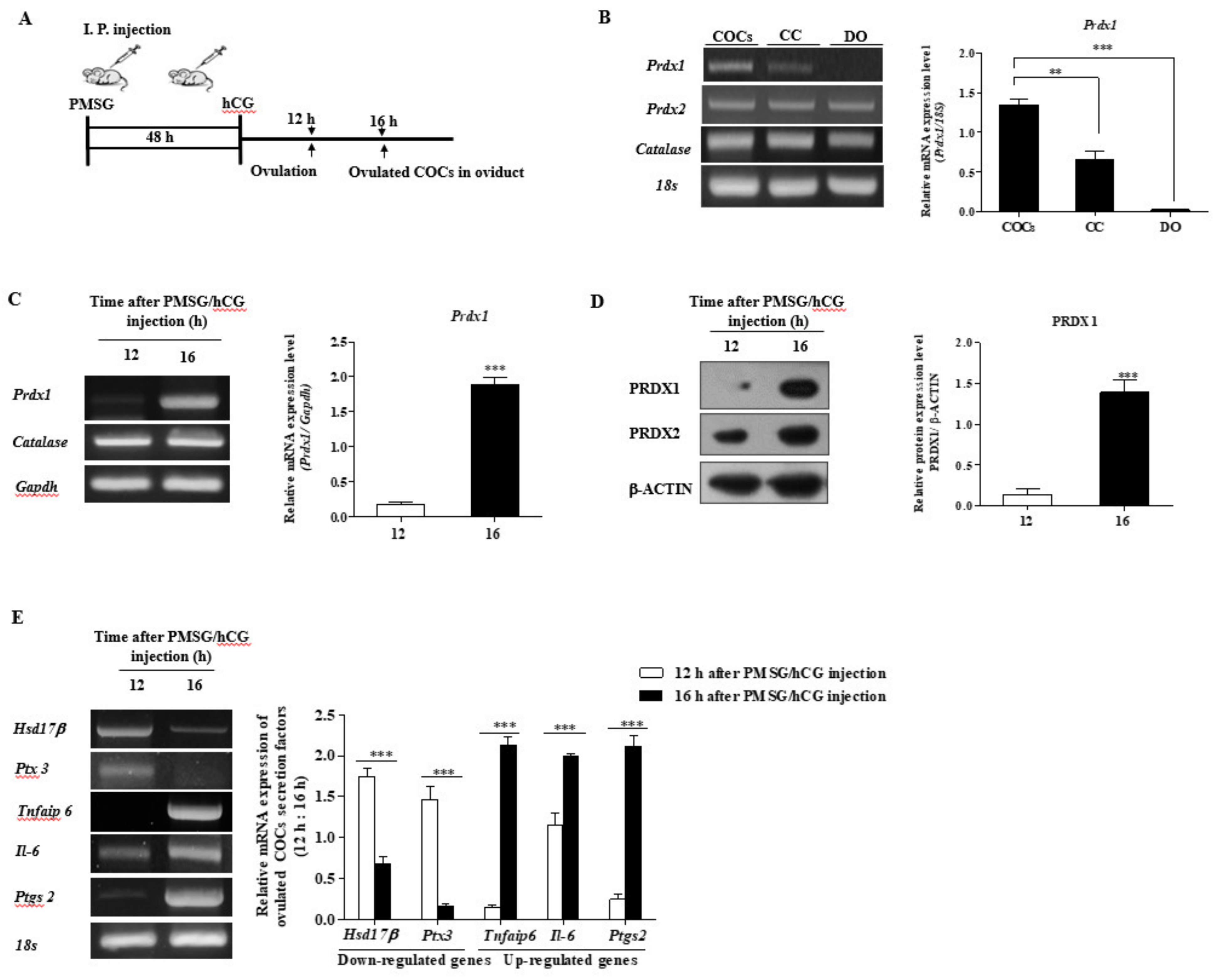
2.1. Expression of PRDX1 Protein Levels and CC Secretion Factors in Ovulated COCs of WT Mice
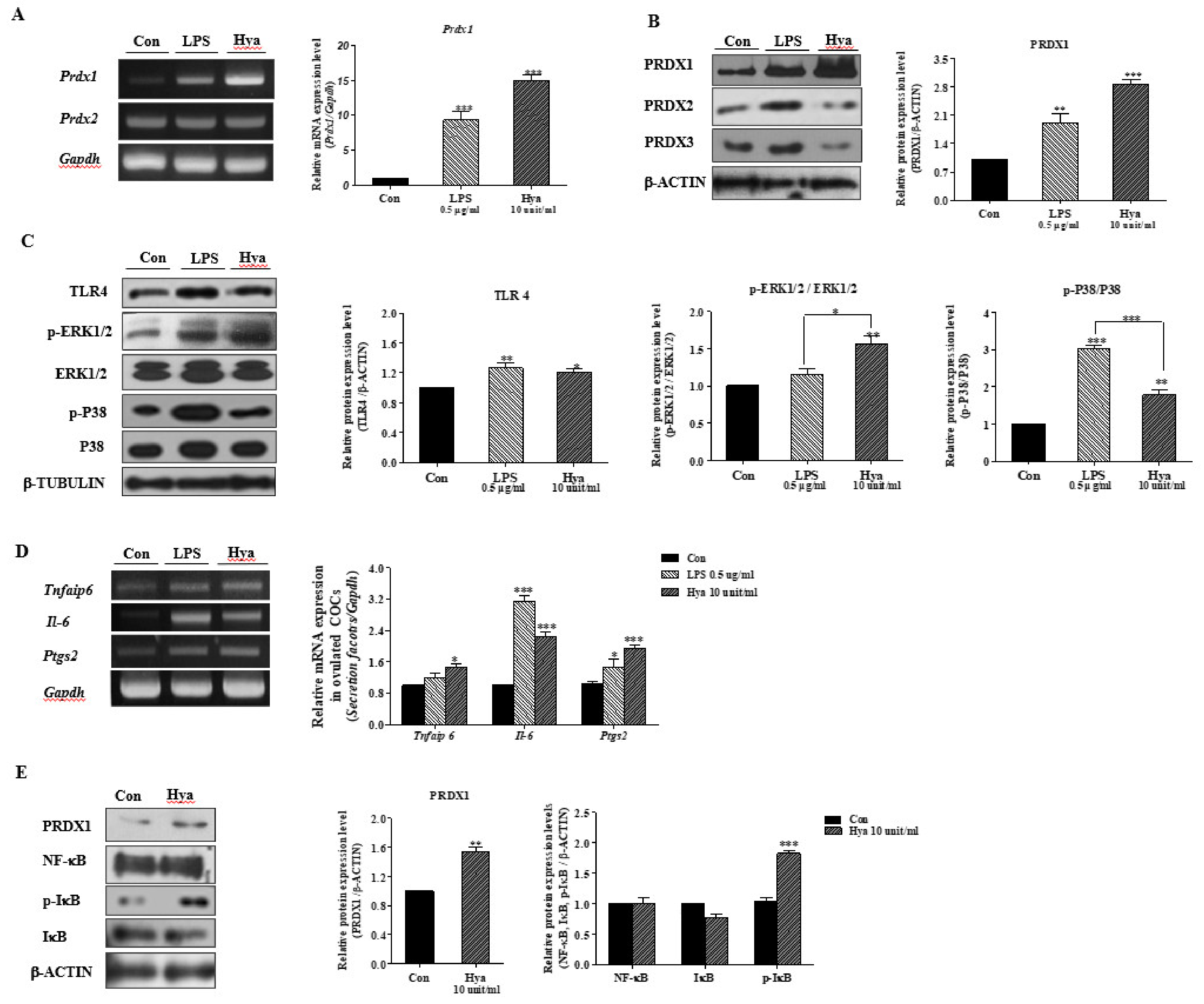
2.2. LPS or Hya Exposure in Ovulated COCs Increased PRDX1, Cumulus Secretion Factors, and the Activity of TLR4/P38/ERK1/2 Signaling Pathways
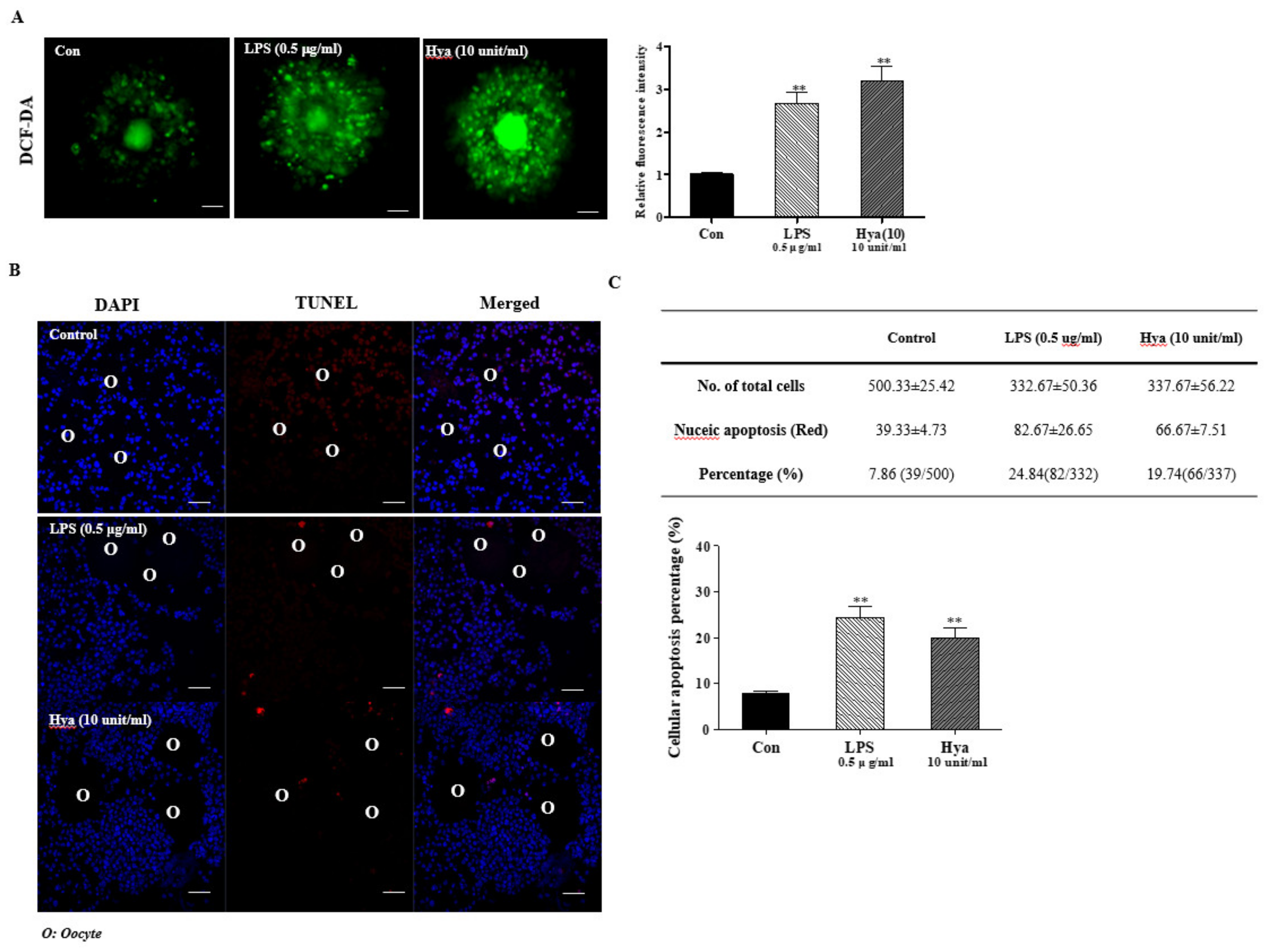
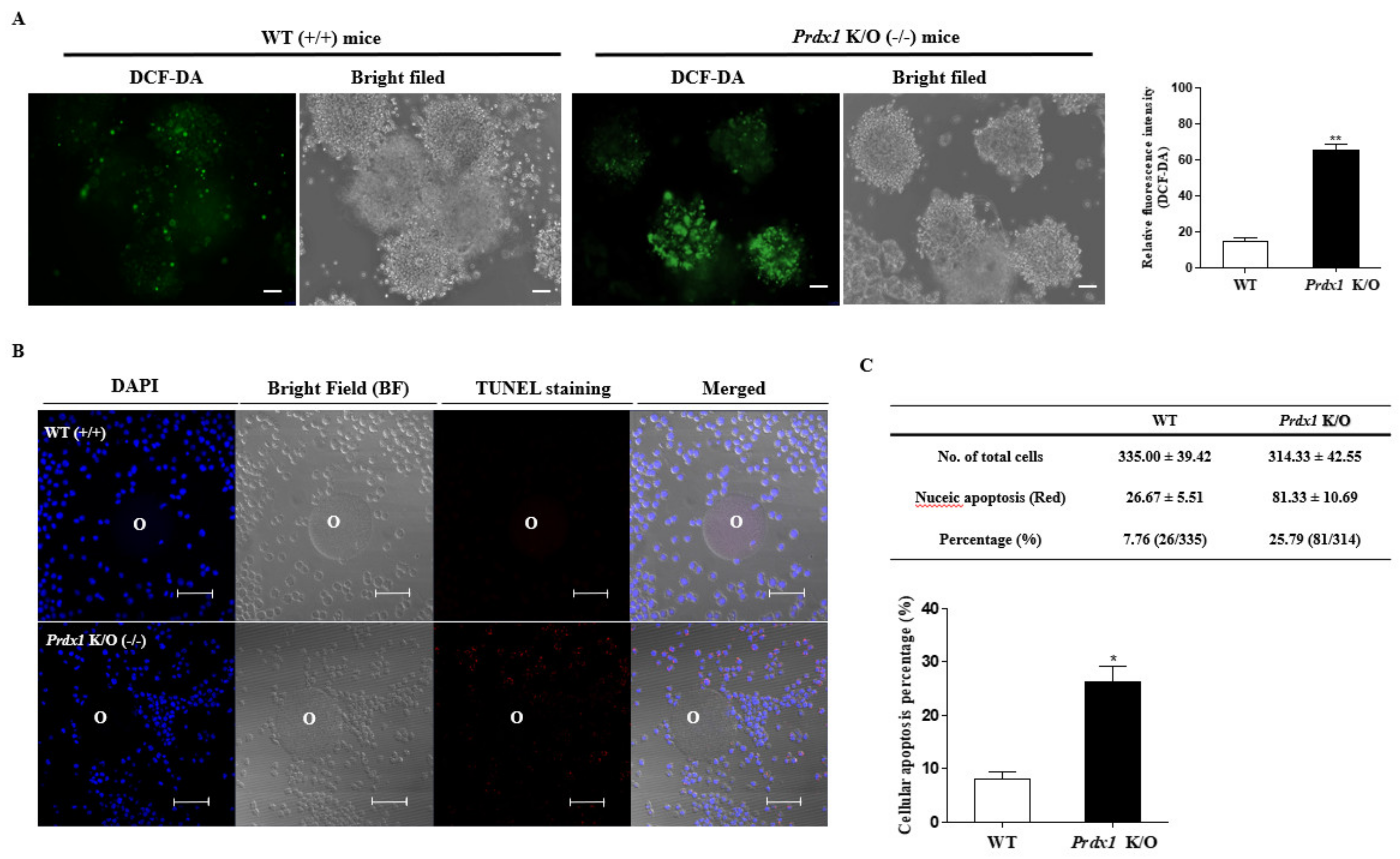
2.3. Induced ROS Production and Cellular Apoptosis in Ovulated COCs after LPS or Hya Treatment
2.4. Correlation between PRDX1 and ERK1/2 Signals in Ovulated COCs
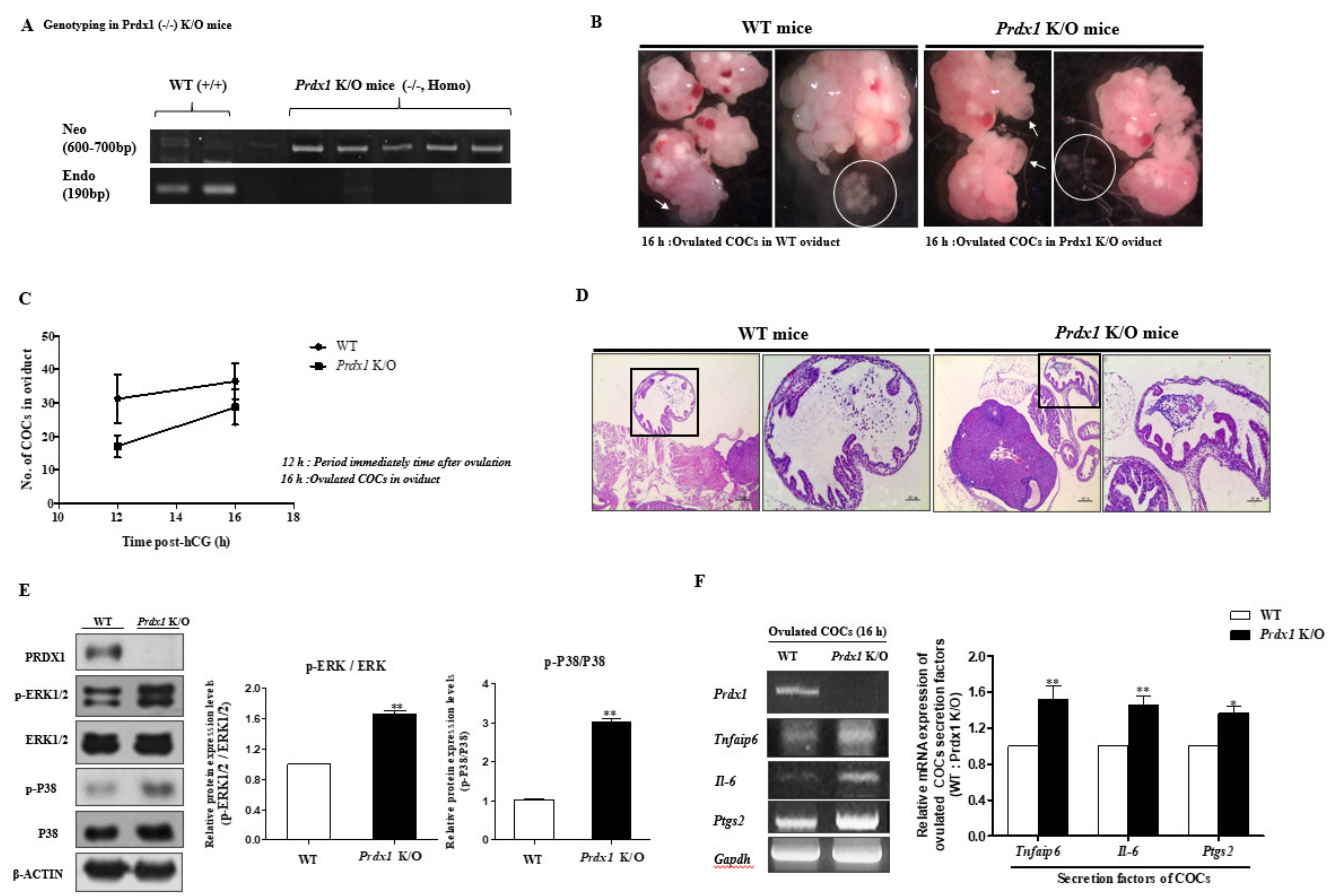
2.5. Obstruction of (Decreasing) Ovulated COC Secretion Function and Expansion via TLR4-Mediated p38 and ERK1/2 Signaling in Prdx1 Knockout (K/O) Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Isolation and Culture of Ovulated COCs
4.4. Measurement of Intracellular ROS Using DCF-DA Staining
4.5. Assessment of Apoptosis in CCs of Ovulated COCs
4.6. RNA Extraction and Reverse Transcription (RT)-PCR
4.7. Protein Extraction and Western Blot Assays
4.8. Transfection of SiRNA for the Mouse Prdx1 Gene
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Akison, L.K.; Alvino, E.R.; Dunning, K.R.; Robker, R.L.; Russell, D.L. Transient invasive migration in mouse cumulus oocyte complexes induced at ovulation by luteinizing hormone. Biol. Reprod. 2012, 86, 125. [Google Scholar] [CrossRef] [Green Version]
- Coy, P.; Garcia-Vazquez, F.A.; Visconti, P.E.; Aviles, M. Roles of the oviduct in mammalian fertilization. Reproduction 2012, 144, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Di Giacomo, M.; Camaioni, A.; Klinger, F.G.; Bonfiglio, R.; Salustri, A. Cyclic AMP-elevating Agents Promote Cumulus Cell Survival and Hyaluronan Matrix Stability, Thereby Prolonging the Time of Mouse Oocyte Fertilizability. J. Biol. Chem. 2016, 291, 3821–3836. [Google Scholar] [CrossRef] [Green Version]
- Girish, K.; Kemparaju, K. The magic glue hyaluronan and its eraser hyaluronidase: A biological overview. Life Sci. 2007, 80, 1921–1943. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.S.; Liu, Z.; Shimada, M. Immune-like mechanisms in ovulation. Trends Endocrinol. Metab. 2008, 19, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Gonzalez, I.; Gonzalez-Robayna, I.; Shimada, M.; Wayne, C.M.; Ochsner, S.A.; White, L.; Richards, J.S. Gene expression profiles of cumulus cell oocyte complexes during ovulation reveal cumulus cells express neuronal and immune-related genes: Does this expand their role in the ovulation process? Mol. Endocrinol. 2006, 20, 1300–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; McFarland-Mancini, M.M.; Funk, H.M.; Husseinzadeh, N.; Mounajjed, T.; Drew, A.F. Toll-like receptor expression in normal ovary and ovarian tumors. Cancer Immunol. Immunother. 2009, 58, 1375–1385. [Google Scholar] [CrossRef]
- Bromfield, J.J.; Sheldon, I.M. Lipopolysaccharide initiates inflammation in bovine granulosa cells via the TLR4 pathway and perturbs oocyte meiotic progression in vitro. Endocrinology 2011, 152, 5029–5040. [Google Scholar] [CrossRef] [Green Version]
- Shimada, M.; Hernandez-Gonzalez, I.; Gonzalez-Robanya, I.; Richards, J.S. Induced expression of pattern recognition receptors in cumulus oocyte complexes: Novel evidence for innate immune-like functions during ovulation. Mol. Endocrinol. 2006, 20, 3228–3239. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Gupta, S.; Sharma, R.K. Role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2005, 3, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devine, P.J.; Perreault, S.D.; Luderer, U. Roles of reactive oxygen species and antioxidants in ovarian toxicity. Biol. Reprod. 2012, 86, 27. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Shkolnik, K.; Tadmor, A.; Ben-Dor, S.; Nevo, N.; Galiani, D.; Dekel, N. Reactive oxygen species are indispensable in ovulation. Proc. Natl. Acad. Sci. USA 2011, 108, 1462–1467. [Google Scholar] [CrossRef] [Green Version]
- Fujii, J.; Iuchi, Y.; Okada, F. Fundamental roles of reactive oxygen species and protective mechanisms in the female reproductive system. Reprod. Biol. Endocrinol. 2005, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, H.A.; Jeong, W.J.; Chang, T.S.; Park, K.J.; Park, S.J.; Yang, J.S.; Rhee, S.G. Reduction of cysteine suffinic acid by sulfiredoxin is specific to 2-Cys peroxiredoxins. J. Biol. Chem. 2005, 280, 3125–3128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Immenschuh, S.; Baumgart-Vogt, E. Peroxiredoxins, oxidative stress, and cell proliferation. Antioxid. Redox Signal. 2005, 7, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.I.; Kang, W.D.; Kim, M.Y.; Cho, M.K.; Chun, S.Y. Expression of peroxiredoxin I regulated by gonadotropins in the rat ovary. Clin. Exp. Reprod. Med. 2011, 38, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Liu, X.H.; Ma, X.; Shen, Y.; Fan, L.; Leng, J.; Liu, J.Y.; Sha, J.H. The protein profile of mouse mature cumulus-oocyte complex. Biochim. Biophys. Acta 2007, 1774, 1477–1490. [Google Scholar] [CrossRef]
- Kim, S.U.; Park, Y.H.; Min, J.S.; Sun, H.N.; Han, Y.H.; Hua, J.M.; Lee, T.H.; Lee, S.R.; Chang, K.T.; Kang, S.W.; et al. Peroxiredoxin I is a ROS/p38 MAPK-dependent inducible antioxidant that regulates NF-kappaB-mediated iNOS induction and microglial activation. J. Neuroimmunol. 2013, 259, 26–36. [Google Scholar] [CrossRef]
- Marei, W.F.; Salavati, M.; Fouladi-Nashta, A.A. Critical role of hyaluronidase-2 during preimplantation embryo development. Mol. Hum. Reprod. 2013, 19, 590–599. [Google Scholar] [CrossRef] [Green Version]
- Deb, G.K.; Dey, S.R.; Bang, J.I.; Lee, J.G.; Kong, I.K. 9-cis Retinoic acid inhibits cumulus cell apoptosis during the maturation of bovine cumulus-oocyte-complexes. J. Anim. Sci. 2012, 90, 1798–1806. [Google Scholar] [CrossRef]
- Son, W.Y.; Das, M.; Shalom-Paz, E.; Holzer, H. Mechanisms of follicle selection and development. Minerva Ginecol. 2011, 63, 89–102. [Google Scholar]
- Samir, M.; Glister, C.; Mattar, D.; Laird, M.; Knight, P.G. Follicular expression of pro-inflammatory cytokines tumour necrosis factor-alpha (TNFalpha), interleukin 6 (IL6) and their receptors in cattle: TNFalpha, IL6 and macrophages suppress thecal androgen production in vitro. Reproduction 2017, 154, 35–49. [Google Scholar] [CrossRef]
- Liu, Z.; Shimada, M.; Richards, J.S. The involvement of the Toll-like receptor family in ovulation. J. Assist. Reprod. Genet. 2008, 25, 223–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, M.; Yanai, Y.; Okazaki, T.; Noma, N.; Kawashima, I.; Mori, T.; Richards, J.S. Hyaluronan fragments generated by sperm-secreted hyaluronidase stimulate cytokine/chemokine production via the TLR2 and TLR4 pathway in cumulus cells of ovulated COCs, which may enhance fertilization. Development 2008, 135, 2001–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCubrey, J.A.; Lahair, M.M.; Franklin, R.A. Reactive oxygen species-induced activation of the MAP kinase signaling pathways. Antioxid. Redox Signal. 2006, 8, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Luo, A.; Hao, X.; Lai, Z.; Ding, T.; Ma, X.; Mayinuer, M.; Shen, W.; Wang, X.; Lu, Y.; et al. Peroxiredoxin 2 inhibits granulosa cell apoptosis during follicle atresia through the NFKB pathway in mice. Biol. Reprod. 2011, 84, 1182–1189. [Google Scholar] [CrossRef] [Green Version]
- Park, J.I.; Jeon, H.J.; Jung, N.K.; Jang, Y.J.; Kim, J.S.; Seo, Y.W.; Jeong, M.; Chae, H.Z.; Chun, S.Y. Periovulatory expression of hydrogen peroxide-induced sulfiredoxin and peroxiredoxin 2 in the rat ovary: Gonadotropin regulation and potential modification. Endocrinology 2012, 153, 5512–5521. [Google Scholar] [CrossRef] [Green Version]
- Banmeyer, I.; Marchand, C.; Verhaeghe, C.; Vucic, B.; Rees, J.F.; Knoops, B. Overexpression of human peroxiredoxin 5 in subcellular compartments of Chinese hamster ovary cells: Effects on cytotoxicity and DNA damage caused by peroxides. Free Radic. Biol. Med. 2004, 36, 65–77. [Google Scholar] [CrossRef]
- West, A.P.; Brodsky, I.E.; Rahner, C.; Woo, D.K.; Erdjument-Bromage, H.; Tempst, P.; Walsh, M.C.; Choi, Y.; Shadel, G.S.; Ghosh, S. TLR signalling augments macrophage bactericidal activity through mitochondrial ROS. Nature 2011, 472, 476–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddell, J.R.; Wang, X.Y.; Minderman, H.; Gollnick, S.O. Peroxiredoxin 1 stimulates secretion of proinflammatory cytokines by binding to TLR4. J. Immunol. 2010, 184, 1022–1030. [Google Scholar] [CrossRef] [PubMed]
- Taniuchi, K.; Furihata, M.; Hanazaki, K.; Iwasaki, S.; Tanaka, K.; Shimizu, T.; Saito, M.; Saibara, T. Peroxiredoxin 1 promotes pancreatic cancer cell invasion by modulating p38 MAPK activity. Pancreas 2015, 44, 331–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, J.M.; Moriarty-Craige, S.; Jones, D.P. Nuclear and cytoplasmic peroxiredoxin-1 differentially regulate NF-kappaB activities. Free Radic. Biol. Med. 2007, 43, 282–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.L.; Zhao, L.X.; Zhang, S.; Du, J.R. Peroxiredoxin 1-mediated activation of TLR4/NF-kappaB pathway contributes to neuroinflammatory injury in intracerebral hemorrhage. Int. Immunopharmacol. 2016, 41, 82–89. [Google Scholar] [CrossRef]
- Yu, F.-Q.; Han, C.-S.; Yang, W.; Jin, X.; Hu, Z.-Y.; Liu, Y.-X. Activation of the p38 MAPK pathway by follicle-stimulating hormone regulates steroidogenesis in granulosa cells differentially. J. Endocrinol. 2005, 186, 85–96. [Google Scholar] [CrossRef]
- Chuderland, D.; Dvashi, Z.; Kaplan-Kraicer, R.; Ben-Meir, D.; Shalgi, R.; Lavi, S. De novo synthesis of protein phosphatase 1A, magnesium dependent, alpha isoform (PPM1A) during oocyte maturation. Cell. Mol. Biol. Lett. 2012, 17, 433–445. [Google Scholar] [CrossRef]
- Schuermann, Y.; Rovani, M.T.; Gasperin, B.; Ferreira, R.; Ferst, J.; Madogwe, E.; Goncalves, P.B.; Bordignon, V.; Duggavathi, R. ERK1/2-dependent gene expression in the bovine ovulating follicle. Sci. Rep. 2018, 8, 16170. [Google Scholar] [CrossRef]
- Salustri, A.; Garlanda, C.; Hirsch, E.; De Acetis, M.; Maccagno, A.; Bottazzi, B.; Doni, A.; Bastone, A.; Mantovani, G.; Beck Peccoz, P.; et al. PTX3 plays a key role in the organization of the cumulus oophorus extracellular matrix and in in vivo fertilization. Development 2004, 131, 1577–1586. [Google Scholar] [CrossRef] [Green Version]
- Takeo, T.; Nakagata, N. Superovulation using the combined administration of inhibin antiserum and equine chorionic gonadotropin increases the number of ovulated oocytes in C57BL/6 female mice. PLoS ONE 2015, 10, e0128330. [Google Scholar] [CrossRef]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Aasada, H.; Yamagata, Y.; et al. The role of melatonin as an antioxidant in the follicle. J. Ovarian Res. 2012, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer | Sequence Reported 5’-3’ | Tm °C | Amplicon Length (bp) |
|---|---|---|---|---|
| Hsd17βB3 (NM_008291.3) | Sense | ATGGAGTCAAGGAGGAAAGGC | 61 | 556 |
| Antisense | GGCTGTAAAGAGGCCAGGG | |||
| Ptx3 (NM_008987) | Sense | GGACAACGAAATAGACAATGGACTT | 55 | 109 |
| Antisense | CGAGTTCTCCAGCATGATGAAC | |||
| Il-6 (NM_001314054.1) | Sense | AGTTGCCTTCTTGGGACTGA | 57 | 223 |
| Antisense | TTCTGCAAGTGCATCATCGT | |||
| Ptgs2 (NM_011198.3) | Sense | TGTACAAGCAGTGGCAAAGG | 53 | 433 |
| Antisense | CAATGTGCAAGATCCACAGC | |||
| Tnfaip6 (NM_009398) | Sense | TTCCATGTCTGTGCTGCTGGATGG | 64 | 330 |
| Antisense | TTTGACCTTGAACATGATCCAGGCT |
| Target | Primer | Sequence Reported 5’-3’ | Tm °C | Amplicon Length (bp) |
|---|---|---|---|---|
| Prdx1 (NM_011034) | Sense | CACCCAAGAAACAAGGAGGA | 53.5 | 343 |
| Antisense | TGGTCCAGTGCTCACTTCTG | |||
| Prdx2 (NM_011563) | Sense | AGGACTTCCGAAAGCTAGGC | 54.1 | 387 |
| Antisense | CCTTGCTGTCATCCACATTG | |||
| Prdx3 (NM_007452) | Sense | CTCTGCCCAAGGAAAGTCAG | 53.3 | 463 |
| Antisense | GACACTCAGGTGCTTGACGA | |||
| Prdx4 (NM_016764) | Sense | TCTCCAAGCCAGCACCTTAT | 53.2 | 429 |
| Antisense | ATCTTCCGACAGGAAGGTCA | |||
| Prdx5 (NM_012021) | Sense | GGCATTTACACCTGGCTGTT | 53.5 | 297 |
| Antisense | AGTGCCTTCACTATGCCGTT | |||
| Prdx6 (NM_007453.4) | Sense | CCTGGAGCAAGGACATCAAT | 53.5 | 326 |
| Antisense | TACCATCACGCTCTCTCCCT | |||
| Catalase (NM_009804) | Sense | GCGTCCAGTGCGCTGTAGA | 59.2 | 199 |
| Antisense | TCAGGGTGGACGTCAGTGAA | |||
| 18s (NR_003278.3) | Sense | GTAACCCGTTGAACCCCATT | 57 | 151 |
| Antisense | CCATCCAATCGGTAGTAGCG | |||
| Gapdh (BC145810.1) | Sense | ACCACAGTCCATGCCATCAC | 55 | 452 |
| Antisense | TCCACCACCCTGTTGCTGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.-J.; Kim, B.; Koo, D.-B.; Lee, D.-S. Peroxiredoxin 1 Controls Ovulation and Ovulated Cumulus–Oocyte Complex Activity through TLR4-Derived ERK1/2 Signaling in Mice. Int. J. Mol. Sci. 2021, 22, 9437. https://doi.org/10.3390/ijms22179437
Park H-J, Kim B, Koo D-B, Lee D-S. Peroxiredoxin 1 Controls Ovulation and Ovulated Cumulus–Oocyte Complex Activity through TLR4-Derived ERK1/2 Signaling in Mice. International Journal of Molecular Sciences. 2021; 22(17):9437. https://doi.org/10.3390/ijms22179437
Chicago/Turabian StylePark, Hyo-Jin, Bokyung Kim, Deog-Bon Koo, and Dong-Seok Lee. 2021. "Peroxiredoxin 1 Controls Ovulation and Ovulated Cumulus–Oocyte Complex Activity through TLR4-Derived ERK1/2 Signaling in Mice" International Journal of Molecular Sciences 22, no. 17: 9437. https://doi.org/10.3390/ijms22179437
APA StylePark, H. -J., Kim, B., Koo, D. -B., & Lee, D. -S. (2021). Peroxiredoxin 1 Controls Ovulation and Ovulated Cumulus–Oocyte Complex Activity through TLR4-Derived ERK1/2 Signaling in Mice. International Journal of Molecular Sciences, 22(17), 9437. https://doi.org/10.3390/ijms22179437