
Does the Rainbow Trout Ovarian Fluid Promote the Spermatozoon on Its Way to the Egg?

 , ,
, ,  ,
,
Abstract
:1. Introduction
- How does the presence of ovarian fluid affect the outcome of in vitro fertilization?
- What is the effect of ovarian fluid in the activation medium on the spermatozoa motility traits, including velocity and linearity of motion?
- Does ovarian fluid have a chemotactic effect on spermatozoa?
- Are changes in spermatozoa motility associated with specific features of the ovarian fluid, including osmolarity and Ca2+ ions?
2. Results
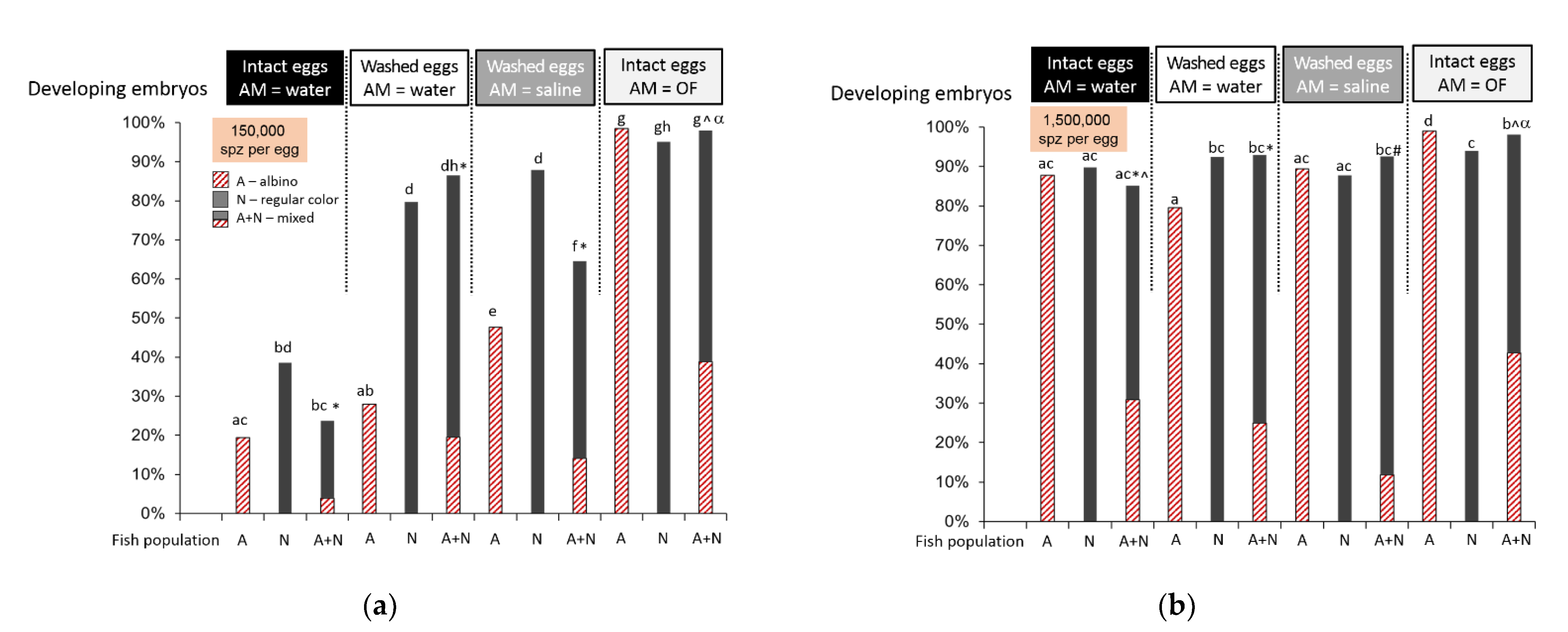
2.1. Ovarian Fluid Enhances the In Vitro Fertilization Outcome
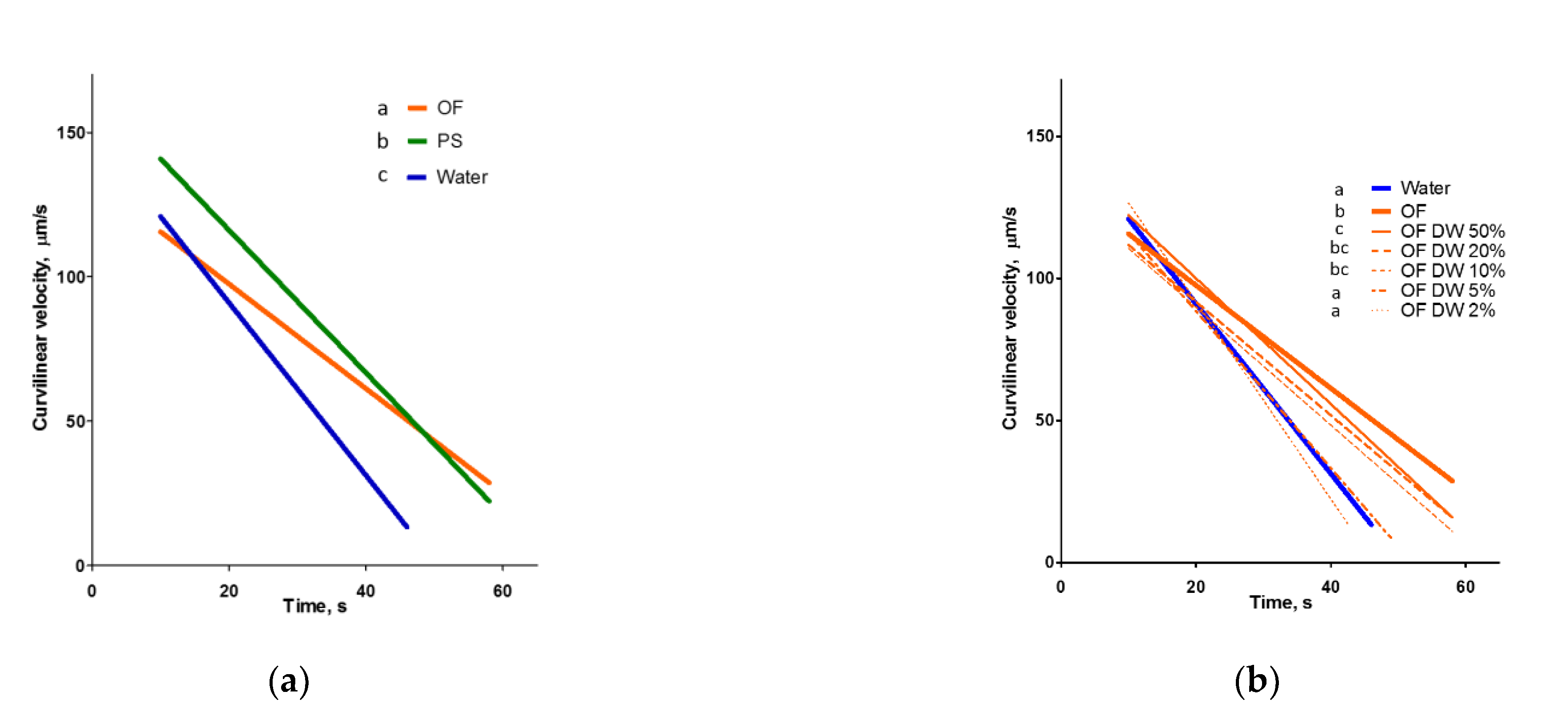
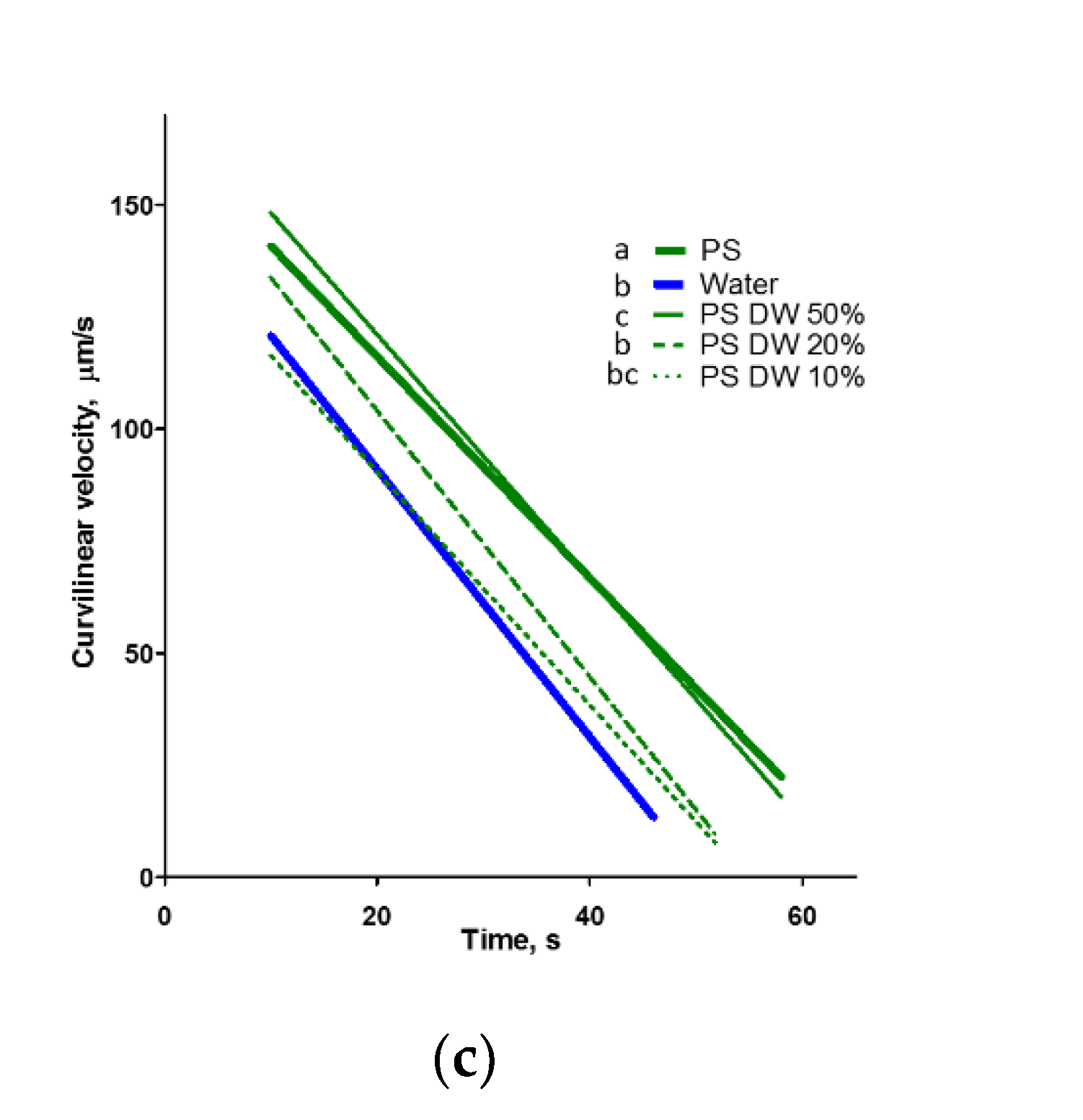
2.2. Ovarian Fluid Prolongs the Motility of Spermatozoa
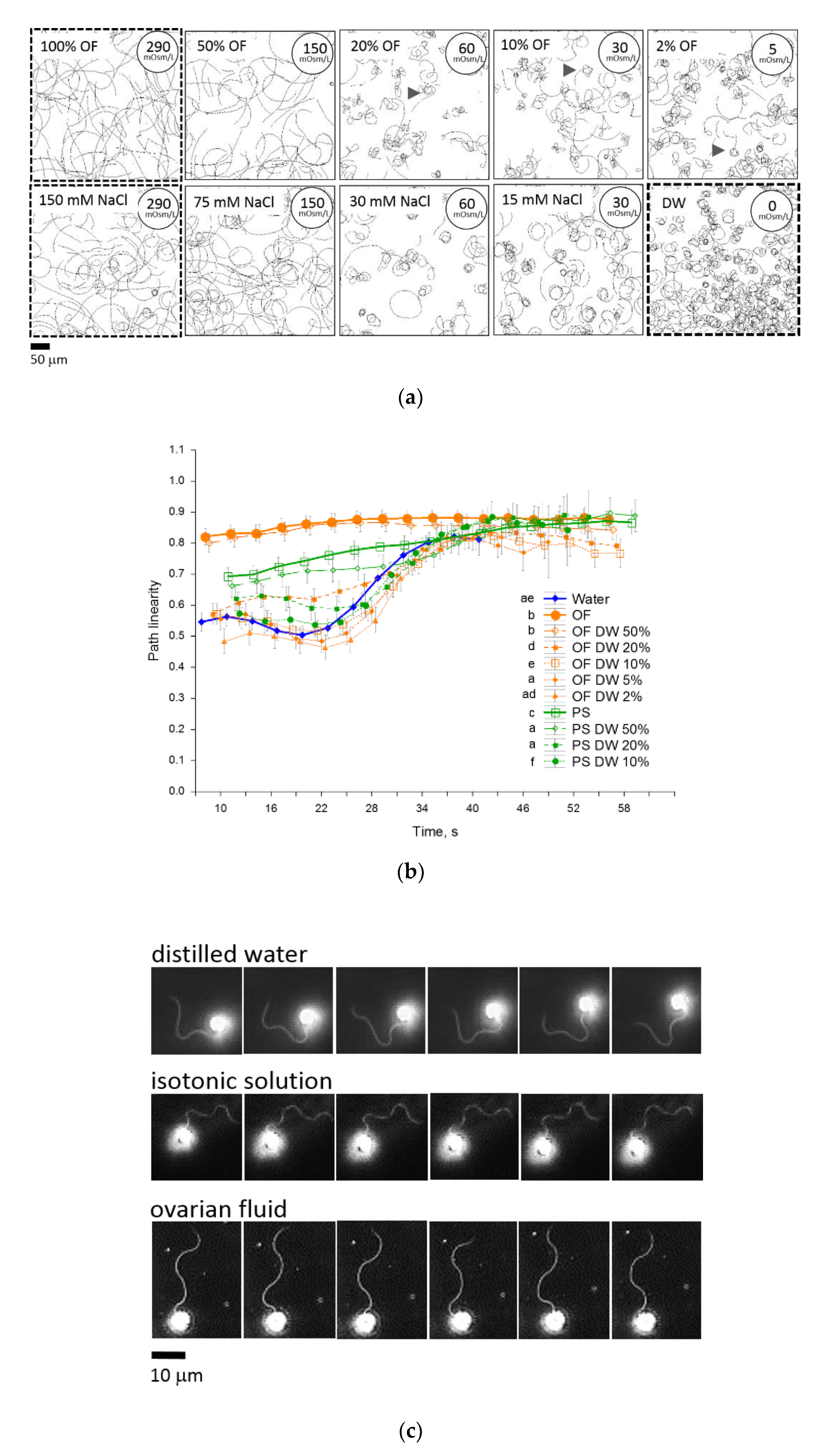
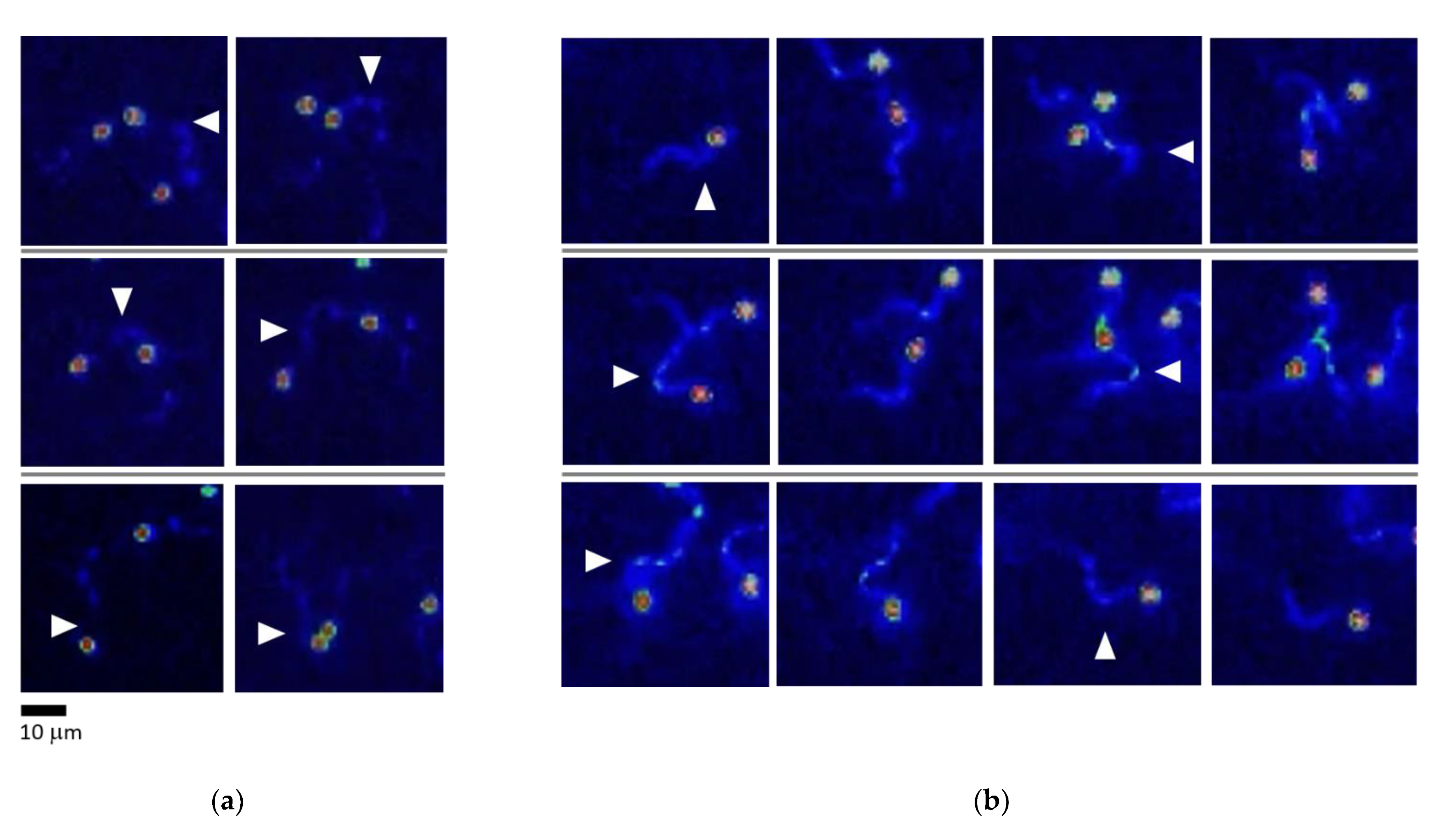
2.3. Ovarian Fluid Straightens Trajectories and Has a Trapping Effect on Rainbow Trout Spermatozoa
2.4. Rainbow Trout Spermatozoa React to an Abrupt Ca2+ Rise in Flagella during Turn-and-Run Behavior
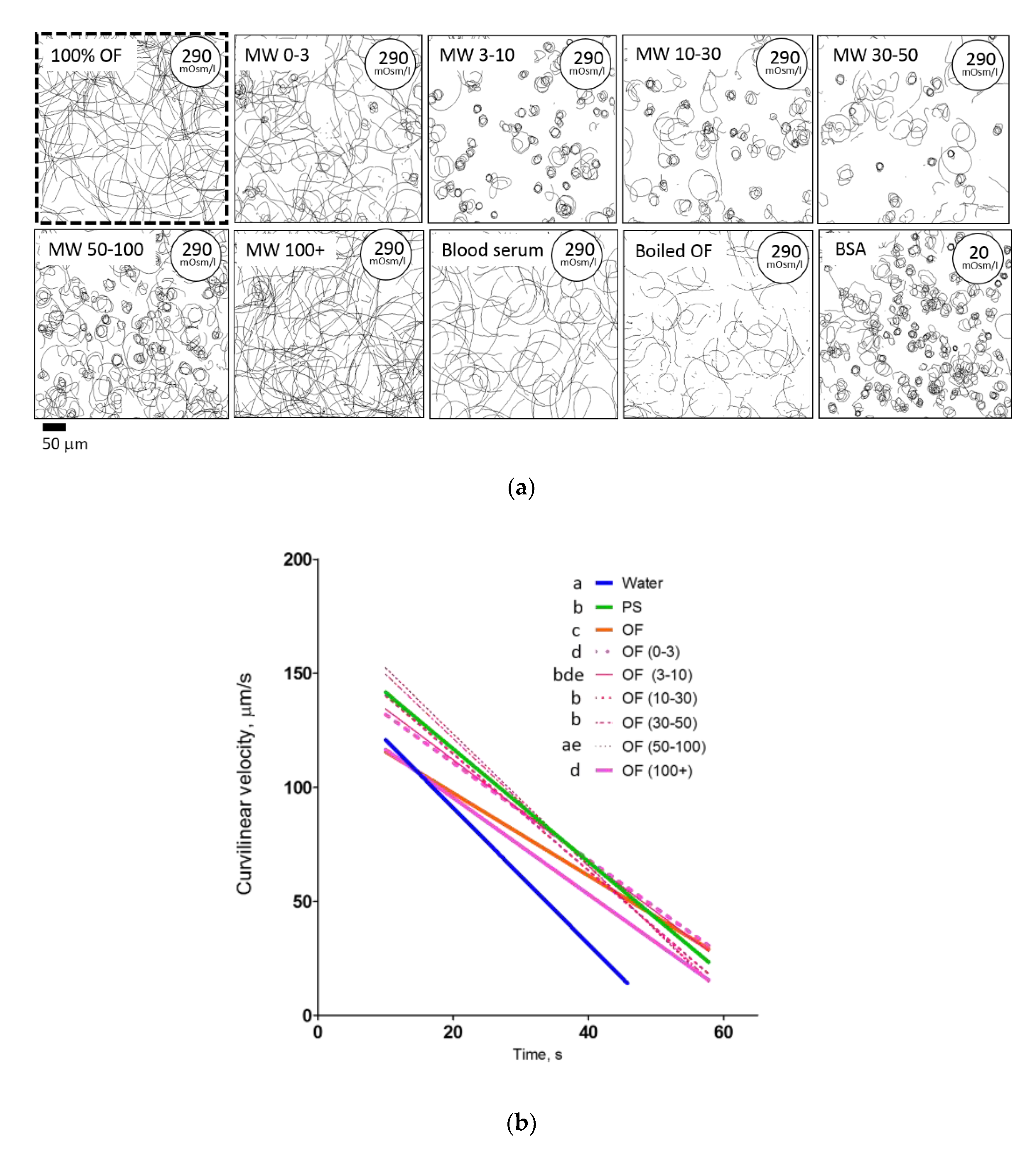
2.5. Isolated Agents of Ovarian Fluid Differently Affect the Behavior of Rainbow Trout Spermatozoa––Effect of Ovarian Fluid Molecular Weight Fractions and Other Protein Solutions
2.6. Medium Osmolarity and Ca2+ Content Have a “Cross Effect” on Motility and Trapping of Rainbow Trout Spermatozoa
3. Discussion
3.1. Rainbow Trout Ovarian Fluid Provides an Optimal Environment for Fertilization
3.2. Rainbow Trout Ovarian Fluid Enhances the Kinetic Traits of Spermatozoa
3.3. Ca2+ Concentration and Osmolarity Have Cross-Effects on Motility Traits
3.4. The Ovarian Fluid Causes the Attraction and Trapping of Spermatozoa
3.5. Effect of Rainbow Trout Ovarian Fluid on Spermatozoa Is in Line with the Effects of Female Factors in Other Externally Fertilizing Species
4. Materials and Methods
4.1. Fish Broodstock—Gametes and Fluids Collection
4.2. Preparation of Media
4.3. Motility Observation and Recording
4.4. Sperm Chemotaxis Tests
4.5. High-Speed Imaging of Spermatozoon Flagellar Beating
4.6. Ca2+ Concentration Imaging
4.7. In Vitro Fertilization
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | |||||
| Medium | R2 | p | A ± SE (Slope of Regression Line) | B ± SE (Intercept with y (VCL) Axis) | C (Intercept with x (Time) Axis) |
| Water | 0.86 | <0.0001 | −3.21 ± 0.08 a | 151.5 ± 2.1 | 47.25 |
| OF | 0.88 | <0.0001 | −1.88 ± 0.03 b | 139.2 ± 1.2 | 73.89 |
| PS | 0.94 | <0.0001 | −2.50 ± 0.03 c | 166.7 ± 1.0 | 66.58 |
| (b) | |||||
| Medium | R2 | p | A ± SE (Slope of Regression Line) | B ± SE (Intercept with y (VCL) Axis) | C (Intercept with x (Time) Axis) |
| Water | 0.86 | <0.0001 | −3.21 ± 0.08 a | 151.5 ± 2.1 | 47.25 |
| PS | 0.94 | <0.0001 | −2.50 ± 0.03 b | 166.7 ± 1.0 | 66.58 |
| PS DW 50% | 0.95 | <0.0001 | −2.72 ± 0.05 c | 173.6 ± 1.7 | 63.75 |
| PS DW 20% | 0.90 | <0.0001 | −3.20 ± 0.08 b | 167.9 ± 2.5 | 52.41 |
| PS DW 10% | 0.86 | <0.0001 | −2.99 ± 0.10 bc | 151.5 ± 3.0 | 50.77 |
| (c) | |||||
| Medium | R2 | p | A ± SE (Slope of Regression Line) | B ± SE (Intercept with y (VCL) Axis) | C (Intercept with x (Time) Axis) |
| Water | 0.86 | <0.0001 | −3.21 ± 0.08 a | 151.5 ± 2.1 | 47.25 |
| OF | 0.88 | <0.0001 | −1.88 ± 0.03 b | 139.2 ± 1.2 | 73.89 |
| OF DW 50% | 0.89 | <0.0001 | −2.19 ± 0.05 c | 144.4 ± 1.8 | 65.95 |
| OF DW 20% | 0.85 | <0.0001 | −2.01 ± 0.06 bc | 130.4 ± 2.2 | 64.76 |
| OF DW 10% | 0.80 | <0.0001 | −2.16 ± 0.08 bc | 130.0 ± 2.9 | 60.17 |
| OF DW 5% | 0.89 | <0.0001 | −3.03 ± 0.10 a | 147.2 ± 2.8 | 48.51 |
| OF DW 2% | 0.87 | <0.0001 | −3.38 ± 0.14 a | 152.5 ± 3.8 | 45.10 |
| (d) | |||||
| Medium | R2 | p | A ± SE (Slope of Regression Line) | B ± SE (Intercept with y (VCL) Axis) | C (Intercept with x (Time) Axis) |
| Water | 0.86 | <0.0001 | −3.21 ± 0.08 a | 151.5 ± 2.1 | 47.25 |
| OF | 0.88 | <0.0001 | −1.88 ± 0.03 b | 139.2 ± 1.2 | 73.89 |
| PS | 0.94 | <0.0001 | −2.50 ± 0.03 c | 166.7 ± 1.0 | 66.58 |
| OF (0–3) | 0.85 | <0.0001 | −2.11 ± 0.06 d | 152.0 ± 2.3 | 71.88 |
| OF (3–10) | 0.84 | <0.0001 | −2.36 ± 0.10 bde | 157.6 ± 3.5 | 66.91 |
| OF (10–30) | 0.93 | <0.0001 | −2.66 ± 0.06 b | 169.4 ± 2.1 | 63.59 |
| OF (30–50) | 0.93 | <0.0001 | −2.63 ± 0.06 b | 172.1 ± 2.0 | 65.45 |
| OF (50–100) | 0.93 | <0.0001 | −2.93 ± 0.06 ac | 181.6 ± 2.1 | 62.03 |
| OF (100+) | 0.85 | <0.0001 | −2.21 ± 0.07 d | 140.3 ± 2.6 | 63.43 |
References
- Hart, N.H. Fertilization in Teleost Fishes: Mechanisms of Sperm-Egg Interactions. Int. Rev. Cytol. 1990, 121, 1–66. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A. Sperm competition games: Sperm size and sperm number under adult control. Proc. R. Soc. B Boil. Sci. 1993, 253, 245–254. [Google Scholar] [CrossRef]
- Thornhill, R. Cryptic Female Choice and Its Implications in the Scorpionfly Harpobittacus nigriceps. Am. Nat. 1983, 122, 765–788. [Google Scholar] [CrossRef]
- Eisenbach, M.; Giojalas, L. Sperm guidance in mammals—An unpaved road to the egg. Nat. Rev. Mol. Cell Biol. 2006, 7, 276–285. [Google Scholar] [CrossRef]
- Morisawa, M.; Yoshida, M. Activation of motility and chemotaxis in the spermatozoa: From invertebrates to humans. Reprod. Med. Biol. 2005, 4, 101–114. [Google Scholar] [CrossRef]
- Yanagimachi, R.; Harumi, T.; Matsubara, H.; Yan, W.; Yuan, S.; Hirohashi, N.; Iida, T.; Yamaha, E.; Arai, K.; Matsubara, T.; et al. Chemical and physical guidance of fish spermatozoa into the egg through the micropyle. Biol. Reprod. 2017, 96, 780–799. [Google Scholar] [CrossRef] [Green Version]
- Cosson, J.; Billard, R.; Gibert, C.; Dreanno, C.; Suquet, M. Ionic factors regulating the motility of fish sperm. In The Male Gamete: From Basic to Clinical Applications; Gagnon, C., Ed.; Cache River Press: St. Louis, MO, USA, 1999; pp. 161–186. [Google Scholar]
- Lillie, F.R. The Production of Sperm Iso-Agglutinins by Ova. Science 1912, 36, 527–530. [Google Scholar] [CrossRef]
- Kaupp, U. 100 years of sperm chemotaxis. J. Gen. Physiol. 2012, 140, 583–586. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Shiba, K.; Sakamoto, A.; Ikenaga, J.; Matsunaga, S.; Inaba, K.; Yoshida, M. Ca2+ efflux via plasma membrane Ca2+-ATPase mediates chemotaxis in ascidian sperm. Sci. Rep. 2018, 8, 16622. [Google Scholar] [CrossRef]
- Hurk, R.V.D.; Peute, J. Cyclic changes in the ovary of the rainbow trout, Salmo gairdneri, with special reference to sites of steroidogenesis. Cell Tissue Res. 1979, 199, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Rosengrave, P.; Gemmell, N.; Metcalf, V.; McBride, K.; Montgomerie, R. A mechanism for cryptic female choice in chinook salmon. Behav. Ecol. 2008, 19, 1179–1185. [Google Scholar] [CrossRef] [Green Version]
- Lahnsteiner, F.; Weismann, T.; Patzner, R. Composition of the ovarian fluid in 4 salmonid species: Oncorhynchus mykiss, Salmo trutta f lacustris, Saivelinus alpinus and Hucho hucho. Reprod. Nutr. Dev. 1995, 35, 465–474. [Google Scholar] [CrossRef]
- Nynca, J.; Arnold, G.J.; Fröhlich, T.; Ciereszko, A. Shotgun proteomics of rainbow trout ovarian fluid. Reprod. Fertil. Dev. 2015, 27, 504–512. [Google Scholar] [CrossRef]
- Rosengrave, P.; Taylor, H.; Montgomerie, R.; Metcalf, V.; McBride, K.; Gemmell, N. Chemical composition of seminal and ovarian fluids of chinook salmon (Oncorhynchus tshawytscha) and their effects on sperm motility traits. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 152, 123–129. [Google Scholar] [CrossRef]
- Lahnsteiner, F. The influence of ovarian fluid on the gamete physiology in the Salmonidae. Fish Physiol. Biochem. 2002, 27, 49–59. [Google Scholar] [CrossRef]
- Dietrich, M.A.; Wojtczak, M.; Słowińska, M.; Dobosz, S.; Kuźmiński, H.; Ciereszko, A. Effects of ovarian fluid on motility characteristics of rainbow trout (Oncorhynchus mykiss Walbaum) spermatozoa. J. Appl. Ichthyol. 2008, 24, 503–507. [Google Scholar] [CrossRef]
- Kanuga, M.; Drew, R.; Wilson-Leedy, J.; Ingermann, R. Subpopulation distribution of motile sperm relative to activation medium in steelhead (Oncorhynchus mykiss). Theriogenology 2012, 77, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Poli, F.; Immler, S.; Gasparini, C. Effects of ovarian fluid on sperm traits and its implications for cryptic female choice in zebrafish. Behav. Ecol. 2019, 30, 1298–1305. [Google Scholar] [CrossRef]
- Myers, J.N.; Senior, A.; Zadmajid, V.; Sørensen, S.R.; Butts, I.A.E. Associations between Ovarian Fluid and Sperm Swimming Trajectories in Marine and Freshwater Teleosts: A Meta-Analysis. Rev. Fish. Sci. Aquac. 2020, 28, 322–339. [Google Scholar] [CrossRef]
- Hatef, A.; Niksirat, H.; Amiri, B.M.; Alavi, S.M.H.; Karami, M. Sperm density, seminal plasma composition and their physiological relationship in the endangered Caspian brown trout (Salmo trutta caspius). Aquac. Res. 2007, 38, 1175–1181. [Google Scholar] [CrossRef]
- Gasparini, C.; Pilastro, A. Cryptic female preference for genetically unrelated males is mediated by ovarian fluid in the guppy. Proc. R. Soc. B Boil. Sci. 2011, 278, 2495–2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvano, P.M.; Johnson, K.; Wilson, C.C.; Pitcher, T.E.; Butts, I.A. Ovarian fluid influences sperm performance in lake trout, Salvelinus namaycush. Reprod. Biol. 2013, 13, 172–175. [Google Scholar] [CrossRef]
- Miller, R.L. Sperm Chemotaxis in Ascidians. Am. Zool. 1982, 22, 827–840. [Google Scholar] [CrossRef]
- Kaupp, U.; Strünker, T. Signaling in Sperm: More Different than Similar. Trends Cell Biol. 2017, 27, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Yoshida, K. Sperm chemotaxis and regulation of flagellar movement by Ca2+. Mol. Hum. Reprod. 2011, 17, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Shiba, K.; Baba, S.A.; Inoue, T.; Yoshida, M. Ca2+ bursts occur around a local minimal concentration of attractant and trigger sperm chemotactic response. Proc. Natl. Acad. Sci. USA 2008, 105, 19312–19317. [Google Scholar] [CrossRef] [Green Version]
- Zadmajid, V.; Myers, J.N.; Sørensen, S.R.; Butts, I.A.E. Ovarian fluid and its impacts on spermatozoa performance in fish: A review. Theriogenology 2019, 132, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Hatef, A.; Niksirat, H.; Alavi, S.M.H. Composition of ovarian fluid in endangered Caspian brown trout, Salmo trutta caspius, and its effects on spermatozoa motility and fertilizing ability compared to freshwater and a saline medium. Fish Physiol. Biochem. 2009, 35, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Hugunin, H.A.; Parsons, J.E.; Nagler, J.J. The influence of coelomic fluid on in vitro fertilization success in rainbow trout (Oncorhynchus mykiss). Aquaculture 2008, 281, 155–157. [Google Scholar] [CrossRef]
- Rime, H.; Guitton, N.; Pineau, C.; Bonnet, E.; Bobe, J.; Jalabert, B. Post-ovulatory ageing and egg quality: A proteomic analysis of rainbow trout coelomic fluid. Reprod. Biol. Endocrinol. 2004, 2, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butts, I.A.; Johnson, K.; Wilson, C.; Pitcher, T. Ovarian fluid enhances sperm velocity based on relatedness in lake trout, Salvelinus namaycush. Theriogenology 2012, 78, 2105–2109.e1. [Google Scholar] [CrossRef]
- Wojtczak, M.; Dietrich, G.J.; Słowińska, M.; Dobosz, S.; Kuźmiński, H.; Ciereszko, A. Ovarian fluid pH enhances motility parameters of rainbow trout (Oncorhynchus mykiss) spermatozoa. Aquaculture 2007, 270, 259–264. [Google Scholar] [CrossRef]
- Kleppe, S.A.; Nordeide, J.T.; Rudolfsen, G.; Figenschou, L.; Larsen, B.; Reiss, K.; Folstad, I. No support for cryptic choice by ovarian fluid in an external fertilizer. Ecol. Evol. 2018, 8, 11763–11774. [Google Scholar] [CrossRef] [PubMed]
- Lahnsteiner, F.; Berger, B.; Weismann, T.; Patzner, R. Determination of semen quality of the rainbow trout, Oncorhynchus mykiss, by sperm motility, seminal plasma parameters, and spermatozoal metabolism. Aquaculture 1998, 163, 163–181. [Google Scholar] [CrossRef]
- Levitan, D.R. Sperm velocity and longevity trade off each other and influence fertilization in the sea urchin Lytechinus variegatus. Proc. R. Soc. B Boil. Sci. 2000, 267, 531–534. [Google Scholar] [CrossRef] [Green Version]
- Taborsky, M. Sperm competition in fish: ‘bourgeois’ males and parasitic spawning. Trends Ecol. Evol. 1998, 13, 222–227. [Google Scholar] [CrossRef]
- Gage, M.; Macfarlane, C.P.; Yeates, S.; Ward, R.G.; Searle, J.B.; Parker, G.A. Spermatozoal Traits and Sperm Competition in Atlantic Salmon Relative Sperm Velocity Is the Primary Determinant of Fertilization Success. Curr. Biol. 2004, 14, 44–47. [Google Scholar] [CrossRef]
- Liljedal, S.; Rudolfsen, G.; Folstad, I. Factors predicting male fertilization success in an external fertilizer. Behav. Ecol. Sociobiol. 2008, 62, 1805–1811. [Google Scholar] [CrossRef]
- Litvak, M.K.; Trippel, E. Sperm motility patterns of Atlantic cod (Gadus morhua) in relation to salinity: Effects of ovarian fluid and egg presence. Can. J. Fish. Aquat. Sci. 1998, 55, 1871–1877. [Google Scholar] [CrossRef]
- Turner, E.; Montgomerie, R. Ovarian fluid enhances sperm movement in Arctic charr. J. Fish Biol. 2002, 60, 1570–1579. [Google Scholar] [CrossRef]
- Rosengrave, P.; Montgomerie, R.; Metcalf, V.; McBride, K.; Gemmell, N.J. Sperm traits in Chinook salmon depend upon activation medium: Implications for studies of sperm competition in fishes. Can. J. Zool. 2009, 87, 920–927. [Google Scholar] [CrossRef]
- Elofsson, H.; Van Look, K.J.W.; Sundell, K.; Sundh, H.; Borg, B. Stickleback sperm saved by salt in ovarian fluid. J. Exp. Biol. 2006, 209, 4230–4237. [Google Scholar] [CrossRef] [Green Version]
- Alavi, S.M.H.; Cosson, J.; Bondarenko, O.; Linhart, O. Sperm motility in fishes: (III) diversity of regulatory signals from membrane to the axoneme. Theriogenology 2019, 136, 143–165. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Cosson, J. Sperm motility in fishes. (II) Effects of ions and osmolality: A review. Cell Biol. Int. 2006, 30, 1–14. [Google Scholar] [CrossRef]
- Ward, G.; Brokaw, C.J.; Garbers, D.L.; Vacquier, V.D. Chemotaxis of Arbacia punctulata spermatozoa to resact, a peptide from the egg jelly layer. J. Cell Biol. 1985, 101, 2324–2329. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Inaba, K.; Morisawa, M. Sperm Chemotaxis during the Process of Fertilization in the Ascidians Ciona savignyi and Ciona intestinalis. Dev. Biol. 1993, 157, 497–506. [Google Scholar] [CrossRef]
- Hirohashi, N.; Alvarez, L.; Shiba, K.; Fujiwara, E.; Iwata, Y.; Mohri, T.; Inaba, K.; Chiba, K.; Ochi, H.; Supuran, C.T.; et al. Sperm from Sneaker Male Squids Exhibit Chemotactic Swarming to CO2. Curr. Biol. 2013, 23, 775–781. [Google Scholar] [CrossRef] [Green Version]
- Hay, D.E. Reproductive Biology of Pacific Herring (Clupea harengus pallasi). Can. J. Fish. Aquat. Sci. 1985, 42, s111–s126. [Google Scholar] [CrossRef]
- Oda, S.; Igarashi, Y.; Manaka, K.I.; Koibuchi, N.; Sakai-Sawada, M.; Sakai, K.; Morisawa, M.; Ohtake, H.; Shimizu, N. Sperm-Activating Proteins Obtained from the Herring Eggs Are Homologous to Trypsin Inhibitors and Synthesized in Follicle Cells. Dev. Biol. 1998, 204, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Yanagimachi, R.; Cherr, G.; Matsubara, T.; Andoh, T.; Harumi, T.; Vines, C.; Pillai, M.; Griffin, F.; Matsubara, H.; Weatherby, T.; et al. Sperm Attractant in the Micropyle Region of Fish and Insect Eggs. Biol. Reprod. 2013, 88, 47. [Google Scholar] [CrossRef] [PubMed]
- Moorehead, W.R.; Biggs, H.G. 2-Amino-2-methyl-1-propanol as the alkalizing agent in an improved continuous flow cresol-phthalein complexone procedure for calcium in serum. Clin. Chem. 1974, 20, 1458–1460. [Google Scholar] [CrossRef]
- Wilson-Leedy, J.G.; Ingermann, R.L. Development of a novel CASA system based on open source software for characterization of zebrafish sperm motility parameters. Theriogenology 2007, 67, 661–672. [Google Scholar] [CrossRef]
- Purchase, C.F.; Earle, P.T. Modifications to the imagej computer assisted sperm analysis plugin greatly improve efficiency and fundamentally alter the scope of attainable data. J. Appl. Ichthyol. 2012, 28, 1013–1016. [Google Scholar] [CrossRef]
- Eisenbach, M. Sperm chemotaxis. Rev. Reprod. 1999, 4, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Cosson, J.; Billard, R.; Cibert, C.; Dreanno, C.; Linhart, O.; Suquet, M. Movements of fish sperm flagella studied by high speed videomicroscopy coupled to computer assisted image analysis. Pol. Arch. Hydrobiol. 1997, 44, 103–113. [Google Scholar]
- Inoue, T. TI Workbench, an integrated software package for electrophysiology and imaging. Microscopy 2018, 67, 129–143. [Google Scholar] [CrossRef]
- Fitzpatrick, J.L.; Simmons, L.W.; Evans, J.P. Complex Patterns of Multivariate Selection on the Ejaculate of a Broadcast Spawning Marine Invertebrate. Evolution 2012, 66, 2451–2460. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]









| Solution | Used for Activation/Chemotaxis (Injected Fluid) Tests | Osmolarity, mOsm/L | pH | |
|---|---|---|---|---|
| Distilled water | activation/chemotaxis | ≈0 | - | |
| Tap water | activation/chemotaxis | ≈0 | - | |
| 10 mM Tris HCl buffer | activation | 10 | 8 | |
| Ovarian fluid | activation/chemotaxis | 290 | ~8 | |
| Ovarian fluid in water | 50% | activation/chemotaxis | 150 | ~8 |
| 20% | activation/chemotaxis | 60 | ~8 | |
| 10% | activation/chemotaxis | 30 | ~8 | |
| 5% | activation/chemotaxis | 15 | ~8 | |
| 2% | activation/chemotaxis | 5 | ~8 | |
| Ovarian fluid in isotonic NaCl solution | 50% | activation/chemotaxis | 290 | ~8 |
| 20% | activation/chemotaxis | 290 | ~8 | |
| 10% | activation/chemotaxis | 290 | ~8 | |
| 5% | activation/chemotaxis | 290 | ~8 | |
| 2% | activation/chemotaxis | 290 | ~8 | |
| NaCl solution (+Tris buffer) | 150 mmol/L | activation/chemotaxis | 300 | 8 |
| 75 mmol/L | activation/chemotaxis | 150 | 8 | |
| 30 mmol/L | activation/chemotaxis | 60 | 8 | |
| 15 mmol/L | activation/chemotaxis | 30 | 8 | |
| EGTA supplement to water | 5 mmol/L | activation/chemotaxis | 10 | - |
| Ca2+ supplement to water | 0.2, 1, 2, 5 mmol/L | activation/chemotaxis | 1, 3, 6, 15 | - |
| Washing of Eggs | Sperm Activation Medium | Male Fish Color | Procedure |
|---|---|---|---|
| Intact eggs | Tap water | Albino | 5 g of eggs were placed into the plastic beaker and supplemented with 8 mL of water together with 0.5 µL sperm, either from albino or from regular color male. In the case of mixed sperm, the albino and normal color male sperm were mixed together, and 0.5 mL were taken from the mixture. |
| Regular color | |||
| Albino + regular color | |||
| Eggs washed thrice with isotonic saline | Tap water | Albino | |
| Regular color | |||
| Albino + regular color | |||
| Eggs washed thrice with isotonic saline | NaCl 0.9% saline | Albino | |
| Regular color | |||
| Albino + regular color | |||
| Intact eggs | 100% ovarian fluid | Albino | |
| Regular color | |||
| Albino + regular color |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kholodnyy, V.; Dzyuba, B.; Rodina, M.; Bloomfield-Gadêlha, H.; Yoshida, M.; Cosson, J.; Boryshpolets, S. Does the Rainbow Trout Ovarian Fluid Promote the Spermatozoon on Its Way to the Egg? Int. J. Mol. Sci. 2021, 22, 9519. https://doi.org/10.3390/ijms22179519
Kholodnyy V, Dzyuba B, Rodina M, Bloomfield-Gadêlha H, Yoshida M, Cosson J, Boryshpolets S. Does the Rainbow Trout Ovarian Fluid Promote the Spermatozoon on Its Way to the Egg? International Journal of Molecular Sciences. 2021; 22(17):9519. https://doi.org/10.3390/ijms22179519
Chicago/Turabian StyleKholodnyy, Vitaliy, Borys Dzyuba, Marek Rodina, Hermes Bloomfield-Gadêlha, Manabu Yoshida, Jacky Cosson, and Sergii Boryshpolets. 2021. "Does the Rainbow Trout Ovarian Fluid Promote the Spermatozoon on Its Way to the Egg?" International Journal of Molecular Sciences 22, no. 17: 9519. https://doi.org/10.3390/ijms22179519
APA StyleKholodnyy, V., Dzyuba, B., Rodina, M., Bloomfield-Gadêlha, H., Yoshida, M., Cosson, J., & Boryshpolets, S. (2021). Does the Rainbow Trout Ovarian Fluid Promote the Spermatozoon on Its Way to the Egg? International Journal of Molecular Sciences, 22(17), 9519. https://doi.org/10.3390/ijms22179519