
Synergistic Effect of Simultaneous versus Sequential Combined Treatment of Histone Deacetylase Inhibitor Valproic Acid with Etoposide on Melanoma Cells




Abstract
:1. Introduction
2. Results
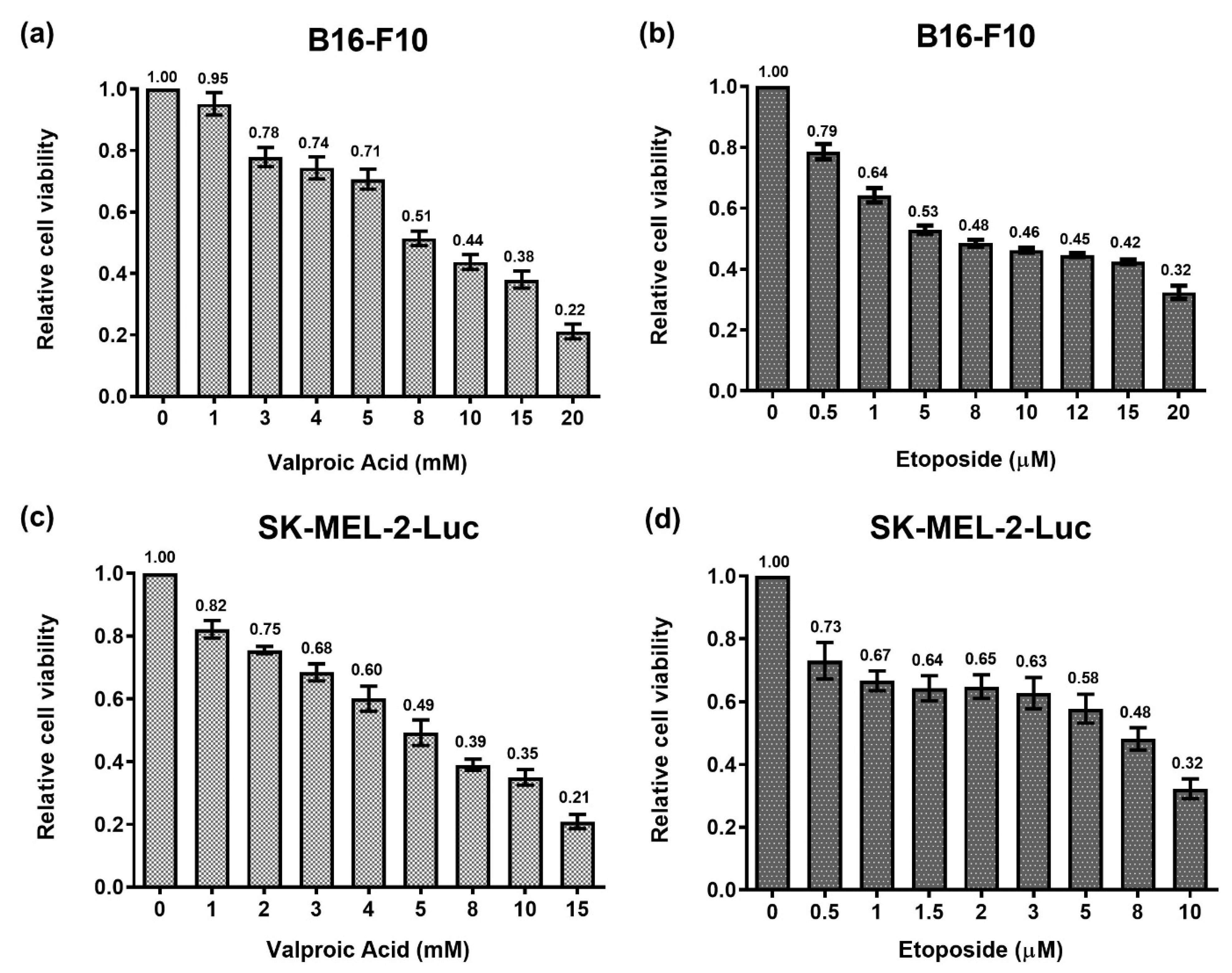
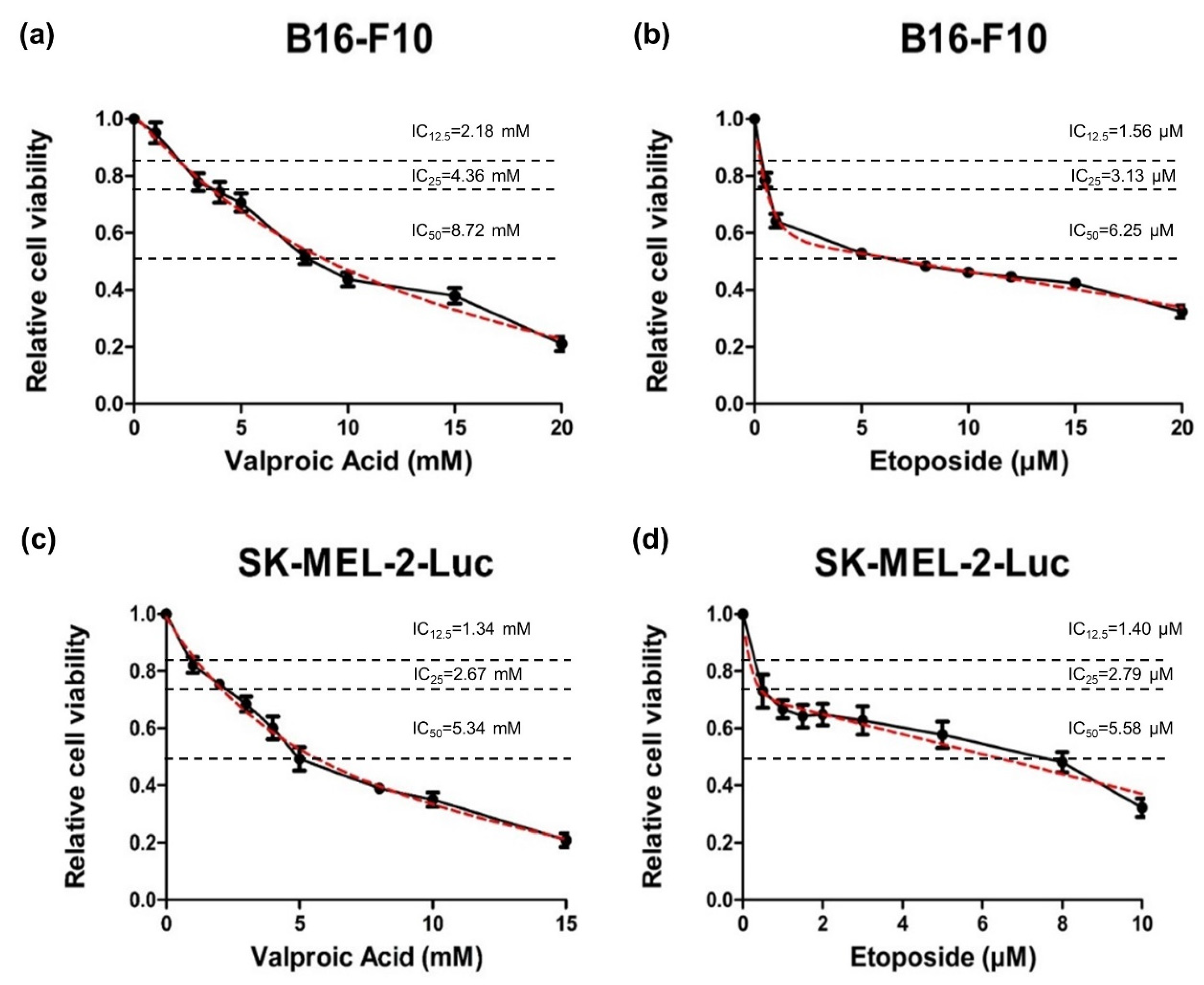
2.1. Analysis of Dose-Response Curves under Single Treatments of VPA or ETO on B16-F10 and SK-MEL-2-Luc Melanoma Cells
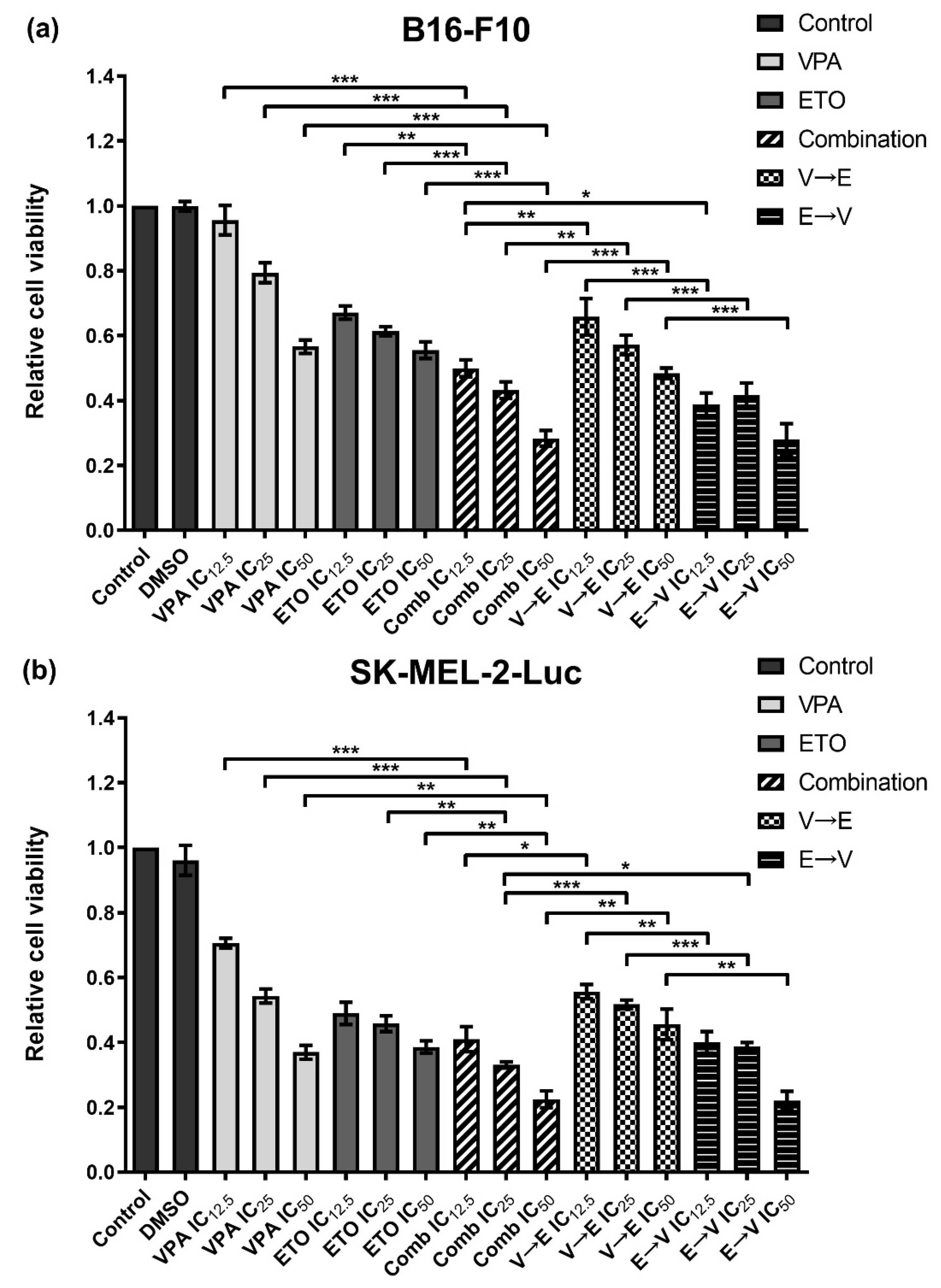
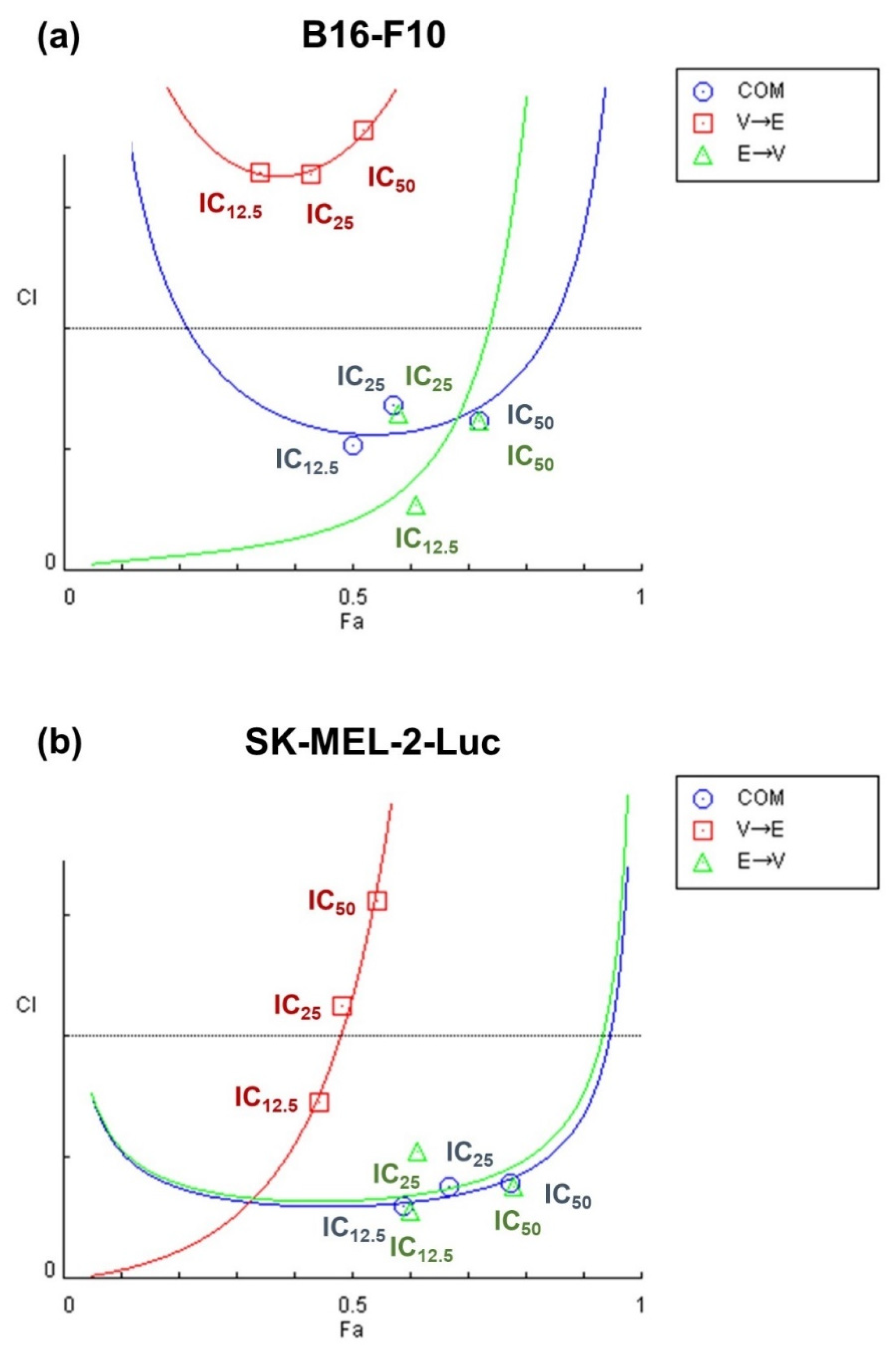
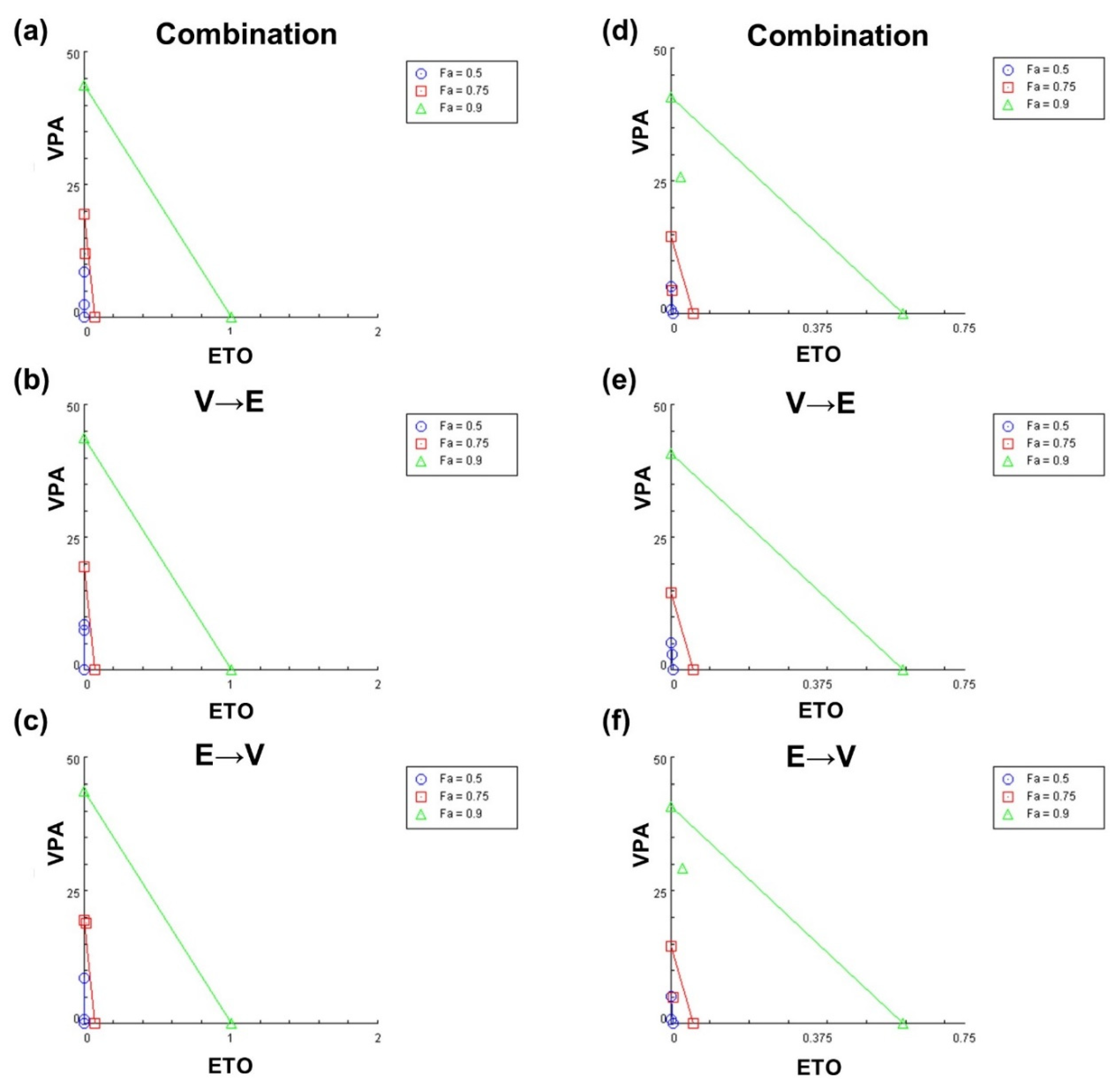
2.2. Simultaneous and Sequential Combined Treatments of VPA and ETO Contributed to Synergistic Inhibitory Effects or Antagonistic Effect
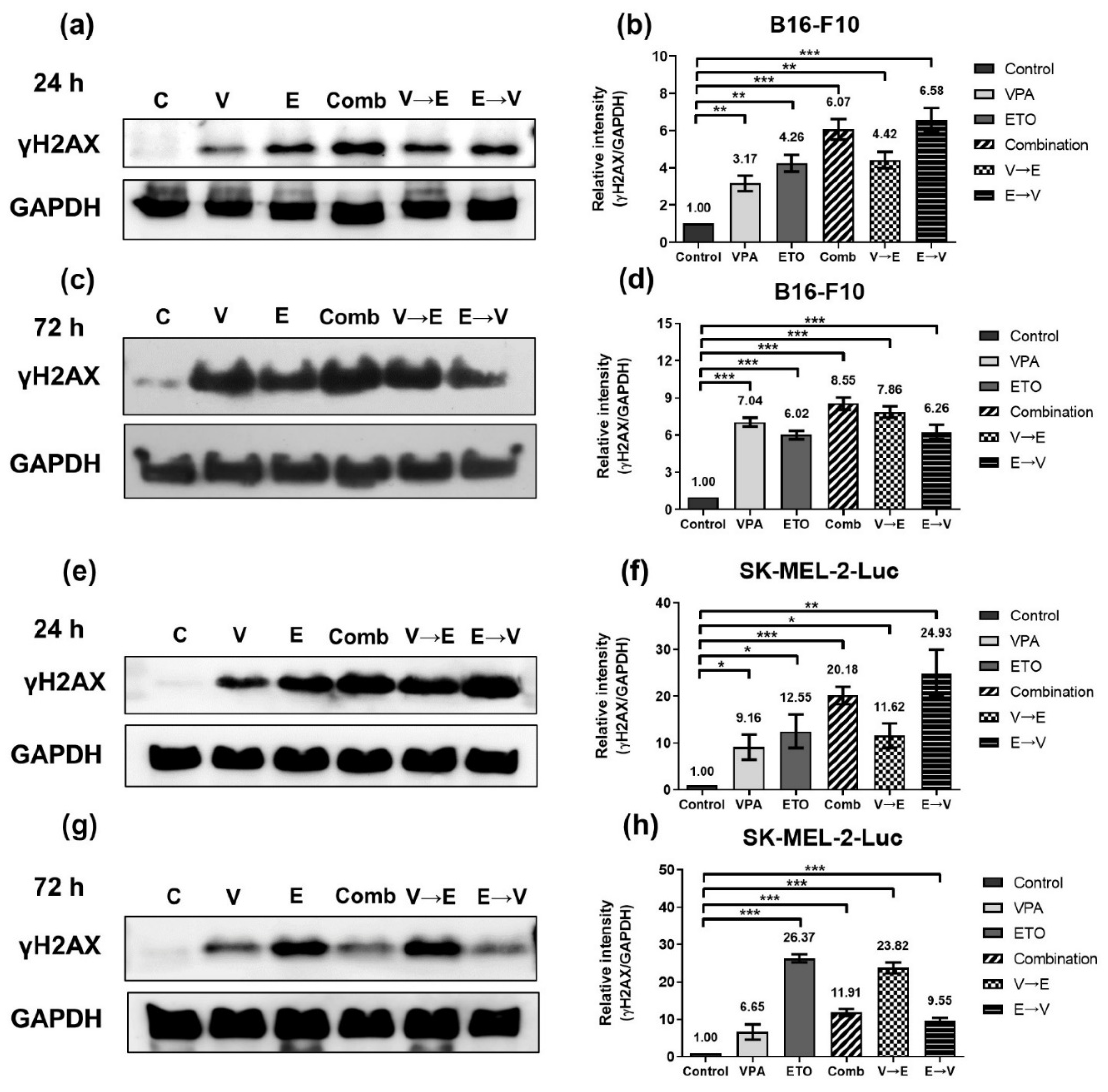
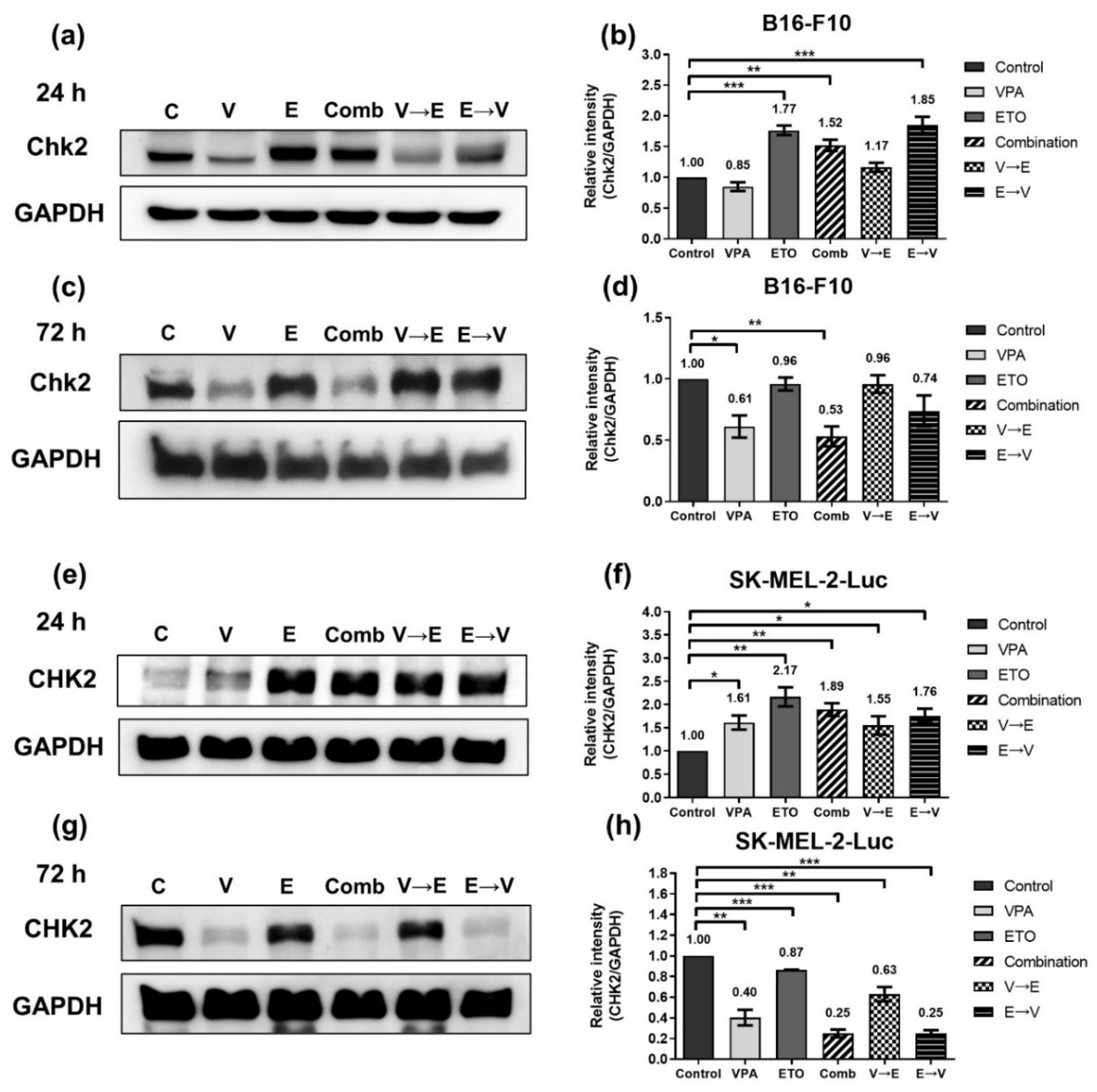
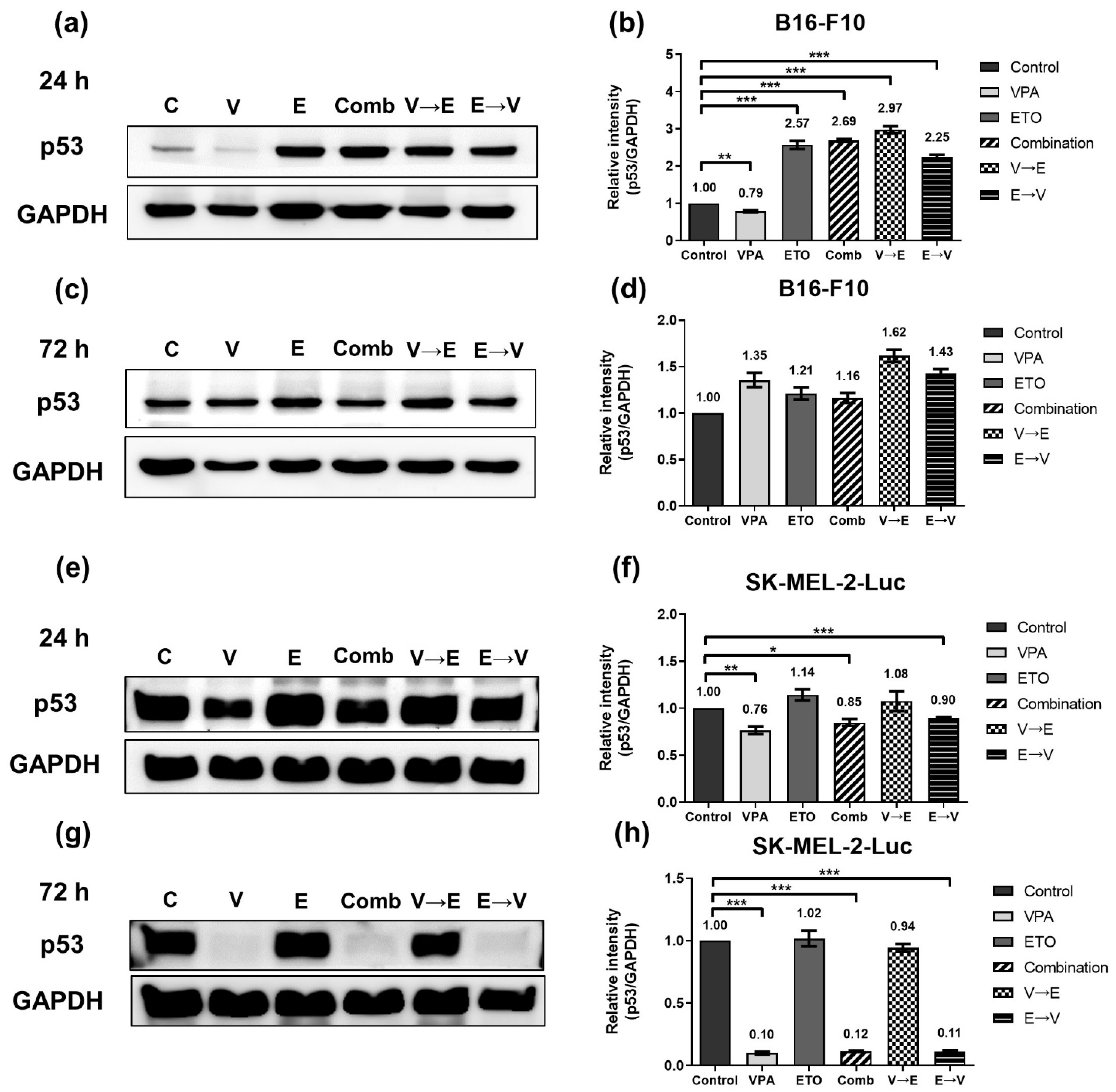
2.3. ETO Dominated the Induction of DNA Double-Strand Breaks (DSBs) under ETO→VPA Sequential Combined Treatment
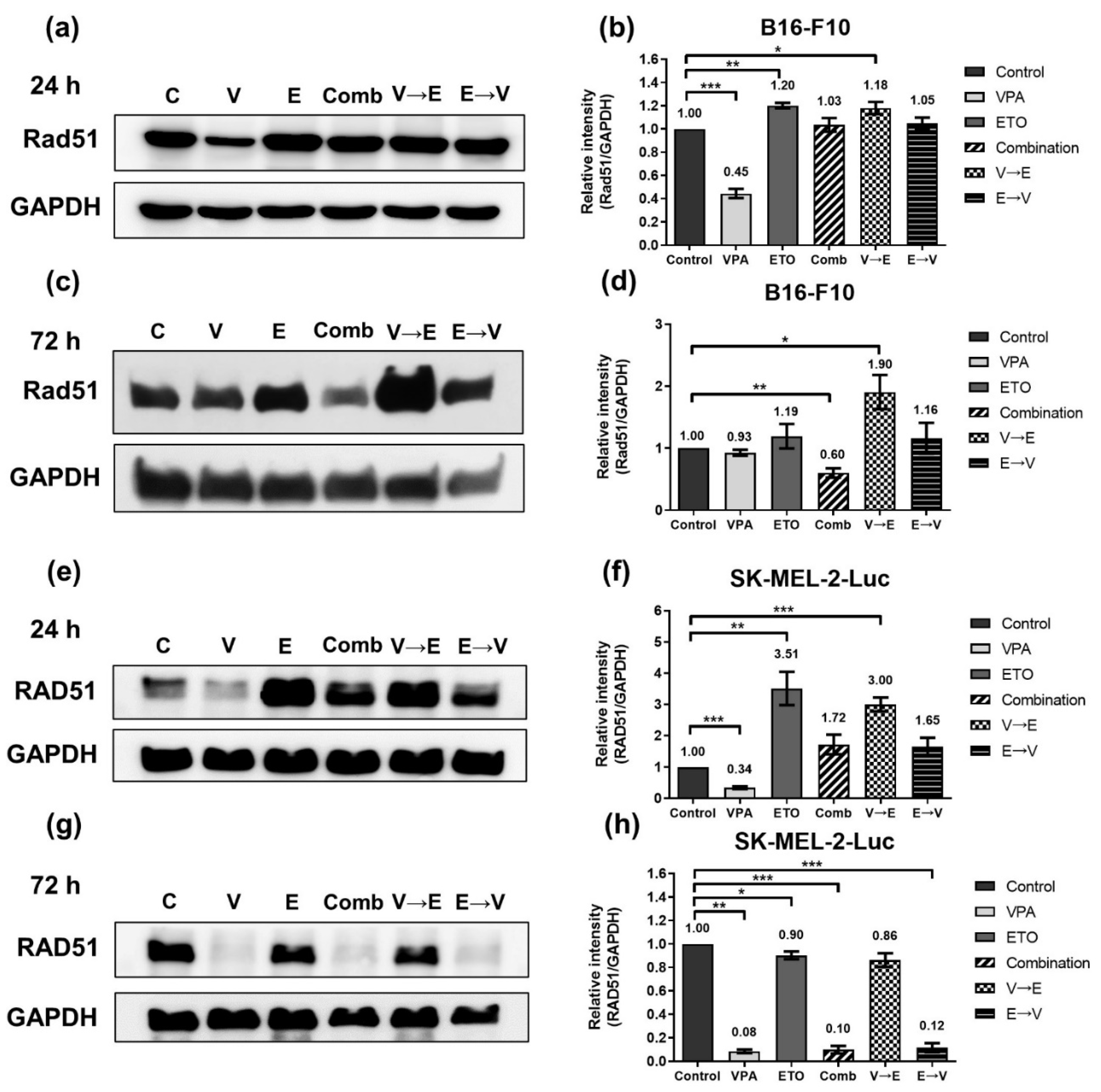
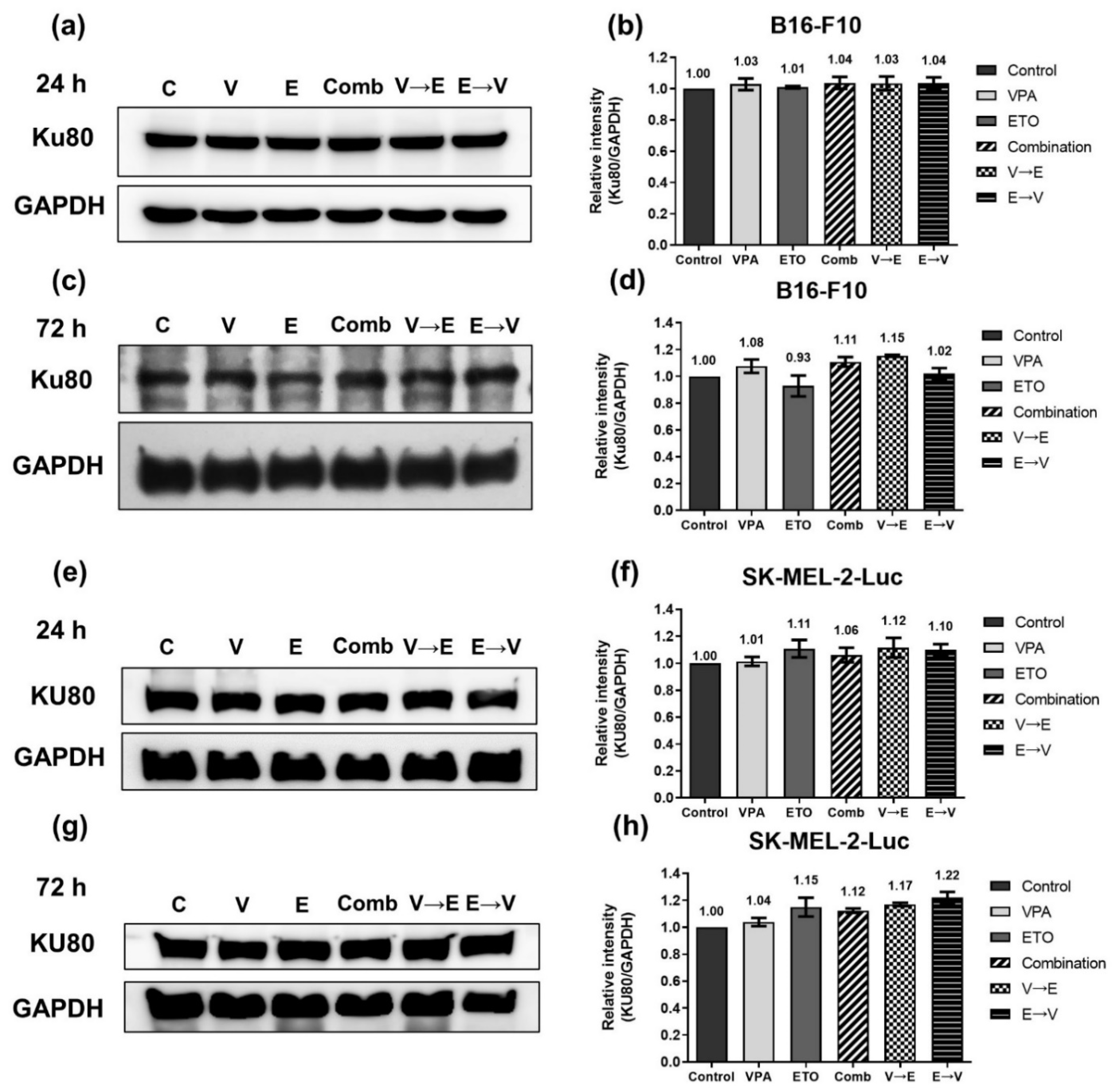
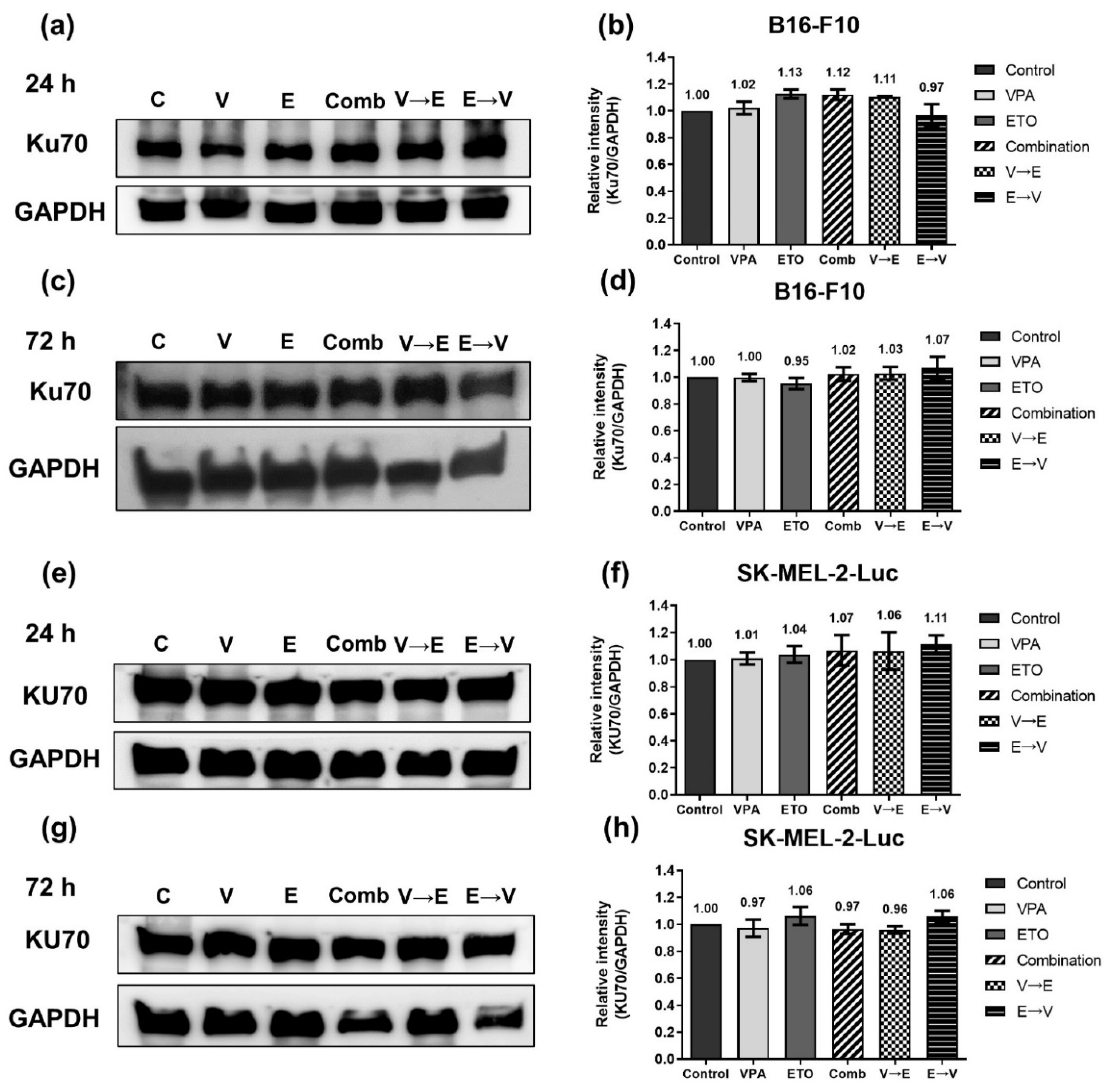
2.4. DNA DSBs Induced by VPA Pretreated Sequential Combined Treatment Were Predominantly Repaired via Homologous Recombination
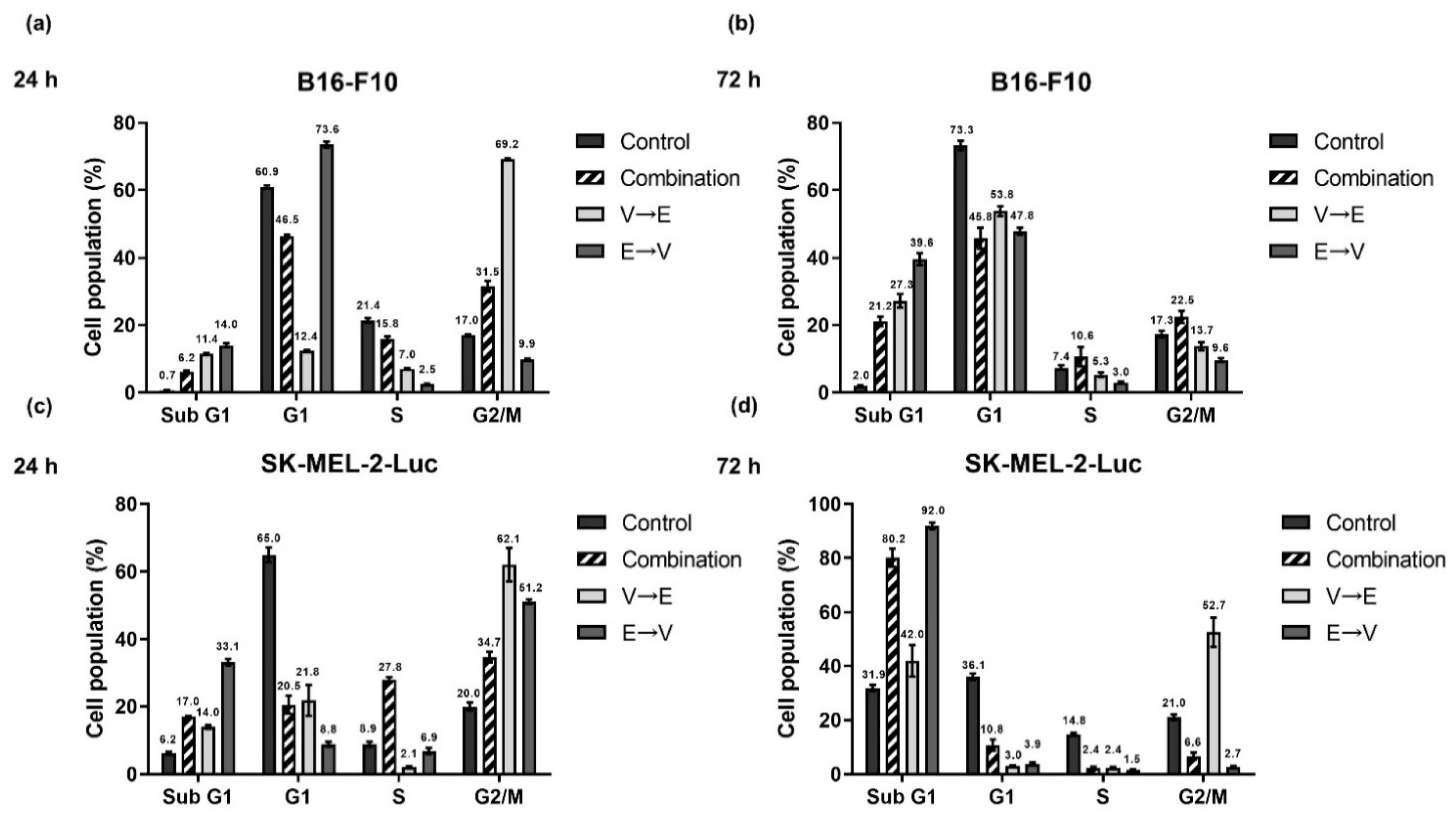
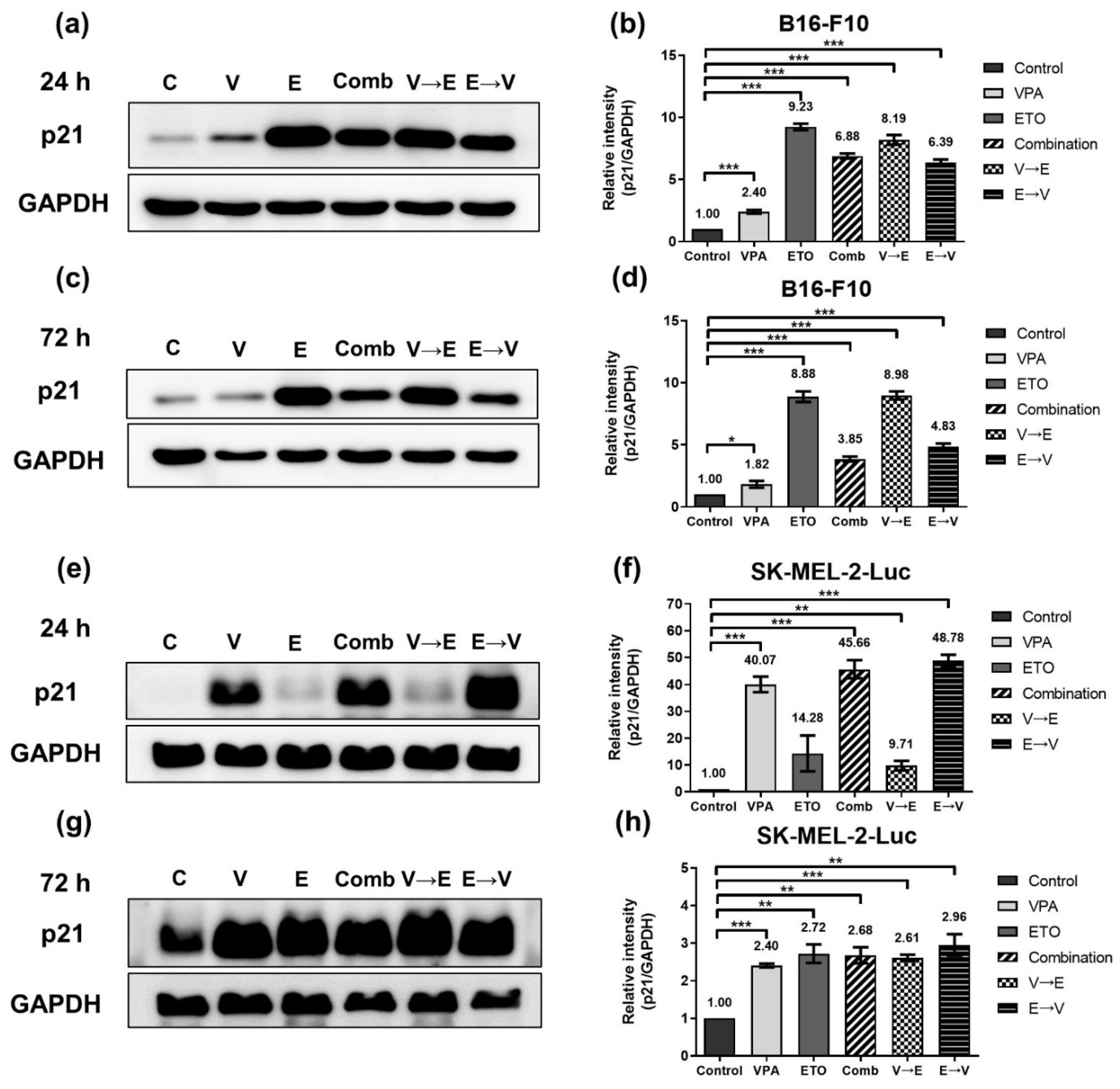
2.5. Different Sequential Orders of VPA and ETO Administration Might Influence the Cell Cycle Regulation
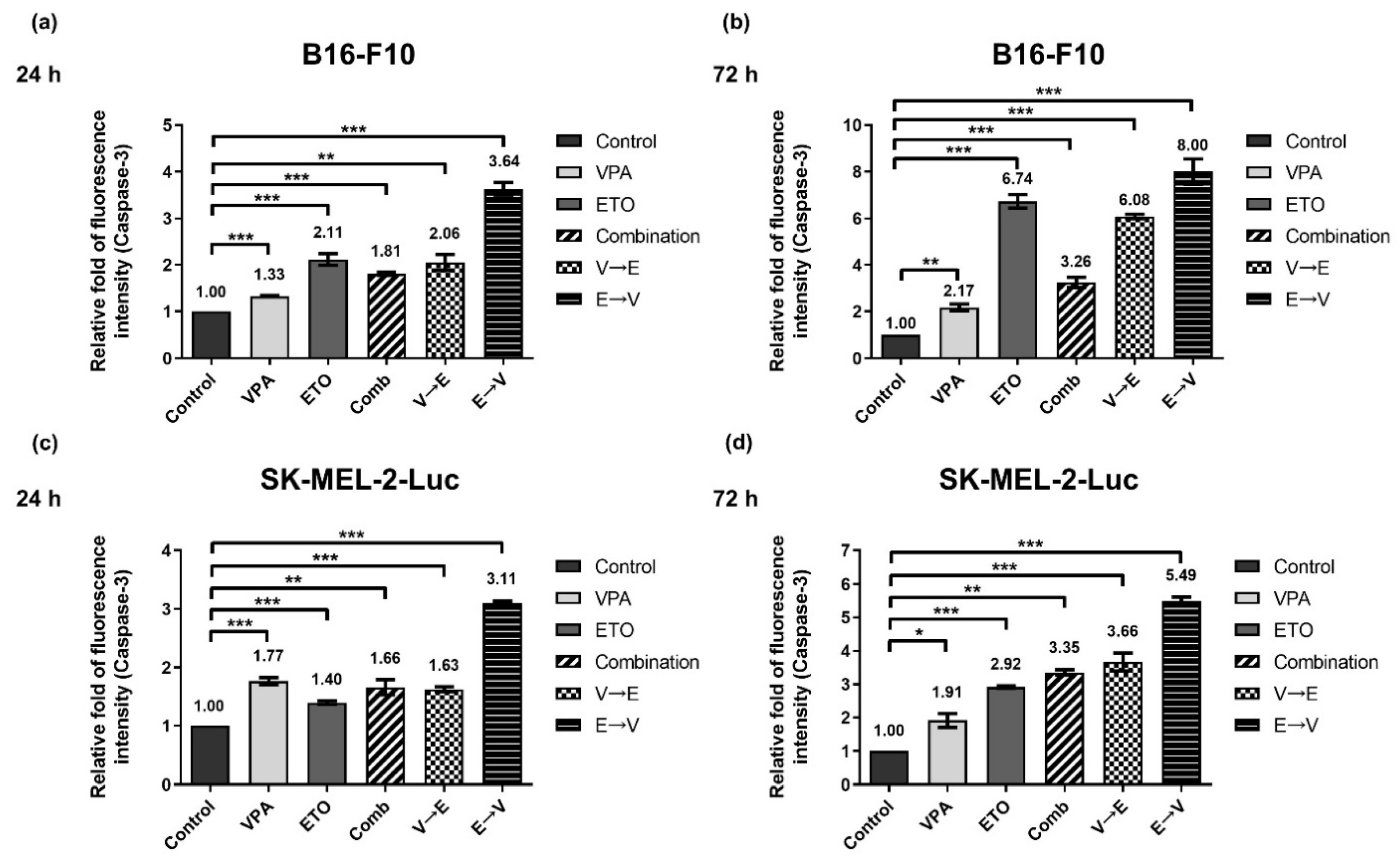
2.6. ETO Pretreated Sequential Combined Treatment Augmented Apoptosis via Caspase-3 Up-Regulation
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Drugs
4.3. Cell Viability Assay and IC50 Determination
4.4. Analysis of Combination Index
4.5. Western Blot Analysis
4.6. Cell Cycle and Caspase-3 Apoptosis Analysis
4.7. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Leonardi, G.C.; Falzone, L.; Salemi, R.; Zanghi, A.; Spandidos, D.A.; McCubrey, J.A.; Candido, S.; Libra, M. Cutaneous melanoma: From pathogenesis to therapy. Int. J. Oncol. 2018, 52, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Barker, C.A.; Lee, N.Y. Radiation therapy for cutaneous melanoma. Derm. Clin. 2012, 30, 525–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Singh, R.K. Resistance to chemotherapy and molecularly targeted therapies: Rationale for combination therapy in malignant melanoma. Curr. Mol. Med. 2011, 11, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Komarova, N.L.; Boland, C.R. Cancer: Calculated treatment. Nature 2013, 499, 291–292. [Google Scholar] [CrossRef] [Green Version]
- Mokhtari, R.B.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [Green Version]
- Terrie, Y. Monitoring Combinationdrug Therapy. Available online: https://www.pharmacytimes.com/publications/issue/2010/january2010/rxfocuscombination-0110 (accessed on 26 May 2021).
- Valentini, A.; Gravina, P.; Federici, G.; Bernardini, S. Valproic acid induces apoptosis, p16INK4A upregulation and sensitization to chemotherapy in human melanoma cells. Cancer Biol. 2007, 6, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Vigushin, D.M.; Coombes, R.C. Histone deacetylase inhibitors in cancer treatment. Anticancer Drugs 2002, 13, 1–13. [Google Scholar] [CrossRef]
- Daud, A.I.; Dawson, J.; DeConti, R.C.; Bicaku, E.; Marchion, D.; Bastien, S.; Hausheer, F.A., III; Lush, R.; Neuger, A.; Sullivan, D.M.; et al. Potentiation of a topoisomerase I inhibitor, karenitecin, by the histone deacetylase inhibitor valproic acid in melanoma: Translational and phase I/II clinical trial. Clin. Cancer Res. 2009, 15, 2479–2487. [Google Scholar] [CrossRef] [Green Version]
- Kalal, B.S.; Pai, V.R.; Behera, S.K.; Somashekarappa, H.M. HDAC2 inhibitor valproic acid increases radiation sensitivity of drug-resistant melanoma cells. Med. Sci. 2019, 7, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudolf, K.; Cervinka, M.; Rudolf, E. Cytotoxicity and mitochondrial apoptosis induced by etoposide in melanoma cells. Cancer Investig. 2009, 27, 704–717. [Google Scholar] [CrossRef] [PubMed]
- Eckschlager, T.; Plch, J.; Stiborova, M.; Hrabeta, J. Histone deacetylase inhibitors as anticancer drugs. Int. J. Mol. Sci. 2017, 18, 1414. [Google Scholar] [CrossRef]
- Das, C.M.; Aguilera, D.; Vasquez, H.; Prasad, P.; Zhang, M.; Wolff, J.E.; Gopalakrishnan, V. Valproic acid induces p21 and topoisomerase-II (alpha/beta) expression and synergistically enhances etoposide cytotoxicity in human glioblastoma cell lines. J. Neuro-Oncol. 2007, 85, 159–170. [Google Scholar] [CrossRef]
- Rocca, A.; Minucci, S.; Tosti, G.; Croci, D.; Contegno, F.; Ballarini, M.; Nole, F.; Munzone, E.; Salmaggi, A.; Goldhirsch, A.; et al. A phase I-II study of the histone deacetylase inhibitor valproic acid plus chemoimmunotherapy in patients with advanced melanoma. Br. J. Cancer 2009, 100, 28–36. [Google Scholar] [CrossRef] [Green Version]
- GraphPad Prism 8 Curve Fitting Guide—Equation: Two Phase Association. Available online: https://www.graphpad.com/guides/prism/8/curve-fitting/reg_exponential_association_2phase.htm (accessed on 26 May 2021).
- Chou, T.C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzym. Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Chou, T.C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharm. Rev. 2006, 58, 621–681. [Google Scholar] [PubMed]
- Montecucco, A.; Zanetta, F.; Biamonti, G. Molecular mechanisms of etoposide. EXCLI J. 2015, 14, 95–108. [Google Scholar] [PubMed]
- Taylor, W.R.; Stark, G.R. Regulation of the G2/M transition by p53. Oncogene 2001, 20, 1803–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsarra, I.A.; Al-Omar, M.; Belal, F. Valproic Acid and sodium valproate: Comprehensive profile. Profiles Drug Subst. Excip. Relat. Methodol. 2005, 32, 209–240. [Google Scholar] [PubMed]
- Brandsma, I.; Gent, D.C. Pathway choice in DNA double strand break repair: Observations of a balancing act. Genome Integr. 2012, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Krumm, A.; Barckhausen, C.; Kucuk, P.; Tomaszowski, K.H.; Loquai, C.; Fahrer, J.; Kramer, O.H.; Kaina, B.; Roos, W.P. Enhanced histone deacetylase activity in malignant melanoma provokes RAD51 and FANCD2-triggered drug resistance. Cancer Res. 2016, 76, 3067–3077. [Google Scholar] [CrossRef] [Green Version]
- Shaltiel, I.A.; Krenning, L.; Bruinsma, W.; Medema, R.H. The same, only different—DNA damage checkpoints and their reversal throughout the cell cycle. J. Cell Sci. 2015, 128, 607–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lizard, G.; Miguet, C.; Gueldry, S.; Monier, S.; Gambert, P. Flow cytometry measurement of DNA fragmentation in the course of cell death via apoptosis. New techniques for evaluation of DNA status for the pathologist. Ann. Pathol. 1997, 17, 61–66. [Google Scholar] [PubMed]
- Chen, K.H.; Lai, Z.Y.; Li, D.Y.; Lin, Y.C.; Chou, F.I.; Chuang, Y.J. Analysis of DNA Damage Responses After Boric Acid-mediated Boron Neutron Capture Therapy in Hepatocellular Carcinoma. Anticancer Res. 2019, 39, 6661–6671. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | B16-F10 | |
| Single-drug treatment | Valproic acid (VPA) | Etoposide (ETO) |
| IC50 value | 8.72 | 6.25 |
| Unit | mM | μM |
| Cell Line | SK-MEL-2-Luc | |
| Single-drug treatment | Valproic acid (VPA) | Etoposide (ETO) |
| IC50 value | 5.34 | 5.58 |
| Unit | mM | μM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shyu, Y.-M.; Liu, L.Y.-M.; Chuang, Y.-J. Synergistic Effect of Simultaneous versus Sequential Combined Treatment of Histone Deacetylase Inhibitor Valproic Acid with Etoposide on Melanoma Cells. Int. J. Mol. Sci. 2021, 22, 10029. https://doi.org/10.3390/ijms221810029
Shyu Y-M, Liu LY-M, Chuang Y-J. Synergistic Effect of Simultaneous versus Sequential Combined Treatment of Histone Deacetylase Inhibitor Valproic Acid with Etoposide on Melanoma Cells. International Journal of Molecular Sciences. 2021; 22(18):10029. https://doi.org/10.3390/ijms221810029
Chicago/Turabian StyleShyu, Yueh-Ming, Lawrence Yu-Min Liu, and Yung-Jen Chuang. 2021. "Synergistic Effect of Simultaneous versus Sequential Combined Treatment of Histone Deacetylase Inhibitor Valproic Acid with Etoposide on Melanoma Cells" International Journal of Molecular Sciences 22, no. 18: 10029. https://doi.org/10.3390/ijms221810029
APA StyleShyu, Y. -M., Liu, L. Y. -M., & Chuang, Y. -J. (2021). Synergistic Effect of Simultaneous versus Sequential Combined Treatment of Histone Deacetylase Inhibitor Valproic Acid with Etoposide on Melanoma Cells. International Journal of Molecular Sciences, 22(18), 10029. https://doi.org/10.3390/ijms221810029