
Dehydrocostus Lactone Attenuates Methicillin-Resistant Staphylococcus aureus-Induced Inflammation and Acute Lung Injury via Modulating Macrophage Polarization

 ,
,
Abstract
:1. Introduction
2. Results
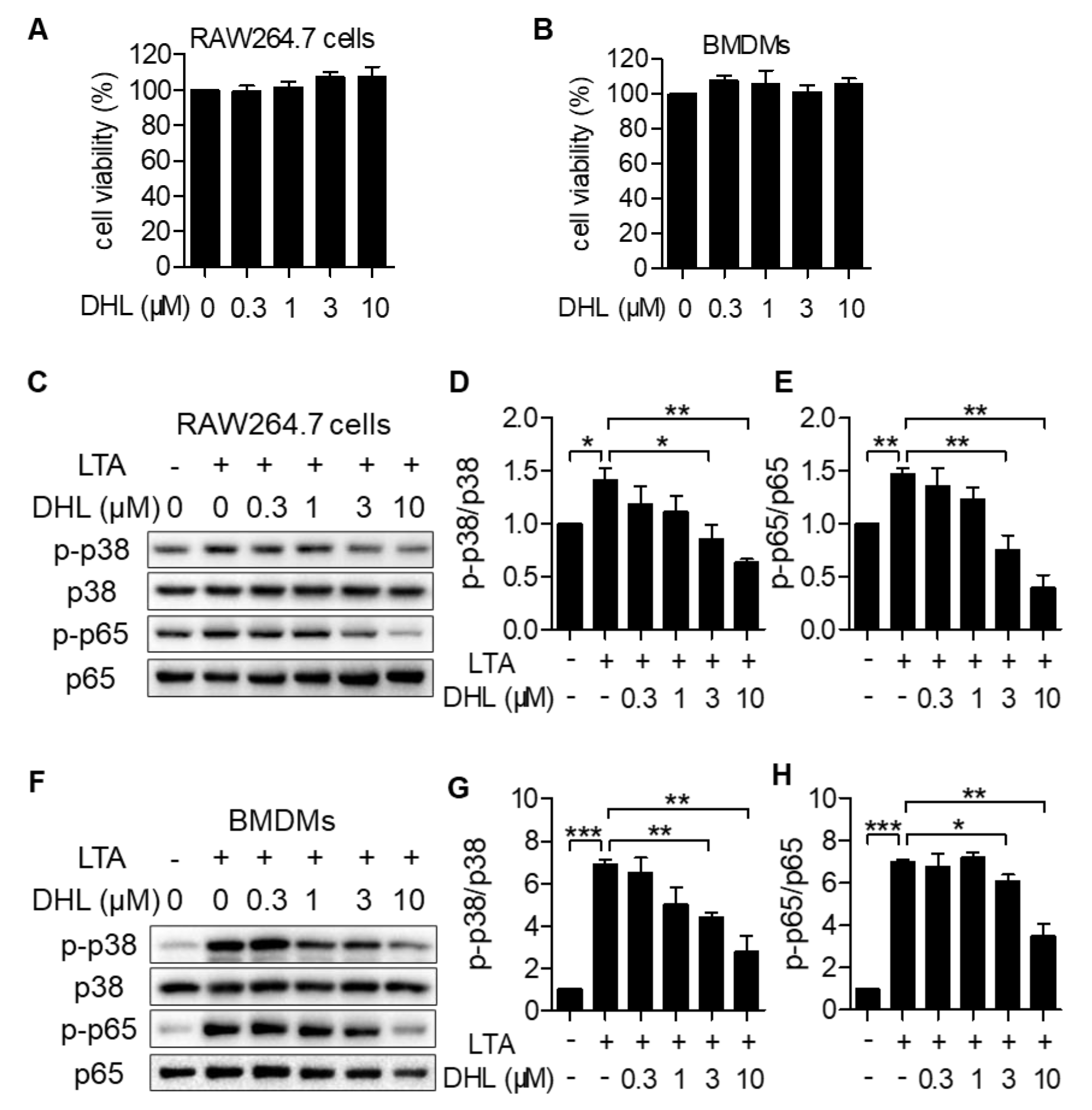
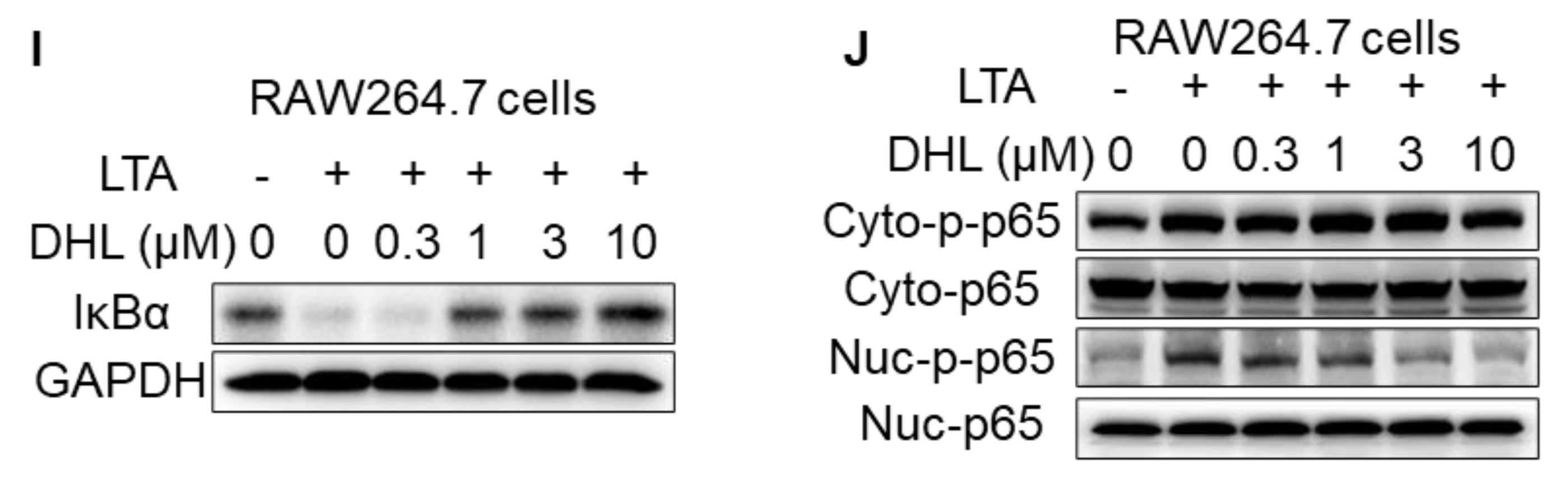
2.1. DHL Attenuates LTA-Induced Phosphorylation of p38 MAPK and NF-κB in RAW264.7 Cells and BMDMs
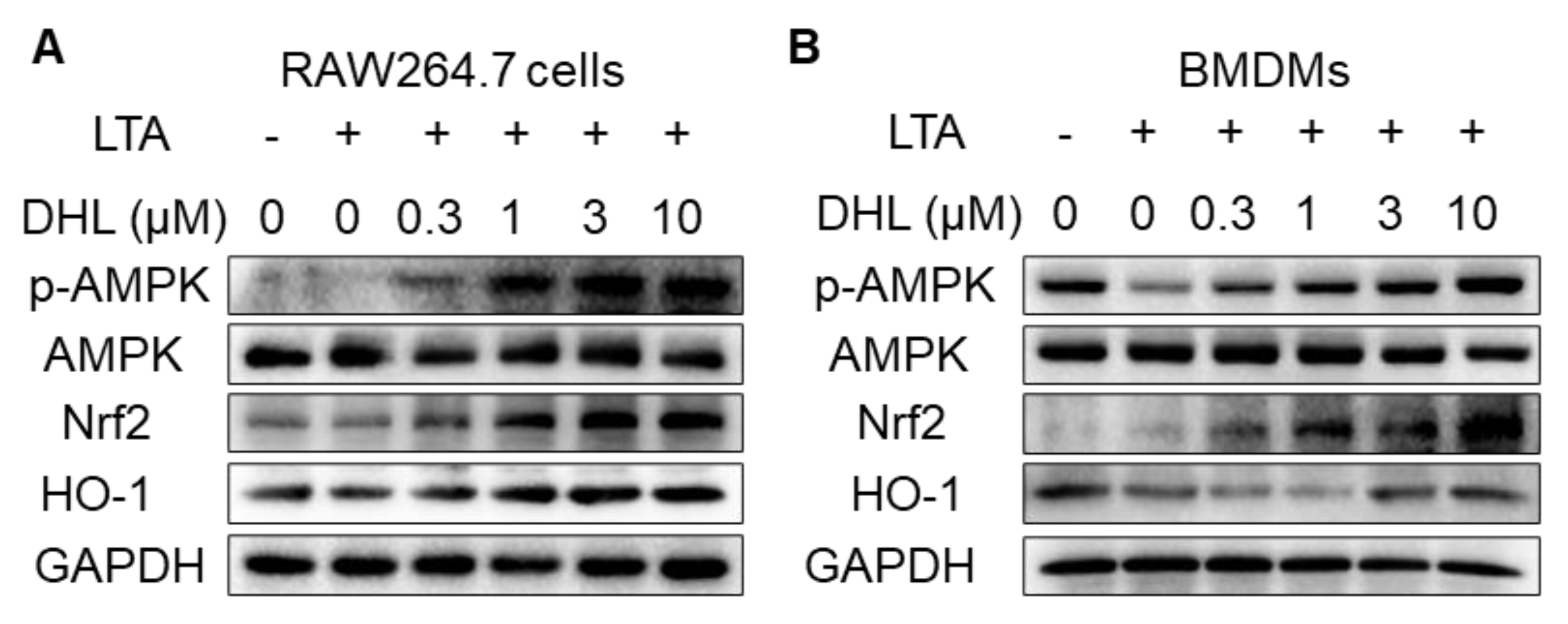
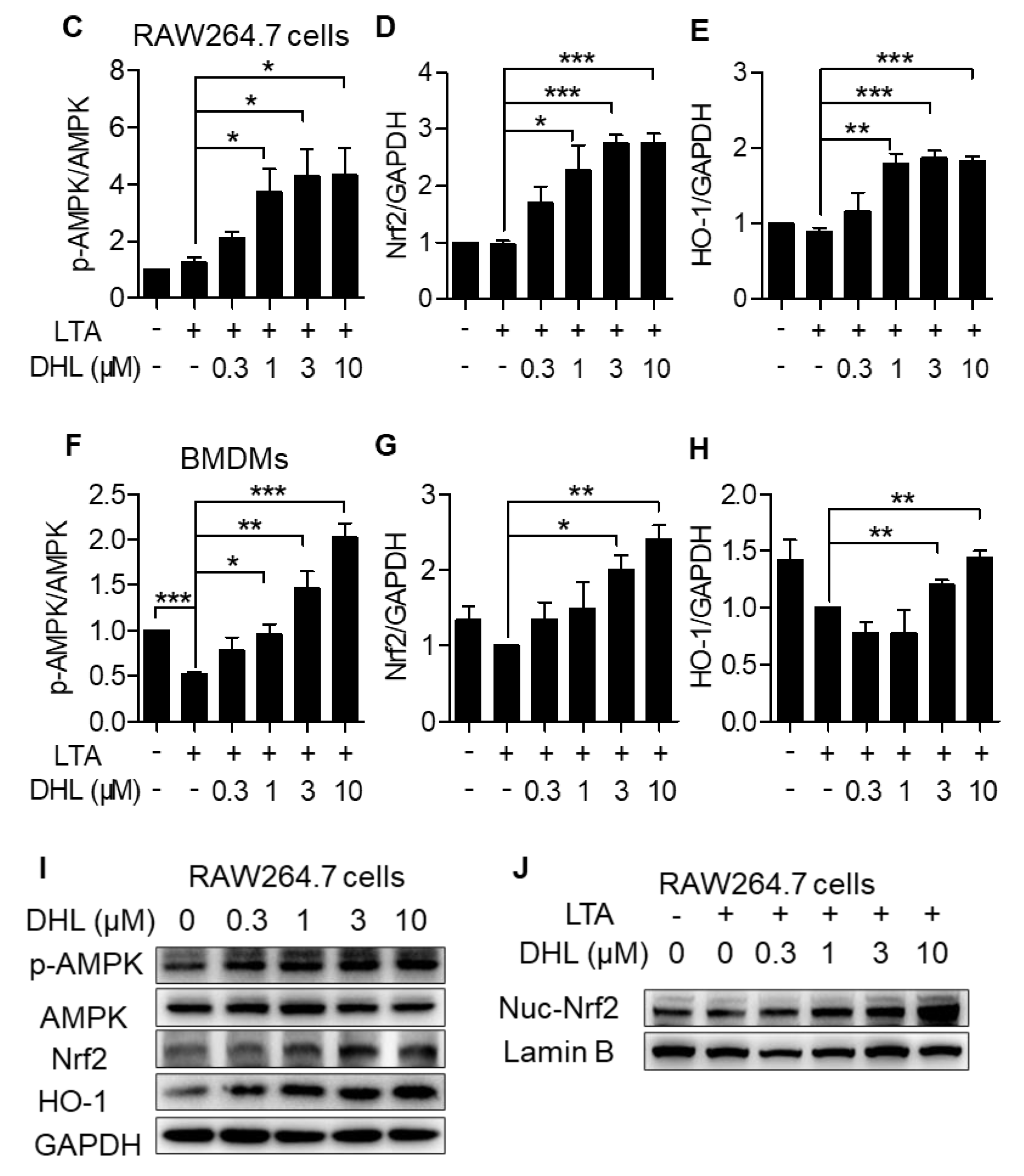
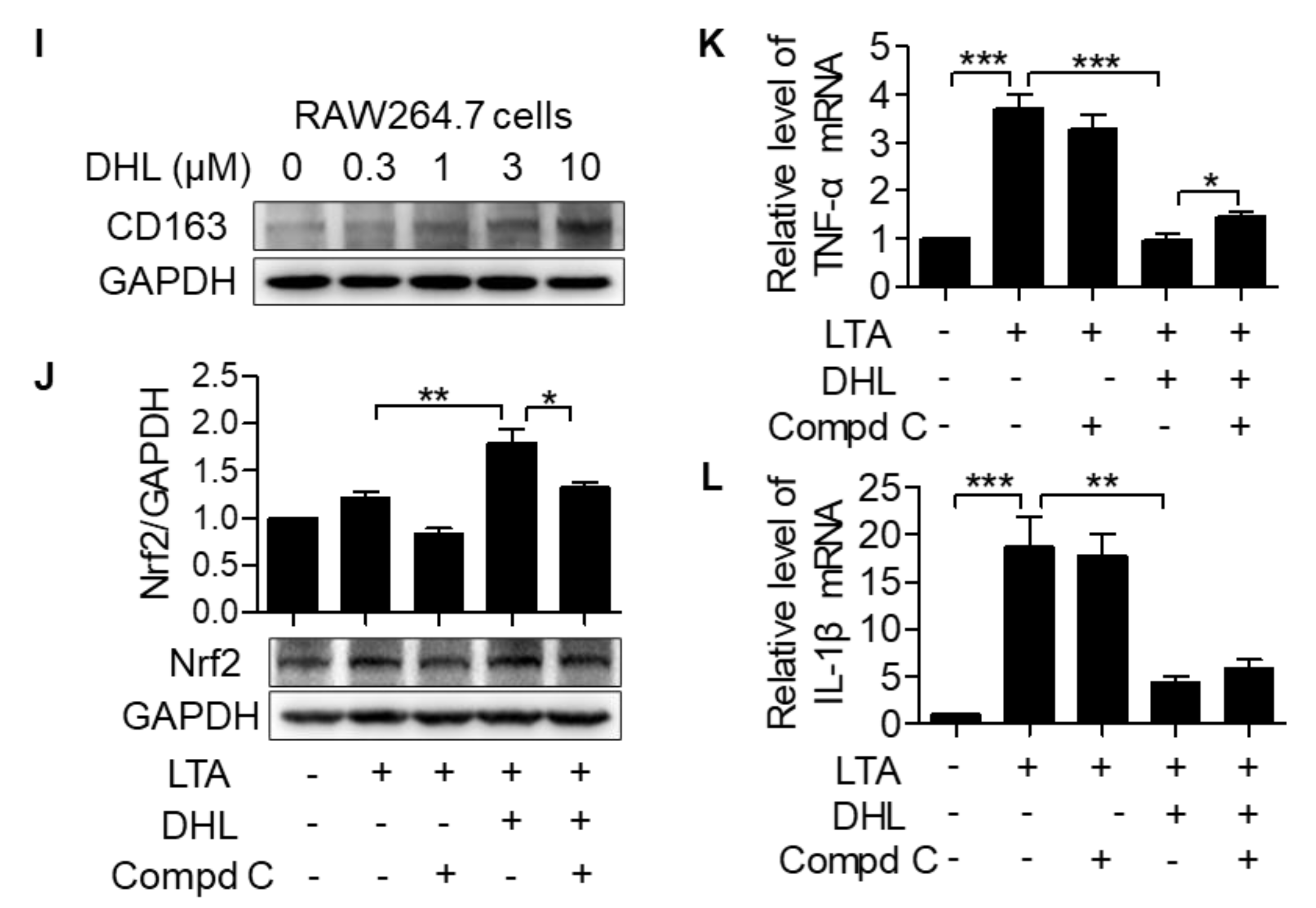
2.2. DHL Promotes the Activation of AMPK/Nrf2 Pathway in LTA-Induced Macrophages
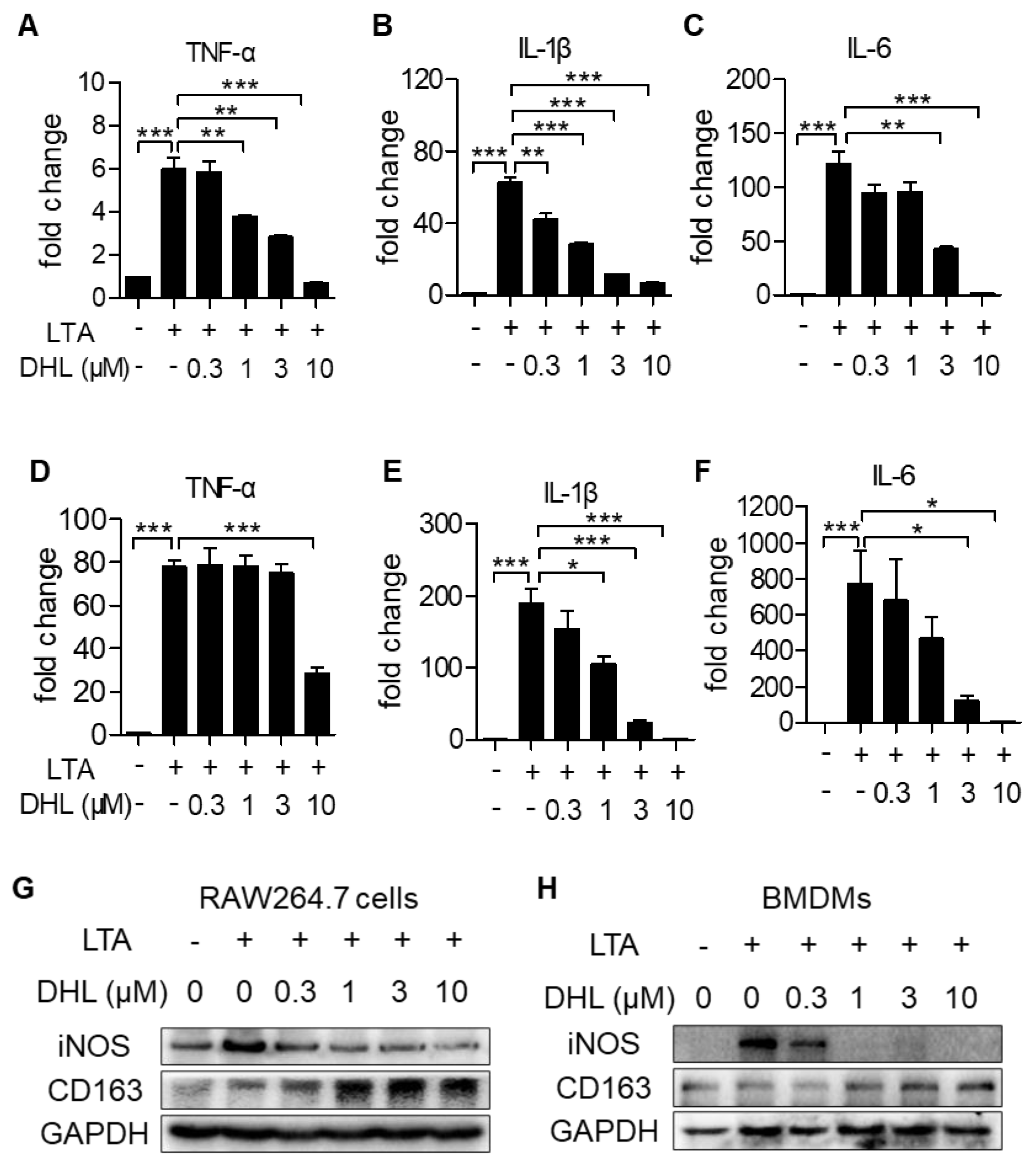
2.3. DHL Suppresses M1 Phenotype Macrophage Activation While Accelerating M2 Macrophage Polarization in Macrophages
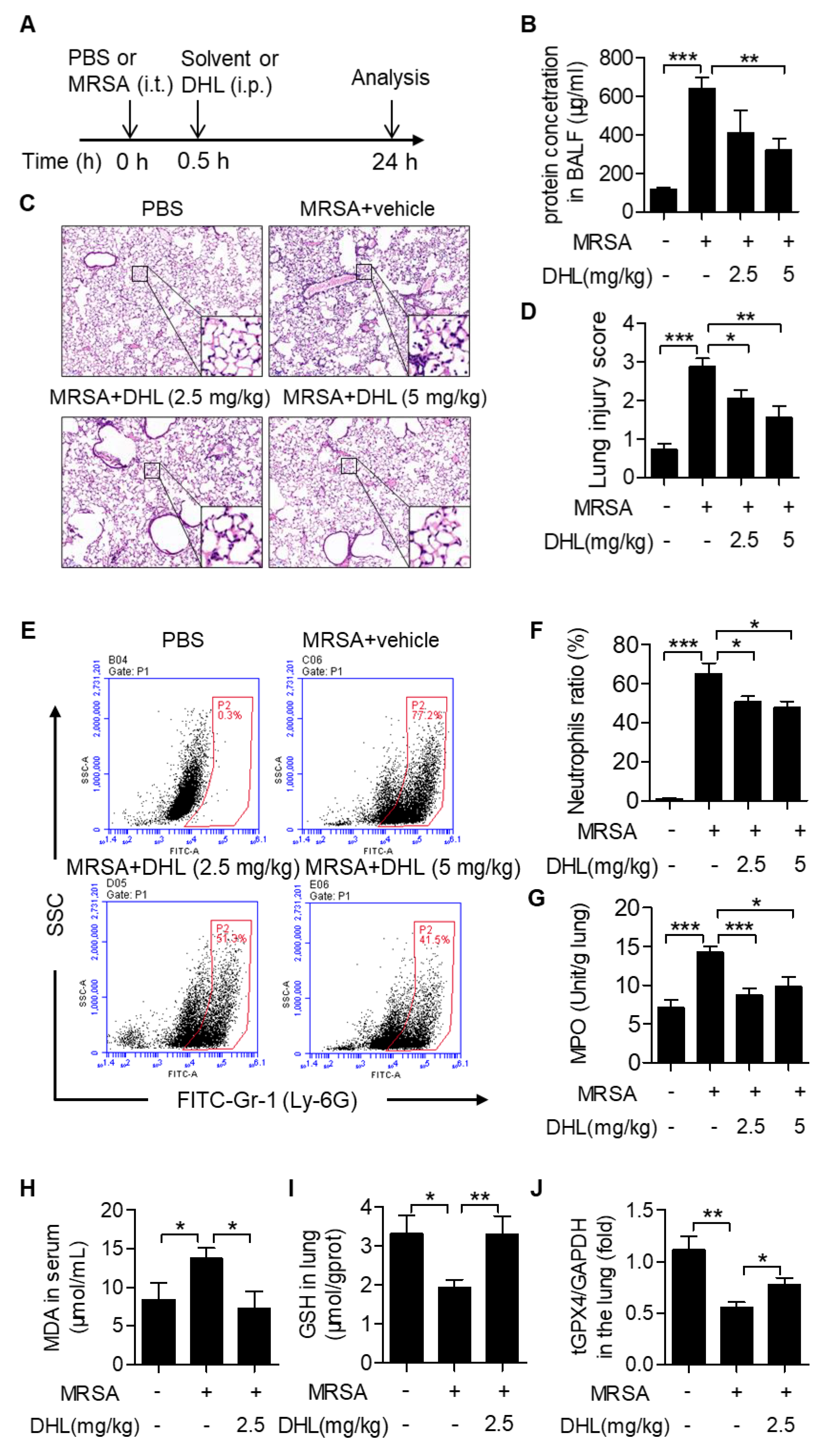
2.4. DHL Ameliorates MRSA-Induced ALI in Mice
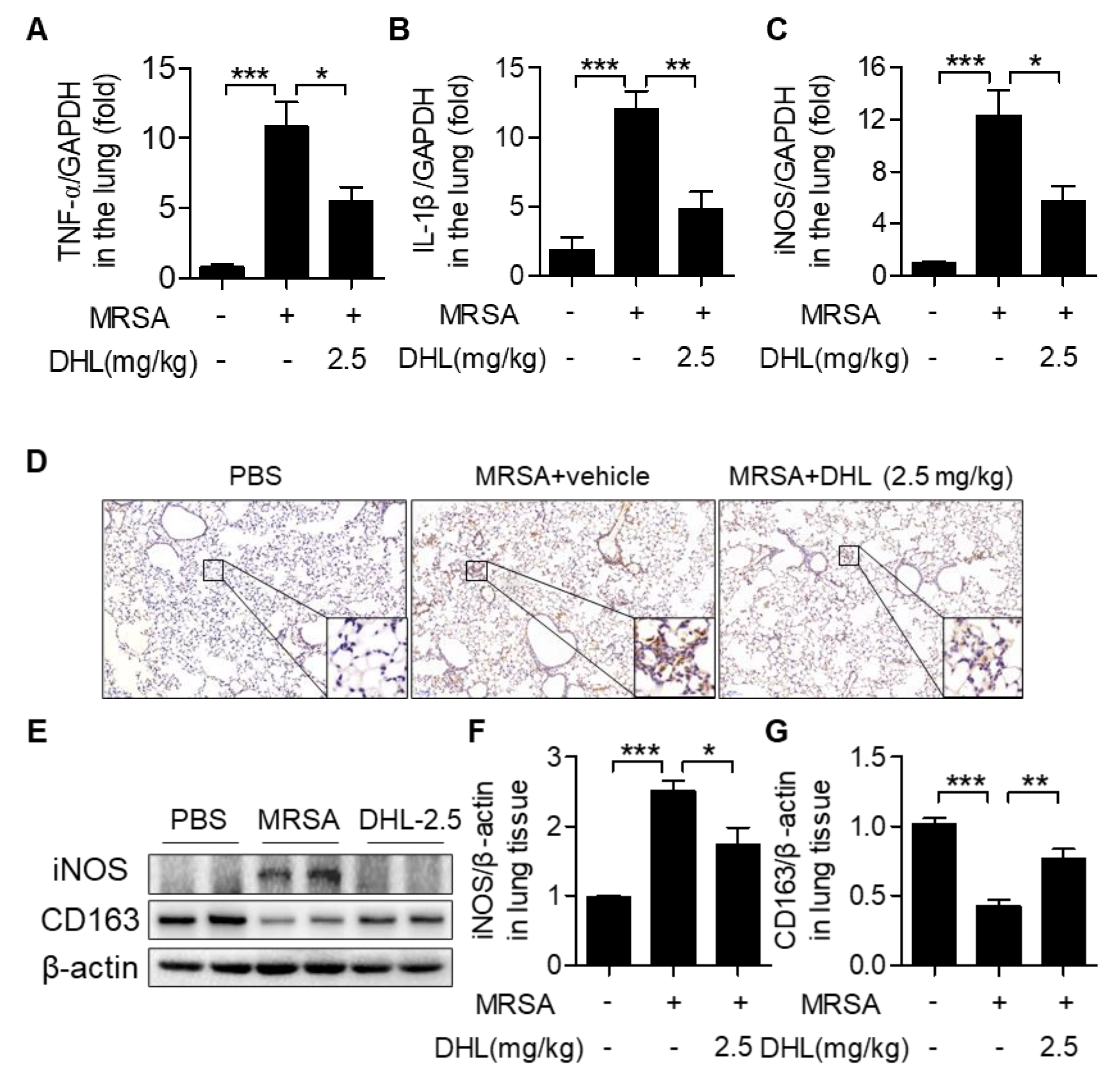
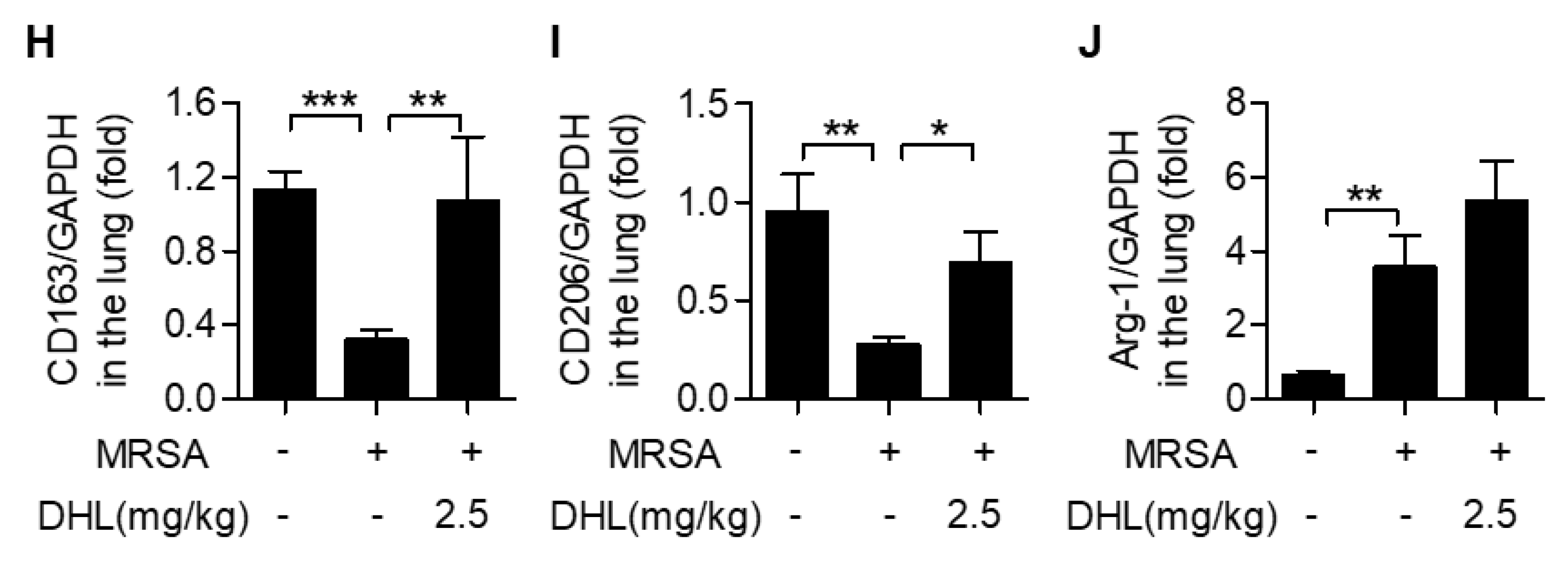
2.5. DHL Modulates Macrophage Polarization in MRSA-Induced ALI in Mice
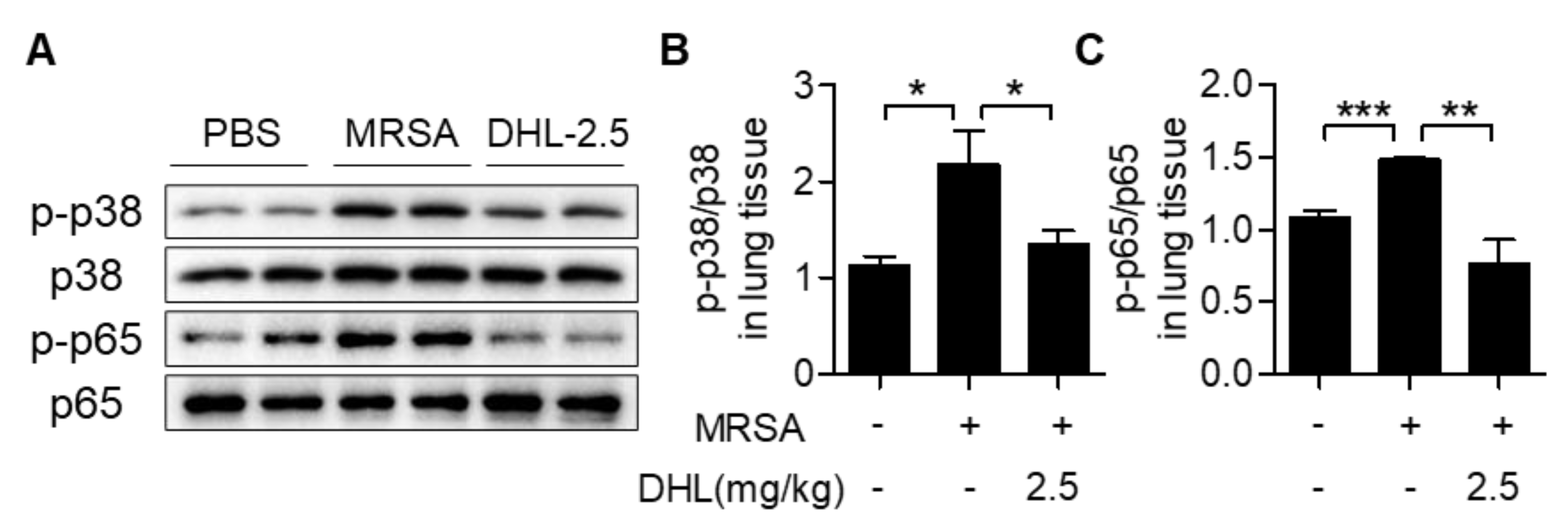
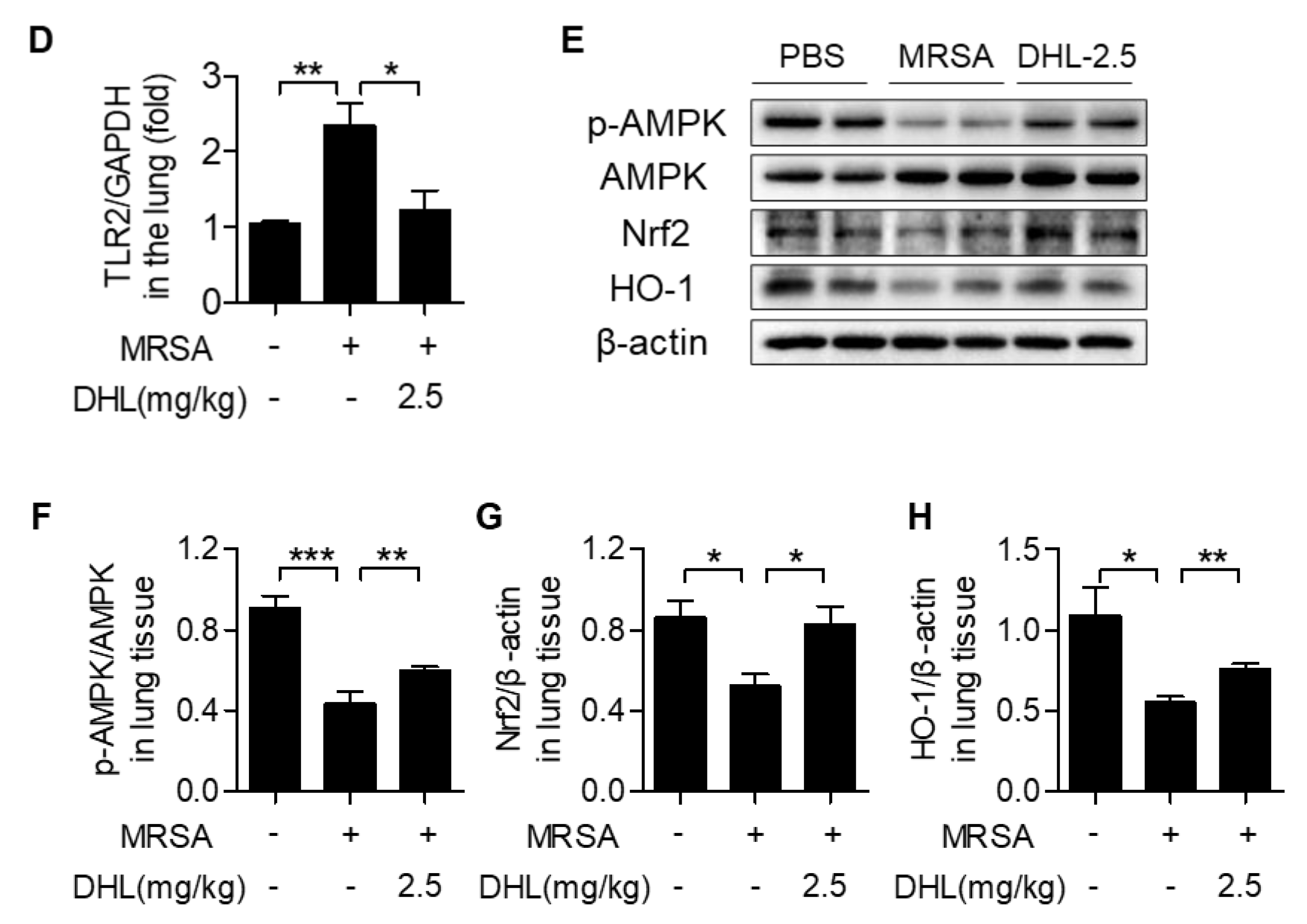
2.6. DHL Inhibits the Phosphorylation of p38 MAPK and NF-κB, but Promotes the Activation of the AMPK/Nrf2 Pathway in an MRSA-Induced ALI Model
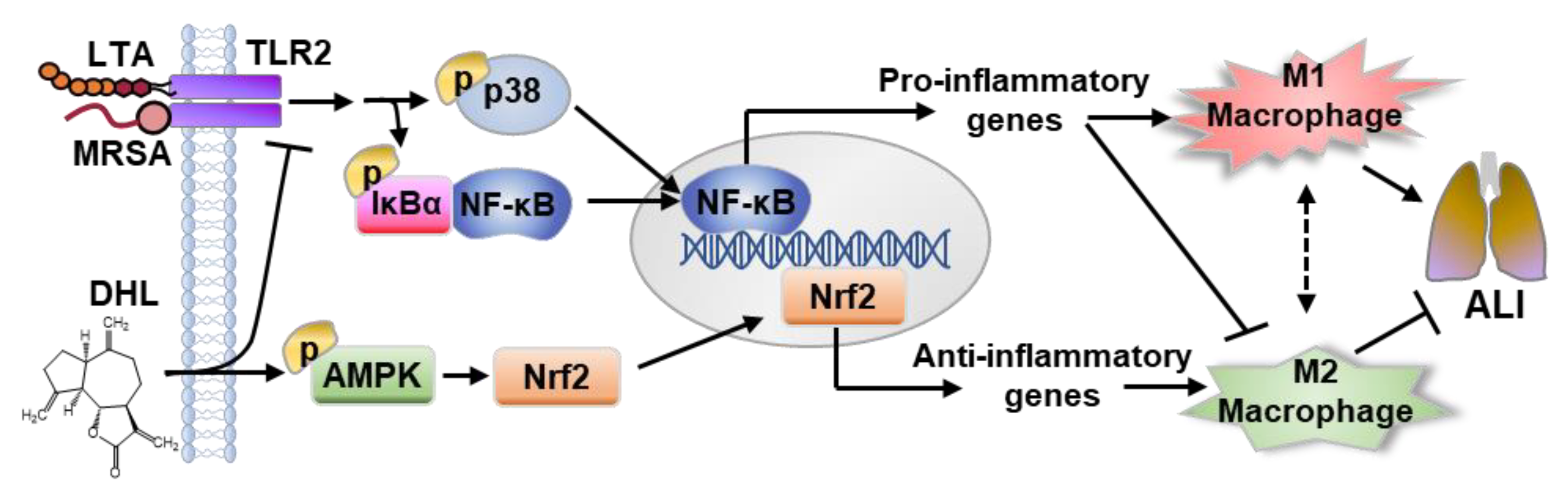
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture and Treatment
4.3. Cell Viability Assay
4.4. Preparation of MRSA
4.5. Animals
4.6. MRSA-Induced ALI Mouse Model
4.7. Lung Histological Assay
4.8. Flow Cytometry Assay
4.9. Myeloperoxidase Activity Assay
4.10. Measurement of MDA and GSH Contents
4.11. Quantitative RT-PCR Analysis
4.12. Immunoblotting
4.13. Immunohistochemistry
4.14. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALI | acute lung injury |
| AMPK | adenosine monophosphate-activated protein kinase |
| ARDS | acute respiratory distress syndrome |
| Arg-1 | arginase-1 |
| BALF | bronchoalveolar lavage fluid |
| BMDM | bone-marrow-derived macrophage |
| GSH | glutathione |
| H&E | hematoxylin and eosin |
| HO-1 | heme oxygenase-1 |
| IκBα | inhibitor of nuclear factor-κB alpha |
| IL-1β | interleukin-1 beta |
| IL-6 | interleukin-6 |
| iNOS | inducible nitric oxide synthase |
| LTA | lipoteichoic acid |
| M1 | classically activated macrophage |
| M2 | alternatively activated macrophage |
| MAPK | mitogen-activated protein kinase |
| MDA | malondialdehyde |
| MPO | myeloperoxidase |
| MRSA | methicillin-resistant Staphylococcus aureus |
| NF-κB | nuclear factor-κB |
| Nrf2 | nuclear factor erythroid 2-releated factor 2 |
| tGPX4 | total glutathione peroxidase 4 |
| TLR2 | toll-like receptor 2 |
| TNF-α | tumor necrosis factor-alpha |
References
- Angus, D.C. The acute respiratory distress syndrome: What’s in a name? JAMA 2012, 307, 2542–2544. [Google Scholar] [CrossRef]
- DeLeo, F.R.; Otto, M.; Kreiswirth, B.N.; Chambers, H.F. Community-associated meticillin-resistant Staphylococcus aureus. Lancet 2010, 375, 1557–1568. [Google Scholar] [CrossRef] [Green Version]
- Klevens, R.M.; Morrison, M.A.; Nadle, J.; Petit, S.; Gershman, K.; Ray, S.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; Townes, J.M.; et al. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 2007, 298, 1763–1771. [Google Scholar] [CrossRef] [Green Version]
- Karki, P.; Ke, Y.; Tian, Y.; Ohmura, T.; Sitikov, A.; Sarich, N.; Montgomery, C.P.; Birukova, A.A. Staphylococcus aureus-induced endothelial permeability and inflammation are mediated by microtubule destabilization. J. Biol. Chem. 2019, 294, 3369–3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, L.K.; Rims, C.R.; Gill, S.E.; McGuire, J.K.; Manicone, A.M. Pulmonary macrophage subpopulations in the induction and resolution of acute lung injury. Am. J. Respir. Cell Mol. Biol. 2012, 47, 417–426. [Google Scholar] [CrossRef]
- Mittal, M.; Tiruppathi, C.; Nepal, S.; Zhao, Y.Y.; Grzych, D.; Soni, D.; Prockop, D.J.; Malik, A.B. TNFalpha-stimulated gene-6 (TSG6) activates macrophage phenotype transition to prevent inflammatory lung injury. Proc. Natl. Acad. Sci. USA 2016, 113, E8151–E8158. [Google Scholar] [CrossRef] [Green Version]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef]
- Di Lorenzo, A.; Bolli, E.; Tarone, L.; Cavallo, F.; Conti, L. Toll-Like Receptor 2 at the Crossroad between Cancer Cells, the Immune System, and the Microbiota. Int J Mol Sci 2020, 21, 9418. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Jiang, K.; Wu, H.; Qiu, C.; Deng, G.; Peng, X. Polydatin reduces Staphylococcus aureus lipoteichoic acid-induced injury by attenuating reactive oxygen species generation and TLR2-NFkappaB signalling. J. Cell. Mol. Med. 2017, 21, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Qian, F.; Deng, J.; Gantner, B.N.; Flavell, R.A.; Dong, C.; Christman, J.W.; Ye, R.D. Map kinase phosphatase 5 protects against sepsis-induced acute lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, L866-74. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zmijewski, J.W.; Lorne, E.; Liu, G.; Park, Y.J.; Tsuruta, Y.; Abraham, E. Activation of AMPK attenuates neutrophil proinflammatory activity and decreases the severity of acute lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 295, L497–L504. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, Y.; Xu, Y.; Ruan, W.; Wang, H.; Zhang, Y.; Saavedra, J.M.; Zhang, L.; Huang, Z.; Pang, T. A Dual AMPK/Nrf2 Activator Reduces Brain Inflammation After Stroke by Enhancing Microglia M2 Polarization. Antioxid. Redox Signal. 2018, 28, 141–163. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Xiao, J.; Zhai, H.; Hao, J. Dimethyl fumarate attenuates experimental autoimmune neuritis through the nuclear factor erythroid-derived 2-related factor 2/hemoxygenase-1 pathway by altering the balance of M1/M2 macrophages. J. Neuroinflammation 2016, 13, 97. [Google Scholar] [CrossRef] [Green Version]
- Porcheray, F.; Viaud, S.; Rimaniol, A.C.; Leone, C.; Samah, B.; Dereuddre-Bosquet, N.; Dormont, D.; Gras, G. Macrophage activation switching: An asset for the resolution of inflammation. Clin. Exp. Immunol. 2005, 142, 481–489. [Google Scholar] [CrossRef]
- Heerspink, H.J.L.; Perco, P.; Mulder, S.; Leierer, J.; Hansen, M.K.; Heinzel, A.; Mayer, G. Canagliflozin reduces inflammation and fibrosis biomarkers: A potential mechanism of action for beneficial effects of SGLT2 inhibitors in diabetic kidney disease. Diabetologia 2019, 62, 1154–1166. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Wu, F.; Shi, Z.; He, B.; Zhao, X.; Wu, H.; Yan, S. Dehydrocostus lactone attenuates osteoclastogenesis and osteoclast-induced bone loss by modulating NF-kappaB signalling pathway. J. Cell. Mol. Med. 2019, 23, 5762–5770. [Google Scholar] [CrossRef]
- Nie, Y.; Wang, Z.; Chai, G.; Xiong, Y.; Li, B.; Zhang, H.; Xin, R.; Qian, X.; Tang, Z.; Wu, J.; et al. Dehydrocostus Lactone Suppresses LPS-induced Acute Lung Injury and Macrophage Activation through NF-kappaB Signaling Pathway Mediated by p38 MAPK and Akt. Molecules 2019, 24, 1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Cui, X.; Zhou, Y.; Chai, G.; Jiang, X.; Ge, G.; Wang, Y.; Sun, H.; Che, H.; Nie, Y.; et al. Dehydrocostus lactone inhibits BLM-induced pulmonary fibrosis and inflammation in mice via the JNK and p38 MAPK-mediated NF-kappaB signaling pathways. Int. Immunopharmacol. 2021, 98, 107780. [Google Scholar] [CrossRef]
- Li, W.; Ma, Y.B.; Mao, Y.Q.; Lin, T. Dehydrocostus lactone suppresses cell growth and induces apoptosis in recombinant human papilloma virus18 HaCaT cells via the PI3K/Akt signaling pathway. Mol. Med. Rep. 2018, 17, 7925–7930. [Google Scholar]
- Lee, H.I.; Lee, J.; Hwang, D.; Lee, G.R.; Kim, N.; Kwon, M.; Lee, H.; Piao, D.; Kim, H.J.; Kim, N.Y.; et al. Dehydrocostus lactone suppresses osteoclast differentiation by regulating NFATc1 and inhibits osteoclast activation through modulating migration and lysosome function. FASEB J. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 9685–9694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Yuan, G.; Lin, X.; Liu, Q.; Xu, J.; Lian, Z.; Song, F.; Zheng, J.; Xie, D.; Chen, L.; et al. Dehydrocostus lactone (DHC) suppresses estrogen deficiency-induced osteoporosis. Biochem. Pharmacol. 2019, 163, 279–289. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, J.; Li, Y.; Han, X.; Gao, K.; Fang, J. Bioassay-guided isolation of dehydrocostus lactone from Saussurea lappa: A new targeted cytosolic thioredoxin reductase anticancer agent. Arch. Biochem. Biophys. 2016, 607, 20–26. [Google Scholar] [CrossRef]
- Chen, Y.; Li, R.; Wang, Z.; Hou, X.; Wang, C.; Ai, Y.; Shi, W.; Zhan, X.; Wang, J.B.; Xiao, X.; et al. Dehydrocostus lactone inhibits NLRP3 inflammasome activation by blocking ASC oligomerization and prevents LPS-mediated inflammation in vivo. Cell. Immunol. 2020, 349, 104046. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Lee, H.J.; Ryu, J.H.; Chung, K.S. Inhibition of LPS-induced NO production and NF-kappaB activation by a sesquiterpene from Saussurea lappa. Arch. Pharmacal Res. 2000, 23, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, I. Role of lipoteichoic acid in infection and inflammation. Lancet. Infect. Dis. 2002, 2, 171–179. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, D.; Li, M.; Wang, B. Costunolide ameliorates lipoteichoic acid-induced acute lung injury via attenuating MAPK signaling pathway. Int. Immunopharmacol. 2018, 61, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; He, H.; Ding, Y.; Liu, S.; Zhang, D.; Wang, J.; Jiang, H.; Zhang, D.; Sun, L.; Ye, R.D.; et al. MK2 mediates macrophage activation and acute lung injury by regulating let-7e miRNA. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L371–L381. [Google Scholar] [CrossRef]
- Song, Y.; Wu, Y.; Li, X.; Shen, Y.; Ding, Y.; Zhu, H.; Liu, F.; Yu, K.; Sun, L.; Qian, F. Protostemonine attenuates alternatively activated macrophage and DRA-induced asthmatic inflammation. Biochem. Pharmacol. 2018, 155, 198–206. [Google Scholar] [CrossRef]
- Chawla, A.; Nguyen, K.D.; Goh, Y.P. Macrophage-mediated inflammation in metabolic disease. Nat. Rev. Immunol. 2011, 11, 738–749. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, Y.; Zhang, P.; Ruan, W.; Zhang, L.; Yuan, S.; Pang, T.; Jia, A.Q. Smiglaside A ameliorates LPS-induced acute lung injury by modulating macrophage polarization via AMPK-PPARgamma pathway. Biochem. Pharmacol. 2018, 156, 385–395. [Google Scholar] [CrossRef]
- Shabani, M.; Sadeghi, A.; Hosseini, H.; Teimouri, M.; Babaei Khorzoughi, R.; Pasalar, P.; Meshkani, R. Resveratrol alleviates obesity-induced skeletal muscle inflammation via decreasing M1 macrophage polarization and increasing the regulatory T cell population. Sci. Rep. 2020, 10, 3791. [Google Scholar] [CrossRef]
- Wang, M.; Wang, L.; Fang, L.; Li, S.; Liu, R. NLRC5 negatively regulates LTA-induced inflammation via TLR2/NF-kappaB and participates in TLR2-mediated allergic airway inflammation. J. Cell. Physiol. 2019, 234, 19990–20001. [Google Scholar] [CrossRef]
- Huang, B.R.; Tsai, C.F.; Lin, H.Y.; Tseng, W.P.; Huang, S.S.; Wu, C.R.; Lin, C.; Yeh, W.L.; Lu, D.Y. Interaction of inflammatory and anti-inflammatory responses in microglia by Staphylococcus aureus-derived lipoteichoic acid. Toxicol. Appl. Pharmacol. 2013, 269, 43–50. [Google Scholar] [CrossRef]
- Jiang, K.F.; Zhao, G.; Deng, G.Z.; Wu, H.C.; Yin, N.N.; Chen, X.Y.; Qiu, C.W.; Peng, X.L. Polydatin ameliorates Staphylococcus aureus-induced mastitis in mice via inhibiting TLR2-mediated activation of the p38 MAPK/NF-kappaB pathway. Acta Pharmacol. Sin. 2017, 38, 211–222. [Google Scholar] [CrossRef]
- Wu, Y.; Nie, Y.; Huang, J.; Qiu, Y.; Wan, B.; Liu, G.; Chen, J.; Chen, D.; Pang, Q. Protostemonine alleviates heat-killed methicillin-resistant Staphylococcus aureus-induced acute lung injury through MAPK and NF-kappaB signaling pathways. Int. Immunopharmacol. 2019, 77, 105964. [Google Scholar] [CrossRef] [PubMed]
- Ge, G.; Bai, J.; Wang, Q.; Liang, X.; Tao, H.; Chen, H.; Wei, M.; Niu, J.; Yang, H.; Xu, Y.; et al. Punicalagin ameliorates collagen-induced arthritis by downregulating M1 macrophage and pyroptosis via NF-kappaB signaling pathway. Sci. China Life Sci. 2021. [Google Scholar] [CrossRef]
- Zhong, J.; Wang, H.; Chen, W.; Sun, Z.; Chen, J.; Xu, Y.; Weng, M.; Shi, Q.; Ma, D.; Miao, C. Ubiquitylation of MFHAS1 by the ubiquitin ligase praja2 promotes M1 macrophage polarization by activating JNK and p38 pathways. Cell Death Dis. 2017, 8, e2763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frasnelli, M.E.; Tarussio, D.; Chobaz-Peclat, V.; Busso, N.; So, A. TLR2 modulates inflammation in zymosan-induced arthritis in mice. Arthritis Res. Ther. 2005, 7, R370-9. [Google Scholar] [CrossRef] [Green Version]
- Quero, L.; Hanser, E.; Manigold, T.; Tiaden, A.N.; Kyburz, D. TLR2 stimulation impairs anti-inflammatory activity of M2-like macrophages, generating a chimeric M1/M2 phenotype. Arthritis Res. Ther. 2017, 19, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Shi, J.; Zhao, L.; Guan, J.; Liu, F.; Huo, G.; Li, B. Lactobacillus plantarum KLDS1.0344 and Lactobacillus acidophilus KLDS1.0901 Mixture Prevents Chronic Alcoholic Liver Injury in Mice by Protecting the Intestinal Barrier and Regulating Gut Microbiota and Liver-Related Pathways. J. Agric. Food Chem. 2021, 69, 183–197. [Google Scholar] [CrossRef]
- Lee, H.I.; Lee, G.R.; Lee, J.; Kim, N.; Kwon, M.; Kim, H.J.; Kim, N.Y.; Park, J.H.; Jeong, W. Dehydrocostus lactone inhibits NFATc1 via regulation of IKK, JNK, and Nrf2, thereby attenuating osteoclastogenesis. BMB Rep. 2020, 53, 218–222. [Google Scholar] [CrossRef]
- Luo, D.; Guo, Y.; Cheng, Y.; Zhao, J.; Wang, Y.; Rong, J. Natural product celastrol suppressed macrophage M1 polarization against inflammation in diet-induced obese mice via regulating Nrf2/HO-1, MAP kinase and NF-kappaB pathways. Aging 2017, 9, 2069–2082. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, C.; Xie, M.; Tang, T.; Wang, Z.; Nie, X. Atorvastatin causes oxidative stress and alteration of lipid metabolism in estuarine goby Mugilogobius abei. Env. Pollut. 2021, 289, 117879. [Google Scholar] [CrossRef]
- Liu, G.; Wu, Y.; Jin, S.; Sun, J.; Wan, B.B.; Zhang, J.; Wang, Y.; Gao, Z.Q.; Chen, D.; Li, S.; et al. Itaconate ameliorates methicillin-resistant Staphylococcus aureus-induced acute lung injury through the Nrf2/ARE pathway. Ann. Transl. Med. 2021, 9, 712. [Google Scholar] [CrossRef] [PubMed]
- Ahn, T.; Kim, C.H.; Kim, T.H.; Chang, J.S.; Jeong, M.Y.; Aditya, K.; Yoon, P.W. What is the Prevalence of Radiographic Hip Findings Associated with Femoroacetabular Impingement in Asymptomatic Asian Volunteers? Clin. Orthop. Relat. Res. 2016, 474, 2655–2661. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Wang, H.; Yang, Y.; Gou, Y.; Wang, Z.; Yang, D.; Li, C. Protective Effects of Silymarin Against D-Gal/LPS-Induced Organ Damage and Inflammation in Mice. Drug Des. Devel. 2021, 15, 1903–1914. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, L.; Xing, Q.; Li, K.; Si, J.; Ma, X.; Mao, L. Sevoflurane inhibits ferroptosis: A new mechanism to explain its protective role against lipopolysaccharide-induced acute lung injury. Life Sci. 2021, 275, 119391. [Google Scholar] [CrossRef]
- Dong, H.; Qiang, Z.; Chai, D.; Peng, J.; Xia, Y.; Hu, R.; Jiang, H. Nrf2 inhibits ferroptosis and protects against acute lung injury due to intestinal ischemia reperfusion via regulating SLC7A11 and HO-1. Aging 2020, 12, 12943–12959. [Google Scholar] [CrossRef]
- Qiu, Y.B.; Wan, B.B.; Liu, G.; Wu, Y.X.; Chen, D.; Lu, M.D.; Chen, J.L.; Yu, R.Q.; Chen, D.Z.; Pang, Q.F. Nrf2 protects against seawater drowning-induced acute lung injury via inhibiting ferroptosis. Respir. Res. 2020, 21, 232. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Chen, D.; Liu, G.; Wu, Y.X.; Gao, Z.Q.; Su, Y.; Yuan, J.N.; Liu, L.; Shan, J.C.; Pang, Q.F.; et al. A protects against methicillin resistant staphylococcus aureus-induced acute lung injury via Nrf2 pathway. Int. Immunopharmacol. 2021, 90, 107221. [Google Scholar] [CrossRef] [PubMed]
- Hoogendijk, A.J.; Pinhancos, S.S.; van der Poll, T.; Wieland, C.W. AMP-activated protein kinase activation by 5-aminoimidazole-4-carbox-amide-1-beta-D-ribofuranoside (AICAR) reduces lipoteichoic acid-induced lung inflammation. J. Biol. Chem. 2013, 288, 7047–7052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.X.; Zeng, S.; Wan, B.B.; Wang, Y.Y.; Sun, H.X.; Liu, G.; Gao, Z.Q.; Chen, D.; Chen, Y.Q.; Lu, M.D.; et al. Sophoricoside attenuates lipopolysaccharide-induced acute lung injury by activating the AMPK/Nrf2 signaling axis. Int. Immunopharmacol. 2021, 90, 107187. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.M.; Lv, J.; Zeng, Q.L.; Shen, S.; Xing, J.Y.; Zhang, Y.Y.; Zhang, Z.H.; Yu, Z.J. AMPK activation ameliorates D-GalN/LPS-induced acute liver failure by upregulating Foxo3A to induce autophagy. Exp. Cell Res. 2017, 358, 335–342. [Google Scholar] [CrossRef]
- Grommes, J.; Soehnlein, O. Contribution of neutrophils to acute lung injury. Mol. Med. 2011, 17, 293–307. [Google Scholar] [CrossRef]
- Zhu, B.; Zhang, R.; Li, C.; Jiang, L.; Xiang, M.; Ye, Z.; Kita, H.; Melnick, A.M.; Dent, A.L.; Sun, J. BCL6 modulates tissue neutrophil survival and exacerbates pulmonary inflammation following influenza virus infection. Proc. Natl. Acad. Sci. USA 2019, 116, 11888–11893. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Onizawa, S.; Nagai, A.; Aoshiba, K. Epithelial cell senescence impairs repair process and exacerbates inflammation after airway injury. Respir. Res. 2011, 12, 78. [Google Scholar] [CrossRef] [Green Version]
- Tsukita, K.; Yano, T.; Tamura, A.; Tsukita, S. Reciprocal Association between the Apical Junctional Complex and AMPK: A Promising Therapeutic Target for Epithelial/Endothelial Barrier Function? Int. J. Mol. Sci. 2019, 20, 6012. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Xiao, C.X.; Su, Z.H.; Huang, M.W.; Qin, M.; Wu, W.J.; Jia, W.W.; Zhu, Y.Z.; Hu, J.F.; Liu, X.H. (-)-7(S)-hydroxymatairesinol protects against tumor necrosis factor-alpha-mediated inflammation response in endothelial cells by blocking the MAPK/NF-kappaB and activating Nrf2/HO-1. Phytomedicine Int. J. Phytother. Phytopharm. 2017, 32, 15–23. [Google Scholar]
- Zhu, Y.; Wang, C.; Luo, J.; Hua, S.; Li, D.; Peng, L.; Liu, H.; Song, L. The protective role of Zingerone in a murine asthma model via activation of the AMPK/Nrf2/HO-1 pathway. Food Funct. 2021, 12, 3120–3131. [Google Scholar] [CrossRef] [PubMed]
- Roussel, L.; Houle, F.; Chan, C.; Yao, Y.; Berube, J.; Olivenstein, R.; Martin, J.G.; Huot, J.; Hamid, Q.; Ferri, L.; et al. IL-17 promotes p38 MAPK-dependent endothelial activation enhancing neutrophil recruitment to sites of inflammation. J. Immunol. 2010, 184, 4531–4537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V. Pulmonary Innate Immune Response Determines the Outcome of Inflammation During Pneumonia and Sepsis-Associated Acute Lung Injury. Front. Immunol. 2020, 11, 1722. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Su, Y.; Wu, K.; Pan, F.; Wang, A. DOCK2 contributes to endotoxemia-induced acute lung injury in mice by activating proinflammatory macrophages. Biochem. Pharmacol. 2021, 184, 114399. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.M.; Peng, C.K.; Yang, S.S.; Shui, H.A.; Huang, K.L. WNK4-SPAK modulates lipopolysaccharide-induced macrophage activation. Biochem. Pharmacol. 2020, 171, 113738. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer | Primer Sequences (5′–3′) |
|---|---|---|
| GAPDH | Forward | TGGCCTTCCGTGTTCCTAC |
| Reverse | GAGTTGCTGTTGAAGTCGCA | |
| TNF-α | Forward | CCTGTAGCCCACGTCGTAG |
| Reverse | GGGAGTAGACAAGGTACAACCC | |
| IL-1β | Forward | GAAATGCCACCTT TTGACAGTG |
| Reverse | TGGATGCTCTCATCAG GACAG | |
| IL-6 | Forward | CTGCAAGAGACTTCCATCCAG |
| Reverse | AGTGGTATAGACAGGTCTG TTGG | |
| tGPX4 | Forward | CGCAGCCGTTCTTATCAATG |
| Reverse | CACTGTGGAAATGGATGAAAGTC | |
| iNOS | Forward | GTTCTCAGCCCAACAATACAAGA |
| Reverse | GTGGACGGGTCGATGTCAC | |
| CD163 | Forward | ATGGGTGGACACAGAATGGTT |
| Reverse | CAGGAGCGTTAGTGACAGCAG | |
| CD206 | Forward | CTCTGTTCAGCTATTGGACGC |
| Reverse | TGGCACTCCCAAACATAATTTGA | |
| Arg-1 | Forward | CTCCAAGCCAAAGTCCTTAGAG |
| Reverse | GGAGCTGTCATTAGGGACATCA | |
| TLR2 | Forward | TCTAAAGTCGATCCGCGACAT |
| Reverse | CTACGGGCAGTGGTGAAAACT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.-X.; Jiang, F.-J.; Liu, G.; Wang, Y.-Y.; Gao, Z.-Q.; Jin, S.-H.; Nie, Y.-J.; Chen, D.; Chen, J.-L.; Pang, Q.-F. Dehydrocostus Lactone Attenuates Methicillin-Resistant Staphylococcus aureus-Induced Inflammation and Acute Lung Injury via Modulating Macrophage Polarization. Int. J. Mol. Sci. 2021, 22, 9754. https://doi.org/10.3390/ijms22189754
Wu Y-X, Jiang F-J, Liu G, Wang Y-Y, Gao Z-Q, Jin S-H, Nie Y-J, Chen D, Chen J-L, Pang Q-F. Dehydrocostus Lactone Attenuates Methicillin-Resistant Staphylococcus aureus-Induced Inflammation and Acute Lung Injury via Modulating Macrophage Polarization. International Journal of Molecular Sciences. 2021; 22(18):9754. https://doi.org/10.3390/ijms22189754
Chicago/Turabian StyleWu, Ya-Xian, Feng-Juan Jiang, Gang Liu, Ying-Ying Wang, Zhi-Qi Gao, Si-Hao Jin, Yun-Juan Nie, Dan Chen, Jun-Liang Chen, and Qing-Feng Pang. 2021. "Dehydrocostus Lactone Attenuates Methicillin-Resistant Staphylococcus aureus-Induced Inflammation and Acute Lung Injury via Modulating Macrophage Polarization" International Journal of Molecular Sciences 22, no. 18: 9754. https://doi.org/10.3390/ijms22189754
APA StyleWu, Y. -X., Jiang, F. -J., Liu, G., Wang, Y. -Y., Gao, Z. -Q., Jin, S. -H., Nie, Y. -J., Chen, D., Chen, J. -L., & Pang, Q. -F. (2021). Dehydrocostus Lactone Attenuates Methicillin-Resistant Staphylococcus aureus-Induced Inflammation and Acute Lung Injury via Modulating Macrophage Polarization. International Journal of Molecular Sciences, 22(18), 9754. https://doi.org/10.3390/ijms22189754