
The Cdc14 Phosphatase Controls Resolution of Recombination Intermediates and Crossover Formation during Meiosis

 , , , , , and
, , , , , and 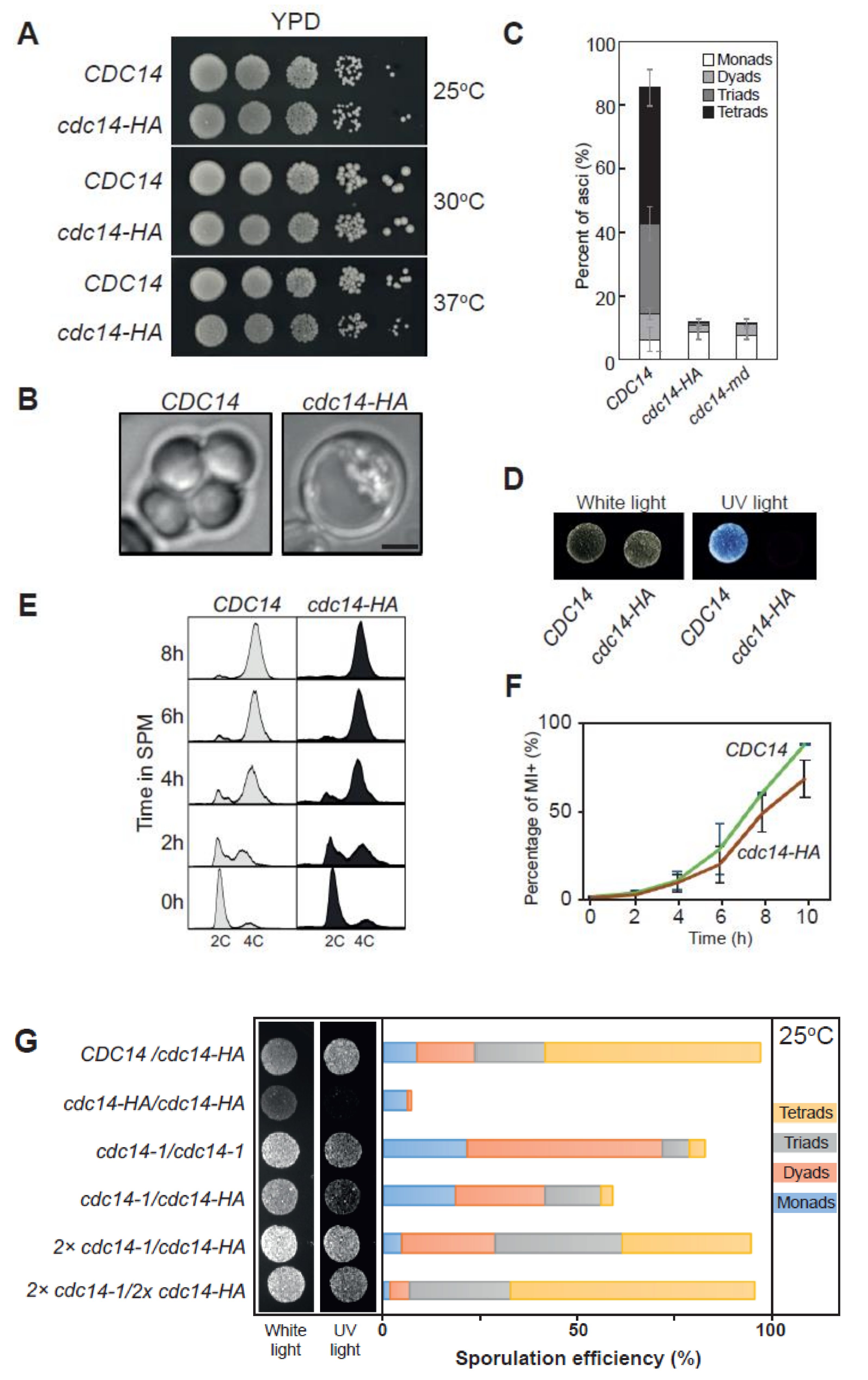{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of a Separation-Of-Function Allele of CDC14 Defective in Sporulation
2.2. Distinct Sporulation-Defective Alleles of CDC14 Block Meiotic Progression at Different Stages
2.3. Cdc14 Protein Levels Are Depleted during Meiosis in cdc14-HA Cells
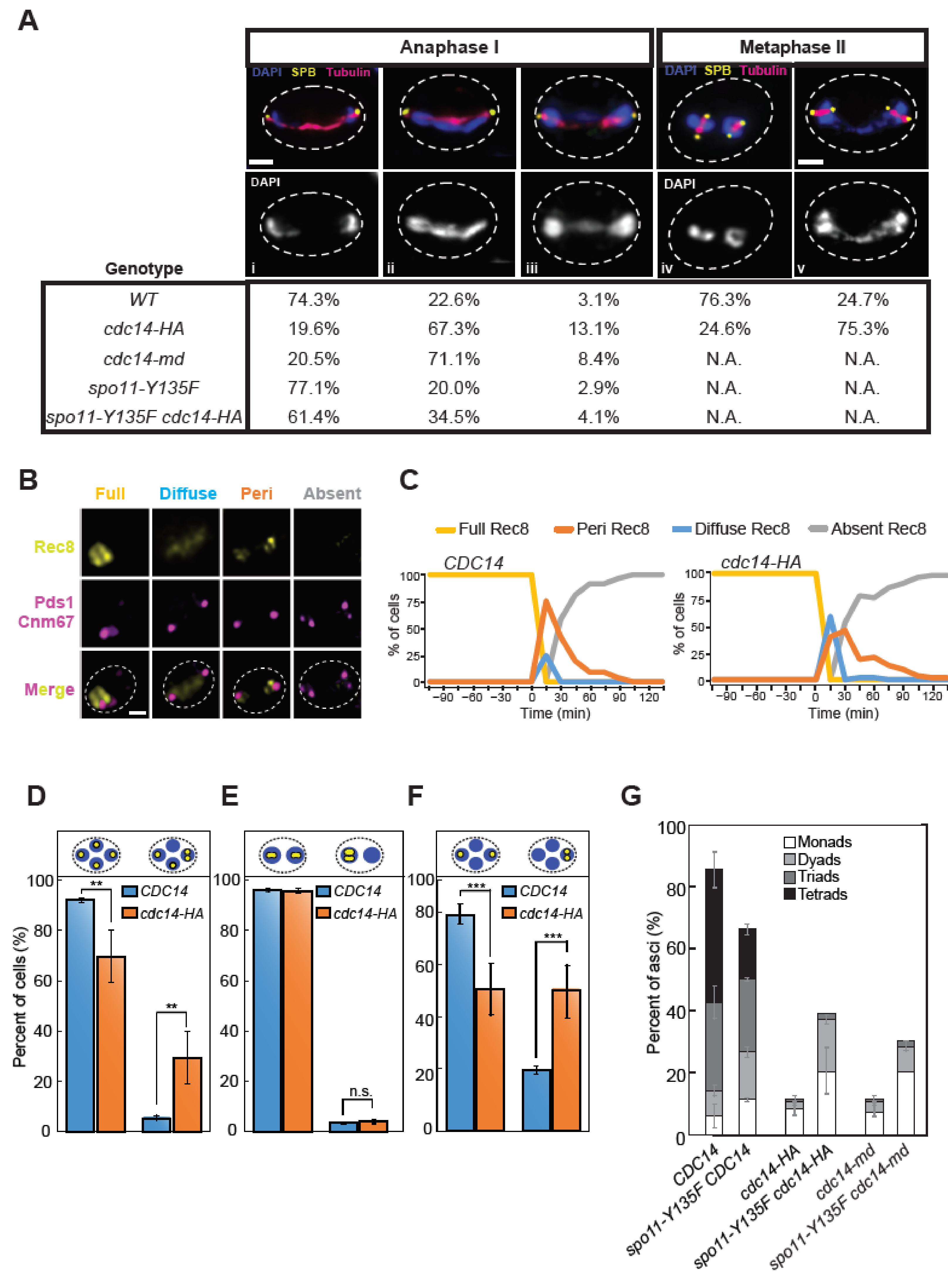
2.4. SPB Integrity Is Compromised in cdc14-HA Cells after the Second Meiotic Division
2.5. Impeded Homolog Separation and Sister Missegregation Are Common Hallmarks of cdc14 Mutations in Meiosis
2.6. Preventing Recombination Alleviates Defective Events in Meiosis-Specific cdc14 Mutants
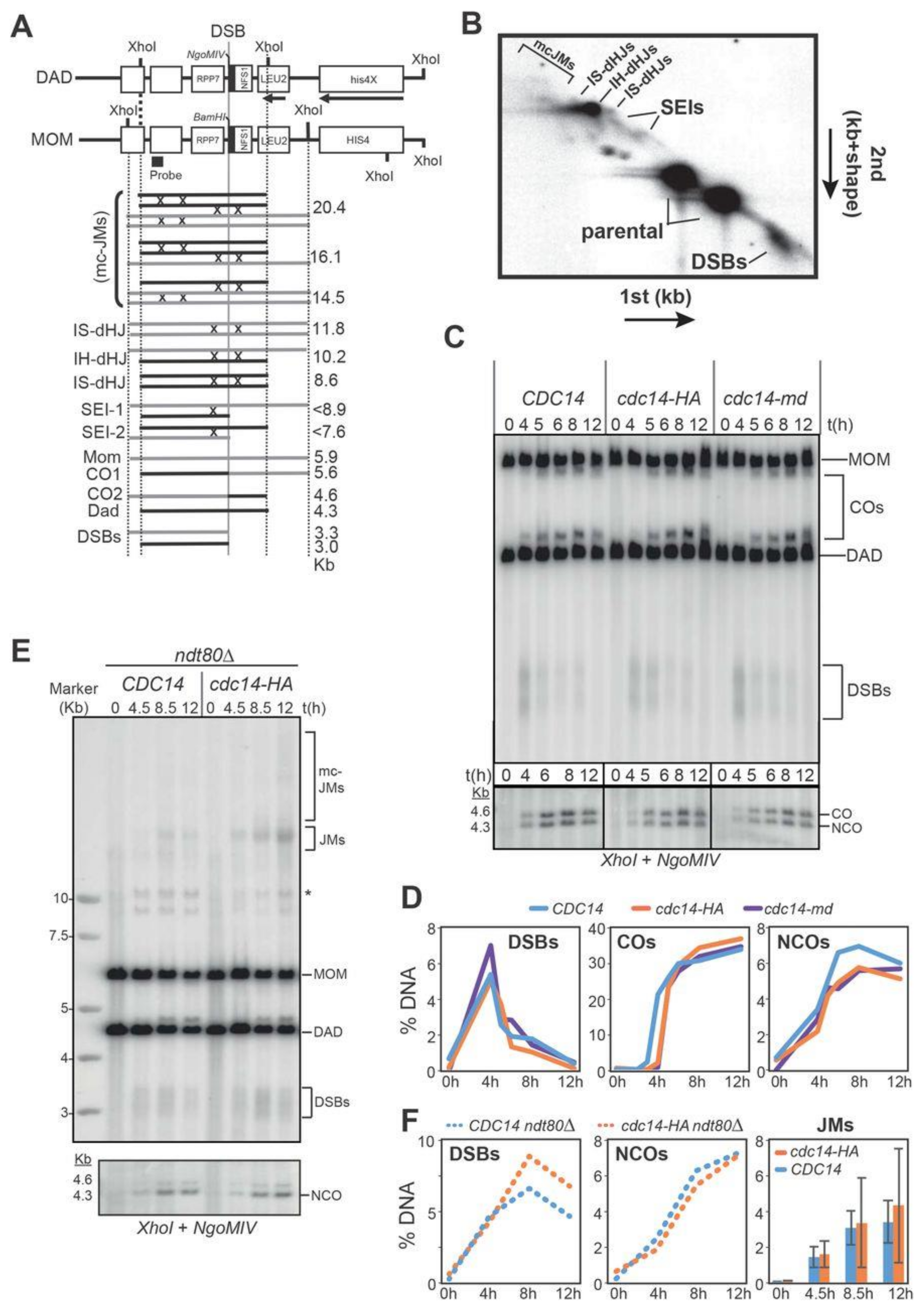
2.7. Meiotic Recombination Is Impaired in cdc14 Mutants
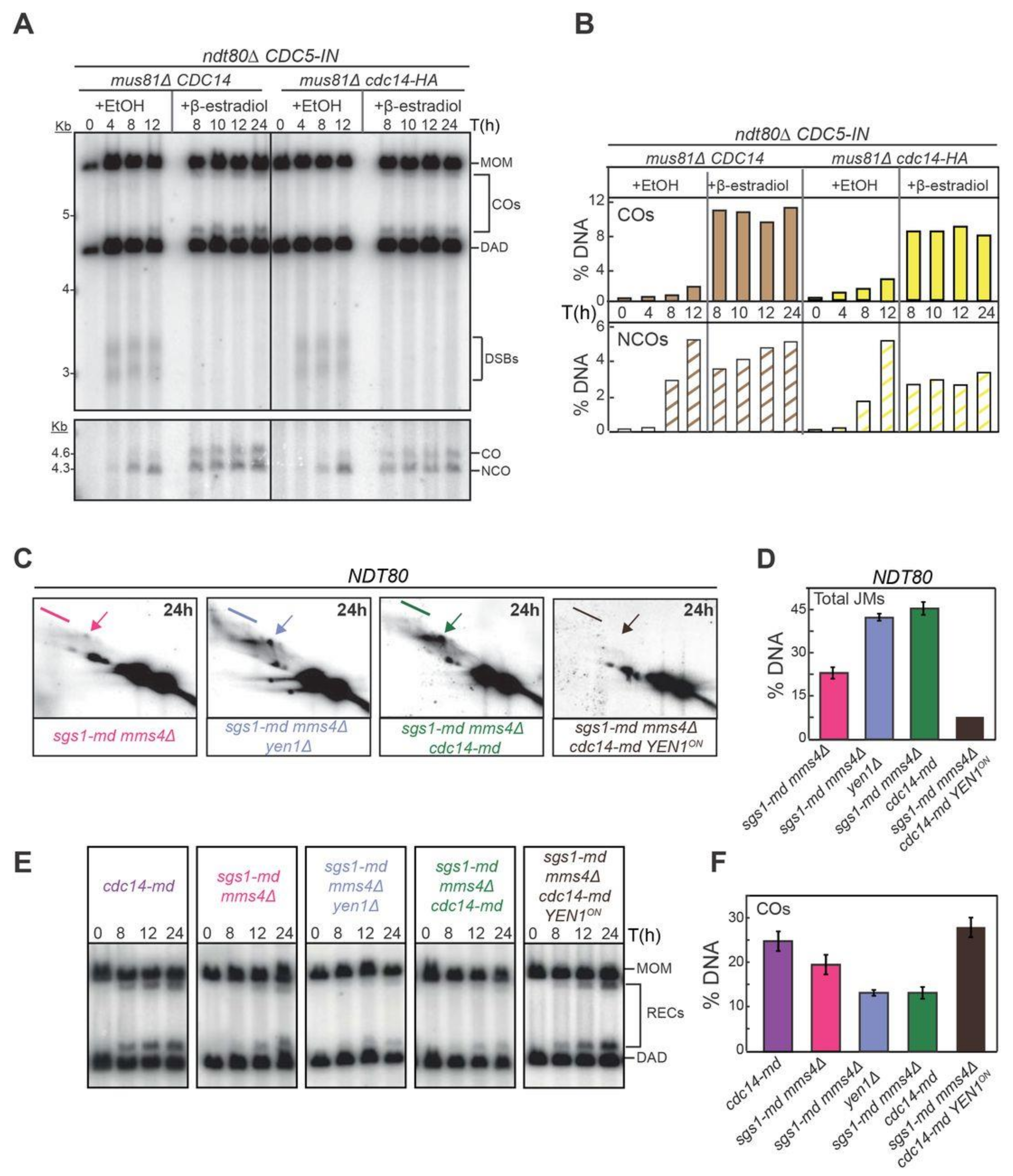
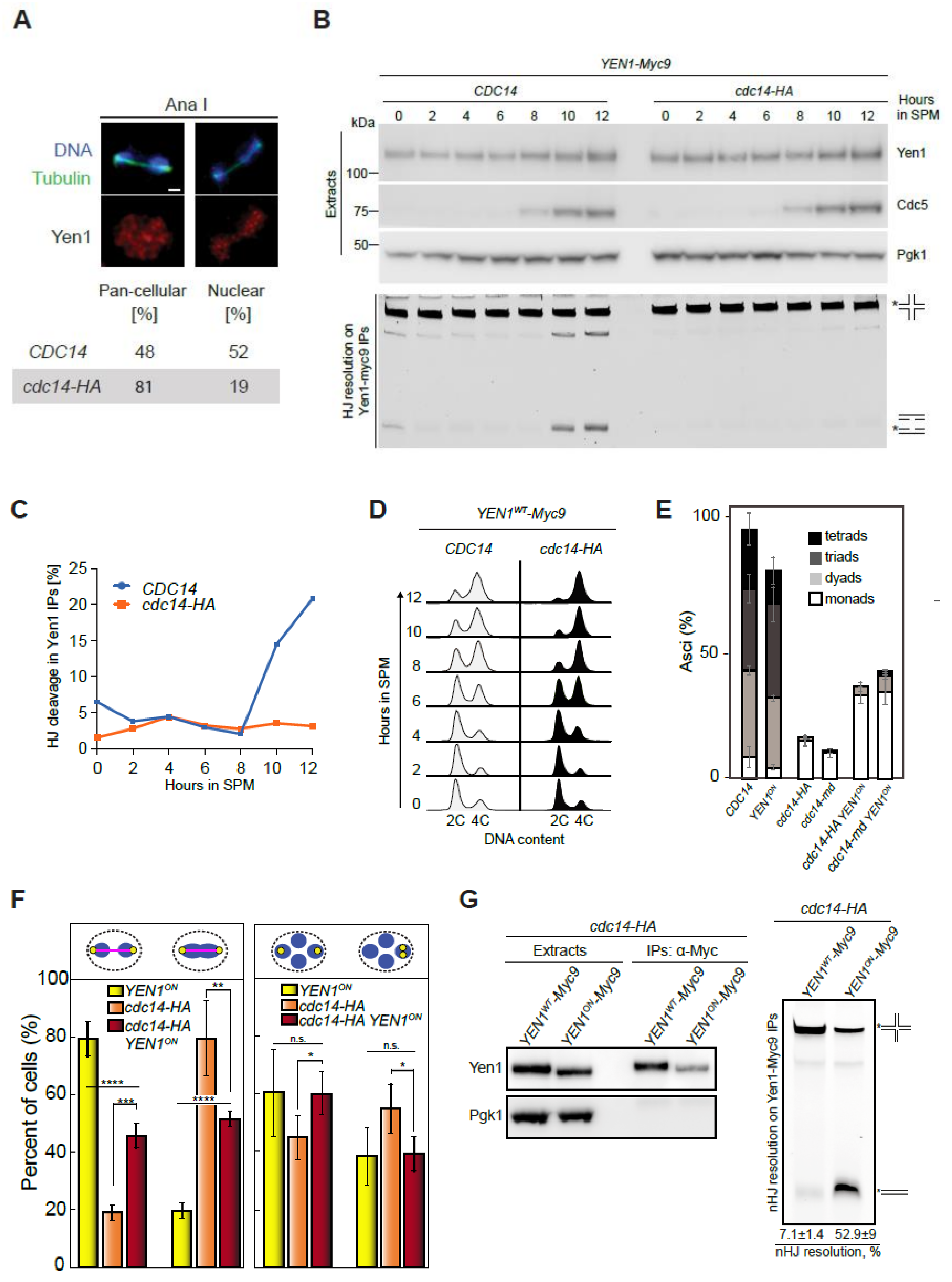
2.8. Yen1 Nuclear Localization and Activity during Meiosis Requires Cdc14 Function
2.9. Constitutively Active Yen1 Suppresses the Accumulation of Aberrant Recombination Intermediates in cdc14 Mutants
2.10. Cdc14 and Yen1 Promote JM Resolution during the First Meiotic Division
3. Discussion
3.1. Novel Insights into Cdc14 Meiotic Functions Using Different cdc14 Alleles
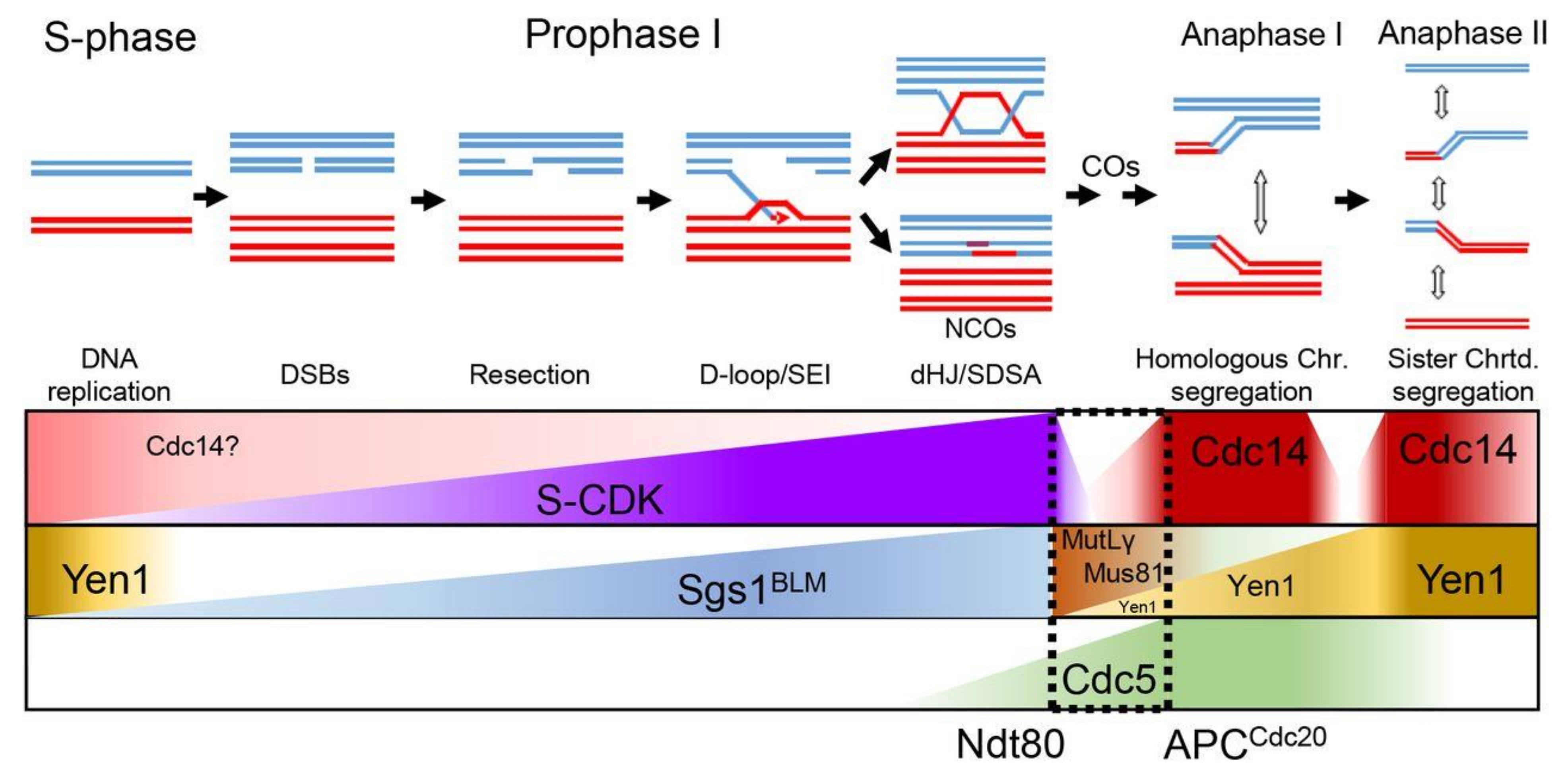
3.2. Multistage CDC14-Dependent Processing of Recombination Intermediates
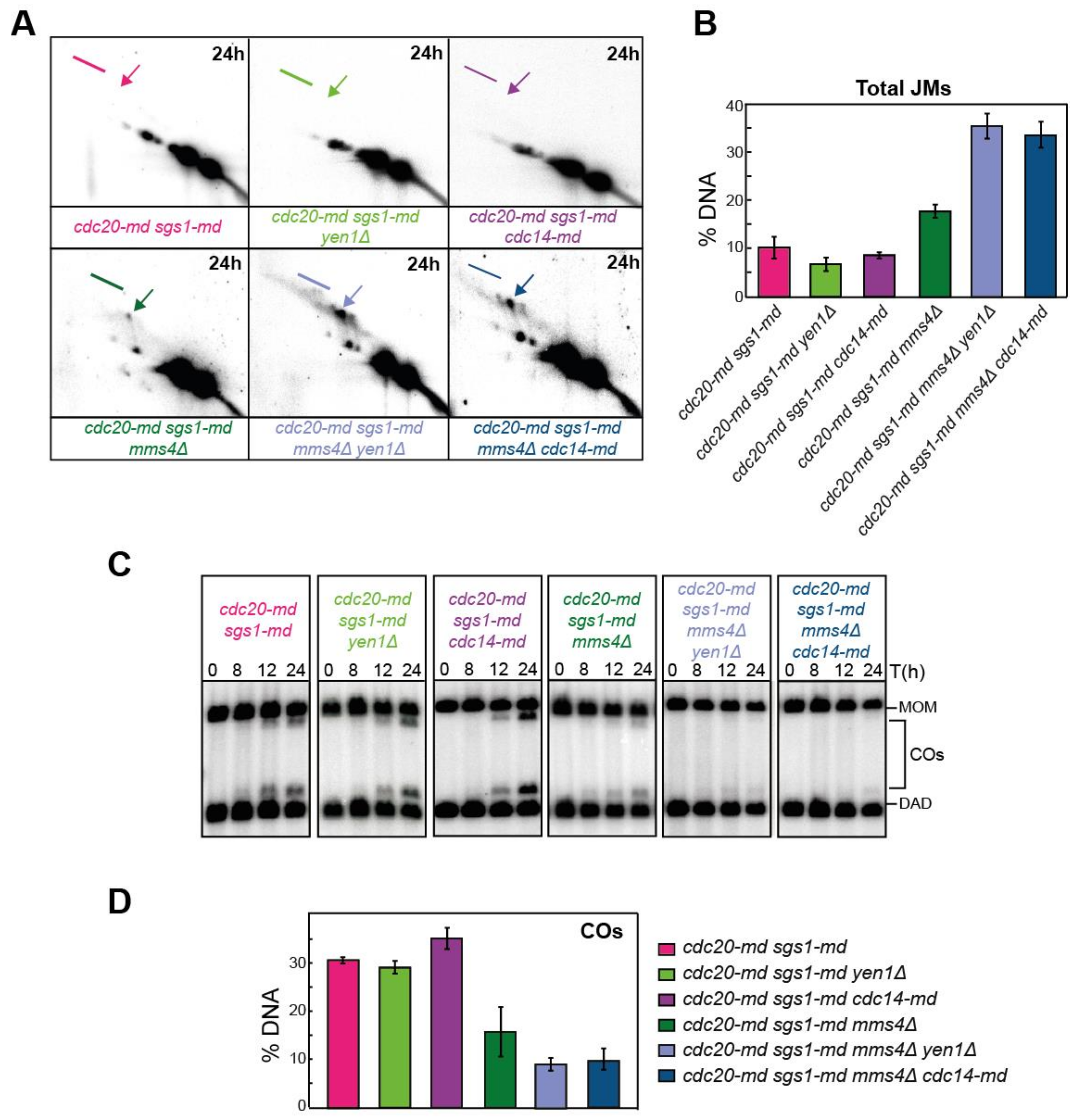
3.3. Alternative Repair Pathways to Process JMs during Meiosis I
4. Materials and Methods
4.1. Yeast Strains and Plasmids
4.2. Synchronous Meiotic Time Courses
4.3. DNA Manipulation, Extraction and Southern Blot Detection
4.4. Time-Lapse Imaging, Immunofluorescence, Microscopy, and Image Analysis
4.5. Protein Extraction, Western Blot Analysis and Antibodies
4.6. Nuclease Assays
4.7. Data Analysis and Biostatistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keeney, S.; Giroux, C.N.; Kleckner, N. Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell 1997, 88, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Bishop, D.K. Multiple mechanisms of meiotic recombination. Cell 2006, 127, 1095–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakharyevich, K.; Ma, Y.; Tang, S.; Hwang, P.Y.; Boiteux, S.; Hunter, N. Temporally and biochemically distinct activities of Exo1 during meiosis: Double-strand break resection and resolution of double Holliday junctions. Mol. Cell 2010, 40, 1001–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakharyevich, K.; Tang, S.; Ma, Y.; Hunter, N. Delineation of joint molecule resolution pathways in meiosis identifies a crossover-specific resolvase. Cell 2012, 149, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, A.; Adam, C.; Rauh, F.; Duroc, Y.; Ranjha, L.; Lombard, B.; Mu, X.; Wintrebert, M.; Loew, D.; Guarne, A.; et al. Exo1 recruits Cdc5 polo kinase to MutLgamma to ensure efficient meiotic crossover formation. Proc. Natl. Acad. Sci. USA 2020, 117, 30577–30588. [Google Scholar] [CrossRef]
- De los Santos, T.; Loidl, J.; Larkin, B.; Hollingsworth, N.M. A role for MMS4 in the processing of recombination intermediates during meiosis in Saccharomyces cerevisiae. Genetics 2001, 159, 1511–1525. [Google Scholar] [CrossRef]
- Oh, S.D.; Lao, J.P.; Taylor, A.F.; Smith, G.R.; Hunter, N. RecQ helicase, Sgs1, and XPF family endonuclease, Mus81-Mms4, resolve aberrant joint molecules during meiotic recombination. Mol. Cell 2008, 31, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Jessop, L.; Lichten, M. Mus81/Mms4 endonuclease and Sgs1 helicase collaborate to ensure proper recombination intermediate metabolism during meiosis. Mol. Cell 2008, 31, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Matos, J.; Blanco, M.G.; Maslen, S.; Skehel, J.M.; West, S.C. Regulatory control of the resolution of DNA recombination intermediates during meiosis and mitosis. Cell 2011, 147, 158–172. [Google Scholar] [CrossRef] [Green Version]
- Arter, M.; Hurtado-Nieves, V.; Oke, A.; Zhuge, T.; Wettstein, R.; Fung, J.C.; Blanco, M.G.; Matos, J. Regulated Crossing-Over Requires Inactivation of Yen1/GEN1 Resolvase during Meiotic Prophase I. Dev. Cell 2018, 45, 785–800. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.D.; Lao, J.P.; Hwang, P.Y.; Taylor, A.F.; Smith, G.R.; Hunter, N. BLM ortholog, Sgs1, prevents aberrant crossing-over by suppressing formation of multichromatid joint molecules. Cell 2007, 130, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Piazza, A.; Wright, W.D.; Heyer, W.D. Multi-invasions Are Recombination Byproducts that Induce Chromosomal Rearrangements. Cell 2017, 170, 760–773. [Google Scholar] [CrossRef] [PubMed]
- De Muyt, A.; Jessop, L.; Kolar, E.; Sourirajan, A.; Chen, J.; Dayani, Y.; Lichten, M. BLM helicase ortholog Sgs1 is a central regulator of meiotic recombination intermediate metabolism. Mol. Cell 2012, 46, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Xaver, M.; Huang, L.; Chen, D.; Klein, F. Smc5/6-Mms21 prevents and eliminates inappropriate recombination intermediates in meiosis. PLoS Genet. 2013, 9, e1004067. [Google Scholar] [CrossRef] [Green Version]
- Copsey, A.; Tang, S.; Jordan, P.W.; Blitzblau, H.G.; Newcombe, S.; Chan, A.C.; Newnham, L.; Li, Z.; Gray, S.; Herbert, A.D.; et al. Smc5/6 coordinates formation and resolution of joint molecules with chromosome morphology to ensure meiotic divisions. PLoS Genet. 2013, 9, e1004071. [Google Scholar] [CrossRef] [Green Version]
- Grigaitis, R.; Susperregui, A.; Wild, P.; Matos, J. Characterization of DNA helicases and nucleases from meiotic extracts of S. cerevisiae. Methods Cell Biol. 2018, 144, 371–388. [Google Scholar]
- San-Segundo, P.A.; Clemente-Blanco, A. Resolvases, Dissolvases, and Helicases in Homologous Recombination: Clearing the Road for Chromosome Segregation. Genes 2020, 11, 71. [Google Scholar] [CrossRef] [Green Version]
- Machin, F. Implications of Metastable Nicks and Nicked Holliday Junctions in Processing Joint Molecules in Mitosis and Meiosis. Genes 2020, 11, 1498. [Google Scholar] [CrossRef]
- Clyne, R.K.; Katis, V.L.; Jessop, L.; Benjamin, K.R.; Herskowitz, I.; Lichten, M.; Nasmyth, K. Polo-like kinase Cdc5 promotes chiasmata formation and cosegregation of sister centromeres at meiosis I. Nat. Cell Biol. 2003, 5, 480–485. [Google Scholar] [CrossRef]
- Sourirajan, A.; Lichten, M. Polo-like kinase Cdc5 drives exit from pachytene during budding yeast meiosis. Genes Dev. 2008, 22, 2627–2632. [Google Scholar] [CrossRef] [Green Version]
- Wild, P.; Susperregui, A.; Piazza, I.; Dorig, C.; Oke, A.; Arter, M.; Yamaguchi, M.; Hilditch, A.T.; Vuina, K.; Chan, K.C.; et al. Network Rewiring of Homologous Recombination Enzymes during Mitotic Proliferation and Meiosis. Mol. Cell 2019, 75, 859–874. [Google Scholar] [CrossRef] [Green Version]
- Ip, S.C.; Rass, U.; Blanco, M.G.; Flynn, H.R.; Skehel, J.M.; West, S.C. Identification of Holliday junction resolvases from humans and yeast. Nature 2008, 456, 357–361. [Google Scholar] [CrossRef]
- Eissler, C.L.; Mazon, G.; Powers, B.L.; Savinov, S.N.; Symington, L.S.; Hall, M.C. The Cdk/cDc14 module controls activation of the Yen1 holliday junction resolvase to promote genome stability. Mol. Cell 2014, 54, 80–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, M.G.; Matos, J.; West, S.C. Dual control of Yen1 nuclease activity and cellular localization by Cdk and Cdc14 prevents genome instability. Mol. Cell 2014, 54, 94–106. [Google Scholar] [CrossRef] [Green Version]
- Grigaitis, R.; Ranjha, L.; Wild, P.; Kasaciunaite, K.; Ceppi, I.; Kissling, V.; Henggeler, A.; Susperregui, A.; Peter, M.; Seidel, R.; et al. Phosphorylation of the RecQ Helicase Sgs1/BLM Controls Its DNA Unwinding Activity during Meiosis and Mitosis. Dev. Cell 2020, 53, 706–723. [Google Scholar] [CrossRef] [PubMed]
- Visintin, R.; Craig, K.; Hwang, E.S.; Prinz, S.; Tyers, M.; Amon, A. The phosphatase Cdc14 triggers mitotic exit by reversal of Cdk-dependent phosphorylation. Mol. Cell 1998, 2, 709–718. [Google Scholar] [CrossRef]
- Culotti, J.; Hartwell, L.H. Genetic control of the cell division cycle in yeast. 3. Seven genes controlling nuclear division. Exp. Cell Res. 1971, 67, 389–401. [Google Scholar] [CrossRef]
- Hartwell, L.H.; Mortimer, R.K.; Culotti, J.; Culotti, M. Genetic Control of the Cell Division Cycle in Yeast: V. Genetic Analysis of cdc Mutants. Genetics 1973, 74, 267–286. [Google Scholar] [CrossRef]
- Hartwell, L.H.; Smith, D. Altered fidelity of mitotic chromosome transmission in cell cycle mutants of S. cerevisiae. Genetics 1985, 110, 381–395. [Google Scholar] [CrossRef]
- D’Amours, D.; Stegmeier, F.; Amon, A. Cdc14 and condensin control the dissolution of cohesin-independent chromosome linkages at repeated DNA. Cell 2004, 117, 455–469. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, M.; Higuchi, T.; Katis, V.L.; Uhlmann, F. Cdc14 phosphatase induces rDNA condensation and resolves cohesin-independent cohesion during budding yeast anaphase. Cell 2004, 117, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Torres-Rosell, J.; Machin, F.; Jarmuz, A.; Aragon, L. Nucleolar segregation lags behind the rest of the genome and requires Cdc14p activation by the FEAR network. Cell Cycle 2004, 3, 496–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.D.; Yong-Gonzalez, V.; Strunnikov, A.V. Cdc14p/FEAR pathway controls segregation of nucleolus in S. cerevisiae by facilitating condensin targeting to rDNA chromatin in anaphase. Cell Cycle 2004, 3, 960–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machin, F.; Torres-Rosell, J.; De Piccoli, G.; Carballo, J.A.; Cha, R.S.; Jarmuz, A.; Aragon, L. Transcription of ribosomal genes can cause nondisjunction. J. Cell Biol. 2006, 173, 893–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente-Blanco, A.; Mayan-Santos, M.; Schneider, D.A.; Machin, F.; Jarmuz, A.; Tschochner, H.; Aragon, L. Cdc14 inhibits transcription by RNA polymerase I during anaphase. Nature 2009, 458, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Blanco, A.; Sen, N.; Mayan-Santos, M.; Sacristan, M.P.; Graham, B.; Jarmuz, A.; Giess, A.; Webb, E.; Game, L.; Eick, D.; et al. Cdc14 phosphatase promotes segregation of telomeres through repression of RNA polymerase II transcription. Nat. Cell Biol. 2011, 13, 1450–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shou, W.; Seol, J.H.; Shevchenko, A.; Baskerville, C.; Moazed, D.; Chen, Z.W.; Jang, J.; Shevchenko, A.; Charbonneau, H.; Deshaies, R.J. Exit from mitosis is triggered by Tem1-dependent release of the protein phosphatase Cdc14 from nucleolar RENT complex. Cell 1999, 97, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Visintin, R.; Hwang, E.S.; Amon, A. Cfi1 prevents premature exit from mitosis by anchoring Cdc14 phosphatase in the nucleolus. Nature 1999, 398, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Toh-e, A. Budding yeast Cdc5 phosphorylates Net1 and assists Cdc14 release from the nucleolus. Biochem. Biophys. Res. Commun. 2002, 294, 687–691. [Google Scholar] [CrossRef]
- Visintin, R.; Stegmeier, F.; Amon, A. The role of the polo kinase Cdc5 in controlling Cdc14 localization. Mol. Biol. Cell 2003, 14, 4486–4498. [Google Scholar] [CrossRef] [Green Version]
- Azzam, R.; Chen, S.L.; Shou, W.; Mah, A.S.; Alexandru, G.; Nasmyth, K.; Annan, R.S.; Carr, S.A.; Deshaies, R.J. Phosphorylation by cyclin B-Cdk underlies release of mitotic exit activator Cdc14 from the nucleolus. Science 2004, 305, 516–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queralt, E.; Lehane, C.; Novak, B.; Uhlmann, F. Downregulation of PP2A(Cdc55) phosphatase by separase initiates mitotic exit in budding yeast. Cell 2006, 125, 719–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahal, R.; Amon, A. The Polo-like kinase Cdc5 interacts with FEAR network components and Cdc14. Cell Cycle 2008, 7, 3262–3272. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Rodriguez, J.A.; Moyano, Y.; Jativa, S.; Queralt, E. Mitotic Exit Function of Polo-like Kinase Cdc5 Is Dependent on Sequential Activation by Cdk1. Cell Rep. 2016, 15, 2050–2062. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Li, W.; Liu, Y.; Amon, A. Cross-compartment signal propagation in the mitotic exit network. eLife 2021, 10, e63645. [Google Scholar] [CrossRef] [PubMed]
- Stegmeier, F.; Visintin, R.; Amon, A. Separase, polo kinase, the kinetochore protein Slk19, and Spo12 function in a network that controls Cdc14 localization during early anaphase. Cell 2002, 108, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Visintin, C.; Tomson, B.N.; Rahal, R.; Paulson, J.; Cohen, M.; Taunton, J.; Amon, A.; Visintin, R. APC/C-Cdh1-mediated degradation of the Polo kinase Cdc5 promotes the return of Cdc14 into the nucleolus. Genes Dev. 2008, 22, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Dulev, S.; De Renty, C.; Mehta, R.; Minkov, I.; Schwob, E.; Strunnikov, A. Essential global role of CDC14 in DNA synthesis revealed by chromosome underreplication unrecognized by checkpoints in cdc14 mutants. Proc. Natl. Acad. Sci. USA 2009, 106, 14466–14471. [Google Scholar] [CrossRef] [Green Version]
- Villoria, M.T.; Ramos, F.; Duenas, E.; Faull, P.; Cutillas, P.R.; Clemente-Blanco, A. Stabilization of the metaphase spindle by Cdc14 is required for recombinational DNA repair. EMBO J. 2017, 36, 79–101. [Google Scholar] [CrossRef]
- Mocciaro, A.; Schiebel, E. Cdc14: A highly conserved family of phosphatases with non-conserved functions? J. Cell Sci. 2010, 123, 2867–2876. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Ernsting, B.R.; Wishart, M.J.; Lohse, D.L.; Dixon, J.E. A family of putative tumor suppressors is structurally and functionally conserved in humans and yeast. J. Biol. Chem. 1997, 272, 29403–29406. [Google Scholar] [CrossRef] [Green Version]
- Rosso, L.; Marques, A.C.; Weier, M.; Lambert, N.; Lambot, M.A.; Vanderhaeghen, P.; Kaessmann, H. Birth and rapid subcellular adaptation of a hominoid-specific CDC14 protein. PLoS Biol. 2008, 6, e140. [Google Scholar] [CrossRef]
- Trautmann, S.; McCollum, D. Cell cycle: New functions for Cdc14 family phosphatases. Curr. Biol. 2002, 12, R733–R735. [Google Scholar] [CrossRef] [Green Version]
- Bassermann, F.; Frescas, D.; Guardavaccaro, D.; Busino, L.; Peschiaroli, A.; Pagano, M. The Cdc14B-Cdh1-Plk1 axis controls the G2 DNA-damage-response checkpoint. Cell 2008, 134, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Mocciaro, A.; Berdougo, E.; Zeng, K.; Black, E.; Vagnarelli, P.; Earnshaw, W.; Gillespie, D.; Jallepalli, P.; Schiebel, E. Vertebrate cells genetically deficient for Cdc14A or Cdc14B retain DNA damage checkpoint proficiency but are impaired in DNA repair. J. Cell Biol. 2010, 189, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Cuervo, H.; Bueno, A. Cds1 controls the release of Cdc14-like phosphatase Flp1 from the nucleolus to drive full activation of the checkpoint response to replication stress in fission yeast. Mol. Biol. Cell 2008, 19, 2488–2499. [Google Scholar] [CrossRef] [Green Version]
- Sharon, G.; Simchen, G. Mixed segregation of chromosomes during single-division meiosis of Saccharomyces cerevisiae. Genetics 1990, 125, 475–485. [Google Scholar] [CrossRef]
- Buonomo, S.B.; Rabitsch, K.P.; Fuchs, J.; Gruber, S.; Sullivan, M.; Uhlmann, F.; Petronczki, M.; Toth, A.; Nasmyth, K. Division of the nucleolus and its release of CDC14 during anaphase of meiosis I depends on separase, SPO12, and SLK19. Dev. Cell 2003, 4, 727–739. [Google Scholar] [CrossRef] [Green Version]
- Marston, A.L.; Lee, B.H.; Amon, A. The Cdc14 phosphatase and the FEAR network control meiotic spindle disassembly and chromosome segregation. Dev. Cell 2003, 4, 711–726. [Google Scholar] [CrossRef] [Green Version]
- Yellman, C.M.; Roeder, G.S. Cdc14 Early Anaphase Release, FEAR, Is Limited to the Nucleus and Dispensable for Efficient Mitotic Exit. PLoS ONE 2015, 10, e0128604. [Google Scholar]
- Kamieniecki, R.J.; Liu, L.; Dawson, D.S. FEAR but not MEN genes are required for exit from meiosis I. Cell Cycle 2005, 4, 1093–1098. [Google Scholar] [CrossRef] [Green Version]
- Pablo-Hernando, M.E.; Arnaiz-Pita, Y.; Nakanishi, H.; Dawson, D.; del Rey, F.; Neiman, A.M.; Vazquez de Aldana, C.R. Cdc15 is required for spore morphogenesis independently of Cdc14 in Saccharomyces cerevisiae. Genetics 2007, 177, 281–293. [Google Scholar] [CrossRef] [Green Version]
- Attner, M.A.; Amon, A. Control of the mitotic exit network during meiosis. Mol. Biol. Cell 2012, 23, 3122–3132. [Google Scholar] [CrossRef] [PubMed]
- Kamieniecki, R.J.; Shanks, R.M.; Dawson, D.S. Slk19p is necessary to prevent separation of sister chromatids in meiosis I. Curr. Biol. 2000, 10, 1182–1190. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Saunders, W.S. The Saccharomyces cerevisiae centromere protein Slk19p is required for two successive divisions during meiosis. Genetics 2000, 155, 577–587. [Google Scholar] [CrossRef]
- Kerr, G.W.; Sarkar, S.; Tibbles, K.L.; Petronczki, M.; Millar, J.B.; Arumugam, P. Meiotic nuclear divisions in budding yeast require PP2ACdc55-mediated antagonism of Net1 phosphorylation by Cdk. J. Cell Biol. 2011, 193, 1157–1166. [Google Scholar] [CrossRef] [Green Version]
- Bizzari, F.; Marston, A.L. Cdc55 coordinates spindle assembly and chromosome disjunction during meiosis. J. Cell Biol. 2011, 193, 1213–1228. [Google Scholar] [CrossRef] [PubMed]
- Nolt, J.K.; Rice, L.M.; Gallo-Ebert, C.; Bisher, M.E.; Nickels, J.T. PP2ACdc55 is required for multiple events during meiosis I. Cell Cycle 2011, 10, 1420–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, C.; Zou, J.; Rappsilber, J.; Marston, A.L. Cdc14 phosphatase directs centrosome re-duplication at the meiosis I to meiosis II transition in budding yeast. Wellcome Open Res. 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borner, G.V.; Kleckner, N.; Hunter, N. Crossover/noncrossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of meiosis. Cell 2004, 117, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Briza, P.; Winkler, G.; Kalchhauser, H.; Breitenbach, M. Dityrosine is a prominent component of the yeast ascospore wall. A proof of its structure. J. Biol. Chem. 1986, 261, 4288–4294. [Google Scholar] [CrossRef]
- Bloom, J.; Cross, F.R. Novel role for Cdc14 sequestration: Cdc14 dephosphorylates factors that promote DNA replication. Mol. Cell Biol. 2007, 27, 842–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schild, D.; Byers, B. Diploid spore formation and other meiotic effects of two cell-division-cycle mutations of Saccharomyces cerevisiae. Genetics 1980, 96, 859–876. [Google Scholar] [CrossRef]
- Lee, B.H.; Amon, A. Role of Polo-like kinase CDC5 in programming meiosis I chromosome segregation. Science 2003, 300, 482–486. [Google Scholar] [CrossRef]
- Straight, A.F.; Belmont, A.S.; Robinett, C.C.; Murray, A.W. GFP tagging of budding yeast chromosomes reveals that protein-protein interactions can mediate sister chromatid cohesion. Curr. Biol. 1996, 6, 1599–1608. [Google Scholar] [CrossRef] [Green Version]
- Michaelis, C.; Ciosk, R.; Nasmyth, K. Cohesins: Chromosomal Proteins that Prevent Premature Separation of Sister Chromatids. Cell 1997, 91, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Katis, V.L.; Lipp, J.J.; Imre, R.; Bogdanova, A.; Okaz, E.; Habermann, B.; Mechtler, K.; Nasmyth, K.; Zachariae, W. Rec8 phosphorylation by casein kinase 1 and Cdc7-Dbf4 kinase regulates cohesin cleavage by separase during meiosis. Dev. Cell 2010, 18, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shonn, M.A.; McCarroll, R.; Murray, A.W. Spo13 protects meiotic cohesin at centromeres in meiosis I. Genes Dev. 2002, 16, 1659–1671. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Alani, E.; Kleckner, N. A pathway for generation and processing of double-strand breaks during meiotic recombination in S. cerevisiae. Cell 1990, 61, 1089–1101. [Google Scholar] [CrossRef]
- Schwacha, A.; Kleckner, N. Identification of joint molecules that form frequently between homologs but rarely between sister chromatids during yeast meiosis. Cell 1994, 76, 51–63. [Google Scholar] [CrossRef]
- Hunter, N.; Kleckner, N. The single-end invasion: An asymmetric intermediate at the double-strand break to double-holliday junction transition of meiotic recombination. Cell 2001, 106, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Martini, E.; Diaz, R.L.; Hunter, N.; Keeney, S. Crossover homeostasis in yeast meiosis. Cell 2006, 126, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Shinohara, M.; Oh, S.D.; Hunter, N.; Shinohara, A. Crossover assurance and crossover interference are distinctly regulated by the ZMM proteins during yeast meiosis. Nat. Genet. 2008, 40, 299–309. [Google Scholar] [CrossRef]
- Allers, T.; Lichten, M. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 2001, 106, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Carlile, T.M.; Amon, A. Meiosis I is established through division-specific translational control of a cyclin. Cell 2008, 133, 280–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Luis, J.; Clemente-Blanco, A.; Aragon, L.; Machin, F. Cdc14 targets the Holliday junction resolvase Yen1 to the nucleus in early anaphase. Cell Cycle 2014, 13, 1392–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mankouri, H.W.; Ashton, T.M.; Hickson, I.D. Holliday junction-containing DNA structures persist in cells lacking Sgs1 or Top3 following exposure to DNA damage. Proc. Natl. Acad. Sci. USA 2011, 108, 4944–4949. [Google Scholar] [CrossRef] [Green Version]
- Bzymek, M.; Thayer, N.H.; Oh, S.D.; Kleckner, N.; Hunter, N. Double Holliday junctions are intermediates of DNA break repair. Nature 2010, 464, 937–941. [Google Scholar] [CrossRef] [Green Version]
- Cromie, G.A.; Hyppa, R.W.; Taylor, A.F.; Zakharyevich, K.; Hunter, N.; Smith, G.R. Single Holliday junctions are intermediates of meiotic recombination. Cell 2006, 127, 1167–1178. [Google Scholar] [CrossRef] [Green Version]
- Liberi, G.; Maffioletti, G.; Lucca, C.; Chiolo, I.; Baryshnikova, A.; Cotta-Ramusino, C.; Lopes, M.; Pellicioli, A.; Haber, J.E.; Foiani, M. Rad51-dependent DNA structures accumulate at damaged replication forks in sgs1 mutants defective in the yeast ortholog of BLM RecQ helicase. Genes Dev. 2005, 19, 339–350. [Google Scholar] [CrossRef] [Green Version]
- Schwacha, A.; Kleckner, N. Identification of double Holliday junctions as intermediates in meiotic recombination. Cell 1995, 83, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Arguello-Miranda, O.; Zagoriy, I.; Mengoli, V.; Rojas, J.; Jonak, K.; Oz, T.; Graf, P.; Zachariae, W. Casein Kinase 1 Coordinates Cohesin Cleavage, Gametogenesis, and Exit from M Phase in Meiosis II. Dev. Cell 2017, 40, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef] [Green Version]
- Katis, V.L.; Matos, J.; Mori, S.; Shirahige, K.; Zachariae, W.; Nasmyth, K. Spo13 facilitates monopolin recruitment to kinetochores and regulates maintenance of centromeric cohesion during yeast meiosis. Curr. Biol. 2004, 14, 2183–2196. [Google Scholar] [CrossRef] [Green Version]
- Talhaoui, I.; Bernal, M.; Mullen, J.R.; Dorison, H.; Palancade, B.; Brill, S.J.; Mazon, G. Slx5-Slx8 ubiquitin ligase targets active pools of the Yen1 nuclease to limit crossover formation. Nat. Commun. 2018, 9, 5016. [Google Scholar] [CrossRef]
- Tang, S.; Wu, M.K.Y.; Zhang, R.; Hunter, N. Pervasive and essential roles of the Top3-Rmi1 decatenase orchestrate recombination and facilitate chromosome segregation in meiosis. Mol. Cell 2015, 57, 607–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shodhan, A.; Medhi, D.; Lichten, M. Noncanonical Contributions of MutLgamma to VDE-Initiated Crossovers During Saccharomyces cerevisiae Meiosis. G3 Genes Genomes Genet. 2019, 9, 1647–1654. [Google Scholar]
- Cannavo, E.; Sanchez, A.; Anand, R.; Ranjha, L.; Hugener, J.; Adam, C.; Acharya, A.; Weyland, N.; Aran-Guiu, X.; Charbonnier, J.B.; et al. Regulation of the MLH1-MLH3 endonuclease in meiosis. Nature 2020, 586, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, D.S.; Owens, S.N.; Honda, M.; Ito, M.; Yang, Y.; Corrigan, M.W.; Chen, L.; Quan, A.L.; Hunter, N. PCNA activates the MutLgamma endonuclease to promote meiotic crossing over. Nature 2020, 586, 623–627. [Google Scholar] [CrossRef]
- Tsuchiya, D.; Yang, Y.; Lacefield, S. Positive feedback of NDT80 expression ensures irreversible meiotic commitment in budding yeast. PLoS Genet. 2014, 10, e1004398. [Google Scholar] [CrossRef] [Green Version]
- Imtiaz, A.; Belyantseva, I.A.; Beirl, A.J.; Fenollar-Ferrer, C.; Bashir, R.; Bukhari, I.; Bouzid, A.; Shaukat, U.; Azaiez, H.; Booth, K.T.; et al. CDC14A phosphatase is essential for hearing and male fertility in mouse and human. Hum. Mol. Genet. 2018, 27, 780–798. [Google Scholar] [CrossRef] [Green Version]
- Longtine, M.S.; McKenzie, A., 3rd; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Jaspersen, S.L.; Charles, J.F.; Tinker-Kulberg, R.L.; Morgan, D.O. A late mitotic regulatory network controlling cyclin destruction in Saccharomyces cerevisiae. Mol. Biol. Cell 1998, 9, 2803–2817. [Google Scholar] [CrossRef] [Green Version]
- Padmore, R.; Cao, L.; Kleckner, N. Temporal comparison of recombination and synaptonemal complex formation during meiosis in S. cerevisiae. Cell 1991, 66, 1239–1256. [Google Scholar] [CrossRef]
- Carballo, J.A.; Panizza, S.; Serrentino, M.E.; Johnson, A.L.; Geymonat, M.; Borde, V.; Klein, F.; Cha, R.S. Budding yeast ATM/ATR control meiotic double-strand break (DSB) levels by down-regulating Rec114, an essential component of the DSB-machinery. PLoS Genet. 2013, 9, e1003545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahuja, J.S.; Borner, G.V. Analysis of meiotic recombination intermediates by two-dimensional gel electrophoresis. Methods Mol. Biol. 2011, 745, 99–116. [Google Scholar]
- Newnham, L.; Jordan, P.W.; Carballo, J.A.; Newcombe, S.; Hoffmann, E. Ipl1/Aurora kinase suppresses S-CDK-driven spindle formation during prophase I to ensure chromosome integrity during meiosis. PLoS ONE 2013, 8, e83982. [Google Scholar] [CrossRef] [PubMed]
- Gimenez-Abian, M.I.; Rozalen, A.E.; Carballo, J.A.; Botella, L.M.; Pincheira, J.; Lopez-Saez, J.F.; De la Torre, C. HSP90 and checkpoint-dependent lengthening of the G2 phase observed in plant cells under hypoxia and cold. Protoplasma 2004, 223, 191–196. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso-Ramos, P.; Álvarez-Melo, D.; Strouhalova, K.; Pascual-Silva, C.; Garside, G.B.; Arter, M.; Bermejo, T.; Grigaitis, R.; Wettstein, R.; Fernández-Díaz, M.; et al. The Cdc14 Phosphatase Controls Resolution of Recombination Intermediates and Crossover Formation during Meiosis. Int. J. Mol. Sci. 2021, 22, 9811. https://doi.org/10.3390/ijms22189811
Alonso-Ramos P, Álvarez-Melo D, Strouhalova K, Pascual-Silva C, Garside GB, Arter M, Bermejo T, Grigaitis R, Wettstein R, Fernández-Díaz M, et al. The Cdc14 Phosphatase Controls Resolution of Recombination Intermediates and Crossover Formation during Meiosis. International Journal of Molecular Sciences. 2021; 22(18):9811. https://doi.org/10.3390/ijms22189811
Chicago/Turabian StyleAlonso-Ramos, Paula, David Álvarez-Melo, Katerina Strouhalova, Carolina Pascual-Silva, George B. Garside, Meret Arter, Teresa Bermejo, Rokas Grigaitis, Rahel Wettstein, Marta Fernández-Díaz, and et al. 2021. "The Cdc14 Phosphatase Controls Resolution of Recombination Intermediates and Crossover Formation during Meiosis" International Journal of Molecular Sciences 22, no. 18: 9811. https://doi.org/10.3390/ijms22189811
APA StyleAlonso-Ramos, P., Álvarez-Melo, D., Strouhalova, K., Pascual-Silva, C., Garside, G. B., Arter, M., Bermejo, T., Grigaitis, R., Wettstein, R., Fernández-Díaz, M., Matos, J., Geymonat, M., San-Segundo, P. A., & Carballo, J. A. (2021). The Cdc14 Phosphatase Controls Resolution of Recombination Intermediates and Crossover Formation during Meiosis. International Journal of Molecular Sciences, 22(18), 9811. https://doi.org/10.3390/ijms22189811