
1,5-Anhydro-D-fructose Protects against Rotenone-Induced Neuronal Damage In Vitro through Mitochondrial Biogenesis

 ,
, 
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
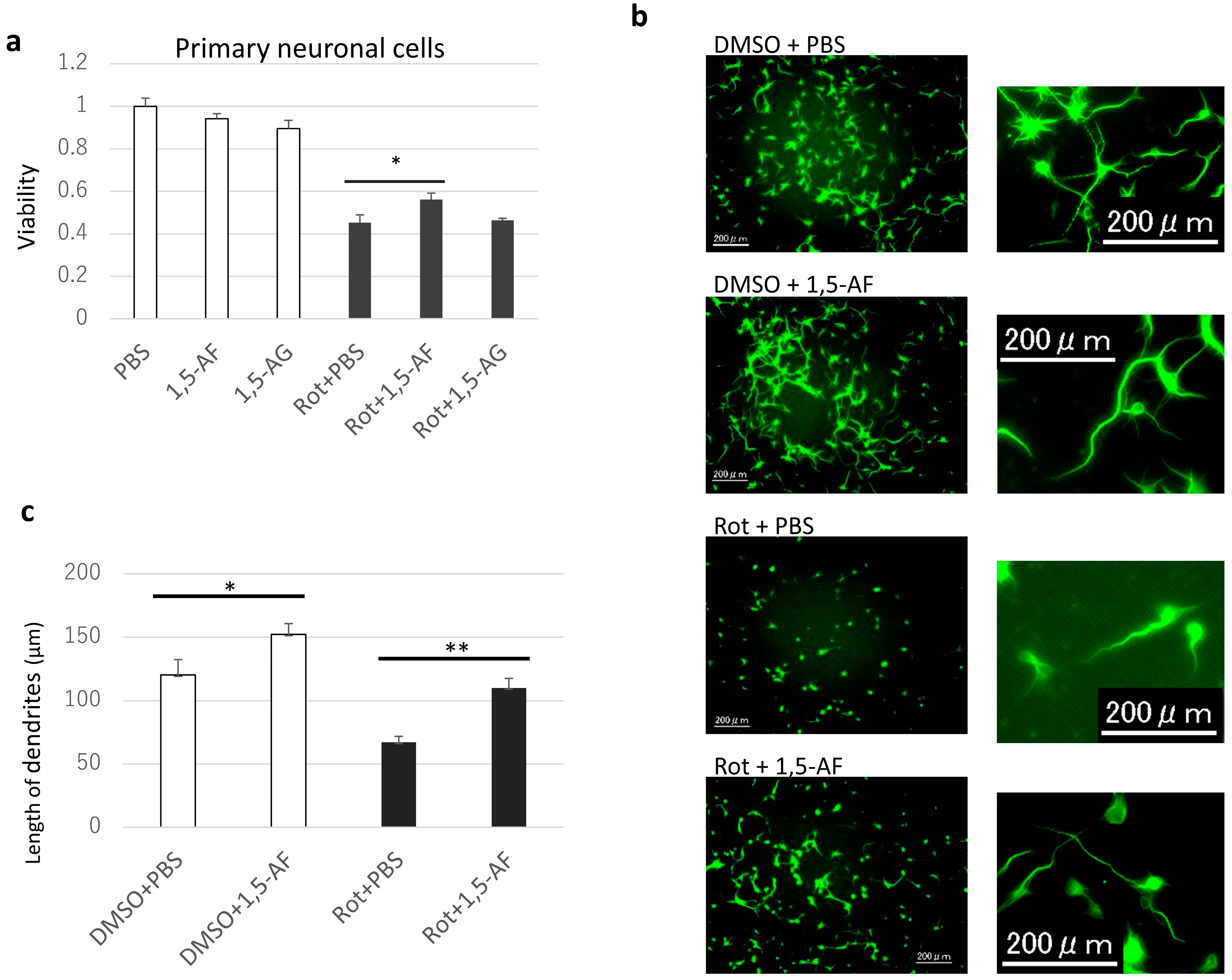
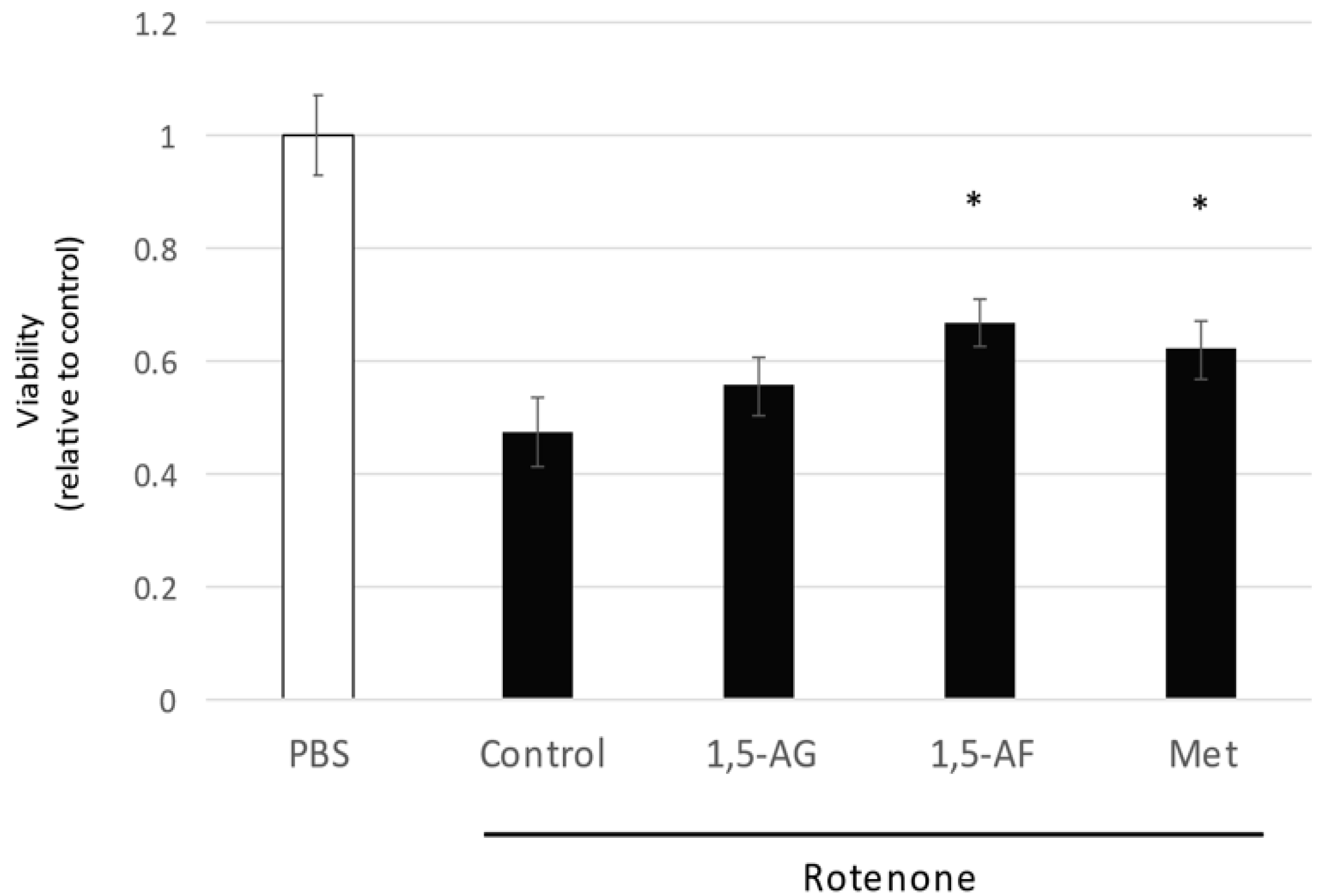
2.1. 1,5-AF Ameliorates Rotenone-Induced Cytotoxic Effects in a Dose-Dependent Manner
2.2. 1,5-AF Upregulates PGC-1α Expression and Deacetylation While Increasing Phosphorylated AMPK
2.3. PGC-1α Silencing Inhibits 1,5-AF-Mediated Protection against Rotenone-Induced Cytotoxicity
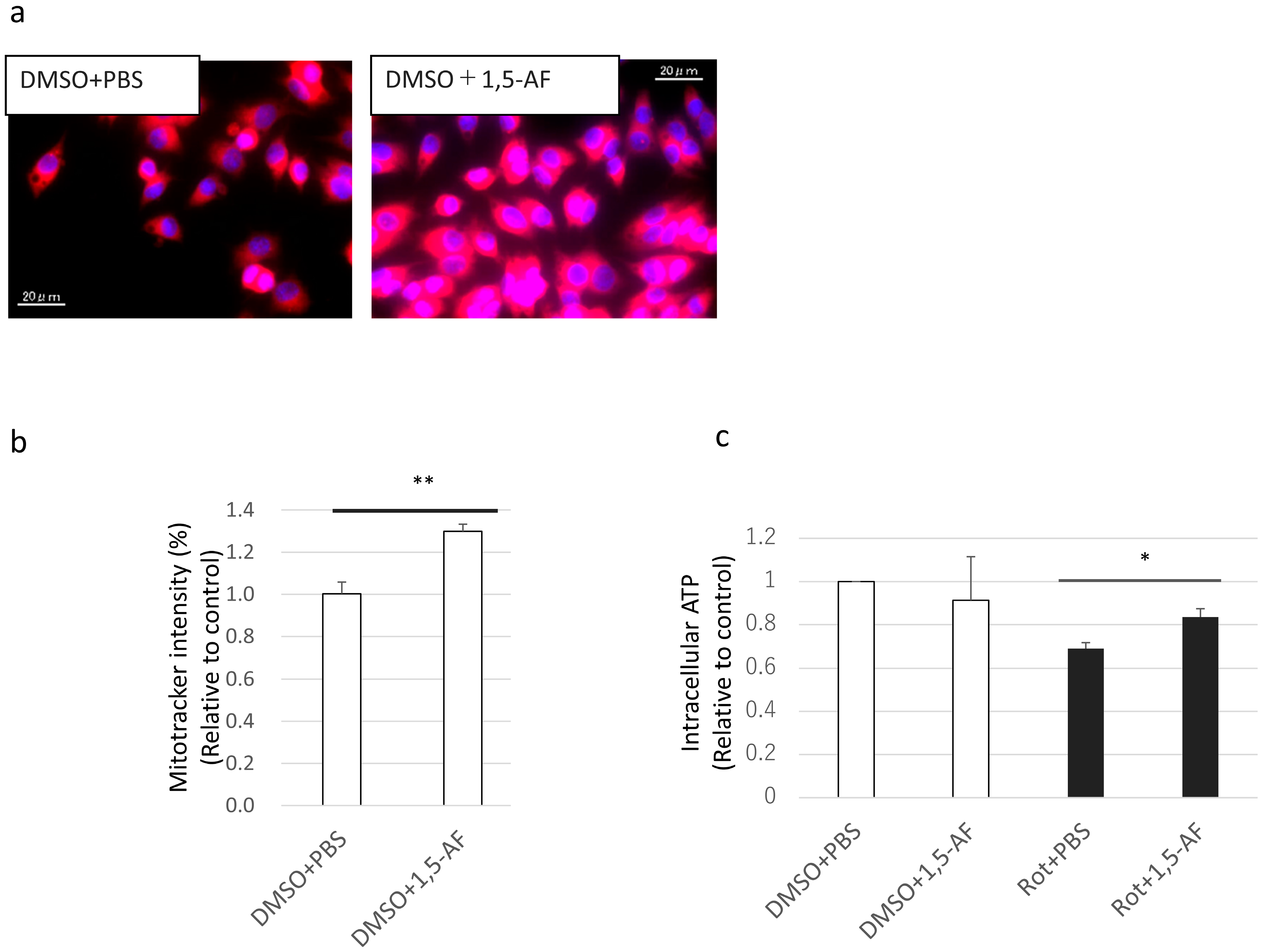
2.4. 1,5-AF Increases MitoTracker Intensity and Activity of Mitochondria
2.5. PGC-1α Silencing Inhibits the Effects of 1,5-AF on Mitochondria
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Stimulation
4.3. MTT Assay to Analyze Cell Viability
4.4. Anti-MAP2 Immunofluorescence Staining
4.5. Immunoblotting
4.6. PGC-1α Acetylation Assays
4.7. Silencing of PGC-1α Using PPARGC1A siRNA for the MTT Assay
4.8. Analysis of Mitochondrial Mass by MitoTracker
4.9. Intracellular ATP Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Lau, L.M.; Giesbergen, P.C.; de Rijk, M.C.; Hofman, A.; Koudstaal, P.J.; Breteler, M.M. Incidence of parkinsonism and Parkinson disease in a general population: The Rotterdam Study. Neurology 2004, 63, 1240–1244. [Google Scholar] [CrossRef] [PubMed]
- Linder, J.; Stenlund, H.; Forsgren, L. Incidence of Parkinson’s disease and parkinsonism in northern Sweden: A population-based study. Mov. Disord. 2010, 25, 341–348. [Google Scholar] [CrossRef]
- Shults, C.W.; Oakes, D.; Kieburtz, K.; Beal, M.F.; Haas, R.; Plumb, S.; Juncos, J.L.; Nutt, J.; Shoulson, I.; Carter, J.; et al. Effects of coenzyme Q10 in early Parkinson disease: Evidence of slowing of the functional decline. Arch. Neurol. 2002, 59, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Jenner, P. Parkinson’s disease, pesticides and mitochondrial dysfunction. Trends Neurosci. 2001, 24, 245–247. [Google Scholar] [CrossRef]
- Ullah, I.; Zhao, L.; Hai, Y.; Fahim, M.; Alwayli, D.; Wang, X.; Li, H. Metal elements and pesticides as risk factors for Parkinson’s disease–A review. Toxicol. Rep. 2021, 8, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Pickrell, A.M.; Youle, R.J. The roles of PINK1, parkin, and mitochondrial fidelity in Parkinson’s disease. Neuron 2015, 85, 257–273. [Google Scholar] [CrossRef] [Green Version]
- Exner, N.; Lutz, A.K.; Haass, C.; Winklhofer, K.F. Mitochondrial dysfunction in Parkinson’s disease: Molecular mechanisms and pathophysiological consequences. EMBO J. 2012, 31, 3038–3062. [Google Scholar] [CrossRef] [Green Version]
- Gibson, G.E.; Huang, H.M. Mitochondrial enzymes and endoplasmic reticulum calcium stores as targets of oxidative stress in neurodegenerative diseases. J. Bioenerg. Biomembr. 2004, 36, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Greene, J.C.; Whitworth, A.J.; Kuo, I.; Andrews, L.A.; Feany, M.B.; Pallanck, L.J. Mitochondrial pathology and apoptotic muscle degeneration in Drosophila parkin mutants. Proc. Natl. Acad. Sci. USA 2003, 100, 4078–4083. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Ragheb, K.; Lawler, G.; Sturgis, J.; Rajwa, B.; Melendez, J.A.; Robinson, J.P. Mitochondrial complex I inhibitor rotenone induces apoptosis through enhancing mitochondrial reactive oxygen species production. J. Biol. Chem. 2003, 278, 8516–8525. [Google Scholar] [CrossRef] [Green Version]
- Inden, M.; Kitamura, Y.; Abe, M.; Tamaki, A.; Takata, K.; Taniguchi, T. Parkinsonian rotenone mouse model: Reevaluation of long-term administration of rotenone in C57BL/6 mice. Biol. Pharm. Bull. 2011, 34, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Liao, Z.; Locascio, J.J.; Lesniak, K.A.; Roderick, S.S.; Watt, M.L.; Eklund, A.C.; Zhang-James, Y.; Kim, P.D.; Hauser, M.A.; et al. PGC-1alpha, a potential therapeutic target for early intervention in Parkinson’s disease. Sci. Transl. Med. 2010, 2, 52ra73. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, A.; Chinta, S.J.; Mallajosyula, J.K.; Rajagopolan, S.; Hanson, I.; Rane, A.; Melov, S.; Andersen, J.K. Selective binding of nuclear alpha-synuclein to the PGC1alpha promoter under conditions of oxidative stress may contribute to losses in mitochondrial function: Implications for Parkinson’s disease. Free Radic. Biol. Med. 2012, 53, 993–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.C.; Lulla, A.; Stahl, M.C.; Wheat, N.D.; Bronstein, J.M.; Sagasti, A. Axon degeneration and PGC-1α-mediated protection in a zebrafish model of α-synuclein toxicity. Dis. Models Mech. 2014, 7, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Fujisue, M.; Yoshinaga, K.; Muroya, K.; Abel, J.I.; Hizukuri, S. Preparation and antioxidant activity of 1,5-anhhydrofructose. J. Appl. Glycosci. 1999, 46, 439. [Google Scholar] [CrossRef] [Green Version]
- Fiskesund, R.; Abeyama, K.; Yoshinaga, K.; Abe, J.; Yuan, Y.; Yu, S. 1,5-anhydro-D-fructose and its derivatives: Biosynthesis, preparation and potential medical applications. Planta Med. 2010, 76, 1635–1641. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, K.; Sarker, K.P.; Maruyama, I.; Hizukuri, S. Antioxidant effects of 1,5-anhydro-D-fructose, a new natural sugar, In Vitro. Planta Med. 2002, 68, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Akanuma, Y.; Morita, M.; Fukuzawa, N.; Yamanouchi, T.; Akanuma, H. Urinary excretion of 1,5-anhydro-D-glucitol accompanying glucose excretion in diabetic patients. Diabetologia 1988, 31, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Kawahara, K.; Matsushita, K.; Nawa, Y.; Shrestha, B.; Kikuchi, K.; Sameshima, H.; Hashiguchi, T.; Maruyama, I. Attenuation of LPS-induced iNOS expression by 1,5-anhydro-d-fructose. Biochem. Biophys. Res. Commun. 2009, 387, 42–46. [Google Scholar] [CrossRef]
- Rena, G.; Hardie, D.G.; Pearson, E.R. The mechanisms of action of metformin. Diabetologia 2017, 60, 1577–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.; Liang, X.; Rogers, C.Q.; Rideout, D.; You, M. Involvement of adiponectin-SIRT1-AMPK signaling in the protective action of rosiglitazone against alcoholic fatty liver in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G364–G374. [Google Scholar] [CrossRef] [Green Version]
- Kukidome, D.; Nishikawa, T.; Sonoda, K.; Imoto, K.; Fujisawa, K.; Yano, M.; Motoshima, H.; Taguchi, T.; Matsumura, T.; Araki, E. Activation of AMP-activated protein kinase reduces hyperglycemia-induced mitochondrial reactive oxygen species production and promotes mitochondrial biogenesis in human umbilical vein endothelial cells. Diabetes 2006, 55, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Song, M.Y.; Jung, H.W.; Kang, S.Y.; Park, Y.K. Atractylenolide III Enhances Energy Metabolism by Increasing the SIRT-1 and PGC1alpha Expression with AMPK Phosphorylation in C2C12 Mouse Skeletal Muscle Cells. Biol. Pharm. Bull. 2017, 40, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Pitkänen, E.; Pitkänen, O.M. Renal tubular reabsorption of 1,5-anhydro-D-glucitol and D-mannose in vivo in the rat. Pflug. Arch. 1992, 420, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Okamoto, K.; Hayashi, Y.; Sheng, M. The importance of dendritic mitochondria in the morphogenesis and plasticity of spines and synapses. Cell 2004, 119, 873–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlgren, H.; Bas-Orth, C.; Freitag, H.E.; Hellwig, A.; Ottersen, O.P.; Bading, H. The nuclear calcium signaling target, activating transcription factor 3 (ATF3), protects against dendrotoxicity and facilitates the recovery of synaptic transmission after an excitotoxic insult. J. Biol. Chem. 2014, 289, 9970–9982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olney, J.W.; Fuller, T.; de Gubareff, T. Acute dendrotoxic changes in the hippocampus of kainate treated rats. Brain Res. 1979, 176, 91–100. [Google Scholar] [CrossRef]
- McNeill, T.H.; Brown, S.A.; Rafols, J.A.; Shoulson, I. Atrophy of medium spiny I striatal dendrites in advanced Parkinson’s disease. Brain Res. 1988, 455, 148–152. [Google Scholar] [CrossRef]
- Zaja-Milatovic, S.; Milatovic, D.; Schantz, A.M.; Zhang, J.; Montine, K.S.; Samii, A.; Deutch, A.Y.; Montine, T.J. Dendritic degeneration in neostriatal medium spiny neurons in Parkinson disease. Neurology 2005, 64, 545–547. [Google Scholar] [CrossRef]
- Chau, M.D.L.; Gao, J.P.; Yang, Q.; Wu, Z.D.; Gromada, J. Fibroblast growth factor 21 regulates energy metabolism by activating the AMPK-SIRT1-PGC-1 alpha pathway. Proc. Natl. Acad. Sci. USA 2010, 107, 12553–12558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canto, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Jager, S.; Handschin, C.; Pierre, J.; Spiegelman, B.M. AMP-activated protein kinase (AMPK) action in skeletal muscle via direct phosphorylation of PGC-1 alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 12017–12022. [Google Scholar] [CrossRef] [Green Version]
- Zong, H.; Ren, J.M.; Young, L.H.; Pypaert, M.; Mu, J.; Birnbaum, M.J.; Shulman, G.I. AMP kinase is required for mitochondrial biogenesis in skeletal muscle in response to chronic energy deprivation. Proc. Natl. Acad. Sci. USA 2002, 99, 15983–15987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenyk-Melody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujii, N.; et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Investig. 2001, 108, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Aryal, B.; Lee, Y. Disease model organism for Parkinson disease: Drosophila melanogaster. BMB Rep. 2019, 52, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.V.; Chaube, B.; Mayengbam, S.S.; Singh, A.; Malvi, P.; Mohammad, N.; Deb, A.; Bhat, M.K. Metformin induced lactic acidosis impaired response of cancer cells towards paclitaxel and doxorubicin: Role of monocarboxylate transporter. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166011. [Google Scholar] [CrossRef]
- DeFronzo, R.; Fleming, G.A.; Chen, K.; Bicsak, T.A. Metformin-associated lactic acidosis: Current perspectives on causes and risk. Metabolism 2016, 65, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Isenberg, J.S.; Klaunig, J.E. Role of the mitochondrial membrane permeability transition (MPT) in rotenone-induced apoptosis in liver cells. Toxicol. Sci. 2000, 53, 340–351. [Google Scholar] [CrossRef] [Green Version]
- Puleston, D. Detection of Mitochondrial Mass, Damage, and Reactive Oxygen Species by Flow Cytometry. Cold Spring Harb. Protoc. 2015, 2015, pdb.prot086298. [Google Scholar] [CrossRef]
- Kametani, S.; Shiga, Y.; Akanuma, H. Hepatic production of 1,5-anhydrofructose and 1,5-anhydroglucitol in rat by the third glycogenolytic pathway. Eur. J. Biochem. 1996, 242, 832–838. [Google Scholar] [CrossRef]
- Webb, J.L.; Ravikumar, B.; Atkins, J.; Skepper, J.N.; Rubinsztein, D.C. alpha-synuclein is degraded by both autophagy and the proteasome. J. Biol. Chem. 2003, 278, 25009–25013. [Google Scholar] [CrossRef] [Green Version]
- Li, D.W.; Yao, M.; Dong, Y.H.; Tang, M.N.; Chen, W.; Li, G.R.; Sun, B.Q. Guanosine exerts neuroprotective effects by reversing mitochondrial dysfunction in a cellular model of Parkinson’s disease. Int. J. Mol. Med. 2014, 34, 1358–1364. [Google Scholar] [CrossRef]
- Kikuchi, K.; Kawahara, K.; Tancharoen, S.; Matsuda, F.; Morimoto, Y.; Ito, T.; Biswas, K.K.; Takenouchi, K.; Miura, N.; Oyama, Y.; et al. The free radical scavenger edaravone rescues rats from cerebral infarction by attenuating the release of high-mobility group box-1 in neuronal cells. J. Pharmacol. Exp. Ther. 2009, 329, 865–874. [Google Scholar] [CrossRef]
- Kambe, Y.; Miyata, A. Role of mitochondrial activation in PACAP dependent neurite outgrowth. J. Mol. Neurosci. 2012, 48, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Radad, K.; Scheller, D.; Rausch, W.D.; Reichmann, H.; Gille, G. Neuroprotective effect of rotigotine against complex I inhibitors, MPP(+) and rotenone, in primary mesencephalic cell culture. Folia Neuropathol. 2014, 52, 179–186. [Google Scholar] [CrossRef]
- Choi, W.S.; Kim, H.W.; Xia, Z. JNK inhibition of VMAT2 contributes to rotenone-induced oxidative stress and dopamine neuron death. Toxicology 2015, 328, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Zeng, T.; Zhang, C.L.; Song, F.Y.; Zhao, X.L.; Xie, K.Q. CMZ Reversed Chronic Ethanol-Induced Disturbance of PPAR-alpha Possibly by Suppressing Oxidative Stress and PGC-1 alpha Acetylation, and Activating the MAPK and GSK3 beta Pathway. PLoS ONE 2014, 9, 15. [Google Scholar] [CrossRef]
- Panta, S.; Yamakuchi, M.; Shimizu, T.; Takenouchi, K.; Oyama, Y.; Koriyama, T.; Kojo, T.; Hashiguchi, T. Low grade inflammation inhibits VEGF induced HUVECs migration in p53 dependent manner. Biochem. Biophys. Res. Commun. 2017, 483, 803–809. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasamo, Y.; Kikuchi, K.; Yamakuchi, M.; Otsuka, S.; Takada, S.; Kambe, Y.; Ito, T.; Kawahara, K.-i.; Arita, K.; Yoshimoto, K.; et al. 1,5-Anhydro-D-fructose Protects against Rotenone-Induced Neuronal Damage In Vitro through Mitochondrial Biogenesis. Int. J. Mol. Sci. 2021, 22, 9941. https://doi.org/10.3390/ijms22189941
Kasamo Y, Kikuchi K, Yamakuchi M, Otsuka S, Takada S, Kambe Y, Ito T, Kawahara K-i, Arita K, Yoshimoto K, et al. 1,5-Anhydro-D-fructose Protects against Rotenone-Induced Neuronal Damage In Vitro through Mitochondrial Biogenesis. International Journal of Molecular Sciences. 2021; 22(18):9941. https://doi.org/10.3390/ijms22189941
Chicago/Turabian StyleKasamo, Yuki, Kiyoshi Kikuchi, Munekazu Yamakuchi, Shotaro Otsuka, Seiya Takada, Yuki Kambe, Takashi Ito, Ko-ichi Kawahara, Kazunori Arita, Koji Yoshimoto, and et al. 2021. "1,5-Anhydro-D-fructose Protects against Rotenone-Induced Neuronal Damage In Vitro through Mitochondrial Biogenesis" International Journal of Molecular Sciences 22, no. 18: 9941. https://doi.org/10.3390/ijms22189941
APA StyleKasamo, Y., Kikuchi, K., Yamakuchi, M., Otsuka, S., Takada, S., Kambe, Y., Ito, T., Kawahara, K. -i., Arita, K., Yoshimoto, K., & Maruyama, I. (2021). 1,5-Anhydro-D-fructose Protects against Rotenone-Induced Neuronal Damage In Vitro through Mitochondrial Biogenesis. International Journal of Molecular Sciences, 22(18), 9941. https://doi.org/10.3390/ijms22189941