
The Positive and Negative Immunoregulatory Role of B7 Family: Promising Novel Targets in Gastric Cancer Treatment


 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. B7-1 (CD80) and B7-2 (CD86)
3. B7-H2 (ICOSL)
4. B7-H1 (PD-L1) and B7-DC (PD-L2)
5. B7-H3 (CD276)
6. B7-H4 (B7x)
7. B7-H5 (VISTA)
8. B7-H6 (NCR3LG1)
9. B7-H7 (HHLA2)
10. Ig-like Domain-Containing Receptor 2 (ILDR2)
11. Treatment of Gastric Cancer Base on B7 Family Inhibition
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [Green Version]
- Hamashima, C.; Systematic Review Group; Systematic Review Group and Guideline Development Group for Gastric Cancer Screening Guidelines. Update version of the Japanese Guidelines for Gastric Cancer Screening. Jpn. J. Clin. Oncol. 2018, 48, 673–683. [Google Scholar] [CrossRef] [Green Version]
- Alsina, M.; Moehler, M.; Hierro, C.; Guardeño, R.; Tabernero, J. Immunotherapy for gastric cancer: A focus on immune checkpoints. Target. Oncol. 2016, 11, 469–477. [Google Scholar] [CrossRef]
- Abdul-Latif, M.; Townsend, K.; Dearman, C.; Shiu, K.K.; Khan, K. Immunotherapy in gastrointestinal cancer: The current scenario and future perspectives. Cancer Treat. Rev. 2020, 88, 102030. [Google Scholar] [CrossRef]
- Su, M.; Lin, Y.; Cui, C.; Tian, X.; Lai, L. ERMAP is a B7 family-related molecule that negatively regulates T cell and macrophage responses. Cell. Mol. Immunol. 2020, 18, 1920–1933. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Dong, C. New B7 family checkpoints in human cancers. Mol. Cancer Ther. 2017, 16, 1203–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Zang, X. Structures of Immune Checkpoints: An Overview on the CD28-B7 Family. In Structural Immunology; Springer: Berlin/Heidelberg, Germany, 2019; Volume 1172, pp. 63–78. [Google Scholar]
- Ye, Q.; Liu, J.; Xie, K. B7 Family proteins in cancer progression: Immunological and non-immunological functions. J. Cancer Treat. Diagn. 2019, 3, 1–6. [Google Scholar] [CrossRef]
- Kong, X. Discovery of new immune checkpoints: Family grows up. In Regulation of Cancer Immune Checkpoints; Springer: Berlin/Heidelberg, Germany, 2020; Volume 1248, pp. 61–82. [Google Scholar]
- Chapoval, A.; Chapoval, S.; Shcherbakova, N.; Shcherbakov, D. Immune checkpoints of the B7 family. Part 2. Representatives of the B7 family B7-H3, B7-H4, B7-H5, B7-H6, B7-H7, and ILDR2 and their receptors. Russ. J. Bioorg. Chem. 2019, 45, 321–334. [Google Scholar] [CrossRef]
- Barrueto, L.; Caminero, F.; Cash, L.; Makris, C.; Lamichhane, P.; Deshmukh, R.R. Resistance to checkpoint inhibition in cancer immunotherapy. Transl. Oncol. 2020, 13, 100738. [Google Scholar] [CrossRef]
- Sadreddini, S.; Baradaran, B.; Aghebati-Maleki, A.; Sadreddini, S.; Shanehbandi, D.; Fotouhi, A.; Aghebati-Maleki, L. Immune checkpoint blockade opens a new way to cancer immunotherapy. J. Cell. Physiol. 2019, 234, 8541–8549. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.F.; Li, Y. Small-molecule targets in tumor immunotherapy. Nat. Prod. Bioprospect. 2018, 8, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, S.; Edidin, M.; Almo, S.C.; Nathenson, S.G. Different cell surface oligomeric states of B7-1 and B7-2: Implications for signaling. Proc. Natl. Acad. Sci. USA 2005, 102, 15569–15574. [Google Scholar] [CrossRef] [Green Version]
- Fleischer, J.; Soeth, E.; Reiling, N.; Grage-Griebenow, E.; Flad, H.D.; Ernst, M. Differential expression and function of CD80 (B7-1) and CD86 (B7-2) on human peripheral blood monocytes. Immunology 1996, 89, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Esensten, J.H.; Helou, Y.A.; Chopra, G.; Weiss, A.; Bluestone, J.A. CD28 Costimulation: From mechanism to therapy. Immunity 2016, 44, 973–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elloso, M.M.; Scott, P. Expression and contribution of B7-1 (CD80) and B7-2 (CD86) in the early immune response to Leishmania major infection. J. Immunol. 1999, 162, 6708–6715. [Google Scholar] [PubMed]
- Brunet, J.F.; Denizot, F.; Luciani, M.F.; Roux-Dosseto, M.; Suzan, M.; Mattei, M.G.; Golstein, P. A new member of the immunoglobulin superfamily—CTLA-4. Nature 1987, 328, 267–270. [Google Scholar] [CrossRef]
- Dariavach, P.; Mattéi, M.G.; Golstein, P.; Lefranc, M.P. Human Ig superfamily CTLA-4 gene: Chromosomal localization and identity of protein sequence between murine and human CTLA-4 cytoplasmic domains. Eur. J. Immunol. 1988, 18, 1901–1905. [Google Scholar] [CrossRef] [PubMed]
- Howard, T.A.; Rochelle, J.M.; Seldin, M.F. Cd28 and Ctla-4, two related members of the Ig supergene family, are tightly linked on proximal mouse chromosome 1. Immunogenetics 1991, 33, 74–76. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, W.; Huang, Y.; Cui, R.; Li, X.; Li, B. Evolving roles for targeting CTLA-4 in cancer immunotherapy. Cellular physiology and biochemistry. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 47, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S.; Carrega, P.; Saverino, D.; Piccioli, P.; Camoriano, M.; Morabito, A.; Dozin, B.; Fontana, V.; Simone, R.; Mortara, L.; et al. CTLA-4 is expressed by human monocyte-derived dendritic cells and regulates their functions. Hum. Immunol. 2010, 71, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Rozanski, C.H.; Arens, R.; Carlson, L.M.; Nair, J.; Boise, L.H.; Chanan-Khan, A.A.; Schoenberger, S.P.; Lee, K.P. Sustained antibody responses depend on CD28 function in bone marrow-resident plasma cells. J. Exp. Med. 2011, 208, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Dustin, M.L. The immunological synapse. Cancer Immunol. Res. 2014, 2, 1023–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchbinder, E.I.; Desai, A. CTLA-4 and PD-1 Pathways: Similarities, differences, and implications of their inhibition. Am. J. Clin. Oncol. 2016, 39, 98–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhardt, J.J.; Sullivan, T.J.; Allison, J.P. CTLA-4 overexpression inhibits T cell responses through a CD28-B7-dependent mechanism. J. Immunol. 2006, 177, 1052–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krummel, M.F.; Allison, J.P. CD28 and CTLA-4 have opposing effects on the response of T cells to stimulation. J. Exp. Med. 1995, 182, 459–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokosuka, T.; Takamatsu, M.; Kobayashi-Imanishi, W.; Hashimoto-Tane, A.; Azuma, M.; Saito, T. Programmed cell death 1 forms negative costimulatory microclusters that directly inhibit T cell receptor signaling by recruiting phosphatase SHP2. J. Exp. Med. 2012, 209, 1201–1217. [Google Scholar] [CrossRef] [Green Version]
- Cetin, B.; Gumusay, O.; Cengiz, M.; Ozet, A. Advances of molecular targeted therapy in gastric cancer. J. Gastrointest. Cancer 2016, 47, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Lens, M.; Ferrucci, P.F.; Testori, A. Anti-CTLA4 monoclonal antibody Ipilimumab in the treatment of metastatic melanoma: Recent findings. Recent Pat. Anti-Cancer Drug Discov. 2008, 3, 105–113. [Google Scholar] [CrossRef]
- Sakate, Y.; Yashiro, M.; Tanaka, H.; Sunami, T.; Kosaka, K.; Hirakawa, K. Usefulness of inhibiting the lymph node metastasis in human gastric carcinoma by B7–1 gene transfection1. J. Surg. Res. 2004, 122, 89–95. [Google Scholar] [CrossRef]
- Sakate, Y.; Yashiro, M.; Tanaka, H.; Kosaka, K.; Ohira, M.; Hirakawa, K. Suppression of peritoneal metastasis in human gastric carcinoma by enhanced immunogenicity of B7-1 transfection. Oncol. Rep. 2004, 12, 53–57. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, D.; Feng, J.; Zhou, Y.; Wang, Q.; Feng, H.; Zhang, J.; Jiang, J. Overexpression of HHLA2 in human clear cell renal cell carcinoma is significantly associated with poor survival of the patients. Cancer Cell Int. 2019, 19, 101. [Google Scholar] [CrossRef]
- Yang, X.; Lv, T.; Qiang, J.; Li, X.; Zheng, C.; Ding, J.; Lim, Z.; Pang, L.; Du, W.; Zhu, P.; et al. Expression and significance of CD28, CTLA-4, CD80 and CD86 in gastric cancer. Biomed. Res. 2017, 28, 10159–10164. [Google Scholar]
- Yao, S.; Zhu, Y.; Zhu, G.; Augustine, M.; Zheng, L.; Goode, D.J.; Broadwater, M.; Ruff, W.; Flies, S.; Xu, H.; et al. B7-h2 is a costimulatory ligand for CD28 in human. Immunity 2011, 34, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Flies, D. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat. Rev. Immunol. 2013, 13, 542. [Google Scholar] [CrossRef] [Green Version]
- Amatore, F.; Gorvel, L.; Olive, D. Role of Inducible Co-Stimulator (ICOS) in cancer immunotherapy. Expert Opin. Biol. Ther. 2020, 20, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Wikenheiser, D.J.; Stumhofer, J.S. ICOS co-stimulation: Friend or foe? Front. Immunol. 2016, 7, 304. [Google Scholar] [CrossRef] [Green Version]
- Khayyamian, S.; Hutloff, A.; Büchner, K.; Gräfe, M.; Henn, V.; Kroczek, R.A.; Mages, H.W. ICOS-ligand, expressed on human endothelial cells, costimulates Th1 and Th2 cytokine secretion by memory CD4+ T cells. Proc. Natl. Acad. Sci. USA 2002, 99, 6198–6203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, H.; Dan, K.; Tamada, K.; Nakamura, K.; Shioi, Y.; Hyodo, H.; Wang, S.D.; Dong, H.; Chen, L.; Ogata, K. Expression of functional B7-H2 and B7.2 costimulatory molecules and their prognostic implications in de novo acute myeloid leukemia. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2005, 11, 5708–5717. [Google Scholar] [CrossRef] [Green Version]
- Klingenberg, R.; Autschbach, F.; Gleissner, C.; Giese, T.; Wambsganss, N.; Sommer, N.; Richter, G.; Katus, H.A.; Dengler, T.J. Endothelial inducible costimulator ligand expression is increased during human cardiac allograft rejection and regulates endothelial cell-dependent allo-activation of CD8+ T cells in vitro. Eur. J. Immunol. 2005, 35, 1712–1721. [Google Scholar] [CrossRef]
- Schenk, A.D.; Gorbacheva, V.; Rabant, M.; Fairchild, R.L.; Valujskikh, A. Effector functions of donor-reactive CD8 memory T cells are dependent on ICOS induced during division in cardiac grafts. Am. J. Transplant. Off. J. Am. Soc. Transplant. Am. Soc. Transpl. Surg. 2009, 9, 64–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linterman, M.A.; Rigby, R.J.; Wong, R.; Silva, D.; Withers, D.; Anderson, G.; Verma, N.K.; Brink, R.; Hutloff, A.; Goodnow, C.C.; et al. Roquin differentiates the specialized functions of duplicated T cell costimulatory receptor genes CD28 and ICOS. Immunity 2009, 30, 228–241. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.S.; Kageyama, R.; Eto, D.; Escobar, T.C.; Johnston, R.J.; Monticelli, L.; Lao, C.; Crotty, S. ICOS receptor instructs T follicular helper cell versus effector cell differentiation via induction of the transcriptional repressor Bcl6. Immunity 2011, 34, 932–946. [Google Scholar] [CrossRef] [Green Version]
- Scott, G.B.; Carter, C.; Parrish, C.; Wood, P.M.; Cook, G. Downregulation of myeloma-induced ICOS-L and regulatory T cell generation by lenalidomide and dexamethasone therapy. Cell. Immunol. 2015, 297, 1–9. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, G.; Chapoval, A.I.; Dong, H.; Tamada, K.; Ni, J.; Chen, L. Costimulation of T cells by B7-H2, a B7-like molecule that binds ICOS. Blood 2000, 96, 2808–2813. [Google Scholar] [CrossRef]
- Park, H.; Li, Z.; Yang, X.O.; Chang, S.H.; Nurieva, R.; Wang, Y.H.; Wang, Y.; Hood, L.; Zhu, Z.; Tian, Q.; et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat. Immunol. 2005, 6, 1133–1141. [Google Scholar] [CrossRef]
- Dong, C.; Juedes, A.E.; Temann, U.A.; Shresta, S.; Allison, J.P.; Ruddle, N.H.; Flavell, R.A. ICOS co-stimulatory receptor is essential for T-cell activation and function. Nature 2001, 409, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Wallin, J.J.; Liang, L.; Bakardjiev, A.; Sha, W.C. Enhancement of CD8+ T cell responses by ICOS/B7h costimulation. J. Immunol. 2001, 167, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmeister, Y.; Lischke, T.; Dahler, A.C.; Mages, H.W.; Lam, K.P.; Coyle, A.J.; Kroczek, R.A.; Hutloff, A. ICOS controls the pool size of effector-memory and regulatory T cells. J. Immunol. 2008, 180, 774–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solinas, C.; Gu-Trantien, C.; Willard-Gallo, K. The rationale behind targeting the ICOS-ICOS ligand costimulatory pathway in cancer immunotherapy. ESMO Open 2020, 5, e000544. [Google Scholar] [CrossRef] [Green Version]
- Nagase, H.; Takeoka, T.; Urakawa, S.; Morimoto-Okazawa, A.; Kawashima, A.; Iwahori, K.; Takiguchi, S.; Nishikawa, H.; Sato, E.; Sakaguchi, S.; et al. ICOS(+) Foxp3(+) TILs in gastric cancer are prognostic markers and effector regulatory T cells associated with Helicobacter pylori. Int. J. Cancer 2017, 140, 686–695. [Google Scholar] [CrossRef]
- Huang, X.-M.; Liu, X.-S.; Lin, X.-K.; Yu, H.; Sun, J.-Y.; Liu, X.-K.; Chen, C.; Jin, H.-L.; Zhang, G.-E.; Shi, X.-X.; et al. Role of plasmacytoid dendritic cells and inducible costimulator-positive regulatory T cells in the immunosuppression microenvironment of gastric cancer. Cancer Sci. 2014, 105, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, Y.; Nakada, C.; Noguchi, T.; Tanigawa, M.; Nguyen, L.T.; Uchida, T.; Hijiya, N.; Matsuura, K.; Fujioka, T.; Seto, M.; et al. MicroRNA-375 is downregulated in gastric carcinomas and regulates cell survival by targeting PDK1 and 14-3-3zeta. Cancer Res. 2010, 70, 2339–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, P.J.; Song, B.; Mishra, P.J.; Wang, Y.; Humeniuk, R.; Banerjee, D.; Merlino, G.; Ju, J.; Bertino, J.R. MiR-24 tumor suppressor activity is regulated independent of p53 and through a target site polymorphism. PLoS ONE 2009, 4, e8445. [Google Scholar] [CrossRef]
- Yang, P.; Tang, R.; Zhu, J.; Zou, L.; Wu, R.; Zhou, H.; Mao, Y.; Li, R.; Hua, D.; Wang, W.; et al. A functional variant at miR-24 binding site in B7-H2 alters susceptibility to gastric cancer in a Chinese Han population. Mol. Immunol. 2013, 56, 98–103. [Google Scholar] [CrossRef]
- Sharpe, A.H.; Wherry, E.J.; Ahmed, R.; Freeman, G.J. The function of programmed cell death 1 and its ligands in regulating autoimmunity and infection. Nat. Immunol. 2007, 8, 239–245. [Google Scholar] [CrossRef]
- Riella, L.V.; Watanabe, T.; Sage, P.T.; Yang, J.; Yeung, M.; Azzi, J.; Vanguri, V.; Chandraker, A.; Sharpe, A.H.; Sayegh, M.H.; et al. Essential role of PDL1 expression on nonhematopoietic donor cells in acquired tolerance to vascularized cardiac allografts. Am. J. Transplant. Off. J. Am. Soc. Transplant. Am. Soc. Transpl. Surg. 2011, 11, 832–840. [Google Scholar] [CrossRef]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, T.; Honjo, T. PD-1 and PD-1 ligands: From discovery to clinical application. Int. Immunol. 2007, 19, 813–824. [Google Scholar] [CrossRef] [Green Version]
- Greaves, P.; Gribben, J.G. The role of B7 family molecules in hematologic malignancy. Blood 2013, 121, 734–744. [Google Scholar] [CrossRef]
- Neel, B.G.; Gu, H.; Pao, L. The ‘Shp’ing news: SH2 domain-containing tyrosine phosphatases in cell signaling. Trends Biochem. Sci. 2003, 28, 284–293. [Google Scholar] [CrossRef]
- Viricel, C.; Ahmed, M.; Barakat, K. Human PD-1 binds differently to its human ligands: A comprehensive modeling study. J. Mol. Graph. Model. 2015, 57, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Freeman, G.J.; Long, A.J.; Iwai, Y.; Bourque, K.; Chernova, T.; Nishimura, H.; Fitz, L.J.; Malenkovich, N.; Okazaki, T.; Byrne, M.C.; et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J. Exp. Med. 2000, 192, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- Butte, M.J.; Keir, M.E.; Phamduy, T.B.; Sharpe, A.H.; Freeman, G.J. Programmed death-1 ligand 1 interacts specifically with the B7-1 costimulatory molecule to inhibit T cell responses. Immunity 2007, 27, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar] [PubMed]
- Taube, J.M.; Anders, R.A.; Young, G.D.; Xu, H.; Sharma, R.; McMiller, T.L.; Chen, S.; Klein, A.P.; Pardoll, D.M.; Topalian, S.L.; et al. Colocalization of inflammatory response with B7-h1 expression in human melanocytic lesions supports an adaptive resistance mechanism of immune escape. Sci. Transl. Med. 2012, 4, 127ra137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crane, C.A.; Panner, A.; Murray, J.C.; Wilson, S.P.; Xu, H.; Chen, L.; Simko, J.P.; Waldman, F.M.; Pieper, R.O.; Parsa, A.T. PI(3) kinase is associated with a mechanism of immunoresistance in breast and prostate cancer. Oncogene 2009, 28, 306–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Li, Y.; Liu, X.; Cao, S.; Harrington, S.M.; Chen, C.; Mansfiled, A.S.; Dronca, R.S.; Park, S.S.; Yan, Y.; et al. B7-H1(PD-L1) confers chemoresistance through ERK and p38 MAPK pathway in tumor cells. bioRxiv 2018, 308601. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Z.; Latchoumanin, O.; Qiao, L. Antagonizing programmed death-1 and programmed death ligand-1 as a therapeutic approach for gastric cancer. Ther. Adv. Gastroenterol. 2016, 9, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Dong, L.; Wang, K.; Zou, H.; Zhao, S.; Wang, Y.; Wang, G. MiR-21 participates in the PD-1/PD-L1 pathway-mediated imbalance of Th17/Treg cells in patients after gastric cancer resection. Ann. Surg. Oncol. 2019, 26, 884–893. [Google Scholar] [CrossRef]
- Zhang, L.; Qiu, M.; Jin, Y.; Ji, J.; Li, B.; Wang, X.; Yan, S.; Xu, R.; Yang, D. Programmed cell death ligand 1 (PD-L1) expression on gastric cancer and its relationship with clinicopathologic factors. Int. J. Clin. Exp. Pathol. 2015, 8, 11084–11091. [Google Scholar]
- Chakrabarti, J.; Holokai, L.; Syu, L.; Steele, N.G.; Chang, J.; Wang, J.; Ahmed, S.; Dlugosz, A.; Zavros, Y. Hedgehog signaling induces PD-L1 expression and tumor cell proliferation in gastric cancer. Oncotarget 2018, 9, 37439–37457. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Wang, H.; Lu, C.; Li, Q.; Xu, B.; Jiang, J.; Wu, C. Expression of costimulatory molecules B7-H1, B7-H4 and Foxp3+ Tregs in gastric cancer and its clinical significance. Int. J. Clin. Oncol. 2015, 20, 273–281. [Google Scholar] [CrossRef]
- Wang, W.; Sun, J.; Li, F.; Li, R.; Gu, Y.; Liu, C.; Yang, P.; Zhu, M.; Chen, L.; Tian, W.; et al. A frequent somatic mutation in CD274 3′-UTR leads to protein over-expression in gastric cancer by disrupting miR-570 binding. Hum. Mutat. 2012, 33, 480–484. [Google Scholar] [CrossRef]
- Imai, Y.; Chiba, T.; Kondo, T.; Kanzaki, H.; Kanayama, K.; Ao, J.; Kojima, R.; Kusakabe, Y.; Nakamura, M.; Saito, T.; et al. Interferon-γ induced PD-L1 expression and soluble PD-L1 production in gastric cancer. Oncol. Lett. 2020, 20, 2161–2168. [Google Scholar] [CrossRef]
- Oki, E.; Okano, S.; Saeki, H.; Umemoto, Y.; Teraishi, K.; Nakaji, Y.; Ando, K.; Zaitsu, Y.; Yamashita, N.; Sugiyama, M.; et al. Protein expression of programmed death 1 ligand 1 and HER2 in gastric carcinoma. Oncology 2017, 93, 387–394. [Google Scholar] [CrossRef]
- Menyhárt, O.; Pongor, L.S.; Győrffy, B. Mutations defining patient cohorts with elevated PD-L1 expression in gastric cancer. Front. Pharm. 2019, 9, 1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, R.; Zhang, P.; Liu, W.; Zeng, X.; Ma, X.; Shi, L.; Wang, T.; Yin, Y.; Chang, W.; Zhang, P.; et al. HDAC is indispensable for IFN-γ-induced B7-H1 expression in gastric cancer. Clin. Epigenet. 2018, 10, 153. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Wang, H.-Y.; Zhao, X.-X.; Chen, J.-N.; Zhang, Y.-W.; Huang, Y.; Xue, L.; Li, H.-G.; Du, H.; Wu, X.-Y.; et al. Expression and prognostic roles of PIK3CA, JAK2, PD-L1, and PD-L2 in Epstein-Barr virus–associated gastric carcinoma. Hum. Pathol. 2016, 53, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Mimura, K.; Kua, L.-F.; Okayama, H.; Min, A.K.T.; Saito, K.; Hanayama, H.; Watanabe, Y.; Saito, M.; Momma, T.; et al. Immune suppression caused by PD-L2 expression on tumor cells in gastric cancer. Gastric Cancer 2020, 23, 961–973. [Google Scholar] [CrossRef]
- Saito, H.; Kuroda, H.; Matsunaga, T.; Osaki, T.; Ikeguchi, M. Increased PD-1 expression on CD4+ and CD8+ T cells is involved in immune evasion in gastric cancer. J. Surg. Oncol. 2013, 107, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Jung, W.Y.; Kang, Y.; Lee, H.; Kim, A.; Kim, H.K.; Shin, B.K.; Kim, B.-H. Programmed death-ligand 1 expression in gastric adenocarcinoma is a poor prognostic factor in a high CD8+ tumor infiltrating lymphocytes group. Oncotarget 2016, 7, 80426–80434. [Google Scholar] [CrossRef] [Green Version]
- Thompson, E.D.; Zahurak, M.; Murphy, A.; Cornish, T.; Cuka, N.; Abdelfatah, E.; Yang, S.; Duncan, M.; Ahuja, N.; Taube, J.M.; et al. Patterns of PD-L1 expression and CD8 T cell infiltration in gastric adenocarcinomas and associated immune stroma. Gut 2017, 66, 794–801. [Google Scholar] [CrossRef]
- Sun, M.; Richards, S.; Prasad, D.V.; Mai, X.M.; Rudensky, A.; Dong, C. Characterization of mouse and human B7-H3 genes. J. Immunol. 2002, 168, 6294–6297. [Google Scholar] [CrossRef] [PubMed]
- Chapoval, A.I.; Ni, J.; Lau, J.S.; Wilcox, R.A.; Flies, D.B.; Liu, D.; Dong, H.; Sica, G.L.; Zhu, G.; Tamada, K.; et al. B7-H3: A costimulatory molecule for T cell activation and IFN-gamma production. Nat. Immunol. 2001, 2, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Mahnke, K.; Ring, S.; Johnson, T.S.; Schallenberg, S.; Schönfeld, K.; Storn, V.; Bedke, T.; Enk, A.H. Induction of immunosuppressive functions of dendritic cells in vivo by CD4+CD25+ regulatory T cells: Role of B7-H3 expression and antigen presentation. Eur. J. Immunol. 2007, 37, 2117–2126. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Dong, C. New checkpoints in cancer immunotherapy. Immunol. Rev. 2017, 276, 52–65. [Google Scholar] [CrossRef]
- Xu, H.; Cheung, I.Y.; Guo, H.-F.; Cheung, N.-K.V. MicroRNA miR-29 modulates expression of immunoinhibitory molecule B7-H3: Potential implications for immune based therapy of human solid tumors. Cancer Res. 2009, 69, 6275–6281. [Google Scholar] [CrossRef] [Green Version]
- Suh, W.K.; Gajewska, B.U.; Okada, H.; Gronski, M.A.; Bertram, E.M.; Dawicki, W.; Duncan, G.S.; Bukczynski, J.; Plyte, S.; Elia, A.; et al. The B7 family member B7-H3 preferentially down-regulates T helper type 1-mediated immune responses. Nat. Immunol. 2003, 4, 899–906. [Google Scholar] [CrossRef]
- Prasad, D.; Nguyen, T.; Li, Z.; Yang, Y.; Duong, J.; Wang, Y.; Dong, C. Murine B7-H3 is a negative regulator of T cells. J. Immunol. 2004, 173, 2500–2506. [Google Scholar] [CrossRef] [Green Version]
- Hofmeyer, K.A.; Ray, A.; Zang, X. The contrasting role of B7-H3. Proc. Natl. Acad. Sci. USA 2008, 105, 10277–10278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veenstra, R.G.; Flynn, R.; Kreymborg, K.; McDonald-Hyman, C.; Saha, A.; Taylor, P.A.; Osborn, M.J.; Panoskaltsis-Mortari, A.; Schmitt-Graeff, A.; Lieberknecht, E.; et al. B7-H3 expression in donor T cells and host cells negatively regulates acute graft-versus-host disease lethality. Blood 2015, 125, 3335–3346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, W.K.; Wang, S.X.; Jheon, A.H.; Moreno, L.; Yoshinaga, S.K.; Ganss, B.; Sodek, J.; Grynpas, M.D.; Mak, T.W. The immune regulatory protein B7-H3 promotes osteoblast differentiation and bone mineralization. Proc. Natl. Acad. Sci. USA 2004, 101, 12969–12973. [Google Scholar] [CrossRef] [Green Version]
- Castriconi, R.; Dondero, A.; Augugliaro, R.; Cantoni, C.; Carnemolla, B.; Sementa, A.R.; Negri, F.; Conte, R.; Corrias, M.V.; Moretta, L.; et al. Identification of 4Ig-B7-H3 as a neuroblastoma-associated molecule that exerts a protective role from an NK cell-mediated lysis. Proc. Natl. Acad. Sci. USA 2004, 101, 12640–12645. [Google Scholar] [CrossRef] [Green Version]
- Hashiguchi, M.; Kobori, H.; Ritprajak, P.; Kamimura, Y.; Kozono, H.; Azuma, M. From the Cover: Triggering receptor expressed on myeloid cell-like transcript 2 (TLT-2) is a counter-receptor for B7-H3 and enhances T cell responses. Proc. Natl. Acad. Sci. USA 2008, 105, 10495–10500. [Google Scholar] [CrossRef] [Green Version]
- Leitner, J.; Klauser, C.; Pickl, W.F.; Stöckl, J.; Majdic, O.; Bardet, A.F.; Kreil, D.P.; Dong, C.; Yamazaki, T.; Zlabinger, G.; et al. B7-H3 is a potent inhibitor of human T-cell activation: No evidence for B7-H3 and TREML2 interaction. Eur. J. Immunol. 2009, 39, 1754–1764. [Google Scholar] [CrossRef]
- Yan, R.; Yang, S.; Gu, A.; Zhan, F.; He, C.; Qin, C.; Zhang, X.; Feng, P. Murine b7-h3 is a co-stimulatory molecule for T cell activation. Monoclon. Antibodies Immunodiagn. Immunother. 2013, 32, 395–398. [Google Scholar] [CrossRef] [Green Version]
- Vigdorovich, V.; Ramagopal, U.A.; Lázár-Molnár, E.; Sylvestre, E.; Lee, J.S.; Hofmeyer, K.A.; Zang, X.; Nathenson, S.G.; Almo, S.C. Structure and T cell inhibition properties of B7 family member, B7-H3. Structure 2013, 21, 707–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.; Cheng, M.; Zhao, Q.; Goldgur, Y.; Cheal, S.M.; Guo, H.F.; Larson, S.M.; Cheung, N.K. Humanized affinity-matured monoclonal antibody 8H9 has potent antitumor activity and binds to FG loop of tumor antigen B7-H3. J. Biol. Chem. 2015, 290, 30018–30029. [Google Scholar] [CrossRef] [Green Version]
- Loo, D.; Alderson, R.F.; Chen, F.Z.; Huang, L.; Zhang, W.; Gorlatov, S.; Burke, S.; Ciccarone, V.; Li, H.; Yang, Y.; et al. Development of an Fc-enhanced anti-B7-H3 monoclonal antibody with potent antitumor activity. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 3834–3845. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.P.; Jiang, J.T.; Tan, M.; Zhu, Y.B.; Ji, M.; Xu, K.F.; Zhao, J.M.; Zhang, G.B.; Zhang, X.G. Relationship between co-stimulatory molecule B7-H3 expression and gastric carcinoma histology and prognosis. World J. Gastroenterol. 2006, 12, 457–459. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Shen, G.; Qiu, J.; Zhao, X.; Gao, Q. Aberrant expression of B7-H3 in gastric adenocarcinoma promotes cancer cell metastasis. Oncol. Rep. 2014, 32, 2086–2092. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yang, X.; Wu, Y.; Zhao, K.; Ye, Z.; Zhu, J.; Xu, X.; Zhao, X.; Xing, C. B7-H3 promotes gastric cancer cell migration and invasion. Oncotarget 2017, 8, 71725–71735. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.-Y.; Wang, J.-T.; Chen, G.; Shan, Z.-G.; Wang, T.-T.; Shen, Y.; Chen, J.; Yan, Z.-B.; Peng, L.-S.; Mao, F.-Y.; et al. Expression, regulation and clinical significance of B7-H3 on neutrophils in human gastric cancer. Clin. Immunol. 2021, 227, 108753. [Google Scholar] [CrossRef]
- Zhan, S.; Liu, Z.; Zhang, M.; Guo, T.; Quan, Q.; Huang, L.; Guo, L.; Cao, L.; Zhang, X. Overexpression of B7-H3 in α-SMA-positive fibroblasts is associated with cancer progression and survival in gastric adenocarcinomas. Front. Oncol. 2020, 9, 1466. [Google Scholar] [CrossRef]
- Li, Y.; Yang, X.; Yao, P.; Shen, W.; Wu, Y.; Ye, Z.; Zhao, K.; Chen, H.; Cao, J.; Xing, C. B7-H3 increases the radioresistance of gastric cancer cells through regulating baseline levels of cell autophagy. Am. J. Transl. Res. 2019, 11, 4438–4449. [Google Scholar] [PubMed]
- Ulase, D.; Behrens, H.-M.; Krueger, S.; Zeissig, S.; Röcken, C. Gastric carcinomas with stromal B7-H3 expression have lower intratumoural CD8+ T cell density. Int. J. Mol. Sci. 2021, 22, 2129. [Google Scholar] [CrossRef] [PubMed]
- Prasad, D.V.; Richards, S.; Mai, X.M.; Dong, C. B7S1, a novel B7 family member that negatively regulates T cell activation. Immunity 2003, 18, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Sica, G.L.; Choi, I.H.; Zhu, G.; Tamada, K.; Wang, S.D.; Tamura, H.; Chapoval, A.I.; Flies, D.B.; Bajorath, J.; Chen, L. B7-H4, a molecule of the B7 family, negatively regulates T cell immunity. Immunity 2003, 18, 849–861. [Google Scholar] [CrossRef] [Green Version]
- Zang, X.; Loke, P.; Kim, J.; Murphy, K.; Waitz, R.; Allison, J.P. B7x: A widely expressed B7 family member that inhibits T cell activation. Proc. Natl. Acad. Sci. USA 2003, 100, 10388–10392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, I.H.; Zhu, G.; Sica, G.L.; Strome, S.E.; Cheville, J.C.; Lau, J.S.; Zhu, Y.; Flies, D.B.; Tamada, K.; Chen, L. Genomic organization and expression analysis of B7-H4, an immune inhibitory molecule of the B7 family. J. Immunol. 2003, 171, 4650–4654. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, N.; Gavrieli, M.; Sedy, J.R.; Yang, J.; Fallarino, F.; Loftin, S.K.; Hurchla, M.A.; Zimmerman, N.; Sim, J.; Zang, X.; et al. BTLA is a lymphocyte inhibitory receptor with similarities to CTLA-4 and PD-1. Nat. Immunol. 2003, 4, 670–679. [Google Scholar] [CrossRef]
- Sedy, J.R.; Gavrieli, M.; Potter, K.G.; Hurchla, M.A.; Lindsley, R.C.; Hildner, K.; Scheu, S.; Pfeffer, K.; Ware, C.F.; Murphy, T.L.; et al. B and T lymphocyte attenuator regulates T cell activation through interaction with herpesvirus entry mediator. Nat. Immunol. 2005, 6, 90–98. [Google Scholar] [CrossRef]
- Arigami, T.; Uenosono, Y.; Hirata, M.; Hagihara, T.; Yanagita, S.; Ishigami, S.; Natsugoe, S. Expression of B7-H4 in blood of patients with gastric cancer predicts tumor progression and prognosis. J. Surg. Oncol. 2010, 102, 748–752. [Google Scholar] [CrossRef]
- Cao, Q.; Wang, Y.; Zheng, D.; Sun, Y.; Wang, Y.; Lee, V.W.; Zheng, G.; Tan, T.K.; Ince, J.; Alexander, S.I.; et al. IL-10/TGF-beta-modified macrophages induce regulatory T cells and protect against adriamycin nephrosis. J. Am. Soc. Nephrol. JASN 2010, 21, 933–942. [Google Scholar] [CrossRef] [Green Version]
- Kryczek, I.; Zou, L.; Rodriguez, P.; Zhu, G.; Wei, S.; Mottram, P.; Brumlik, M.; Cheng, P.; Curiel, T.; Myers, L.; et al. B7-H4 expression identifies a novel suppressive macrophage population in human ovarian carcinoma. J. Exp. Med. 2006, 203, 871–881. [Google Scholar] [CrossRef]
- Kryczek, I.; Wei, S.; Zou, L.; Zhu, G.; Mottram, P.; Xu, H.; Chen, L.; Zou, W. Cutting edge: Induction of B7-H4 on APCs through IL-10: Novel suppressive mode for regulatory T cells. J. Immunol. 2006, 177, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Rahbar, R.; Lin, A.; Ghazarian, M.; Yau, H.L.; Paramathas, S.; Lang, P.A.; Schildknecht, A.; Elford, A.R.; Garcia-Batres, C.; Martin, B.; et al. B7-H4 expression by nonhematopoietic cells in the tumor microenvironment promotes antitumor immunity. Cancer Immunol. Res. 2015, 3, 184–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangaj, D.; Lanitis, E.; Zhao, A.; Joshi, S.; Cheng, Y.; Sandaltzopoulos, R.; Ra, H.J.; Danet-Desnoyers, G.; Powell, D.J., Jr.; Scholler, N. Novel recombinant human b7-h4 antibodies overcome tumoral immune escape to potentiate T-cell antitumor responses. Cancer Res. 2013, 73, 4820–4829. [Google Scholar] [CrossRef] [Green Version]
- Salceda, S.; Tang, T.; Kmet, M.; Munteanu, A.; Ghosh, M.; Macina, R.; Liu, W.; Pilkington, G.; Papkoff, J. The immunomodulatory protein B7-H4 is overexpressed in breast and ovarian cancers and promotes epithelial cell transformation. Exp. Cell Res. 2005, 306, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ruan, M.; Liu, Y.; Zhu, Y.; Fu, D.; Wu, K.; Zhang, Q. B7H4 expression in tumor cells impairs CD8 T cell responses and tumor immunity. Cancer Immunol. Immunother. 2020, 69, 163–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Zhu, Y.; Wu, C.; Shen, Y.; Wei, W.; Chen, L.; Zheng, X.; Sun, J.; Lu, B.; Zhang, X. Tumor expression of B7-H4 predicts poor survival of patients suffering from gastric cancer. Cancer Immunol. Immunother. 2010, 59, 1707–1714. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.-G.; Yan, Z.-B.; Peng, L.-S.; Cheng, P.; Teng, Y.-S.; Mao, F.-Y.; Fan, K.; Zhuang, Y.; Zhao, Y.-L. Granulocyte-macrophage colony-stimulating factor-activated neutrophils express B7-H4 that correlates with gastric cancer progression and poor patient survival. J. Immunol. Res. 2021, 2021, 6613247. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Li, Z. B7-H4 is predictive of poor prognosis in patients with gastric cancer. Med. Sci. Monit. 2016, 22, 4233–4237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.; Zhou, Y.; Li, C.; Yang, L. Silencing of B7-H4 suppresses the tumorigenicity of the MGC-803 human gastric cancer cell line and promotes cell apoptosis via the mitochondrial signaling pathway. Int. J. Oncol. 2018, 52, 1267–1276. [Google Scholar] [CrossRef] [Green Version]
- Arigami, T.; Uenosono, Y.; Ishigami, S.; Hagihara, T.; Haraguchi, N.; Natsugoe, S. Clinical significance of the B7-H4 coregulatory molecule as a novel prognostic marker in gastric cancer. World J. Surg. 2011, 35, 2051. [Google Scholar] [CrossRef] [PubMed]
- Maskey, N.; Li, K.; Hu, M.; Xu, Z.; Peng, C.; Yu, F.; Cao, H.; Chen, J.; Li, Y.; Yang, G. Impact of neoadjuvant chemotherapy on lymphocytes and co-inhibitory B7-H4 molecule in gastric cancer: Low B7-H4 expression associates with favorable prognosis. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2014, 35, 11837–11843. [Google Scholar] [CrossRef]
- Deng, J.; Le Mercier, I.; Kuta, A.; Noelle, R.J. A New VISTA on combination therapy for negative checkpoint regulator blockade. J. ImmunoTher. Cancer 2016, 4, 86. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhang, X.; Li, E.; Zhang, G.; Wang, X.; Tang, T.; Bai, X.; Liang, T. VISTA: An immune regulatory protein checking tumor and immune cells in cancer immunotherapy. J. Hematol. Oncol. 2020, 13, 83. [Google Scholar] [CrossRef]
- Mehta, N.; Maddineni, S.; Mathews, I.I.; Sperberg, A.P.; Huang, P.-S.; Cochran, J.R. Structure and functional binding epitope of V-domain Ig suppressor of T-cell activation (VISTA). bioRxiv 2019, 597716. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Tekle, C.; Chen, Y.W.; Kristian, A.; Zhao, Y.; Zhou, M.; Liu, Z.; Ding, Y.; Wang, B.; Mælandsmo, G.M.; et al. B7-H3 silencing increases paclitaxel sensitivity by abrogating Jak2/Stat3 phosphorylation. Mol. Cancer Ther. 2011, 10, 960–971. [Google Scholar] [CrossRef] [Green Version]
- Le Mercier, I.; Chen, W.; Lines, J.L.; Day, M.; Li, J.; Sergent, P.; Noelle, R.J.; Wang, L. VISTA Regulates the development of protective antitumor immunity. Cancer Res. 2014, 74, 1933–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borggrewe, M.; Grit, C.; Den Dunnen, W.F.A.; Burm, S.M.; Bajramovic, J.J.; Noelle, R.J.; Eggen, B.J.L.; Laman, J.D. VISTA expression by microglia decreases during inflammation and is differentially regulated in CNS diseases. Glia 2018, 66, 2645–2658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ElTanbouly, M.A.; Croteau, W.; Noelle, R.J.; Lines, J.L. VISTA: A novel immunotherapy target for normalizing innate and adaptive immunity. Semin. Immunol. 2019, 42, 101308. [Google Scholar] [CrossRef]
- Yoon, K.W.; Byun, S.; Kwon, E.; Hwang, S.Y.; Chu, K.; Hiraki, M.; Jo, S.H.; Weins, A.; Hakroush, S.; Cebulla, A.; et al. Control of signaling-mediated clearance of apoptotic cells by the tumor suppressor p53. Science 2015, 349, 1261669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, J.; Li, J.; Sarde, A.; Lines, J.L.; Lee, Y.C.; Qian, D.C.; Pechenick, D.A.; Manivanh, R.; Le Mercier, I.; Lowrey, C.H.; et al. Hypoxia-induced VISTA promotes the suppressive function of myeloid-derived suppressor cells in the tumor microenvironment. Cancer Immunol. Res. 2019, 7, 1079–1090. [Google Scholar] [CrossRef] [Green Version]
- ElTanbouly, M.A.; Schaafsma, E.; Noelle, R.J.; Lines, J.L. VISTA: Coming of age as a multi-lineage immune checkpoint. Clin. Exp. Immunol. 2020, 200, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Nowak, E.C.; Lines, J.L.; Varn, F.S.; Deng, J.; Sarde, A.; Mabaera, R.; Kuta, A.; Le Mercier, I.; Cheng, C.; Noelle, R.J. Immunoregulatory functions of VISTA. Immunol. Rev. 2017, 276, 66–79. [Google Scholar] [CrossRef]
- Flies, D.B.; Han, X.; Higuchi, T.; Zheng, L.; Sun, J.; Ye, J.J.; Chen, L. Coinhibitory receptor PD-1H preferentially suppresses CD4+ T cell-mediated immunity. J. Clin. Investig. 2014, 124, 1966–1975. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wu, G.; Manick, B.; Hernandez, V.; Renelt, M.; Erickson, C.; Guan, J.; Singh, R.; Rollins, S.; Solorz, A.; et al. VSIG-3 as a ligand of VISTA inhibits human T-cell function. Immunology 2019, 156, 74–85. [Google Scholar] [CrossRef] [Green Version]
- Johnston, R.J.; Su, L.J.; Pinckney, J.; Critton, D.; Boyer, E.; Krishnakumar, A.; Corbett, M.; Rankin, A.L.; Dibella, R.; Campbell, L.; et al. VISTA is an acidic pH-selective ligand for PSGL-1. Nature 2019, 574, 565–570. [Google Scholar] [CrossRef]
- Liu, J.; Yuan, Y.; Chen, W.; Putra, J.; Suriawinata, A.A.; Schenk, A.D.; Miller, H.E.; Guleria, I.; Barth, R.J.; Huang, Y.H.; et al. Immune-checkpoint proteins VISTA and PD-1 nonredundantly regulate murine T-cell responses. Proc. Natl. Acad. Sci. USA 2015, 112, 6682–6687. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Xu, Z.; Chen, S.; Lv, H.; Wang, Y.; Wang, X.; Mo, S.; Shi, C.; Wei, S.; Hu, L.; et al. Overexpression of B7H5/CD28H is associated with worse survival in human gastric cancer. J. Cell. Mol. Med. 2020, 24, 1360–1369. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, P.; Carvalho, J.; Rocha, S.; Azevedo, M.; Reis, I.; Camilo, V.; Sousa, B.; Valente, S.; Paredes, J.; Almeida, R.; et al. Dies1/VISTA expression loss is a recurrent event in gastric cancer due to epigenetic regulation. Sci. Rep. 2016, 6, 34860. [Google Scholar] [CrossRef] [Green Version]
- Böger, C.; Behrens, H.-M.; Krüger, S.; Röcken, C. The novel negative checkpoint regulator VISTA is expressed in gastric carcinoma and associated with PD-L1/PD-1: A future perspective for a combined gastric cancer therapy? Oncoimmunology 2017, 6, e1293215. [Google Scholar] [CrossRef]
- Brandt, C.S.; Baratin, M.; Yi, E.C.; Kennedy, J.; Gao, Z.; Fox, B.; Haldeman, B.; Ostrander, C.D.; Kaifu, T.; Chabannon, C.; et al. The B7 family member B7-H6 is a tumor cell ligand for the activating natural killer cell receptor NKp30 in humans. J. Exp. Med. 2009, 206, 1495–1503. [Google Scholar] [CrossRef] [Green Version]
- Kaifu, T.; Escalière, B.; Gastinel, L.N.; Vivier, E.; Baratin, M. B7-H6/NKp30 interaction: A mechanism of alerting NK cells against tumors. Cell. Mol. Life Sci. CMLS 2011, 68, 3531–3539. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Q.; Mariuzza, R.A. Structure of the human activating natural cytotoxicity receptor NKp30 bound to its tumor cell ligand B7-H6. J. Exp. Med. 2011, 208, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Matta, J.; Baratin, M.; Chiche, L.; Forel, J.M.; Cognet, C.; Thomas, G.; Farnarier, C.; Piperoglou, C.; Papazian, L.; Chaussabel, D.; et al. Induction of B7-H6, a ligand for the natural killer cell-activating receptor NKp30, in inflammatory conditions. Blood 2013, 122, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Wang, J.; Ke, X. Knockdown of B7-H6 inhibits tumor progression and enhances chemosensitivity in B-cell non-Hodgkin lymphoma. Int. J. Oncol. 2016, 48, 1561–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, F.; Xie, X.; Wang, L.; Su, Q.; Jia, F.; Ye, Y.; Zang, L.; Wang, J.; Li, H.; Quan, Y.; et al. B7-H6 expression is induced by lipopolysaccharide and facilitates cancer invasion and metastasis in human gliomas. Int. Immunopharmacol. 2018, 59, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zeng, T.; Xiao, Z.; Hu, Q.; Li, Y.; Tan, X.; Yue, H.; Wang, W.; Tan, H.; Zou, J. Immunological role and underlying mechanisms of B7-H6 in tumorigenesis. Clin. Chim. Acta 2020, 502, 191–198. [Google Scholar] [CrossRef]
- Cao, G.; Wang, J.; Zheng, X.; Wei, H.; Tian, Z.; Sun, R. Tumor therapeutics work as stress inducers to enhance tumor sensitivity to natural killer (NK) cell cytolysis by up-regulating NKp30 ligand B7-H6. J. Biol. Chem. 2015, 290, 29964–29973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Narni-Mancinelli, E.; Cantoni, C.; Li, Y.; Guia, S.; Gauthier, L.; Chen, Q.; Moretta, A.; Vély, F.; Eisenstein, E.; et al. Structural insights into the inhibitory mechanism of an antibody against B7-H6, a stress-induced cellular ligand for the natural killer cell receptor NKp30. J. Mol. Biol. 2016, 428, 4457–4466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gacerez, A.T.; Hua, C.K.; Ackerman, M.E.; Sentman, C.L. Chimeric antigen receptors with human scFvs preferentially induce T cell anti-tumor activity against tumors with high B7H6 expression. Cancer Immunol. Immunother. 2018, 67, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-J.; Shen, J.; Zhang, G.-B.; Chen, W.-C. B7-H6 protein expression has no prognostic significance in human gastric carcinoma. Pathol. Oncol. Res. 2014, 20, 203–207. [Google Scholar] [CrossRef]
- Li, D.; Xiang, S.; Shen, J.; Xiao, M.; Zhao, Y.; Wu, X.; Du, F.; Ji, H.; Li, M.; Zhao, Q.; et al. Comprehensive understanding of B7 family in gastric cancer: Expression profile, association with clinicopathological parameters and downstream targets. Int. J. Biol. Sci. 2020, 16, 568–582. [Google Scholar] [CrossRef] [Green Version]
- Janakiram, M.; Chinai, J.M.; Fineberg, S.; Fiser, A.; Montagna, C.; Medavarapu, R.; Castano, E.; Jeon, H.; Ohaegbulam, K.C.; Zhao, R.; et al. Expression, clinical significance, and receptor identification of the newest B7 family member HHLA2 protein. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2015, 21, 2359–2366. [Google Scholar] [CrossRef] [Green Version]
- Mager, D.L.; Hunter, D.G.; Schertzer, M.; Freeman, J.D. Endogenous retroviruses provide the primary polyadenylation signal for two new human genes (HHLA2 and HHLA3). Genomics 1999, 59, 255–263. [Google Scholar] [CrossRef]
- Zhao, R.; Chinai, J.M.; Buhl, S.; Scandiuzzi, L.; Ray, A.; Jeon, H.; Ohaegbulam, K.C.; Ghosh, K.; Zhao, A.; Scharff, M.D.; et al. HHLA2 is a member of the B7 family and inhibits human CD4 and CD8 T-cell function. Proc. Natl. Acad. Sci. USA 2013, 110, 9879–9884. [Google Scholar] [CrossRef] [Green Version]
- Flajnik, M.F.; Tlapakova, T.; Criscitiello, M.F.; Krylov, V.; Ohta, Y. Evolution of the B7 family: Co-evolution of B7H6 and NKp30, identification of a new B7 family member, B7H7, and of B7’s historical relationship with the MHC. Immunogenetics 2012, 64, 571–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Yao, S.; Iliopoulou, B.P.; Han, X.; Augustine, M.M.; Xu, H.; Phennicie, R.T.; Flies, S.J.; Broadwater, M.; Ruff, W.; et al. B7-H5 costimulates human T cells via CD28H. Nat. Commun. 2013, 4, 2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Freeman, G.J. A new B7:CD28 family checkpoint target for cancer immunotherapy: HHLA2. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2015, 21, 2201–2203. [Google Scholar] [CrossRef] [Green Version]
- Janakiram, M.; Chinai, J.M.; Zhao, A.; Sparano, J.A.; Zang, X. HHLA2 and TMIGD2: New immunotherapeutic targets of the B7 and CD28 families. Oncoimmunology 2015, 4, e1026534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimonosono, M.; Arigami, T.; Yanagita, S.; Matsushita, D.; Uchikado, Y.; Kijima, Y.; Kurahara, H.; Kita, Y.; Mori, S.; Sasaki, K.; et al. The association of human endogenous retrovirus-H long terminal repeat-associating protein 2 (HHLA2) expression with gastric cancer prognosis. Oncotarget 2018, 9, 22069–22078. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Tang, L.; Chang, H.; Huo, S.; Li, Y. HHLA2 overexpression is a novel biomarker of malignant status and poor prognosis in gastric cancer. Hum. Cell 2020, 33, 116–122. [Google Scholar] [CrossRef]
- Hecht, I.; Toporik, A.; Podojil, J.R.; Vaknin, I.; Cojocaru, G.; Oren, A.; Aizman, E.; Liang, S.C.; Leung, L.; Dicken, Y.; et al. ILDR2 is a novel B7-like protein that negatively regulates T cell responses. J. Immunol. 2018, 200, 2025–2037. [Google Scholar] [CrossRef] [Green Version]
- Dokmanovic-Chouinard, M.; Chung, W.K.; Chevre, J.-C.; Watson, E.; Yonan, J.; Wiegand, B.; Bromberg, Y.; Wakae, N.; Wright, C.V.; Overton, J.; et al. Positional cloning of “Lisch-like”, a candidate modifier of susceptibility to type 2 diabetes in mice. PLoS Genet. 2008, 4, e1000137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashi, T.; Tokuda, S.; Kitajiri, S.; Masuda, S.; Nakamura, H.; Oda, Y.; Furuse, M. Analysis of the ‘angulin’ proteins LSR, ILDR1 and ILDR2--tricellulin recruitment, epithelial barrier function and implication in deafness pathogenesis. J. Cell Sci. 2013, 126, 966–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podojil, J.R.; Hecht, I.; Chiang, M.Y.; Vaknin, I.; Barbiro, I.; Novik, A.; Neria, E.; Rotman, G.; Miller, S.D. ILDR2-Fc is a novel regulator of immune homeostasis and inducer of antigen-specific immune tolerance. J. Immunol. 2018, 200, 2013–2024. [Google Scholar] [CrossRef] [Green Version]
- Huetter, J.; Gritzan, U.; Gutcher, I.; Doecke, W.-D.; Luetke-Eversloh, M.V.; Golfier, S.; Roider, H.G.; Frisk, A.-L.; Hunter, J.; Pow, A.; et al. Characterization of BAY 1905254, an immune checkpoint inhibitor targeting the immunoglobulin-like domain containing receptor 2 (ILDR2). Cancer Immunol. Res. 2020, 8, 895–911. [Google Scholar] [CrossRef] [Green Version]
- Lipson, E.J.; Drake, C.G. Ipilimumab: An anti-CTLA-4 antibody for metastatic melanoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 6958–6962. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Qin, L.; Ren, M.; Sun, H. Effects of combination of anti-CTLA-4 and anti-PD-1 on gastric cancer cells proliferation, apoptosis and metastasis. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 49, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Ralph, C.; Elkord, E.; Burt, D.J.; O’Dwyer, J.F.; Austin, E.B.; Stern, P.L.; Hawkins, R.E.; Thistlethwaite, F.C. Modulation of lymphocyte regulation for cancer therapy: A phase II trial of tremelimumab in advanced gastric and esophageal adenocarcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 1662–1672. [Google Scholar] [CrossRef] [Green Version]
- Janjigian, Y.Y.; Ott, P.A.; Calvo, E.; Kim, J.W.; Ascierto, P.A.; Sharma, P.; Peltola, K.J.; Jaeger, D.; Evans, T.R.J.; De Braud, F.G.; et al. Nivolumab ± ipilimumab in pts with advanced (adv)/metastatic chemotherapy-refractory (CTx-R) gastric (G), esophageal (E), or gastroesophageal junction (GEJ) cancer: CheckMate 032 study. J. Clin. Oncol. 2017, 35, 4014. [Google Scholar] [CrossRef]
- Muro, K.; Chung, H.C.; Shankaran, V.; Geva, R.; Catenacci, D.; Gupta, S.; Eder, J.P.; Golan, T.; Le, D.T.; Burtness, B.; et al. Pembrolizumab for patients with PD-L1-positive advanced gastric cancer (KEYNOTE-012): A multicentre, open-label, phase 1b trial. Lancet. Oncol. 2016, 17, 717–726. [Google Scholar] [CrossRef]
- Kang, Y.-K.; Boku, N.; Satoh, T.; Ryu, M.-H.; Chao, Y.; Kato, K.; Chung, H.; Chen, J.-S.; Muro, K.; Kang, W.; et al. Nivolumab in patients with advanced gastric or gastro-oesophageal junction cancer refractory to, or intolerant of, at least two previous chemotherapy regimens (ONO-4538-12, ATTRACTION-2): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 390, 2461–2471. [Google Scholar] [CrossRef]
- Tabernero, J.; Van Cutsem, E.; Bang, Y.-J.; Fuchs, C.S.; Wyrwicz, L.; Lee, K.W.; Kudaba, I.; Garrido, M.; Chung, H.C.; Castro Salguero, H.R. Pembrolizumab with or without chemotherapy versus chemotherapy for advanced gastric or gastroesophageal junction (G/GEJ) adenocarcinoma: The phase III KEYNOTE-062 study. J. Clin. Oncol. 2019, 37, LBA4007. [Google Scholar] [CrossRef]
- Chung, H.C.; Arkenau, H.-T.; Lee, J.; Rha, S.Y.; Oh, D.-Y.; Wyrwicz, L.; Kang, Y.-K.; Lee, K.-W.; Bauer, T.M.; Lee, S.S.; et al. Abstract CT111: Avelumab (anti-PD-L1) as first-line maintenance (1L mn) or second-line (2L) therapy in patients with advanced gastric or gastroesophageal junction cancer (GC/GEJC): Updated phase Ib results from the JAVELIN Solid Tumor trial. Cancer Res. 2018, 78, CT111. [Google Scholar] [CrossRef]
- Ohtsu, A.; Tabernero, J.; Bang, Y.-J.; Fuchs, C.S.; Sun, L.; Wang, Z.; Csiki, I.; Koshiji, M.; Van Cutsem, E. Pembrolizumab (MK-3475) versus paclitaxel as second-line therapy for advanced gastric or gastroesophageal junction (GEJ) adenocarcinoma: Phase 3 KEYNOTE-061 study. J. Clin. Oncol. 2016, 34, TPS183. [Google Scholar] [CrossRef]
- Fuchs, C.S.; Doi, T.; Jang, R.W.; Muro, K.; Satoh, T.; Machado, M.; Sun, W.; Jalal, S.I.; Shah, M.A.; Metges, J.-P.; et al. Safety and Efficacy of pembrolizumab monotherapy in patients with previously treated advanced gastric and gastroesophageal junction cancer: Phase 2 clinical KEYNOTE-059 trial. JAMA Oncol. 2018, 4, e180013. [Google Scholar] [CrossRef]
- Bang, Y.J.; Ruiz, E.Y.; Van Cutsem, E.; Lee, K.W.; Wyrwicz, L.; Schenker, M.; Alsina, M.; Ryu, M.H.; Chung, H.C.; Evesque, L.; et al. Phase III, randomised trial of avelumab versus physician’s choice of chemotherapy as third-line treatment of patients with advanced gastric or gastro-oesophageal junction cancer: Primary analysis of JAVELIN Gastric 300. Ann. Oncol. 2018, 29, 2052–2060. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhang, F.; Zhou, N.; Gu, Y.-M.; Zhang, Y.-T.; He, Y.-D.; Wang, L.; Yang, L.-X.; Zhao, Y.; Li, Y.-M. Efficacy and safety of immune checkpoint inhibitors in advanced gastric or gastroesophageal junction cancer: A systematic review and meta-analysis. Oncoimmunology 2019, 8, e1581547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marhelava, K.; Pilch, Z.; Bajor, M.; Graczyk-Jarzynka, A.; Zagozdzon, R. Targeting negative and positive immune checkpoints with monoclonal antibodies in therapy of cancer. Cancers 2019, 11, 1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, C.F.; Proverbs-Singh, T.A.; Postow, M.A. Treatment of the immune-related adverse effects of immune checkpoint inhibitors: A review. JAMA Oncol 2016, 2, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Avelumab in Third-Line Gastric Cancer (JAVELIN Gastric 300). 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT02625623 (accessed on 24 November 2020).
- ClinicalTrials.gov. Avelumab in First-Line Maintenance Gastric Cancer (JAVELIN Gastric 100). 2021. Available online: https://clinicaltrials.gov/ct2/show/NCT02625610 (accessed on 24 June 2021).
- ClinicalTrials.gov. Nivolumab, Ipilimumab and OTSGC-A24 Therapeutic Peptide Vaccine in Gastric Cancer—A Combination Immunotherapy Phase Ib Study (da VINci). 2019. Available online: https://clinicaltrials.gov/ct2/show/NCT03784040 (accessed on 8 April 2019).
- ClinicalTrials.gov. Checkpoint Inhibitor and Radiotherapy for Recurrent Gastric Cancer (CIRCUIT). 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT03453164 (accessed on 15 July 2020).
- ClinicalTrials.gov. Combination of Nivolumab and Ipilimumab in Breast, Ovarian and Gastric Cancer Patients. 2019. Available online: https://clinicaltrials.gov/ct2/show/NCT03342417 (accessed on 31 May 2019).
- ClinicalTrials.govl. Study of Adjuvant ONO-4538 with Resected Gastric Cancer. 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT03006705 (accessed on 22 September 2021).
- ClinicalTrials.gov. Postoperative Immunotherapy vs Standard Chemotherapy for Gastric Cancer with High Risk for Recurrence (VESTIGE). 2021. Available online: https://clinicaltrials.gov/ct2/show/NCT03443856 (accessed on 5 February 2021).
- ClinicalTrials.gov. A Study of Pembrolizumab (MK-3475) in Participants with Recurrent or Metastatic Gastric or Gastroesophageal Junction Adenocarcinoma (MK-3475-059/KEYNOTE-059). 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT02335411 (accessed on 30 July 2021).
- ClinicalTrials.gov. Study of Pembrolizumab (MK-3475) as First-Line Monotherapy and Combination Therapy for Treatment of Advanced Gastric or Gastroesophageal Junction Adenocarcinoma (MK-3475-062/KEYNOTE-062). 2021. Available online: https://clinicaltrials.gov/ct2/show/NCT02494583 (accessed on 10 March 2021).
- ClinicalTrials.gov. A Study of DSP-7888 Dosing Emulsion in Combination with Immune Checkpoint Inhibitors in Adult Patients with Advanced Solid Tumors. 2021. Available online: https://clinicaltrials.gov/ct2/show/NCT03311334 (accessed on 10 September 2021).
- ClinicalTrials.gov. FT500 as Monotherapy and in Combination with Immune Checkpoint Inhibitors in Subjects with Advanced Solid Tumors. 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT03841110 (accessed on 2 November 2020).
- Mazloom, A.; Ghalehsari, N.; Gazivoda, V.; Nimkar, N.; Paul, S.; Gregos, P.; Rateshwar, J.; Khan, U. Role of immune checkpoint inhibitors in gastrointestinal malignancies. J. Clin. Med. 2020, 9, 2533. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
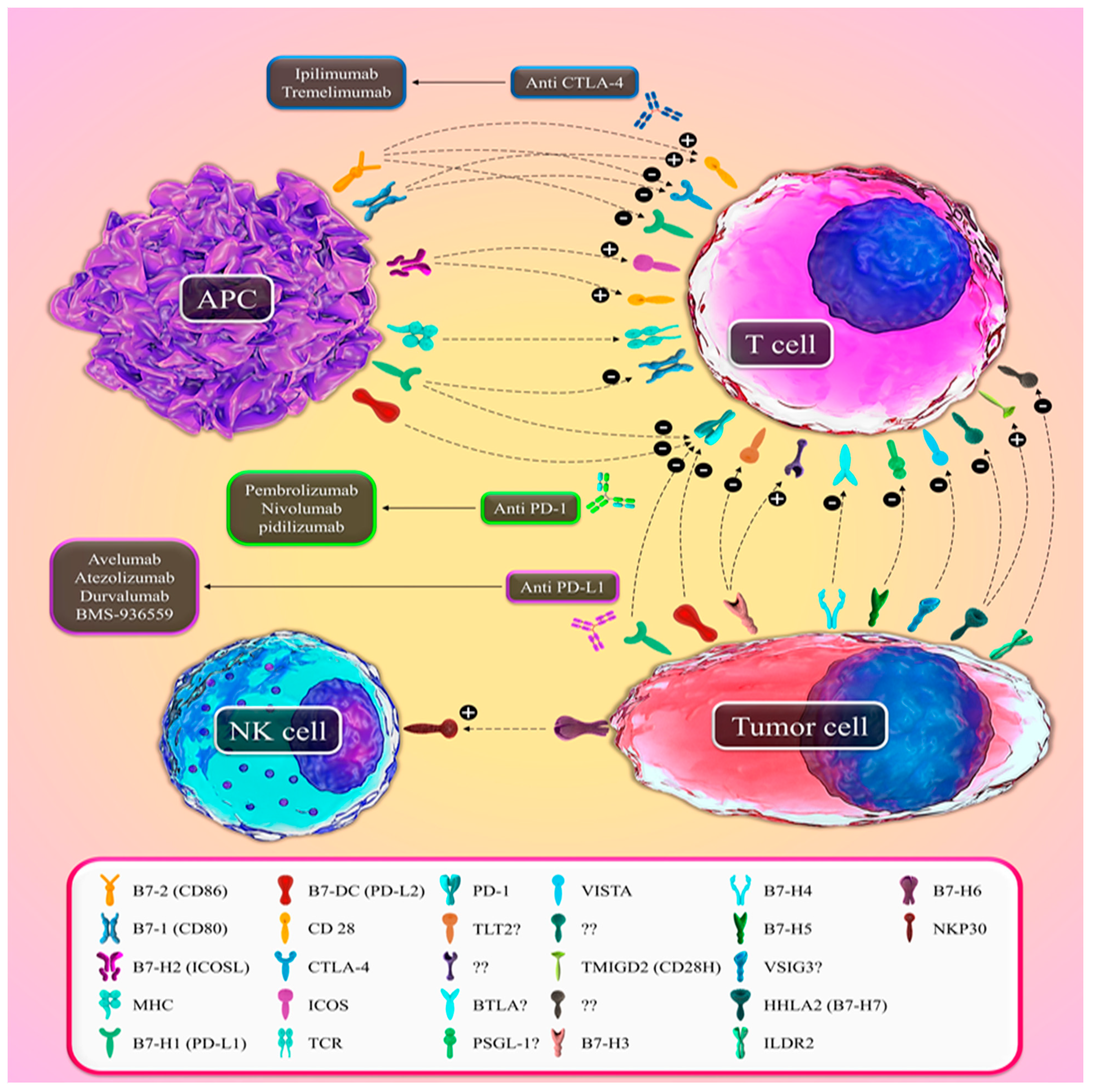
| B7 Family Ligands | Alias Name | Corresponding Receptor(s) | Function |
|---|---|---|---|
| B7-1 | CD80 | CD28/CTLA-4 | Co-stimulatory/Co-inhibitory |
| B7-2 | CD86 | CD28/CTLA-4 | Co-stimulatory/Co-inhibitory |
| B7-H1 | PD-L1/CD274 | PD-1(CD279)/CD80 | Co-inhibitory |
| B7-DC | PD-L2/CD273 | PD-1 (CD279) | Co-inhibitory |
| B7-H2 | ICOSL | ICOS/CD28 | Co-stimulatory |
| B7-H3 | CD276/B7RP-2 | TLT-2 (?) | Co-stimulatory/Co-inhibitory |
| B7-H4 | B7S1/B7x | BTLA (?) | Co-inhibitory |
| B7-H5 | VISTA/PD-1H | PSGL-1 (?) | Co-inhibitory |
| B7-H6 | NCR3LG1 | NKp30 | Co-stimulatory |
| B7-H7 | HHLA2 | CD28H/TMIGD2 | Co-stimulatory/Co-inhibitory |
| ILDR2 | - | (?) | Co-inhibitory |
| Target | Antibody | Combination Treatment | Identifier | Phase | Cancer | Status | Reference |
|---|---|---|---|---|---|---|---|
| Anti-PD-L1 | Avelumab | Irinotecan Paclitaxel | NCT02625623 | III | GC | Completed | [187] |
| Anti-PD-L1 | Avelumab | Oxaliplatin 5-FU Leucovorin Capecitabine | NCT02625610 | III | GC | Active, not recruiting | [188] |
| Anti-PD-1 Anti-CTLA-4 | Nivolumab Ipilimumab | OTSGC-A24 | NCT03784040 | I | GC | Recruiting | [189] |
| Anti-PD-L1 | Nivolumab | Radiotherapy | NCT03453164 | I/II | GC | Active, recruiting | [190] |
| Anti-PD-1 Anti-CTLA-4 | Nivolumab Ipilimumab | - | NCT03342417 | II | GC, Ovarian cancer, Breast Cancer | Terminated (Slow patient accrual) | [191] |
| Anti-PD-L1 | Nivolumab | Tegafur/gimeracil/oteracil Oxaliplatin Capecitabine Placebo | NCT03006705 | III | GC | Recruiting | [192] |
| Anti-PD-1 Anti-CTLA-4 | Nivolumab Ipilimumab | chemotherapy | NCT03443856 | II | GC, gastroesophageal junction adenocarcinoma | Recruiting | [193] |
| Anti-PD-L1 | Pembrolizumab | Cisplatin 5-FU Capecitabine | NCT02335411 | II | GC, gastroesophageal junction adenocarcinoma | Active, not recruiting | [194] |
| Anti-PD-L1 | Pembrolizumab | Cisplatin 5-FU Capecitabine Placebo | NCT02494583 | III | GC | Active, not recruiting | [195] |
| Anti-PD-L1 | Pembrolizumab Nivolumab | DSP-7888 Dosing Emulsion | NCT03311334 | I/II | Melanoma, GC, Colorectal Cancer, etc. | Recruiting | [196] |
| Anti-PD-L1 | Pembrolizumab Nivolumab | FT500 Atezolizumab Cyclophosphamide Fludarabine IL-2 | NCT03841110 | I | GC, Colorectal Cancer, Hepatocellular carcinoma, Small Cell Lung Cancer, Renal Cell Carcinoma, etc. | Recruiting | [197] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolandi, N.; Derakhshani, A.; Hemmat, N.; Baghbanzadeh, A.; Asadzadeh, Z.; Afrashteh Nour, M.; Brunetti, O.; Bernardini, R.; Silvestris, N.; Baradaran, B. The Positive and Negative Immunoregulatory Role of B7 Family: Promising Novel Targets in Gastric Cancer Treatment. Int. J. Mol. Sci. 2021, 22, 10719. https://doi.org/10.3390/ijms221910719
Bolandi N, Derakhshani A, Hemmat N, Baghbanzadeh A, Asadzadeh Z, Afrashteh Nour M, Brunetti O, Bernardini R, Silvestris N, Baradaran B. The Positive and Negative Immunoregulatory Role of B7 Family: Promising Novel Targets in Gastric Cancer Treatment. International Journal of Molecular Sciences. 2021; 22(19):10719. https://doi.org/10.3390/ijms221910719
Chicago/Turabian StyleBolandi, Nadia, Afshin Derakhshani, Nima Hemmat, Amir Baghbanzadeh, Zahra Asadzadeh, Mina Afrashteh Nour, Oronzo Brunetti, Renato Bernardini, Nicola Silvestris, and Behzad Baradaran. 2021. "The Positive and Negative Immunoregulatory Role of B7 Family: Promising Novel Targets in Gastric Cancer Treatment" International Journal of Molecular Sciences 22, no. 19: 10719. https://doi.org/10.3390/ijms221910719
APA StyleBolandi, N., Derakhshani, A., Hemmat, N., Baghbanzadeh, A., Asadzadeh, Z., Afrashteh Nour, M., Brunetti, O., Bernardini, R., Silvestris, N., & Baradaran, B. (2021). The Positive and Negative Immunoregulatory Role of B7 Family: Promising Novel Targets in Gastric Cancer Treatment. International Journal of Molecular Sciences, 22(19), 10719. https://doi.org/10.3390/ijms221910719