Reduced Graphene Oxides Modulate the Expression of Cell Receptors and Voltage-Dependent Ion Channel Genes of Glioblastoma Multiforme

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
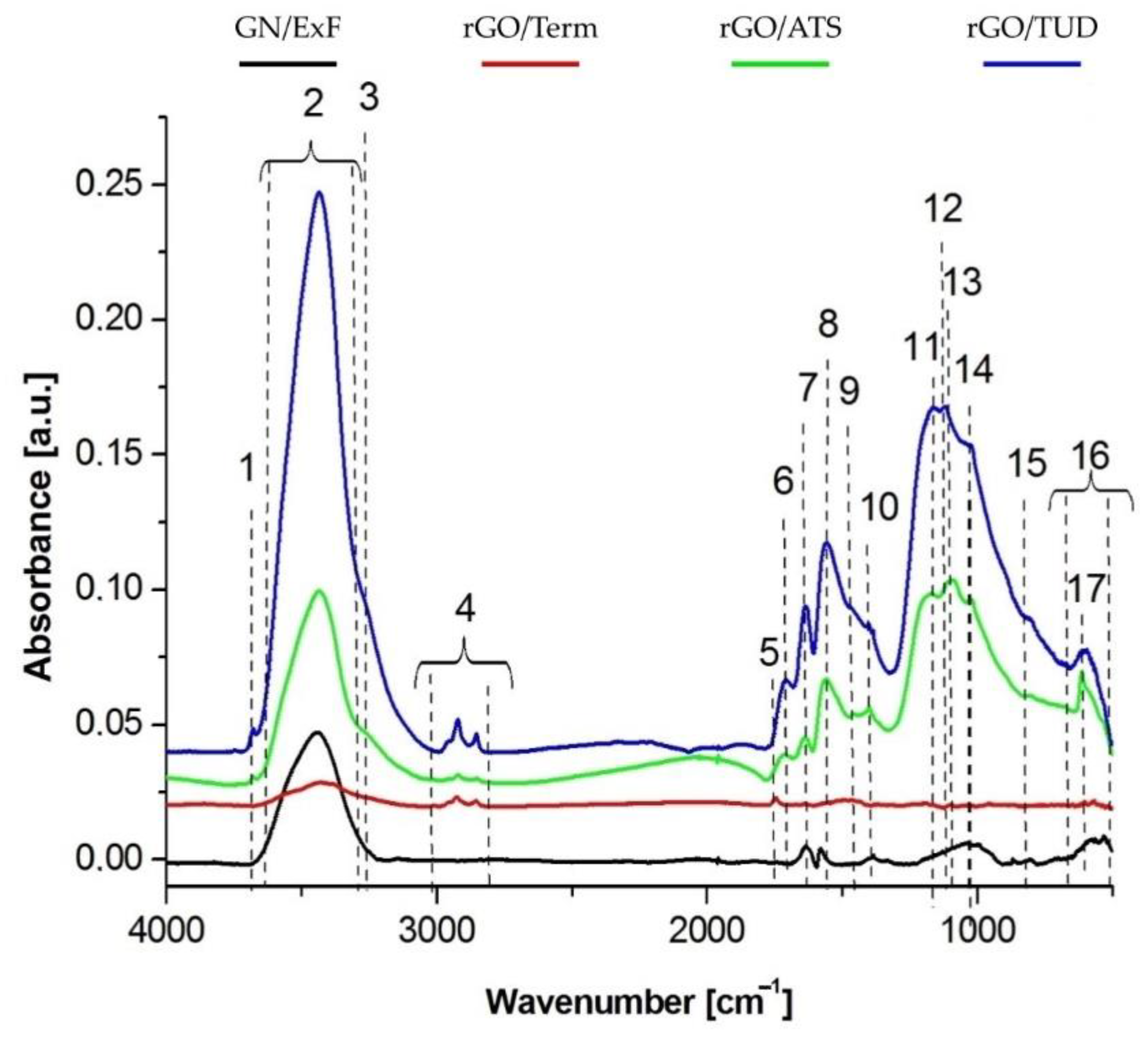
2.1. Physicochemical Characterization of GN and rGO Flakes
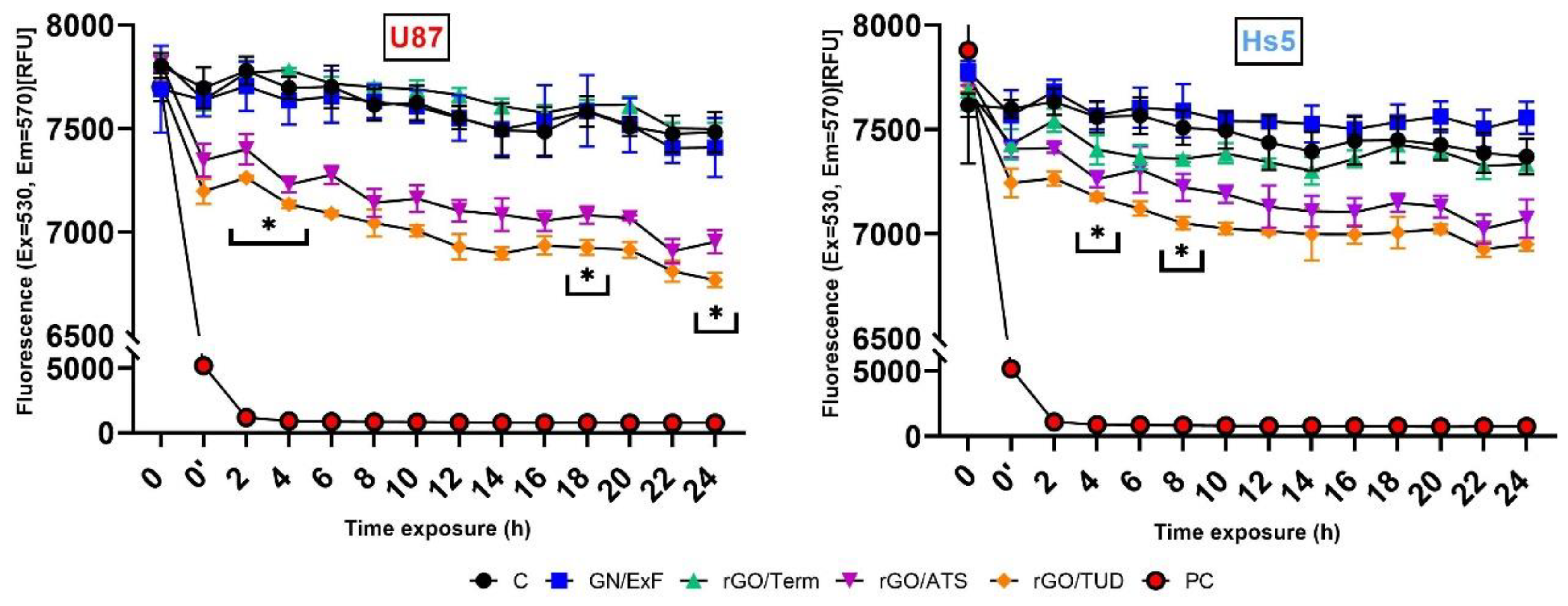
2.2. Membrane Potential
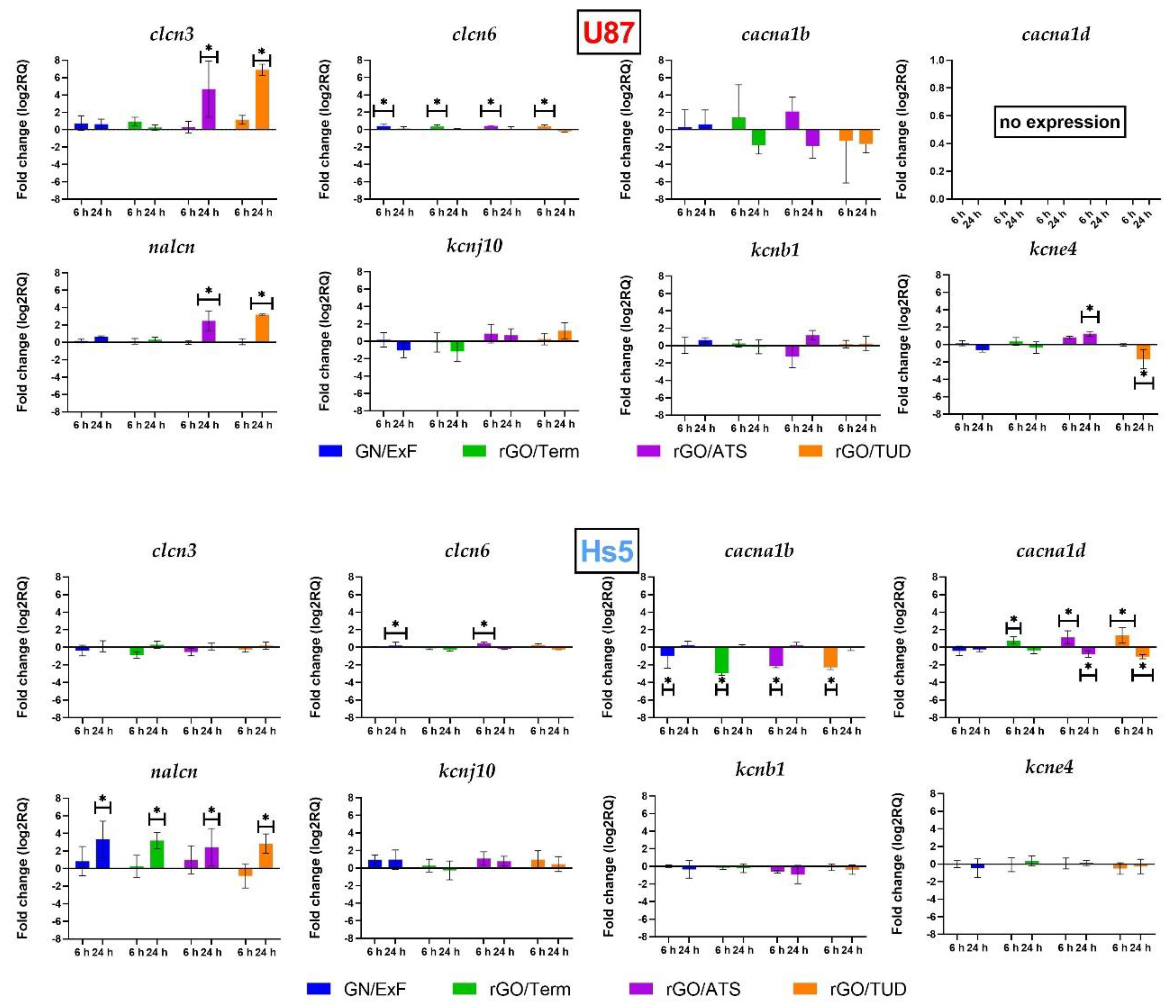
2.3. Gene Expression of Voltage-Gated Ion Channels
2.3.1. Chloride Channels
2.3.2. Calcium Channels
2.3.3. Sodium Channels
2.3.4. Potassium Channels
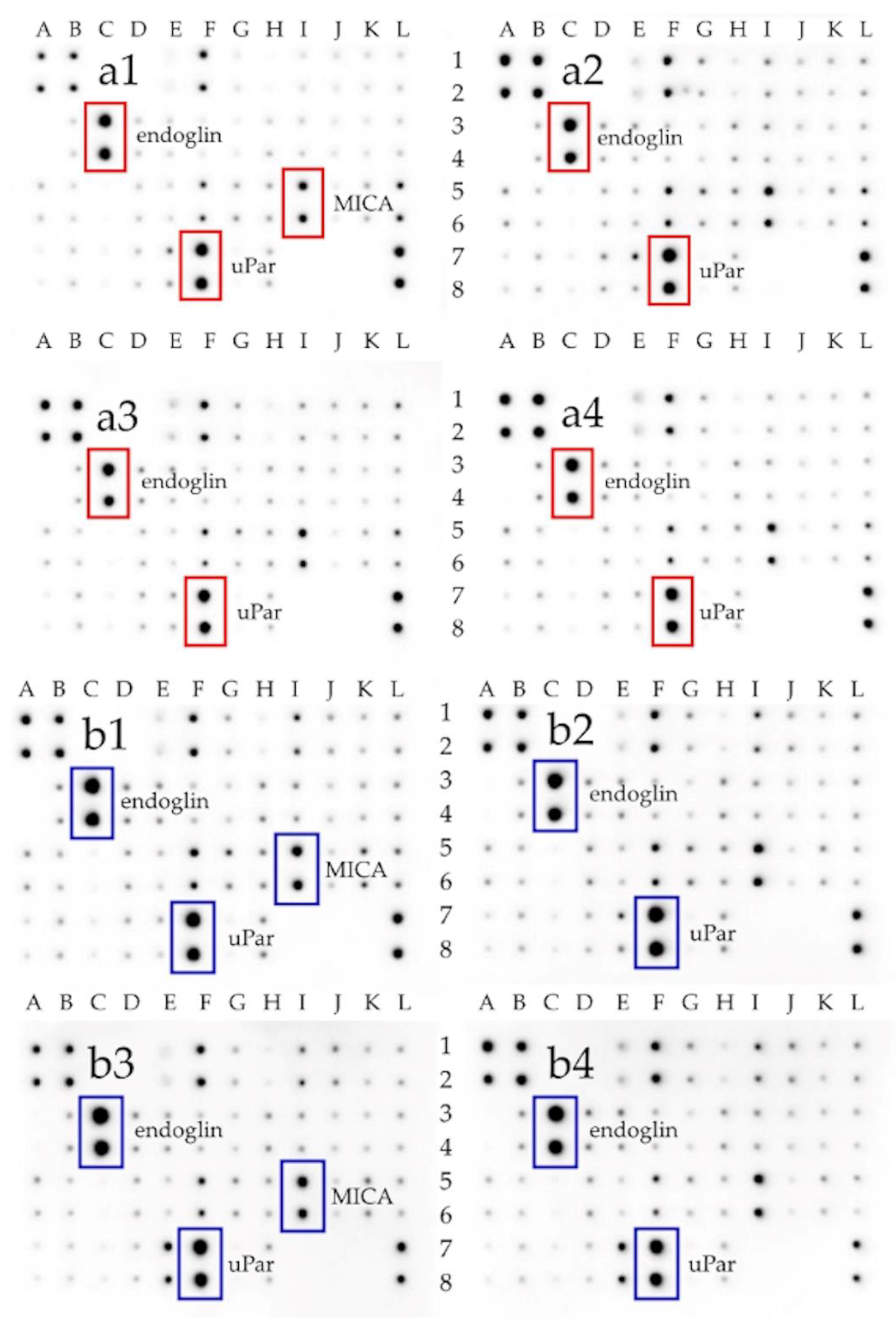
2.4. Expression of Membrane Receptors
3. Discussion
4. Materials and Methods
4.1. Production and Preparation of GN and rGO
4.2. Characterization of GN and rGOs
4.2.1. Fourier Transform IR (FT-IR) Spectrometer Analysis
4.2.2. Scanning Electron Microscopy of Flakes
4.3. Cell Cultures
4.4. Cell Membrane Potential Assay
4.5. Isolation of Total RNA and cDNA Synthesis
4.6. Gene Expression
4.7. Human Receptor Antibody Array
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wen, P.Y.; Reardon, D.A. Neuro-oncology in 2015: Progress in glioma diagnosis, classification and treatment. Nat. Rev. Neurol. 2016, 12, 69–70. [Google Scholar] [CrossRef]
- Zhang, Y.; Cruickshanks, N.; Yuan, F.; Wang, B.; Pahuski, M.; Wulfkuhle, J.; Gallagher, I.; Koeppel, A.F.; Hatef, S.; Papanicolas, C.; et al. Targetable T-type Calcium Channels Drive Glioblastoma. Cancer Res. 2017, 77, 3479–3490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.-H.; Jang, C.; Xiao, S.; Ishigami, M.; Fuhrer, M.S. Intrinsic and extrinsic performance limits of graphene devices on SiO2. Nat. Nanotechnol. 2008, 3, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Lahiri, I.; Seelaboyina, R.; Kang, Y.S. Synthesis of Graphene and Its Applications: A Review. Crit. Rev. Solid State Mater. Sci. 2010, 35, 52–71. [Google Scholar] [CrossRef]
- Geim, A.K.; Novoselov, K.S. The rise of graphene. Nat. Mater. 2007, 6, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Compton, O.C.; Nguyen, S.T. Graphene Oxide, Highly Reduced Graphene Oxide, and Graphene: Versatile Building Blocks for Carbon-Based Materials. Small 2010, 6, 711–723. [Google Scholar] [CrossRef]
- Acik, M.; Mattevi, C.; Gong, C.; Lee, G.; Cho, K.; Chhowalla, M.; Chabal, Y.J. The Role of Intercalated Water in Multilayered Graphene Oxide. ACS Nano 2010, 4, 5861–5868. [Google Scholar] [CrossRef]
- Gao, W. The chemistry of graphene oxide. In Graphene Oxide: Reduction Recipes, Spectroscopy, and Applications; Springer International Publishing: Cham, Switzerland, 2015; ISBN 9783319155005. [Google Scholar]
- Grodzik, M.; Sawosz, E.; Wierzbicki, M.; Orlowski, P.; Hotowy, A.; Niemiec, T.; Szmidt, M.; Mitura, K.; Chwalibog, A. Na-noparticles of carbon allotropes inhibit glioblastoma multiforme angiogenesis in ovo. Int. J. Nanomed. 2011, 6, 3041–3048. [Google Scholar]
- Wierzbicki, M.; Sawosz, E.; Grodzik, M.; Kutwin, M.; Jaworski, S.; Chwalibog, A. Comparison of anti-angiogenic properties of pristine carbon nanoparticles. Nanoscale Res. Lett. 2013, 8, 195. [Google Scholar] [CrossRef] [Green Version]
- Chwalibog, A.; Jaworski, S.; Sawosz, E.; Grodzik, M.; Winnicka, A.; Prasek, M.; Wierzbicki, M. In vitro evaluation of the effects of graphene platelets on glioblastoma multiforme cells. Int. J. Nanomed. 2013, 8, 413–420. [Google Scholar] [CrossRef] [Green Version]
- Jaworski, S.; Sawosz, E.; Kutwin, M.; Wierzbicki, M.; Hinzmann, M.; Grodzik, M.; Winnicka, A.; Lipińska, L.; Wlodyga, K.; Chwalibog, A. In vitro and in vivo effects of graphene oxide and reduced graphene oxide on glioblastoma. Int. J. Nanomed. 2015, 10, 1585–1596. [Google Scholar] [CrossRef] [Green Version]
- Jaworski, S.; Strojny, B.; Sawosz, E.; Wierzbicki, M.; Grodzik, M.; Kutwin, M.; Daniluk, K.; Chwalibog, A. Degradation of Mitochondria and Oxidative Stress as the Main Mechanism of Toxicity of Pristine Graphene on U87 Glioblastoma Cells and Tumors and HS-5 Cells. Int. J. Mol. Sci. 2019, 20, 650. [Google Scholar] [CrossRef] [Green Version]
- Szczepaniak, J.; Strojny, B.; Sawosz, E.; Jaworski, S.; Jagiello, J.; Winkowska, M.; Szmidt, M.; Wierzbicki, M.; Sosnowska, M.; Balaban, J.; et al. Effects of Reduced Graphene Oxides on Apoptosis and Cell Cycle of Glioblastoma Multiforme. Int. J. Mol. Sci. 2018, 19, 3939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, C.; King, A.; Simon, T.; Giamas, G. Graphene-Induced Transdifferentiation of Cancer Stem Cells as a Therapeutic Strategy against Glioblastoma. ACS Biomater. Sci. Eng. 2020, 6, 3258–3269. [Google Scholar] [CrossRef]
- Bondar, O.V.; Saifullina, D.V.; Shakhmaeva, I.I.; Mavlyutova, I.I.; Abdullin, T.I. Monitoring of the Zeta Potential of Human Cells upon Reduction in Their Viability and Interaction with Polymers. Acta Nat. 2012, 4, 78–81. [Google Scholar] [CrossRef]
- Marmo, A.A.; Morris, D.M.; Schwalke, M.A.; Iliev, I.G.; Rogers, S. Electrical Potential Measurements in Human Breast Cancer and Benign Lesions. Tumor Biol. 1994, 15, 147–152. [Google Scholar] [CrossRef]
- Fiorillo, M.; Verre, A.F.; Iliut, M.; Peiris-Pagés, M.; Ozsvari, B.; Gandara, R.; Cappello, A.R.; Sotgia, F.; Vijayaraghavan, A.; Lisanti, M.P. Graphene oxide selectively targets cancer stem cells, across multiple tumor types: Implications for non-toxic cancer treatment, via “differentiation-based nano-therapy”. Oncotarget 2015, 6, 3553–3562. [Google Scholar] [CrossRef] [Green Version]
- Pollak, J.; Rai, K.G.; Funk, C.C.; Arora, S.; Lee, E.; Zhu, J.; Price, N.D.; Paddison, P.J.; Ramirez, J.-M.; Rostomily, R.C. Ion channel expression patterns in glioblastoma stem cells with functional and therapeutic implications for malignancy. PLoS ONE 2017, 12, e0172884. [Google Scholar] [CrossRef]
- Simon, O.J.; Müntefering, T.; Grauer, O.; Meuth, S.G. The role of ion channels in malignant brain tumors. J. Neuro-Oncol. 2015, 125, 225–235. [Google Scholar] [CrossRef]
- Parsons, D.W.; Jones, S.; Zhang, X.; Lin, J.C.-H.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Siu, I.-M.; Gallia, G.L.; et al. An Integrated Genomic Analysis of Human Glioblastoma Multiforme. Science 2008, 321, 1807–1812. [Google Scholar] [CrossRef] [Green Version]
- Joshi, A.D.; Parsons, D.W.; Velculescu, V.E.; Riggins, G.J. Sodium ion channel mutations in glioblastoma patients correlate with shorter survival. Mol. Cancer 2011, 10, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuddapah, V.A.; Robel, S.; Watkins, S.; Sontheimer, H. A neurocentric perspective on glioma invasion. Nat. Rev. Neurosci. 2014, 15, 455–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, A.K.; Liu, X.; Sontheimer, H. Role for calcium-activated potassium channels (BK) in growth control of human malignant glioma cells. J. Neurosci. Res. 2004, 78, 224–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loryuenyong, V.; Totepvimarn, K.; Eimburanapravat, P.; Boonchompoo, W.; Buasri, A. Preparation and Characterization of Reduced Graphene Oxide Sheets via Water-Based Exfoliation and Reduction Methods. Adv. Mater. Sci. Eng. 2013, 2013, 923403. [Google Scholar] [CrossRef] [Green Version]
- Emiru, T.F.; Ayele, D.W. Controlled synthesis, characterization and reduction of graphene oxide: A convenient method for large scale production. Egypt. J. Basic Appl. Sci. 2017, 4, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.; Zhang, Y.; Shan, C.; Zhang, X.; Gao, G.; Pan, B. Flat Graphene-Enhanced Electron Transfer Involved in Redox Reactions. Environ. Sci. Technol. 2017, 51, 8597–8605. [Google Scholar] [CrossRef]
- Cone, C.D. Unified theory on the basic mechanism of normal mitotic control and oncogenesis. J. Theor. Biol. 1971, 30, 151–181. [Google Scholar] [CrossRef]
- Tokuoka, S.; Morioka, H. The membrane potential of the human cancer and related cells. I. Gan 1957, 48, 353–354. [Google Scholar] [CrossRef]
- Johnstone, B.M. Micro-Electrode Penetration of Ascites Tumour Cells. Nature 1959, 183, 411. [Google Scholar] [CrossRef]
- Yang, M.; Brackenbury, W.J. Membrane potential and cancer progression. Front. Physiol. 2013, 4, 185. [Google Scholar] [CrossRef] [Green Version]
- Molenaar, R.J. Ion Channels in Glioblastoma. ISRN Neurol. 2011, 2011, 590249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001; ISBN 0878933212. [Google Scholar]
- Wang, R.; Gurguis, C.I.; Gu, W.; Ko, E.A.; Lim, I.; Bang, H.; Zhou, T.; Ko, J.-H. Ion channel gene expression predicts survival in glioma patients. Sci. Rep. 2015, 5, 11593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sontheimer, H. An Unexpected Role for Ion Channels in Brain Tumor Metastasis. Exp. Biol. Med. 2008, 233, 779–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, N.N.; Wang, C.-Y.; Chen, C.-F.; Sun, Z.; Lai, M.-D.; Lin, Y.-C. Voltage-gated calcium channels: Novel targets for cancer therapy. Oncol. Lett. 2017, 14, 2059–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solé, L.; Roura-Ferrer, M.; Pérez-Verdaguer, M.; Oliveras, A.; Calvo, M.; Fernández-Fernández, J.M.; Felipe, A. KCNE4 suppresses Kv1.3 currents by modulating trafficking, surface expression and channel gating. J. Cell Sci. 2009, 122, 3738–3748. [Google Scholar] [CrossRef] [Green Version]
- Ouwerkerk, R.; Jacobs, M.A.; Macura, K.J.; Wolff, A.C.; Stearns, V.; Mezban, S.D.; Khouri, N.F.; Bluemke, D.A.; Bottomley, P.A. Elevated tissue sodium concentration in malignant breast lesions detected with non-invasive 23Na MRI. Breast Cancer Res. Treat. 2007, 106, 151–160. [Google Scholar] [CrossRef]
- Cahoy, J.D.; Emery, B.; Kaushal, A.; Foo, L.C.; Zamanian, J.L.; Christopherson, K.S.; Xing, Y.; Lubischer, J.L.; Krieg, P.A.; Krupenko, S.A.; et al. A Transcriptome Database for Astrocytes, Neurons, and Oligodendrocytes: A New Resource for Understanding Brain Development and Function. J. Neurosci. 2008, 28, 264–278. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Segura, E.; Cañizares, F.J.; Cubero, M.A.; Warley, A.; Campos, A. Changes in Elemental Content During Apoptotic Cell Death Studied by Electron Probe X-Ray Microanalysis. Exp. Cell Res. 1999, 253, 454–462. [Google Scholar] [CrossRef] [Green Version]
- Skepper, J.N.; Karydis, I.; Garnett, M.R.; Hegyi, L.; Hardwick, S.J.; Warley, A.; Mitchinson, M.J.; Cary, N.R.B. Changes in elemental concentrations are associated with early stages of apoptosis in human monocyte-macrophages exposed to oxidized low-density lipoprotein: An X-ray microanalytical study. J. Pathol. 1999, 188, 100–106. [Google Scholar] [CrossRef]
- Arrebola, F.; Fernández-Segura, E.; Campos, A.; Crespo, P.V.; Skepper, J.N.; Warley, A. Changes in intracellular electrolyte concentrations during apoptosis induced by UV irradiation of human myeloblastic cells. Am. J. Physiol. Cell Physiol. 2006, 290, C638–C649. [Google Scholar] [CrossRef] [Green Version]
- Arrebola, F.; Zabiti, S.; Cañizares, F.J.; Cubero, M.A.; Crespo, P.V.; Fernández-Segura, E. Changes in intracellular sodium, chlorine, and potassium concentrations in staurosporine-induced apoptosis. J. Cell. Physiol. 2005, 204, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Poët, M.; Kornak, U.; Schweizer, M.; Zdebik, A.A.; Scheel, O.; Hoelter, S.; Wurst, W.; Schmitt, A.; Fuhrmann, J.C.; Planells-Cases, R.; et al. Lysosomal storage disease upon disruption of the neuronal chloride transport protein ClC-6. Proc. Natl. Acad. Sci. USA 2006, 103, 13854–13859. [Google Scholar] [CrossRef] [Green Version]
- Neagoe, I.; Stauber, T.; Fidzinski, P.; Bergsdorf, E.-Y.; Jentsch, T.J. The Late Endosomal ClC-6 Mediates Proton/Chloride Countertransport in Heterologous Plasma Membrane Expression. J. Biol. Chem. 2010, 285, 21689–21697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, B.A.; Chimenti, M.S.; Jacobson, M.P.; Barber, D.L. Dysregulated pH: A perfect storm for cancer progression. Nat. Rev. Cancer 2011, 11, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Damaghi, M.; Wojtkowiak, J.W.; Gillies, R.J. pH sensing and regulation in cancer. Front. Physiol. 2013, 4, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuyama, S.; Reed, J.C. Mitochondria-dependent apoptosis and cellular pH regulation. Cell Death Differ. 2000, 7, 1155–1165. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Qu, C.; Han, C.; Chen, M.-M.; An, L.-J.; Zou, W. Potassium channels and their role in glioma: A mini review. Mol. Membr. Biol. 2019, 35, 76–85. [Google Scholar] [CrossRef]
- Du, J.; Haak, L.L.; Phillips-Tansey, E.; Russell, J.T.; McBain, C.J. Frequency-dependent regulation of rat hippocampal somato-dendritic excitability by the K+ channel subunit Kv2.1. J. Physiol. 2000, 522, 19–31. [Google Scholar] [CrossRef]
- Biasiotta, A.; D’Arcangelo, D.; Passarelli, F.; Nicodemi, E.M.; Facchiano, A. Ion channels expression and function are strongly modified in solid tumors and vascular malformations. J. Transl. Med. 2016, 14, 285. [Google Scholar] [CrossRef] [Green Version]
- McCrossan, Z.A.; Abbott, G.W. The MinK-related peptides. Neuropharmacology 2004, 47, 787–821. [Google Scholar] [CrossRef]
- Li, Y.; Sung, Y.U.; McDonald, T.V. Voltage-Gated Potassium Channels: Regulation by Accessory Subunits. Neuroscientist 2006, 12, 199–210. [Google Scholar] [CrossRef]
- Kanda, V.A.; Abbott, G.W. KCNE Regulation of K+ Channel Trafficking—A Sisyphean Task? Front. Physiol. 2012, 3, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.-Y.; Lai, M.-D.; Phan, N.N.; Sun, Z.; Lin, Y.-C. Meta-Analysis of Public Microarray Datasets Reveals Voltage-Gated Calcium Gene Signatures in Clinical Cancer Patients. PLoS ONE 2015, 10, e0125766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.; Bi, M.; Wang, L.; Kang, Z.; Ling, L.; Zhao, C. CLC-3 channels in cancer (Review). Oncol. Rep. 2015, 33, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lui, V.C.H.; Lung, S.S.S.; Pu, J.K.S.; Hung, K.N.; Leung, G.K.K. Invasion of human glioma cells is regulated by multiple chlo-ride channels including ClC-3. Anticancer Res. 2010, 30, 4515–4524. [Google Scholar]
- Montuori, N.; Cosimato, V.; Rinaldi, L.; Rea, V.E.A.; Alfano, D.; Ragno, P. uPAR regulates pericellular proteolysis through a mechanism involving integrins and fMLF-receptors. Thromb. Haemost. 2013, 109, 309–318. [Google Scholar] [CrossRef]
- Raghu, H.; Lakka, S.S.; Gondi, C.S.; Mohanam, S.; Dinh, D.H.; Gujrati, M.; Rao, J.S. Suppression of uPA and uPAR Attenuates Angiogenin Mediated Angiogenesis in Endothelial and Glioblastoma Cell Lines. PLoS ONE 2010, 5, e12458. [Google Scholar] [CrossRef] [Green Version]
- Huai, Q.; Zhou, A.; Lin, L.; Mazar, A.P.; Parry, G.C.; Callahan, J.; Shaw, D.E.; Furie, B.; Furie, B.C.; Huang, M. Crystal structures of two human vitronectin, urokinase and urokinase receptor complexes. Nat. Struct. Mol. Biol. 2008, 15, 422–423. [Google Scholar] [CrossRef] [Green Version]
- Høyer-Hansen, G.; Rønne, E.; Solberg, H.; Behrendt, N.; Ploug, M.; Lund, L.R.; Ellis, V.; Danø, K. Urokinase plasminogen activator cleaves its cell surface receptor releasing the ligand-binding domain. J. Biol. Chem. 1992, 267, 18224–18229. [Google Scholar]
- Høyer-Hansen, G.; Behrendt, N.; Ploug, M.; Danø, K.; Preissner, K.T. The intact urokinase receptor is required for efficient vitronectin binding: Receptor cleavage prevents ligand interaction. FEBS Lett. 1997, 420, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.J.; Tilly, H.; Ward, J.H.; MacArthur, D.C.; Lowe, J.; Coyle, B.; Grundy, R.G. CD105 (Endoglin) exerts prognostic effects via its role in the microvascular niche of paediatric high grade glioma. Acta Neuropathol. 2012, 124, 99–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birlik, B.; Canda, S.; Ozer, E. Tumour vascularity is of prognostic significance in adult, but not paediatric astrocytomas. Neuropathol. Appl. Neurobiol. 2006, 32, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Barresi, V.; Cerasoli, S.; Vitarelli, E.; Tuccari, G. Density of microvessels positive for CD105 (endoglin) is related to prognosis in meningiomas. Acta Neuropathol. 2007, 114, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Muenzner, P.; Rohde, M.; Kneitz, S.; Hauck, C.R. CEACAM engagement by human pathogens enhances cell adhesion and counteracts bacteria-induced detachment of epithelial cells. J. Cell Biol. 2005, 170, 825–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aithal, M.G.S.; Rajeswari, N. Validation of Housekeeping Genes for Gene Expression Analysis in Glioblastoma Using Quantitative Real-Time Polymerase Chain Reaction. Brain Tumor Res. Treat. 2015, 3, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-Y.; Wang, W.; Liu, Y.-W.; Li, M.-Y.; Liang, T.-Y.; Li, J.-Y.; Hu, H.-M.; Lu, Y.; Yao, C.; Ye, Y.-Y.; et al. Role of KCNB1 in the prognosis of gliomas and autophagy modulation. Sci. Rep. 2017, 7, 14. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Wavenumber [cm−1] | Type of Bond | Sample | |||
|---|---|---|---|---|---|---|
| GN/ExF | rGO/Term | rGO/ATS | rGO/TUD | |||
| 1 | 3680 | O-H (stretch) | - | - | - | + (w) |
| 2 | 3600–3000 | O-H (stretch) | + (m) | + (w) | + (m) | + (s) |
| 3 | 3280 | N-H (stretch) | - | - | + (m) | + (m) |
| 4 | 3000–2800 | C-H (stretch) | - | + (m) | + (m) | + (s) |
| 5 | 1747 | C=O (stretch) | - | + (w) | - | - |
| 6 | 1711 | C=O (stretch) | - | - | + (m) | + (s) |
| 7 | 1640 | C=C (stretch) | + (m) | - | + (m) | + (s) |
| 8 | 1563 | COO- (stretch) | + (w) | - | + (m) | + (s) |
| 9 | 1465 | C-H (deformation) | - | + (w) | + (w) | + (m) |
| 10 | 1400 | -OH (in-plane bendig) | + (w) | + (w) | + (w) | + (s) |
| 11 | 1174 | C-O-C (stretch) | + (w) | - | + (m) | + (s) |
| 12 | 1124 | C-NH2/C-N (stretch) | - | - | + (m) | + (w) |
| 13 | 1100 | C=S (stretch) | - | - | + (w) | + (w) |
| 14 | 1018 | Carbon ring | - | + (w) | +(m) | +(m) |
| 15 | 800 | CH2=CH (deformation) | - | - | + (w) | + (m) |
| 16 | 659–511 | carbon ring | + (w) | + (m) | + (m) | + (m) |
| 17 | 617 | C-S (stretch)/S-O (scissors) | - | - | + (s) | + (m) |
| Genes | Forward Primers (5′-3′) | Reverse Primers (5′-3′) | References |
|---|---|---|---|
| clcn3 | CTGTGCCGCCTCTAAGCC | ACTGTAGTTCGACTCGCTGAA | Primer blast |
| clcn6 | ACCTGGAAGTTTTGGAGACCAT | TGAGTTGGGTGAAGAGTCGC | Primer blast |
| cacna1b | CCCTTGCTGTCAACATCTGGT | GGATGGGTGAGGAGTTGGC | Primer blast |
| cacna1d | ACTCGGGCTATCCAGAAGTAG | CTTGCCCAAAGAAAAGACTGC | Primer blast |
| nalcn | CGCCGTAGACTGTGGTTTTG | AATGACGCTGATGATGGCAC | Primer blast |
| kcnj10 | TCAGAAGACGGGCGAAACAA | GCGAGCCTAAGCAAGACTCA | Primer blast |
| kcnb1 | CCATTCTGCCATACTATGTCACC | AGCAAGCCCAACTCATTGTAG | [68] |
| kcne4 | CACCGCTACCTGAAAACCCT | TTGATCGTGGCAGAGTGAGC | Primer blast |
| rpl13a | CATAGGAAGCTGGGAGCAAG | GCCCTCCAATCAGTCTTCTG | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczepaniak, J.; Jagiello, J.; Wierzbicki, M.; Nowak, D.; Sobczyk-Guzenda, A.; Sosnowska, M.; Jaworski, S.; Daniluk, K.; Szmidt, M.; Witkowska-Pilaszewicz, O.; et al. Reduced Graphene Oxides Modulate the Expression of Cell Receptors and Voltage-Dependent Ion Channel Genes of Glioblastoma Multiforme. Int. J. Mol. Sci. 2021, 22, 515. https://doi.org/10.3390/ijms22020515
Szczepaniak J, Jagiello J, Wierzbicki M, Nowak D, Sobczyk-Guzenda A, Sosnowska M, Jaworski S, Daniluk K, Szmidt M, Witkowska-Pilaszewicz O, et al. Reduced Graphene Oxides Modulate the Expression of Cell Receptors and Voltage-Dependent Ion Channel Genes of Glioblastoma Multiforme. International Journal of Molecular Sciences. 2021; 22(2):515. https://doi.org/10.3390/ijms22020515
Chicago/Turabian StyleSzczepaniak, Jaroslaw, Joanna Jagiello, Mateusz Wierzbicki, Dorota Nowak, Anna Sobczyk-Guzenda, Malwina Sosnowska, Slawomir Jaworski, Karolina Daniluk, Maciej Szmidt, Olga Witkowska-Pilaszewicz, and et al. 2021. "Reduced Graphene Oxides Modulate the Expression of Cell Receptors and Voltage-Dependent Ion Channel Genes of Glioblastoma Multiforme" International Journal of Molecular Sciences 22, no. 2: 515. https://doi.org/10.3390/ijms22020515
APA StyleSzczepaniak, J., Jagiello, J., Wierzbicki, M., Nowak, D., Sobczyk-Guzenda, A., Sosnowska, M., Jaworski, S., Daniluk, K., Szmidt, M., Witkowska-Pilaszewicz, O., Strojny-Cieslak, B., & Grodzik, M. (2021). Reduced Graphene Oxides Modulate the Expression of Cell Receptors and Voltage-Dependent Ion Channel Genes of Glioblastoma Multiforme. International Journal of Molecular Sciences, 22(2), 515. https://doi.org/10.3390/ijms22020515