A Blunted Sympathetic Function and an Enhanced Nitrergic Activity Contribute to Reduce Mesenteric Resistance in Hyperthyroidism

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Animal Evolution
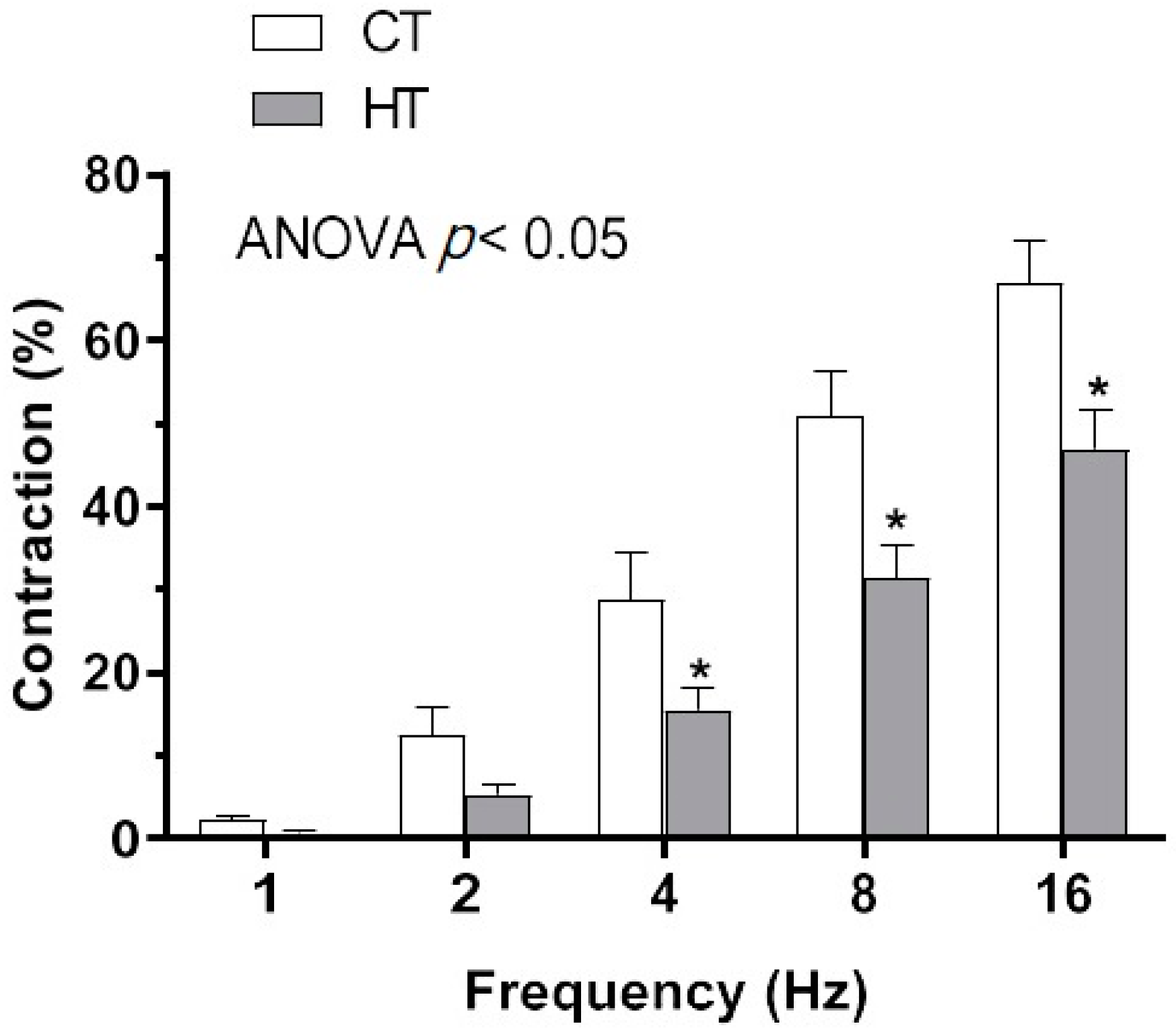
2.2. Effect of Hyperthyroidism in Electrical Field Stimulation (EFS)-Induced Vasoconstriction
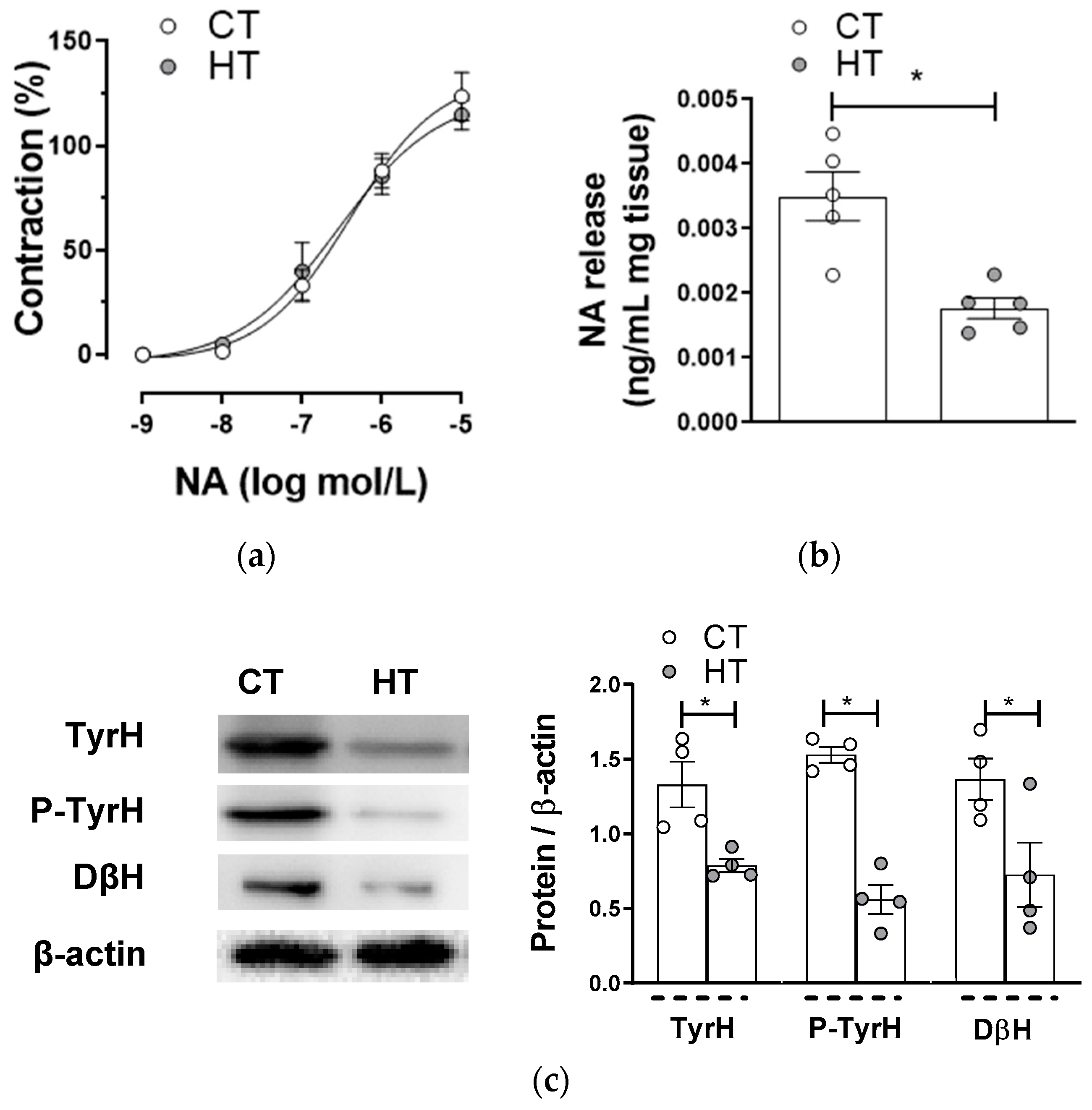
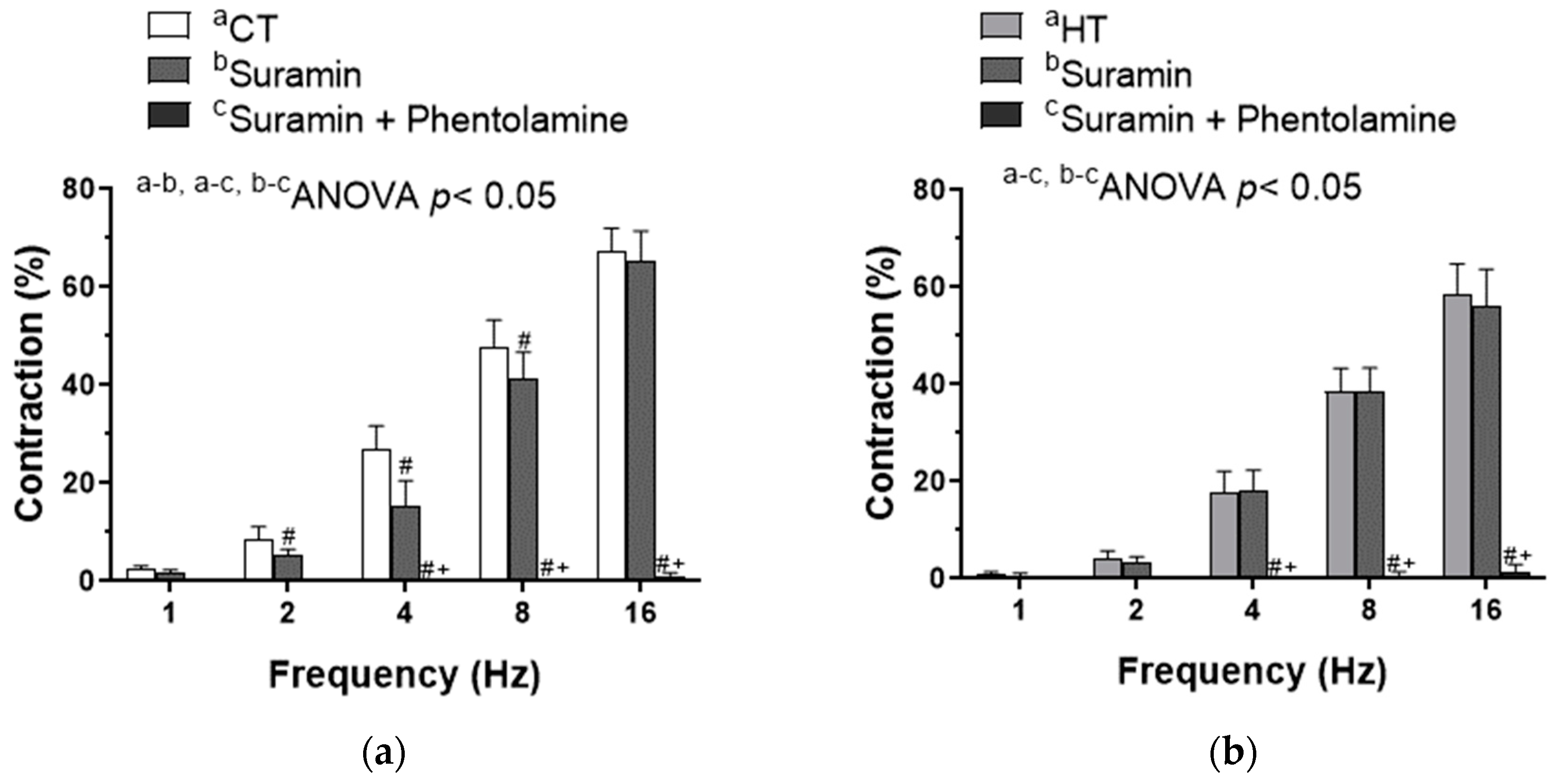
2.3. Effect of Hyperthyroidism in the Sympathetic Neurotransduction in Mesenteric Vascular Innervation
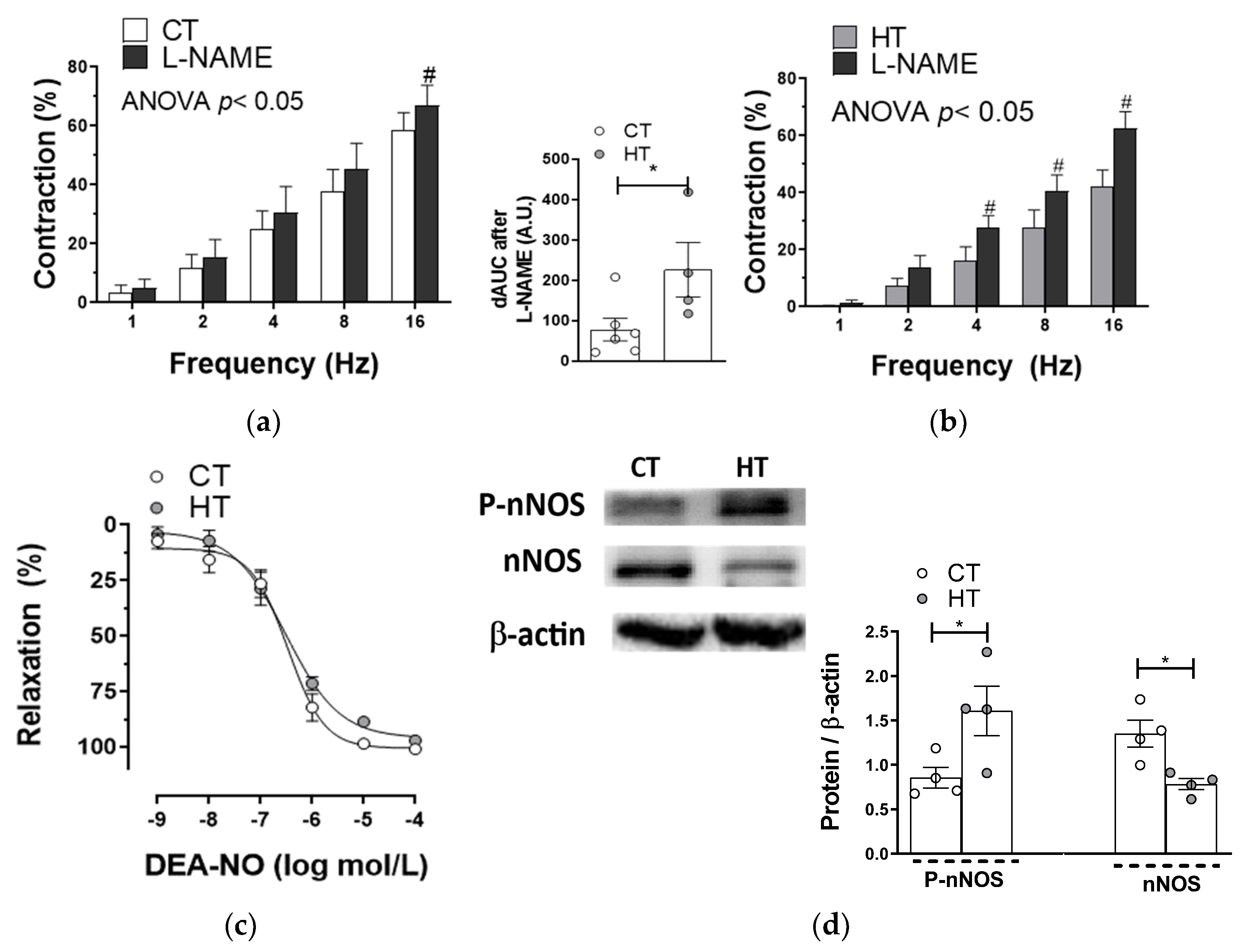
2.4. Participation of the Nitrergic Component in Vascular Responses to EFS
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Blood Pressure Measurements
4.3. Animal Euthanasia and Sample Collection
4.4. Citrate Synthase Activity
4.5. Vascular Reactivity
4.6. Neurotransmitter Release
4.7. PKA and PKC Activity Assays
4.8. Western Blot Analysis
4.9. Drugs and Antibodies Used
4.10. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CT | Control rats |
| DβH | Dopamine β Hydroxylase |
| DAF | Diaminofluorescein |
| dAUC | Differences in area under the curve |
| DEA-NO | Diethylamine NONOate |
| EFS | Electrical field stimulation |
| HT | Hyperthyroidic rats |
| KHS | Krebs–Henseleit solution |
| L-NAME | Nω-Nitro-L-arginine methyl ester |
| NA | Noradrenaline |
| nNOS | Neuronal nitric oxide synthase |
| NO | Nitric oxide |
| PI3K | Phosphatydil inositol 3 phosphate kinase |
| PKA | Protein Kinase A |
| PKC. | Protein Kinase C |
| S.E.M. | Standard error of the media |
| SMA | Superior mesenteric artery |
| TyrH | Tyroxine hydroxylase |
| TTX | Tetrodotoxin |
| VSMC | Vascular smooth muscle cells |
References
- Axelband, F.; Dias, J.; Ferrão, F.M.; Einicker-Lamas, M. Nongenomic signaling pathways triggered by thyroid hormones and their metabolite 3-iodothyronamine on the cardiovascular system. J. Cell Physiol. 2011, 226, 21–28. [Google Scholar] [CrossRef]
- Sinha, R.; Yen, P.M. Cellular Action of Thyroid Hormone; Feingold, K.R., Anawalt, B., Boyce, A., Eds.; Endotext [Internet]; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK285568/?report=classic (accessed on 10 November 2020).
- LiVolsi, V.A.; Baloch, Z.W. The Pathology of Hyperthyroidism. Front. Endocrinol. 2018, 9, 737. [Google Scholar] [CrossRef]
- De Leo, S.; Lee, S.Y.; Braverman, L.E. Hyperthyroidism. Lancet 2016, 388, 906–918. [Google Scholar] [CrossRef] [Green Version]
- Ertec, S.; Cicero, A.F. Hyperthyroidism and cardiovascular complications: A narrative review on the basis of pathophysiology. Arch. Med. Sci. 2013, 9, 944–952. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, G.; Makowski, K.; Michałkiewicz, D.; Kowal, J.; Ruchala, M.; Szczepanek, E.; Gielerak, G. The influence of subclinical hyperthyroidism on blood pressure, heart rate variability, and prevalence of arrhythmias. Thyroid 2012, 22, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Ojamaa, K.; Klemperer, J.D.; Klein, I. Acute effects of thyroid hormone on vascular smooth muscle. Thyroid 1996, 6, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Sepúlveda, M.A.; Ceravolo, G.S.; Fortes, Z.B.; Carvalho, M.H.; Tostes, R.C.; Laurindo, F.R.; Webb, R.C.; Barreto-Chaves, M.L. Thyroid hormone stimulates NO production via activation of the PI3K/Akt pathway in vascular myocytes. Cardiovasc. Res. 2010, 85, 560–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichiki, T. Thyroid Hormone and Vascular Remodeling. J. Atheroscler. Thromb. 2016, 23, 266–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jabbar, A.; Pingitore, A.; Pearce, S.H.; Zaman, A.; Iervasi, G.; Razvi, S. Thyroid hormones and cardiovascular disease. Nat. Rev. Cardiol. 2017, 14, 39–55. [Google Scholar]
- Fukuyama, K.; Ichiki, T.; Takeda, K.; Tokunou, T.; Iino, N.; Masuda, S.; Ishibashi, M.; Egashira, K.; Shimokawa, H.; Hirano, K.; et al. Downregulation of vascular angiotensin II type 1 receptor by thyroid hormone. Hypertension 2003, 41, 598–603. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Jing, M.; Gao, Y.; Liu, C.; Liu, L.; Jia, H.; Liu, P.; Chang, M. Study on the relationship between hyperthyroidism and vascular endothelial cell damage. Sci. Rep. 2020, 10, 6992. [Google Scholar] [CrossRef] [PubMed]
- Klein, I.; Ojamaa, K. Thyroid hormone and the cardiovascular system. N. Engl. J. Med. 2001, 344, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Ferrari, S.M.; Corrado, A.; Di Domenicantonio, A.; Fallahi, P. Autoimmune Thyroid Disorders. Autoimmun. Rev. 2015, 14, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.M.; Ruffilli, I.; Elia, G.; Ragusa, F.; Paparo, S.R.; Patrizio, A.; Mazzi, V.; Antonelli, A.; Fallahi, P. Chemokines in hyperthyroidism. J. Clin. Transl. Endocrinol. 2019, 16, 100196. [Google Scholar] [CrossRef]
- Cassado, A.D.A.; D’Império Lima, M.R.; Ramalho Bortoluci, K. Revisiting Mouse Peritoneal Macrophages: Heterogeneity, Development, and Function. Front. Immunol. 2015, 6, 225. [Google Scholar] [CrossRef] [Green Version]
- Martín, A.; Pérez-Girón, J.V.; Hernanz, R.; Palacios, R.; Briones, A.M.; Fortuño, A.; Zalba, G.; Salaices, M.; Alonso, M.J. Peroxisome proliferator-activated receptor-γ activation reduces cyclooxygenase-2 expression in vascular smooth muscle cells from hypertensive rats by interfering with oxidative stress. J. Hypertens. 2012, 30, 315–326. [Google Scholar] [CrossRef]
- Orejudo, M.; García-Redondo, A.B.; Rodrigues-Diez, R.R.; Rodrigues-Díez, R.; Santos-Sanchez, L.; Tejera-Muñoz, A.; Egido, J.; Selgas, R.; Salaices, M.; Briones, A.M.; et al. Interleukin-17A induces vascular remodeling of small arteries and blood pressure elevation. Clin. Sci. 2020, 134, 513–527. [Google Scholar] [CrossRef]
- Xavier, F.E.; Blanco-Rivero, J.; Avendaño, M.S.; Sastre, E.; Yela, R.; Velázquez, K.; Salaices, M.; Balfagón, G. Aldosterone alters the participation of endothelial factors in noradrenaline vasoconstriction differently in resistance arteries from normotensive and hypertensive rats. Eur. J. Pharmacol. 2011, 654, 280–288. [Google Scholar] [CrossRef]
- Caracuel, L.; Sastre, E.; Llévenes, P.; Prieto, I.; Funes, T.; Aller, M.Á.; Arias, J.; Balfagón, G.; Blanco-Rivero, J. Acute-on-chronic liver disease enhances phenylephrine-induced endothelial nitric oxide release in rat mesenteric resistance arteries through enhanced PKA, PI3K/AKT and cGMP signalling pathways. Sci. Rep. 2019, 9, 6993. [Google Scholar] [CrossRef]
- Sastre, E.; Márquez-Rodas, I.; Blanco-Rivero, J.; Balfagon, G. Perivascular innervation of the superior mesenteric artery: Pathophysiological implications. Rev. Neurol. 2010, 50, 727–737. [Google Scholar]
- Xavier, F.E. Nitrergic perivascular innervation in health and diseases: Focus on vascular tone regulation. Acta Physiol. 2020, 26, 13484. [Google Scholar]
- Aras-Lopez, R.; Blanco-Rivero, J.; Xavier, F.E.; Salaices, M.; Ferrer, M.; Balfagón, G. Dexamethasone decreases contraction to electrical field stimulation in mesenteric arteries from spontaneously hypertensive rats through decreases in thromboxane A2 release. J. Pharmacol. Exp. Ther. 2007, 322, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Rivero, J.; Sastre, E.; Caracuel, L.; Granado, M.; Balfagón, G. Breast feeding increases vasoconstriction induced by electrical field stimulation in rat mesenteric artery. Role of neuronal nitric oxide and ATP. PLoS ONE 2013, 8, e53802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sastre, E.; Blanco-Rivero, J.; Caracuel, L.; Callejo, M.; Balfagón, G. Alterations in perivascular sympathetic and nitrergic innervation function induced by late pregnancy in rat mesenteric arteries. PLoS ONE 2015, 10, e0126017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sastre, E.; Caracuel, L.; Blanco-Rivero, J.; Callejo, M.; Xavier, F.E.; Balfagón, G. Biphasic Effect of Diabetes on Neuronal Nitric Oxide Release in Rat Mesenteric Arteries. PLoS ONE 2016, 11, e0156793. [Google Scholar] [CrossRef] [Green Version]
- Llévenes, P.; Balfagón, G.; Blanco-Rivero, J. Thyroid hormones affect nitrergic innervation function in rat mesenteric artery: Role of the PI3K/AKT pathway. Vascul. Pharmacol. 2018, 108, 36–45. [Google Scholar] [CrossRef]
- Silva, J.E.; Bianco, S.D. Thyroid-adrenergic interactions: Physiological and clinical implications. Thyroid 2008, 18, 157–165. [Google Scholar] [CrossRef]
- Hong, W.; Li, G.; Nie, Y.; Zou, L.; Zhang, X.; Liu, S.; Li, G.; Xu, H.; Zhang, C.P.; Liang, S. Potential Involvement of P2 Receptors in the Pathological Processes of Hyperthyroidism: A Pilot Study. Ann. Clin. Lab. Sci. 2016, 46, 254–259. [Google Scholar]
- Gumanova, N.G.; Deev, A.D.; Klimushina, M.V.; Kots, A.Y.; Shalnova, S.A. Serum nitrate and nitrite are associated with the prevalence of various chronic diseases except cancer. Int. Angiol. 2017, 36, 160–166. [Google Scholar]
- Serfozo, Z.; Kiss, P.B.; Kukor, Z.; Lontay, B.; Palatka, K.; Varga, V.; Erdodi, F.; Elekes, K. Thyroid hormones affect the level and activity of nitric oxide synthase in rat cerebral cortex during postnatal development. Neurochem. Res. 2008, 33, 569–578. [Google Scholar] [CrossRef]
- Yousefzadeh, N.; Jeddi, S.; Ghasemi, A. Impaired Cardiovascular Function in Male Rats with Hypo and Hyperthyroidism: Involvement of Imbalanced Nitric Oxide Synthase Levels. Endocr. Metab. Immune Disord. Drug Targets 2020. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Alonso, M.J.; Salaices, M.; Marin, J.; Balfagón, G. Increase in neurogenic nitric oxide metabolism by endothelin-1 in mesenteric arteries from hypertensive rats. J. Cardiovasc. Pharmacol. 2000, 36, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Alonso, M.J.; Salaices, M.; Marin, J.; Balfagón, G. Angiotensin II increases neurogenic nitric oxide metabolism in mesenteric arteries from hypertensive rats. Life Sci. 2001, 68, 1169–1179. [Google Scholar] [CrossRef]
- Llévenes, P.; Rodrigues-Díez, R.; Cros-Brunsó, L.; Prieto, M.I.; Casaní, L.; Balfagón, G.; Blanco-Rivero, J. Beneficial Effect of a Multistrain Synbiotic Prodefen® Plus on the Systemic and Vascular Alterations Associated with Metabolic Syndrome in Rats: The Role of the Neuronal Nitric Oxide Synthase and Protein Kinase, A. Nutrients 2020, 12, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco-Rivero, J.; Balfagón, G.; Ferrer, M. Male castration increases neuronal nitric oxide synthase activity in the rat mesenteric artery through protein kinase C activation. J. Vasc. Res. 2005, 42, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Hellström, L.; Wahrenberg, H.; Reynisdottir, S.; Arner, P. Catecholamine-induced adipocyte lipolysis in human hyperthyroidism. J. Clin. Endocrinol. Metab. 1997, 82, 159–166. [Google Scholar] [CrossRef]
- Dalkjaer Riis, A.L.; Lunde Jørgensen, J.O.; Gjedde, S.; Nørrelund, H.; Jurik, A.G.; Nair, K.S.; Ivarsen, P.; Weeke, J.; Møller, N. Whole body and forearm substrate metabolism in hyperthyroidism: Evidence of increased basal muscle protein breakdown. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E1067–E1073. [Google Scholar] [CrossRef]
- Dalkjaer Riis, A.L.; Lunde Jørgensen, J.O.; Ivarsen, P.; Fryskyt, J.; Weeke, J.; Møller, N. Increased protein turnover and proteolysis is an early and primary feature of short-term experimental hyperthyroidism in healthy women. J. Clin. Endocrinol. Metab. 2008, 93, 3999–4005. [Google Scholar] [CrossRef] [Green Version]
- Peterson, M.E.; Castellano, C.A.; Rishniw, M. Evaluation of Body Weight, Body Condition, and Muscle Condition in Cats with Hyperthyroidism. J. Vet. Intern. Med. 2016, 30, 1780–1789. [Google Scholar] [CrossRef]
- Marinello, P.C.; Bernardes, S.S.; Guarnier, F.A.; Da Silva, F.N.X.; Borges, F.H.; Lopes, N.M.D.; Simão, A.N.C.; Armani, A.; Cecchini, R.; Cecchini, A.L. Isoflavin-β modifies muscle oxidative stress and prevents a thyrotoxicosis-induced loss of muscle mass in rats. Muscle Nerve 2017, 56, 975–981. [Google Scholar] [CrossRef]
- Szlejf, C.; Suemoto, C.K.; Janovsky, C.C.P.S.; Barreto, S.M.; Diniz, M.D.F.H.; Lotufo, P.A.; Bensenor, I.M. Thyroid Function and Sarcopenia: Results from the ELSA-Brasil Study. J. Am. Geriatr. Soc. 2020, 68, 1545–1553. [Google Scholar] [CrossRef]
- Egan, B.M.; Kjeldsen, S.E.; Grassi, G.; Esler, M.; Mancia, G. The global burden of hypertension exceeds 1.4 billion people: Should a systolic blood pressure target below 130 become the universal standard? J. Hypertens. 2019, 37, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Takala, J. Determinants of splanchnic blood flow. Br. J. Anaesth. 1996, 77, 50–58. [Google Scholar] [CrossRef] [PubMed]
- McAllister, R.M.; Grossenburg, V.D.; Delp, M.D.; Laughlin, M.H. Effects of hyperthyroidism on vascular contractile and relaxation responses. Am. J. Physiol. 1998, 274, E946–E953. [Google Scholar] [CrossRef] [PubMed]
- Utkan, T.; Yusuf, S.; Zafer, U.N.; Kemal, M.Y. The influence of experimental hyperthyroidism on responsiveness in rabbit aortic smooth muscle. Environ. Toxicol. Pharmacol. 1997, 3, 7–11. [Google Scholar] [CrossRef]
- Ozdemirci, S.; Yildiz, F.; Utkan, T.; Ulak, G.; Cetinaslan, B.; Erden, F.; Gacar, N. Impaired neurogenic and endothelium-dependent relaxant responses of corpus cavernosum smooth muscle from hyperthyroid rabbits. Eur. J. Pharmacol. 2001, 428, 105–111. [Google Scholar] [CrossRef]
- Lohmeier, T.E. The sympathetic nervous system and long-term blood pressure regulation. Am. J. Hypertens. 2001, 14, 147S–154S. [Google Scholar] [CrossRef] [Green Version]
- Claustre, J.; Balende, C.; Pujol, J.F. Influence of the thyroid hormone status on tyrosine hydroxylase in central and peripheral catecholaminergic structures. Neurochem. Int. 1996, 28, 277–281. [Google Scholar] [CrossRef]
- Pennacchio, G.E.; Neira, F.J.; Soaje, M.; Jahn, G.A.; Valdez, S.R. Effect of hyperthyroidism on circulating prolactin and hypothalamic expression of tyrosine hydroxylase, prolactin signaling cascade members and estrogen and progesterone receptors during late pregnancy and lactation in the rat. Mol. Cell Endocrinol. 2017, 442, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Yun, F.; Zhao, H.; Zhang, S.; Liu, Z.; Wang, X.; Wang, D.; Peng, W.; Li, A.; Xiu, C.; et al. Atrial sympathetic remodeling in experimental hyperthyroidism and hypothyroidism rats. Int. J. Cardiol. 2015, 187, 148–150. [Google Scholar] [CrossRef]
- Grassby, P.F.; McNeill, J.H. Hyperthyroidism induces supersensitivity to biogenic amines in rat vascular tissue via a pre- and a postjunctional mechanism. J. Pharmacol. Exp. Ther. 1998, 244, 1027–1035. [Google Scholar]
- Honda, H.; Iwata, T.; Mochizuki, T.; Kogo, H. Changes in vascular reactivity induced by acute hyperthyroidism in isolated rat aortae. Gen. Pharmacol. 2000, 34, 429–434. [Google Scholar] [CrossRef]
- Napoli, R.; Guardasole, V.; Angelini, V.; Zarra, E.; Terracciano, D.; D’Anna, C.; Matarazzo, M.; Oliviero, U.; Macchia, V.; Saccà, L. Acute effects of triiodothyronine on endothelial function in human subjects. J. Clin. Endocrinol Metab. 2007, 92, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Racchi, H.; Irarrázabal, M.J.; Howard, M.; Morán, S.; Zalaquett, R.; Huidobro-Toro, J.P. Adenosine 5′-triphosphate and neuropeptide Y are co-transmitters in conjunction with noradrenaline in the human saphenous vein. Br. J. Pharmacol. 1999, 126, 1175–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goonetilleke, L.; Ralevic, V.; Dunn, W.R. Influence of pressure on adenosine triphosphate function as a sympathetic neurotransmitter in small mesenteric arteries from the spontaneously hypertensive rat. J. Hypertens. 2013, 31, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Sousa, J.B.; Vieira-Rocha, M.S.; Sá, C.; Ferreirinha, F.; Correia-de-Sá, P.; Fresco, P.; Diniz, C. Lack of endogenous adenosine tonus on sympathetic neurotransmission in spontaneously hypertensive rat mesenteric artery. PLoS ONE 2014, 9, e105540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shatarat, A.; Dunn, W.R.; Ralevic, V. Raised tone reveals ATP as a sympathetic neurotransmitter in the porcine mesenteric arterial bed. Purinergic Signal. 2014, 10, 639–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwata, T.; Honda, H. Acute hyperthyroidism alters adrenoceptor- and muscarinic receptor-mediated responses in isolated rat renal and femoral arteries. Eur. J. Pharmacol. 2004, 493, 191–199. [Google Scholar] [CrossRef]
- Deng, J.; Zhao, R.; Zhang, Z.; Wang, J. Changes in vasoreactivity of rat large- and medium-sized arteries induced by hyperthyroidism. Exp. Toxicol. Pathol. 2010, 62, 317–322. [Google Scholar] [CrossRef]
- Zhang, L.; Medina, M.P.; Hernández, V.S.; Estrada, F.S.; Vega-González, A. Vasopressinergic network abnormalities potentiate conditioned anxious state of rats subjected to maternal hyperthyroidism. Neurosci. 2010, 168, 416–428. [Google Scholar] [CrossRef]
- Blanco-Rivero, J.; Roque, F.R.; Sastre, E.; Caracuel, L.; Couto, G.K.; Avendaño, M.S.; Paula, S.M.; Rossoni, L.V.; Salaices, M.; Balfagón, G. Aerobic exercise training increases neuronal nitric oxide release and bioavailability and decreases noradrenaline release in mesenteric artery from spontaneously hypertensive rats. J. Hypertens. 2013, 31, 916–926. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.C.; Owman, C. Contractile response and amine receptor mechanism in isolated middle cerebral artery of the cat. Brain Res. 1971, 27, 33–42. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CT | HT | |
|---|---|---|
| Body weight gain (g) | 34.1 ± 4.6 | 21.3 ± 4.9 * |
| Tibia length (cm) | 3.6 ± 0.2 | 3.7 ± 0.2 |
| Systolic blood pressure (mm Hg) | 107.1 ± 1.3 | 133.0 ± 3.0 * |
| Heart weight/tibia length (g/cm tibia) | 0.31 ± 0.01 | 0.33 ± 0.01 |
| Adipose tissue (g/cm tibia) | 3.19 ± 0.1 | 2.43 ± 0.2 * |
| Soleus (g/cm tibia) | 41.06 ± 3.2 | 41.72 ± 4.2 |
| Citrate synthase activity (nmol/min mg protein) | 31.9 ± 6.4 | 126.6 ± 49.7 * |
| 1 Hz | 2 Hz | 4 Hz | 8 Hz | 16 Hz | |
|---|---|---|---|---|---|
| CT | 2.1 ± 0.5 | 12.3 ± 3.4 | 28.8 ± 5.6 | 50.9 ± 5.5 | 66.9 ± 5.1 |
| + TTX | 0 | 0 | 0 | 0.31 ± 0.1 * | 2.5 ± 0.6 * |
| HT | 0.66 ± 0.26 | 5.21 ± 1.19 | 15.45 ± 2.66 | 31.44 ±3.79 | 46.9 ± 4.7 |
| + TTX | 0 | 0 | 0 | 0.2 ± 0.1 * | 2.0 ± 0.2 * |
| CT | HT | |
|---|---|---|
| H89 | 58.6 ± 3.5 | 49.3 ± 5.4 |
| Calphostin C | 51.1 ± 10.9 | 80.8 ± 3.2 * |
| LY294002 | 52.7 ± 5.1 | 69.6 ± 4.2 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cros-Brunsó, L.; Camacho-Rodríguez, L.; Martínez-González, Á.; Llévenes, P.; Salaices, M.; García-Redondo, A.B.; Blanco-Rivero, J. A Blunted Sympathetic Function and an Enhanced Nitrergic Activity Contribute to Reduce Mesenteric Resistance in Hyperthyroidism. Int. J. Mol. Sci. 2021, 22, 570. https://doi.org/10.3390/ijms22020570
Cros-Brunsó L, Camacho-Rodríguez L, Martínez-González Á, Llévenes P, Salaices M, García-Redondo AB, Blanco-Rivero J. A Blunted Sympathetic Function and an Enhanced Nitrergic Activity Contribute to Reduce Mesenteric Resistance in Hyperthyroidism. International Journal of Molecular Sciences. 2021; 22(2):570. https://doi.org/10.3390/ijms22020570
Chicago/Turabian StyleCros-Brunsó, Laia, Laura Camacho-Rodríguez, Ángel Martínez-González, Pablo Llévenes, Mercedes Salaices, Ana Belen García-Redondo, and Javier Blanco-Rivero. 2021. "A Blunted Sympathetic Function and an Enhanced Nitrergic Activity Contribute to Reduce Mesenteric Resistance in Hyperthyroidism" International Journal of Molecular Sciences 22, no. 2: 570. https://doi.org/10.3390/ijms22020570
APA StyleCros-Brunsó, L., Camacho-Rodríguez, L., Martínez-González, Á., Llévenes, P., Salaices, M., García-Redondo, A. B., & Blanco-Rivero, J. (2021). A Blunted Sympathetic Function and an Enhanced Nitrergic Activity Contribute to Reduce Mesenteric Resistance in Hyperthyroidism. International Journal of Molecular Sciences, 22(2), 570. https://doi.org/10.3390/ijms22020570