Regulation of Anti-Apoptotic SOD2 and BIRC3 in Periodontal Cells and Tissues

 , , ,
, , ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
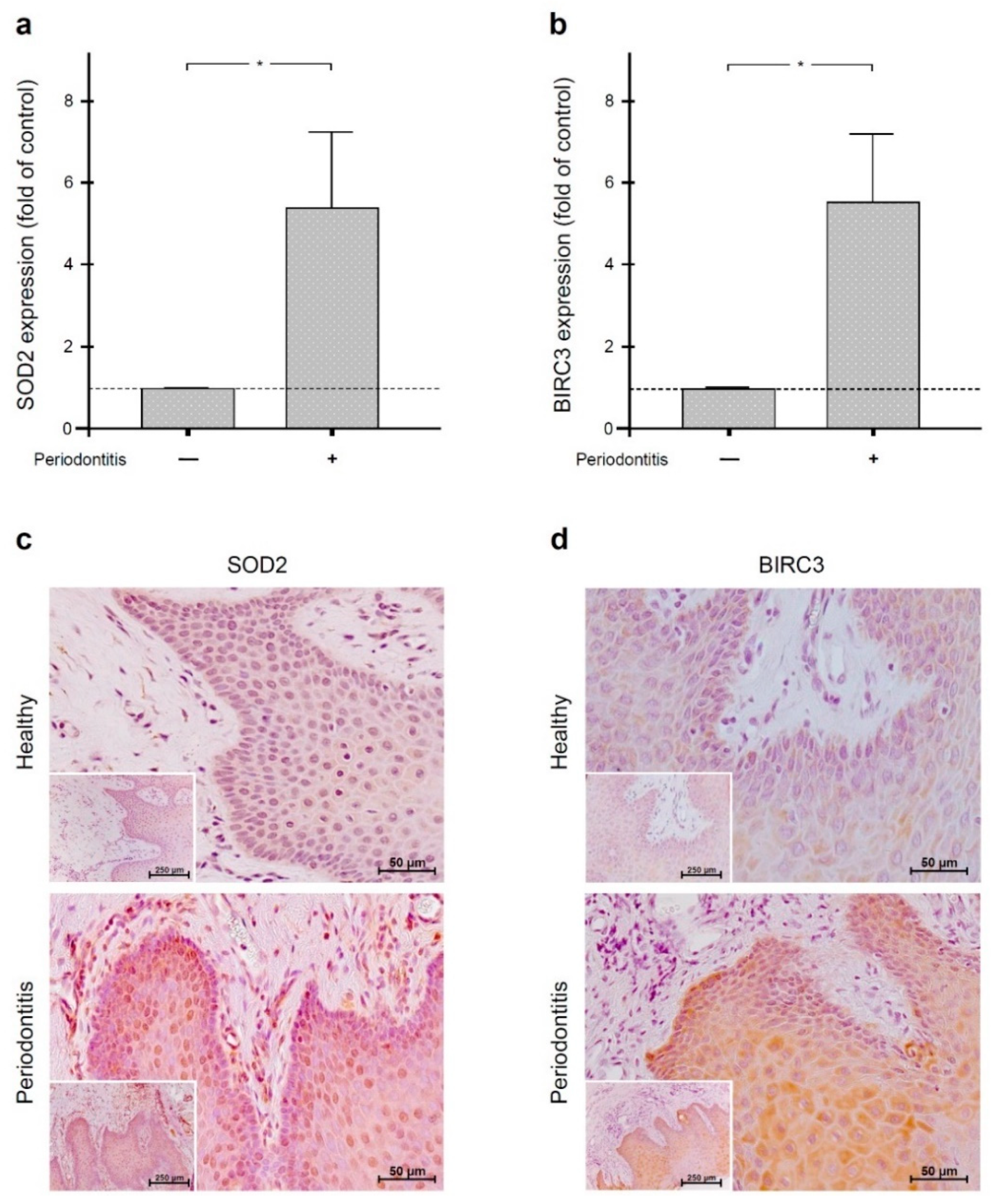
2.1. Regulation of SOD2 and BIRC3 by Periodontal Infection in Human Gingiva
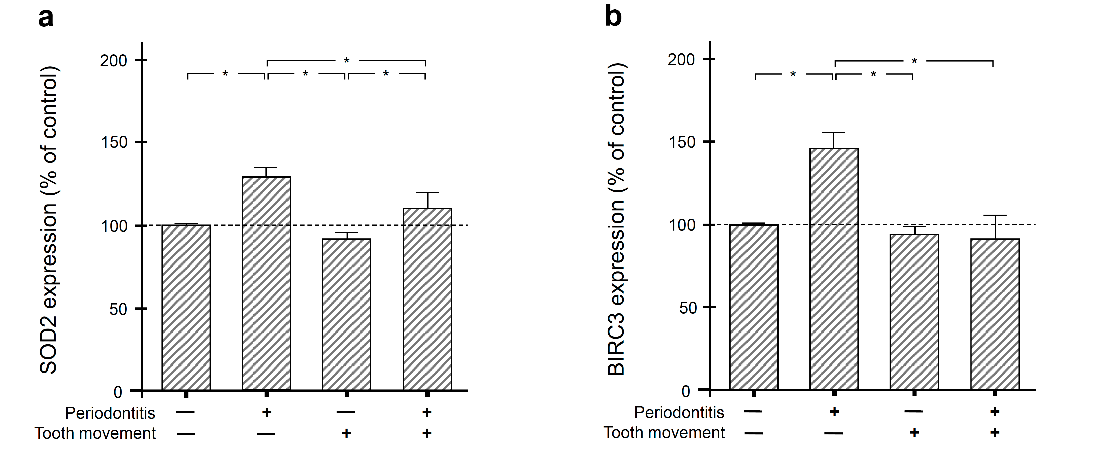
2.2. Regulation of SOD2 and BIRC3 by Periodontal Infection and/or Orthodontic Tooth Movement in Rat Gingiva
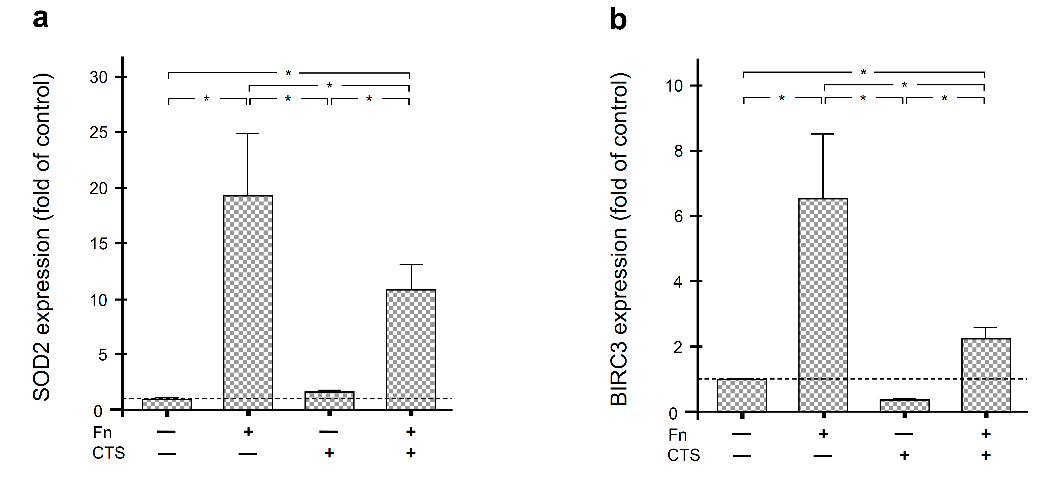
2.3. Regulation of SOD2 and BIRC3 by F. nucleatum and/or Biomechanical Forces in Human Periodontal Fibroblasts
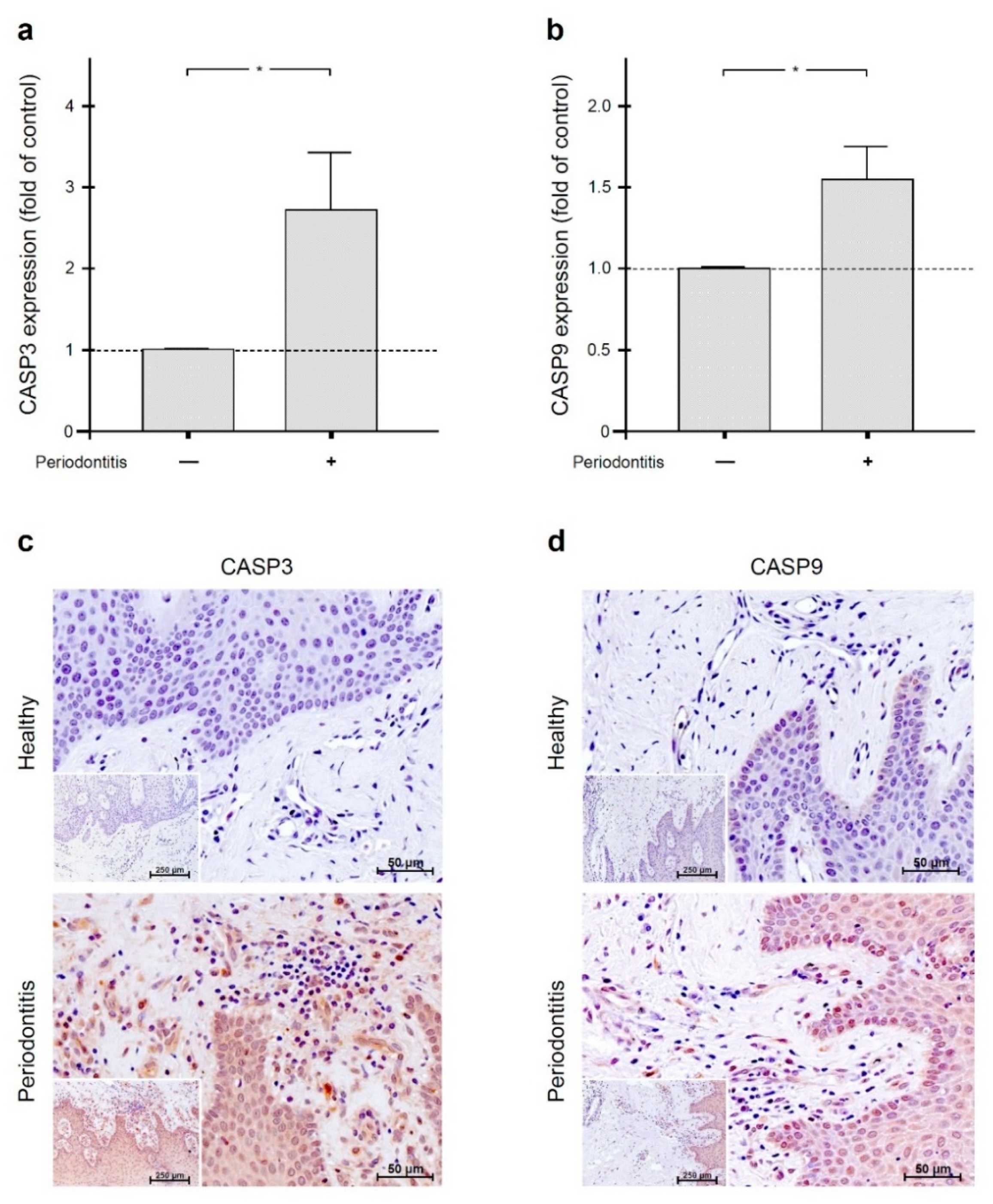
2.4. Increased Gingival Levels of Apoptotic Markers in Periodontal Infection
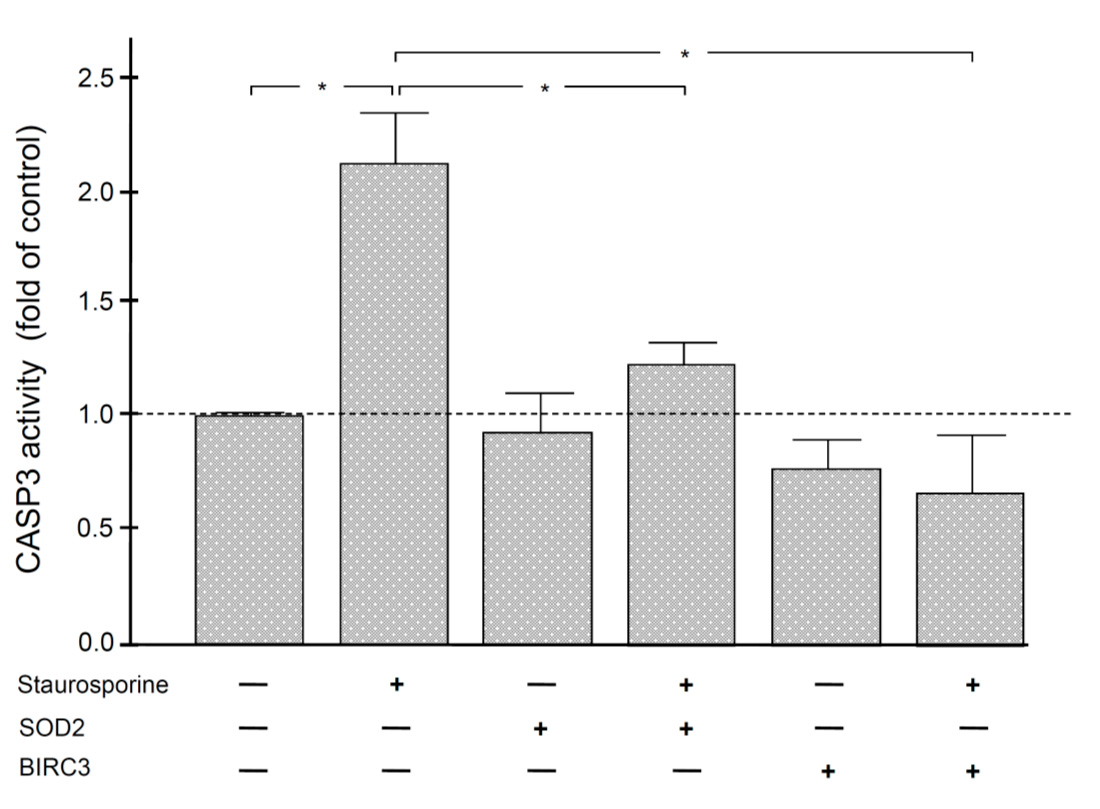
2.5. Anti-Apoptotic Effect of SOD2 and BIRC3 on Human Periodontal Fibroblasts
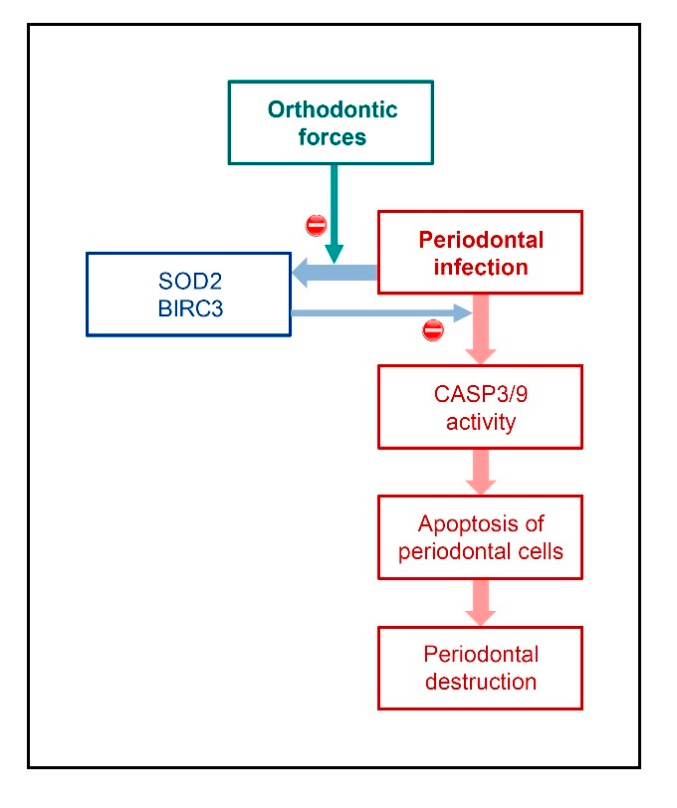
3. Discussion
4. Materials and Methods
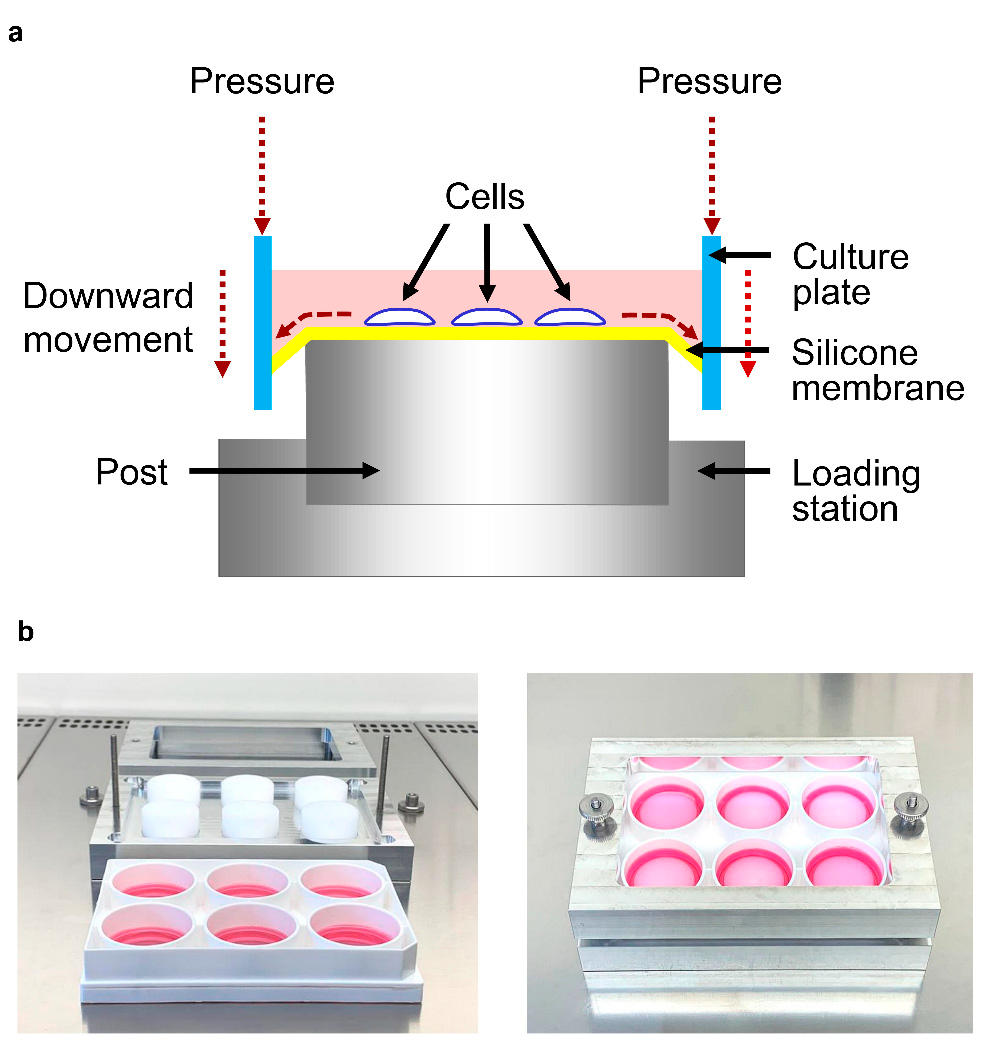
4.1. Culture and Treatment of Human Periodontal Fibroblasts
4.2. Human Gingival Biopsies from Periodontally Healthy and Diseased Subjects
4.3. Rat Gingival Biopsies and Animal Model
4.4. Analyses of Gene Expressions by Quantitative Real-Time Polymerase Chain Reaction (PCR)
4.5. Protein Analysis by Immunohistochemistry
4.6. Measurement of CASP3 Activity
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATCC | American Type Culture Collection |
| BIRC3 | baculoviral IAP repeat-containing protein 3 |
| CASP | caspase |
| cDNA | complementary desoxyribonucleic acid |
| CTS | constant tensile strain |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DNA | desoxyribonucleic acid |
| EDTA | ethylenediaminetetraacetic acid |
| FBS | fetal bovine serum |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| HRP | horseradish peroxidase |
| OD | optical density |
| PCR | polymerase chain reaction |
| PDL | periodontal ligament |
| RNA | ribonucleic acid |
| SOD2 | superoxide dismutase 2 |
References
- Kilian, M.; Chapple, I.L.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome—An update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Signat, B.; Roques, C.; Poulet, P.; Duffaut, D. Fusobacterium nucleatum in periodontal health and disease. Curr. Issues Mol. Biol. 2011, 13, 25–36. [Google Scholar] [PubMed]
- Han, Y.W. Fusobacterium nucleatum: A commensal-turned pathogen. Curr. Opin. Microbiol. 2015, 23, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rickard, A.H.; Gilbert, P.; High, N.J.; Kolenbrander, P.E.; Handley, P.S. Bacterial coaggregation: An integral process in the development of multi-species biofilms. Trends Microbiol. 2003, 11, 94–100. [Google Scholar] [CrossRef]
- Bradshaw, D.J.; Marsh, P.D.; Watson, G.K.; Allison, C. Role of Fusobacterium nucleatum and coaggregation in anaerobe survival in planktonic and biofilm oral microbial communities during aeration. Infect. Immun. 1998, 66, 4729–4732. [Google Scholar] [CrossRef] [Green Version]
- Diaz, P.I.; Zilm, P.S.; Rogers, A.H. Fusobacterium nucleatum supports the growth of Porphyromonas gingivalis in oxygenated and carbon-dioxide-depleted environments. Microbiology 2002, 148, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Meyle, J.; Chapple, I. Molecular aspects of the pathogenesis of periodontitis. Periodontology 2000 2015, 69, 7–17. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Chavakis, T.; Lambris, J.D. Current understanding of periodontal disease pathogenesis and targets for host-modulation therapy. Periodontology 2000 2020, 84, 14–34. [Google Scholar] [CrossRef]
- Pan, W.; Yang, L.; Li, J.; Xue, L.; Wei, W.; Ding, H.; Deng, S.; Tian, Y.; Yue, Y.; Wang, M.; et al. Traumatic occlusion aggravates bone loss during periodontitis and activates Hippo-YAP pathway. J. Clin. Periodontol. 2019, 46, 438–447. [Google Scholar] [CrossRef]
- Nakatsu, S.; Yoshinaga, Y.; Kuramoto, A.; Nagano, F.; Ichimura, I.; Oshino, K.; Yoshimura, A.; Yano, Y.; Hara, Y. Occlusal trauma accelerates attachment loss at the onset of experimental periodontitis in rats. J. Periodontal. Res. 2014, 49, 314–322. [Google Scholar] [CrossRef]
- Li, Y.; Jacox, L.A.; Little, S.H.; Ko, C.C. Orthodontic tooth movement: The biology and clinical implications. Kaohsiung J. Med. Sci. 2018, 34, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Sun, M.; Xie, Y.; Shu, R. mTOR Inhibition Rejuvenates the Aging Gingival Fibroblasts through Alleviating Oxidative Stress. Oxid. Med. Cell. Longev. 2017, 2017, 6292630. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.; Kim, T.J.; Lee, J.M.; Kim, D.Y. SOD2 is upregulated in periodontitis to reduce further inflammation progression. Oral Dis. 2018, 24, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Sun, T.; Yang, F.; Yuan, J.; Yang, M.; Kang, W.; Tang, D.; Zhang, J.; Feng, Q. The Pathogenic Effects of Fusobacterium nucleatum on the Proliferation, Osteogenic Differentiation, and Transcriptome of Osteoblasts. Front. Cell Dev. Biol. 2020, 8, 807. [Google Scholar] [CrossRef]
- Kitase, Y.; Yokozeki, M.; Fujihara, S.; Izawa, T.; Kuroda, S.; Tanimoto, K.; Moriyama, K.; Tanaka, E. Analysis of gene expression profiles in human periodontal ligament cells under hypoxia: The protective effect of CC chemokine ligand 2 to oxygen shortage. Arch. Oral Biol 2009, 54, 618–624. [Google Scholar] [CrossRef]
- Che, M.; Wang, R.; Li, X.; Wang, H.Y.; Zheng, X.F.S. Expanding roles of superoxide dismutases in cell regulation and cancer. Drug Discov. Today 2016, 21, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Flynn, J.M.; Melov, S. SOD2 in mitochondrial dysfunction and neurodegeneration. Free Radic. Biol. Med. 2013, 62, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Palazzotti, B.; Pani, G.; Colavitti, R.; De Leo, M.E.; Bedogni, B.; Borrello, S.; Galeotti, T. Increased growth capacity of cervical-carcinoma cells over-expressing manganous superoxide dismutase. Int. J. Cancer 1999, 82, 145–150. [Google Scholar] [CrossRef]
- Lalaoui, N.; Vaux, D.L. Recent advances in understanding inhibitor of apoptosis proteins. F1000Research 2018, 7, F1000 Faculty Rev-1889. [Google Scholar] [CrossRef]
- Silke, J.; Vucic, D. IAP family of cell death and signaling regulators. Methods Enzymol. 2014, 545, 35–65. [Google Scholar] [CrossRef]
- Erekat, N.S. Apoptosis and its Role in Parkinson’s Disease. In Parkinson’s Disease: Pathogenesis and Clinical Aspects; Stoker, T.B., Greenland, J.C., Eds.; Codon Publications: Brisbane, Australia, 2018; Chapter 4. [Google Scholar]
- Boatright, K.M.; Salvesen, G.S. Mechanisms of caspase activation. Curr. Opin. Cell Biol. 2003, 15, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Salvesen, G.S.; Riedl, S.J. Caspase mechanisms. Adv. Exp. Med. Biol. 2008, 615, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Zhou, T.; Yang, W.L.; Liu, J.; Shao, L.Q. Programmed cell death in periodontitis: Recent advances and future perspectives. Oral Dis. 2017, 23, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Calenic, B.; Yaegaki, K.; Murata, T.; Imai, T.; Aoyama, I.; Sato, T.; Ii, H. Oral malodorous compound triggers mitochondrial-dependent apoptosis and causes genomic DNA damage in human gingival epithelial cells. J. Periodontal Res. 2010, 45, 31–37. [Google Scholar] [CrossRef] [PubMed]
- González, O.A.; Stromberg, A.J.; Huggins, P.M.; Gonzalez-Martinez, J.; Novak, M.J.; Ebersole, J.L. Apoptotic genes are differentially expressed in aged gingival tissue. J. Dent. Res. 2011, 90, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Sancilio, S.; Gallorini, M.; Cataldi, A.; di Giacomo, V. Cytotoxicity and apoptosis induction by e-cigarette fluids in human gingival fibroblasts. Clin. Oral Investig. 2016, 20, 477–483. [Google Scholar] [CrossRef]
- Graves, D.T.; Liu, R.; Oates, T.W. Diabetes-enhanced inflammation and apoptosis: Impact on periodontal pathosis. Periodontology 2000 2007, 45, 128–137. [Google Scholar] [CrossRef]
- Harrel, S.K.; Nunn, M.E.; Hallmon, W.W. Is there an association between occlusion and periodontal destruction? Yes—Occlusal forces can contribute to periodontal destruction. J. Am. Dent. Assoc. 2006, 137, 1380–1392. [Google Scholar] [CrossRef]
- Harrel, S.K.; Nunn, M.E. The effect of occlusal discrepancies on periodontitis. II. Relationship of occlusal treatment to the progression of periodontal disease. J. Periodontol. 2001, 72, 495–505. [Google Scholar] [CrossRef]
- Fu, J.H.; Yap, A.U. Occlusion and periodontal disease--where is the link? Singap. Dent. J. 2007, 29, 22–33. [Google Scholar]
- Botelho, J.; Machado, V.; Proença, L.; Rua, J.; Martins, L.; Alves, R.; Cavacas, M.A.; Manfredini, D.; Mendes, J.J. Relationship between self-reported bruxism and periodontal status: Findings from a cross-sectional study. J. Periodontol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Manfredini, D.; Ahlberg, J.; Mura, R.; Lobbezoo, F. Bruxism is unlikely to cause damage to the periodontium: Findings from a systematic literature assessment. J. Periodontol. 2015, 86, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Perlitsh, M.J. Letter to the Editor: Re: Bruxism is Unlikely to Cause Damage to the Periodontium: Findings from a Systematic Literature Assessment. J. Periodontol. 2016, 87, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Milward, M.R.; Chapple, I.L.; Wright, H.J.; Millard, J.L.; Matthews, J.B.; Cooper, P.R. Differential activation of NF-kappaB and gene expression in oral epithelial cells by periodontal pathogens. Clin. Exp. Immunol. 2007, 148, 307–324. [Google Scholar] [CrossRef]
- Milward, M.R.; Chapple, I.L.; Carter, K.; Matthews, J.B.; Cooper, P.R. Micronutrient modulation of NF-κB in oral keratinocytes exposed to periodontal bacteria. Innate Immun. 2013, 19, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Sztukowska, M.; Ojo, A.; Scott, D.A.; Wang, H.; Lamont, R.J. FOXO responses to Porphyromonas gingivalis in epithelial cells. Cell. Microbiol. 2015, 17, 1605–1617. [Google Scholar] [CrossRef] [Green Version]
- Offenbacher, S.; Barros, S.P.; Paquette, D.W.; Winston, J.L.; Biesbrock, A.R.; Thomason, R.G.; Gibb, R.D.; Fulmer, A.W.; Tiesman, J.P.; Juhlin, K.D.; et al. Gingival transcriptome patterns during induction and resolution of experimental gingivitis in humans. J. Periodontol. 2009, 80, 1963–1982. [Google Scholar] [CrossRef]
- Duarte, P.M.; Napimoga, M.H.; Fagnani, E.C.; Santos, V.R.; Bastos, M.F.; Ribeiro, F.V.; Araújo, V.C.; Demasi, A.P. The expression of antioxidant enzymes in the gingivae of type 2 diabetics with chronic periodontitis. Arch. Oral Biol. 2012, 57, 161–168. [Google Scholar] [CrossRef]
- Wright, H.J.; Chapple, I.L.; Matthews, J.B.; Cooper, P.R. Fusobacterium nucleatum regulation of neutrophil transcription. J. Periodontal Res. 2011, 46, 1–12. [Google Scholar] [CrossRef]
- Xu, C.; Hao, Y.; Wei, B.; Ma, J.; Li, J.; Huang, Q.; Zhang, F. Apoptotic gene expression by human periodontal ligament cells following cyclic stretch. J. Periodontal Res. 2011, 46, 742–748. [Google Scholar] [CrossRef]
- Mao, S.; Park, Y.; Hasegawa, Y.; Tribble, G.D.; James, C.E.; Handfield, M.; Stavropoulos, M.F.; Yilmaz, O.; Lamont, R.J. Intrinsic apoptotic pathways of gingival epithelial cells modulated by Porphyromonas gingivalis. Cell. Microbiol. 2007, 9, 1997–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chukkapalli, S.S.; Velsko, I.M.; Rivera-Kweh, M.F.; Zheng, D.; Lucas, A.R.; Kesavalu, L. Polymicrobial Oral Infection with Four Periodontal Bacteria Orchestrates a Distinct Inflammatory Response and Atherosclerosis in ApoE null Mice. PLoS ONE 2015, 10, e0143291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugueno, I.M.; Batool, F.; Korah, L.; Benkirane-Jessel, N.; Huck, O. Porphyromonas gingivalis Differentially Modulates Apoptosome Apoptotic Peptidase Activating Factor 1 in Epithelial Cells and Fibroblasts. Am. J. Pathol 2018, 188, 404–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, H.; Bartold, P.M.; Dharmapatni, A.A.; Holding, C.A.; Haynes, D.R. Inhibition of apoptosis in periodontitis. J. Dent. Res. 2010, 89, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Bantel, H.; Beikler, T.; Flemmig, T.F.; Schulze-Osthoff, K. Caspase activation is involved in chronic periodontitis. FEBS Lett. 2005, 579, 5559–5564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamonal, J.; Bascones, A.; Acevedo, A.; Blanco, E.; Silva, A. Apoptosis in chronic adult periodontitis analyzed by in situ DNA breaks, electron microscopy, and immunohistochemistry. J. Periodontol. 2001, 72, 517–525. [Google Scholar] [CrossRef]
- Pradeep, A.R.; Suke, D.K.; Prasad, M.V.; Singh, S.P.; Martande, S.S.; Nagpal, K.; Naik, S.B.; Guruprasad, C.N.; Raju, A.P.; Singh, P.; et al. Expression of key executioner of apoptosis caspase-3 in periodontal health and disease. J. Investig. Clin. Dent. 2016, 7, 174–179. [Google Scholar] [CrossRef]
- Aral, K.; Aral, C.A.; Kapila, Y. The role of caspase-8, caspase-9, and apoptosis inducing factor in periodontal disease. J. Periodontol. 2019, 90, 288–294. [Google Scholar] [CrossRef]
- Abuhussein, H.; Bashutski, J.D.; Dabiri, D.; Halubai, S.; Layher, M.; Klausner, C.; Makhoul, H.; Kapila, Y. The role of factors associated with apoptosis in assessing periodontal disease status. J. Periodontol. 2014, 85, 1086–1095. [Google Scholar] [CrossRef]
- Moffatt, C.E.; Whitmore, S.E.; Griffen, A.L.; Leys, E.J.; Lamont, R.J. Filifactor alocis interactions with gingival epithelial cells. Mol. Oral Microbiol. 2011, 26, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; de Brito Bezerra, B.; Pacios, S.; Andriankaja, O.; Li, Y.; Tsiagbe, V.; Schreiner, H.; Fine, D.H.; Graves, D.T. Aggregatibacter actinomycetemcomitans infection enhances apoptosis in vivo through a caspase-3-dependent mechanism in experimental periodontitis. Infect. Immun. 2012, 80, 2247–2256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhuang, J.; Zhao, D.; Xu, C. Interaction between caspase-3 and caspase-5 in the stretch-induced programmed cell death in the human periodontal ligament cells. J. Cell. Physiol. 2019, 234, 13571–13581. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhao, D.; Zhuang, J.; Zhang, F.; Xu, C. Caspase-8 and caspase-9 functioned differently at different stages of the cyclic stretch-induced apoptosis in human periodontal ligament cells. PLoS ONE 2016, 11, e0168268. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Xu, C.; Sun, S.Y.; Zhang, F.Q. Cyclic stretching force induces apoptosis in human periodontal ligament cells via caspase-9. Arch. Oral Biol. 2009, 54, 864–870. [Google Scholar] [CrossRef]
- Tsuda, H.; Ning, Z.; Yamaguchi, Y.; Suzuki, N. Programmed cell death and its possible relationship with periodontal disease. J. Oral Sci. 2012, 54, 137–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nokhbehsaim, M.; Keser, S.; Nogueira, A.V.; Cirelli, J.A.; Jepsen, S.; Jäger, A.; Eick, S.; Deschner, J. Beneficial effects of adiponectin on periodontal ligament cells under normal and regenerative conditions. J. Diabetes Res. 2014, 2014, 796565. [Google Scholar] [CrossRef] [Green Version]
- Nokhbehsaim, M.; Keser, S.; Nogueira, A.V.; Jäger, A.; Jepsen, S.; Cirelli, J.A.; Bourauel, C.; Eick, S.; Deschner, J. Leptin effects on the regenerative capacity of human periodontal cells. Int. J. Endocrinol. 2014, 2014, 180304. [Google Scholar] [CrossRef]
- Damanaki, A.; Nokhbehsaim, M.; Eick, S.; Götz, W.; Winter, J.; Wahl, G.; Jäger, A.; Jepsen, S.; Deschner, J. Regulation of NAMPT in human gingival fibroblasts and biopsies. Mediat. Inflamm. 2014, 2014, 912821. [Google Scholar] [CrossRef]
- Rath-Deschner, B.; Memmert, S.; Damanaki, A.; Nokhbehsaim, M.; Eick, S.; Cirelli, J.A.; Götz, W.; Deschner, J.; Jäger, A.; Nogueira, A.V.B. CXCL1, CCL2, and CCL5 modulation by microbial and biomechanical signals in periodontal cells and tissues-in vitro and in vivo studies. Clin. Oral Investig. 2020, 24, 3661–3670. [Google Scholar] [CrossRef] [Green Version]
- Rath-Deschner, B.; Memmert, S.; Damanaki, A.; de Molon, R.S.; Nokhbehsaim, M.; Eick, S.; Kirschneck, C.; Cirelli, J.A.; Deschner, J.; Jäger, A.; et al. CXCL5, CXCL8, and CXCL10 regulation by bacteria and mechanical forces in periodontium. Ann. Anat. 2020, 234, 151648. [Google Scholar] [CrossRef]
- Memmert, S.; Damanaki, A.; Weykopf, B.; Rath-Deschner, B.; Nokhbehsaim, M.; Götz, W.; Gölz, L.; Till, A.; Deschner, J.; Jäger, A. Autophagy in periodontal ligament fibroblasts under biomechanical loading. Cell Tissue Res. 2019, 378, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, A.V.; de Molon, R.S.; Nokhbehsaim, M.; Deschner, J.; Cirelli, J.A. Contribution of biomechanical forces to inflammation-induced bone resorption. J. Clin. Periodontol. 2017, 44, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Schminke, B.; Vom Orde, F.; Gruber, R.; Schliephake, H.; Bürgers, R.; Miosge, N. The pathology of bone tissue during peri-implantitis. J. Dent. Res. 2015, 94, 354–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rath-Deschner, B.; Nogueira, A.V.B.; Memmert, S.; Nokhbehsaim, M.; Augusto Cirelli, J.; Eick, S.; Miosge, N.; Kirschneck, C.; Kesting, M.; Deschner, J.; et al. Regulation of Anti-Apoptotic SOD2 and BIRC3 in Periodontal Cells and Tissues. Int. J. Mol. Sci. 2021, 22, 591. https://doi.org/10.3390/ijms22020591
Rath-Deschner B, Nogueira AVB, Memmert S, Nokhbehsaim M, Augusto Cirelli J, Eick S, Miosge N, Kirschneck C, Kesting M, Deschner J, et al. Regulation of Anti-Apoptotic SOD2 and BIRC3 in Periodontal Cells and Tissues. International Journal of Molecular Sciences. 2021; 22(2):591. https://doi.org/10.3390/ijms22020591
Chicago/Turabian StyleRath-Deschner, Birgit, Andressa Vilas Boas Nogueira, Svenja Memmert, Marjan Nokhbehsaim, Joni Augusto Cirelli, Sigrun Eick, Nicolai Miosge, Christian Kirschneck, Marco Kesting, James Deschner, and et al. 2021. "Regulation of Anti-Apoptotic SOD2 and BIRC3 in Periodontal Cells and Tissues" International Journal of Molecular Sciences 22, no. 2: 591. https://doi.org/10.3390/ijms22020591
APA StyleRath-Deschner, B., Nogueira, A. V. B., Memmert, S., Nokhbehsaim, M., Augusto Cirelli, J., Eick, S., Miosge, N., Kirschneck, C., Kesting, M., Deschner, J., Jäger, A., & Damanaki, A. (2021). Regulation of Anti-Apoptotic SOD2 and BIRC3 in Periodontal Cells and Tissues. International Journal of Molecular Sciences, 22(2), 591. https://doi.org/10.3390/ijms22020591