Molecular and Structural Evolution of Cytochrome P450 Aromatase



Abstract
:1. Introduction
2. Results
2.1. Multiple Sequence Alignment
2.1.1. Structural Conservation
2.1.2. Functional Conservation
2.1.3. Conservation of the Substrate Recognition Sites (SRSs)
2.1.4. Consensus Sequence for Post-Translational Modifications
2.1.5. Interaction with the Redox Partner
2.2. Homology Modeling of Evolutionarily Old Aromatase
3. Discussion
4. Materials and Methods
4.1. Multiple Sequence and Structural Alignments
4.2. Homology Modeling
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thompson, E.A.; Siiteri, P.K. Utilization of oxygen and reduced nicotinamide adenine dinucleotide phosphate by human placental microsomes during aromatization of androstenedione. J. Biol. Chem. 1974, 249, 5364–5372. [Google Scholar] [CrossRef]
- Simpson, E.R.; Mahendroo, M.S.; Means, G.D.; Kilgore, M.W.; Hinshelwood, M.M.; Graham-Lorence, S.; Amarneh, B.; Ito, Y.; Fisher, C.R.; Michael, M.D.; et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocr. Rev. 1994, 15, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Coon, M.J. Cytochrome P450: Nature’s most versatile biological catalyst. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Cytochrome p450 enzymes in the generation of commercial products. Nat. Rev. Drug. Discov. 2002, 1, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Di Nardo, G.; Gilardi, G. Natural compounds as pharmaceuticals: The key role of cytochromes p450 reactivity. Trends Biochem. Sci. 2020, 45, 511–525. [Google Scholar] [CrossRef]
- Nelson, D.R.; Goldstone, J.V.; Stegeman, J.J. The Cytochrome P450 Genesis Locus: The Origin and Evolution of Animal Cytochrome P450s. Philos. Trans. R. Soc. Lond B Biol. Sci. 2013, 368, 20120474. [Google Scholar] [CrossRef] [Green Version]
- Callard, G.V.; Tarrant, A.M.; Novillo, A.; Yacci, P.; Ciaccia, L.; Vajda, S.; Chuang, G.-Y.; Kozakov, D.; Greytak, S.R.; Sawyer, S.; et al. Evolutionary origins of the estrogen signaling system: Insights from amphioxus. J. Steroid Biochem. Mol. Biol. 2011, 127, 176–188. [Google Scholar] [CrossRef] [Green Version]
- Callard, G.V.; Pudney, J.A.; Kendall, S.L.; Reinboth, R. In vitro conversion of androgen to estrogen in amphioxus gonadal tissues. Gen. Comp. Endocrinol. 1984, 56, 53–58. [Google Scholar] [CrossRef]
- Mizuta, T.; Kubokawa, K. Presence of sex steroids and cytochrome P450 genes in amphioxus. Endocrinology 2007, 148, 3554–3565. [Google Scholar] [CrossRef] [Green Version]
- Kishida, M.; Callard, G.V. Distinct cytochrome P450 aromatase isoforms in zebrafish (Danio rerio) brain and ovary are differentially programmed and estrogen regulated during early development. Endocrinology 2001, 142, 740–750. [Google Scholar] [CrossRef]
- Tchoudakova, A.; Kishida, M.; Wood, E.; Callard, G.V. Promoter characteristics of two cyp19 genes differentially expressed in the brain and ovary of teleost fish. J. Steroid Biochem. Mol. Biol. 2001, 78, 427–439. [Google Scholar] [CrossRef]
- Conley, A.J.; Corbin, C.J.; Hughes, A.L. Adaptive evolution of mammalian aromatases: Lessons from Suiformes. J. Exp. Zool. Part A: Ecol. Genet. Physiol. 2009, 311, 346–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbin, C.J.; Hughes, A.L.; Heffelfinger, J.R.; Berger, T.; Waltzek, T.B.; Roser, J.F.; Santos, T.C.; Miglino, M.A.; Oliveira, M.F.; Braga, F.C.; et al. Evolution of suiform aromatases: Ancestral duplication with conservation of tissue-specific expression in the collared peccary (Pecari tayassu). J. Mol. Evol. 2007, 65, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.J.; Hewitt, S.C.; Arao, Y.; Korach, K.S. Estrogen hormone biology. Curr. Top. Dev. Biol. 2017, 125, 109–146. [Google Scholar] [CrossRef]
- McEwen, B.S.; Milner, T.A. Understanding the broad influence of sex hormones and sex differences in the brain. J. Neurosci. Res. 2017, 95, 24–39. [Google Scholar] [CrossRef] [Green Version]
- Galea, L.A.M.; Frick, K.M.; Hampson, E.; Sohrabji, F.; Choleris, E. Why estrogens matter for behavior and brain health. Neurosci. Biobehav. Rev. 2017, 76, 363–379. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.; Laurent, M.R.; Dubois, V.; Claessens, F.; O’Brien, C.A.; Bouillon, R.; Vanderschueren, D.; Manolagas, S.C. Estrogens and androgens in skeletal physiology and pathophysiology. Physiol. Rev. 2017, 97, 135–187. [Google Scholar] [CrossRef]
- Brincat, M.P.; Baron, Y.M.; Galea, R. Estrogens and the skin. Climacteric 2005, 8, 110–123. [Google Scholar] [CrossRef]
- Knowlton, A.A.; Lee, A.R. Estrogen and the cardiovascular system. Pharmacol. Ther. 2012, 135, 54–70. [Google Scholar] [CrossRef] [Green Version]
- Bernasochi, G.B.; Rupasinghe, T.T.W.; Bell, J.R.; Roessner, U.; Boon, W.C.; Delbridge, L.M.D. A novel mass spectrometric methodology facilitates quantification of testosterone and progesterone, but not estrogens in cardiac and adipose tissues. J. Mol. Cell. Cardiol. 2020, 140, 11–12. [Google Scholar] [CrossRef]
- Wang, H.; Li, R.; Hu, Y. The alternative noncoding exons 1 of aromatase (Cyp19) gene modulate gene expression in a posttranscriptional manner. Endocrinology 2009, 150, 3301–3307. [Google Scholar] [CrossRef] [PubMed]
- Piferrer, F.; Blázquez, M. Aromatase distribution and regulation in fish. Fish Physiol. Biochem. 2005, 31, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Cornil, C.A.; Ball, G.F.; Balthazart, J. The dual action of estrogen hypothesis. Trends Neurosci. 2015, 38, 408–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coumailleau, P.; Pellegrini, E.; Adrio, F.; Diotel, N.; Cano-Nicolau, J.; Nasri, A.; Vaillant, C.; Kah, O. Aromatase, estrogen receptors and brain development in fish and amphibians. Biochim. Biophys. Acta 2015, 1849, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Pieau, C.; Dorizzi, M. Oestrogens and temperature-dependent sex determination in reptiles: All is in the gonads. J. Endocrinol. 2004, 181, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Buemio, A.; Chu, R.; Vafaee, M.; Crews, D. Epigenetic control of gonadal aromatase (cyp19a1) in temperature-dependent sex determination of red-eared slider turtles. PLoS ONE 2013, 8, e63599. [Google Scholar] [CrossRef] [Green Version]
- Flament, S. Sex Reversal in Amphibians. Sex Dev. 2016, 10, 267–278. [Google Scholar] [CrossRef]
- Nakamura, M.; Kobayashi, T.; Chang, X.-T.; Nagahama, Y. Gonadal Sex Differentiation in Teleost Fish. J. Exp. Zool. 1998, 281, 362–372. [Google Scholar] [CrossRef]
- Sunobe, T.; Nakamura, M.; Kobayashi, Y.; Kobayashi, T.; Nagahama, Y. Aromatase immunoreactivity and the role of enzymes in steroid pathways for inducing sex change in the hermaphrodite gobiid fish Trimma Okinawae. Comp. Biochem. Physiol. 2005, 141, 54–59. [Google Scholar] [CrossRef]
- Baravalle, R.; Ciaramella, A.; Baj, F.; Di Nardo, G.; Gilardi, G. Identification of endocrine disrupting chemicals acting on human aromatase. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 88–96. [Google Scholar] [CrossRef]
- Zhang, C.; Schilirò, T.; Gea, M.; Bianchi, S.; Spinello, A.; Magistrato, A.; Gilardi, G.; Di Nardo, G. Molecular basis for endocrine disruption by pesticides targeting aromatase and estrogen receptor. Int. J. Environ. Res. Public Health 2020, 17, 5664. [Google Scholar] [CrossRef] [PubMed]
- Kloas, W. Amphibians as a model for the study of endocrine disruptors. Int. Rev. Cytol. 2002, 216, 1–57. [Google Scholar] [CrossRef] [PubMed]
- Scholz, S.; Renner, P.; Belanger, S.E.; Busquet, F.; Davi, R.; Demeneix, B.A.; Denny, J.S.; Léonard, M.; McMaster, M.E.; Villeneuve, D.L.; et al. Alternatives to in vivo tests to detect endocrine disrupting chemicals (edcs) in fish and amphibians—Screening for estrogen, androgen and thyroid hormone disruption. Crit. Rev. Toxicol. 2013, 43, 45–72. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Mak, P.; Tchoudakova, A.; Callard, G.; Chen, S. Different catalytic properties and inhibitor responses of the goldfish brain and ovary aromatase isozymes. Gen. Comp. Endocrinol. 2001, 123, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.K.; Chiang, E.F.; Hsiao, P.H.; Chung, B. Phylogeny, expression and enzyme activity of zebrafish cyp19 (P450 aromatase) genes. J. Steroid Biochem. Mol. Biol. 2001, 79, 299–303. [Google Scholar] [CrossRef]
- Hasemann, C.A.; Kurumbail, R.G.; Boddupalli, S.S.; Peterson, J.A.; Deisenhofer, J. Structure and function of cytochromes P450: A comparative analysis of three crystal structures. Structure 1995, 3, 41–62. [Google Scholar] [CrossRef]
- Ghosh, D.; Griswold, J.; Erman, M.; Pangborn, W. Structural basis for androgen specificity and oestrogen synthesis in human aromatase. Nature 2009, 457, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Di Nardo, G.; Breitner, M.; Bandino, A.; Ghosh, D.; Jennings, G.K.; Hackett, J.C.; Gilardi, G. Evidence for an elevated aspartate pKa in the active site of human aromatase. J. Biol. Chem. 2015, 290, 1186–1196. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, S.; Li, H.; Poulos, T.L. Structural basis for effect or control and redox partner recognition in cytochrome P450. Science 2013, 340, 1227–1230. [Google Scholar] [CrossRef]
- Liou, S.-H.; Mahomed, M.; Lee, Y.-T.; Goodin, D.B. Effector roles of putidaredoxin on cytochrome P450cam conformational states. J. Am. Chem. Soc. 2016, 138, 10163–10172. [Google Scholar] [CrossRef]
- Hollingsworth, S.A.; Batabyal, D.; Nguyen, B.D.; Poulos, T.L. Conformational selectivity in cytochrome P450 redox partner interactions. Proc. Natl. Acad. Sci. USA 2016, 113, 8723–8728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Catucci, G.; Di Nardo, G.; Gilardi, G. Effector role of cytochrome P450 reductase for androstenedione binding to human aromatase. Int. J. Biol. Macromol. 2020, 164, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Kim, B.H.; Grishin, N.V. PROMALS3D: A tool for multiple protein sequence and structure alignments. Nucleic Acids Res. 2008, 36, 2295–2300. [Google Scholar] [CrossRef] [PubMed]
- Di Nardo, G.; Breitner, M.; Sadeghi, S.J.; Castrignanò, S.; Mei, G.; Di Venere, A.; Nicolai, E.; Allegra, P.; Gilardi, G. Dynamics and flexibility of human aromatase probed by FTIR and time resolved fluorescence spectroscopy. PLoS ONE 2013, 8, e82118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulos, T.L. Cytochrome P450 flexibility. Proc. Natl. Acad. Sci. USA 2003, 100, 13121–13122. [Google Scholar] [CrossRef] [Green Version]
- Kalsotra, A.; Strobel, H.W. Cytochrome P450 4F subfamily: At the crossroads of eicosanoid and drug metabolism. Pharm. Ther. 2006, 112, 589–611. [Google Scholar] [CrossRef]
- Mast, N.; Norcross, R.; Andersson, U.; Shou, M.; Nakayama, K.; Bjorkhem, I.; Pikuleva, I.A. Broad substrate specificity of human cytochrome P450 46A1 which initiates cholesterol degradation in the brain. Biochemistry 2003, 42, 14284–14292. [Google Scholar] [CrossRef]
- Lepesheva, G.I.; Waterman, M.R. Sterol 14α-demethylase cytochrome P450 (CYP51), a P450 in all biological kingdoms. Biochim. Biophys. Acta 2007, 1770, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Ray, W.J.; Bain, G.; Yao, M.; Gottlieb, D.I. CYP26, a novel mammalian cytochrome P450, is induced by retinoic acid and defines a new family. J. Biol. Chem. 1997, 272, 18702–18708. [Google Scholar] [CrossRef] [Green Version]
- Cali, J.J.; Russell, D.W. Characterization of human sterol 27-hydroxylase. A mitochondrial cytochrome P-450 that catalyzes multiple oxidation reaction in bile acid biosynthesis. J. Biol. Chem. 1991, 266, 7774–7778. [Google Scholar] [CrossRef]
- Baravalle, R.; Di Nardo, G.; Bandino, A.; Barone, I.; Catalano, S.; Andò, S.; Gilardi, G. Impact of R264C and R264H polymorphisms in human aromatase function. J. Steroid. Biochem. Mol. Biol. 2017, 167, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Catalano, S.; Barone, I.; Giordano, C.; Rizza, P.; Qi, H.; Gu, G.; Malivindi, R.; Bonofiglio, D.; Andò, S. Rapid estradiol/ERα signaling enhances aromatase enzymatic activity in breast cancer cells. Mol. Endocrinol. 2009, 23, 1634–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Harada, N. Post-translational dual regulation of cytochrome P450 aromatase at the catalytic and protein levels by phosphorylation/dephosphorylation. FEBS J. 2014, 281, 4830–4840. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.W.; Shin, I.; Kagawa, N.; Evans, D.B.; Waterman, M.R.; Arteaga, C.L. Aromatase is phosphorylated in situ at serine-118. J. Steroid. Biochem. Mol. Biol. 2008, 112, 95–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, T.; Tateishi, T.; Hatano, M.; Fujii-Kuriyama, Y. Probing the role of lysines and arginines in the catalytic function of cytochrome P450d by site-directed mutagenesis. Interaction with NADPH-cytochrome P450 reductase. J. Biol. Chem. 1991, 266, 3372–3375. [Google Scholar] [CrossRef]
- Shen, S.J.; Strobel, H.W. Role of lysine and arginine residues of cytochrome P450 in the interaction between cytochrome P4502B1 and NADPH-cytochrome P450 reductase. Arch. Biochem. Biophys. 1993, 304, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Šrejber, M.; Navrátilová, V.; Paloncýová, M.; Bazgier, V.; Berka, K.; Anzenbacher, P.; Otyepka, M. Membrane-attached mammalian cytochromes P450: An overview of the membrane’s effects on structure, drug binding, and interactions with redox partners. J. Inorg. Biochem. 2018, 183, 117–136. [Google Scholar] [CrossRef]
- Hong, Y.; Li, H.; Ye, J.; Miki, Y.; Yuan, Y.-C.; Sasano, H.; Evans, D.B.; Chen, S. Epitope characterization of an aromatase monoclonal antibody suitable for the assessment of intratumoral aromatase activity. PLoS ONE 2009, 4, e8050. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Rashid, R.; Chen, S. Binding features of steroidal and nonsteroidal inhibitors. Steroids 2011, 76, 802–806. [Google Scholar] [CrossRef] [Green Version]
- Ritacco, I.; Spinello, A.; Ippoliti, E.; Magistrato, A. Post-translational regulation of CYP450s metabolism as revealed by all-atoms simulations of the aromatase enzyme. J. Chem. Inf. Model. 2019, 59, 2930–2940. [Google Scholar] [CrossRef]
- Ritacco, I.; Saltalamacchia, A.; Spinello, A.; Ippoliti, E.; Magistrato, A. All-atom simulations disclose how cytochrome reductase reshapes the substrate access/egress routes of its partner CYP450s. J. Phys. Chem. Lett. 2020, 11, 1189–1193. [Google Scholar] [CrossRef]
- Yoshimoto, F.K.; Guengerich, F.P. Mechanism of the third oxidative step in the conversion of androgens to estrogens by cytochrome P450 19A1 steroid aromatase. J. Am. Chem. Soc. 2014, 136, 15016–15025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatri, Y.; Luthra, A.; Duggal, R.; Sligar, S.G. Kinetic solvent isotope effect in steady-state turnover by CYP19A1 suggests involvement of Compound 1 for both hydroxylation and aromatization steps. FEBS Lett. 2014, 588, 3117–3122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, J.; Di Nardo, G.; Griswold, J.; Egbuta, C.; Jiang, W.; Gilardi, G.; Ghosh, D. Structural basis for the functional roles of critical residues in human cytochrome P450 aromatase. Biochemistry 2013, 52, 5821–5829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Nardo, G.; Gilardi, G. Human aromatase: Perspectives in biochemistry and biotechnology: Human Aromatase. Biotechnol. Appl. Biochem. 2013, 60, 92–101. [Google Scholar] [CrossRef]
- Parween, S.; Rojas Velazquez, M.N.; Udhane, S.S.; Kagawa, N.; Pandey, A.V. Variability in loss of multiple enzyme activities due to the human genetic variation P284T located in the flexible hinge region of NADPH cytochrome P450 oxidoreductase. Front. Pharmacol. 2019, 10, 1187. [Google Scholar] [CrossRef] [Green Version]
- Hemimi, N.; Shaafie, I.; Alshawa, H. The study of the impact of genetic polymorphism of aromatase (CYP19) enzyme and the susceptibility to polycystic ovary syndrome (575.5). FASEB J. 2014, 28, 575-5. [Google Scholar] [CrossRef]
- Jin, J.L.; Sun, J.; Ge, H.J.; Cao, Y.X.; Wu, X.K.; Liang, F.J.; Sun, H.X.; Ke, L.; Yi, L.; Wu, Z.W.; et al. Association between CYP19 gene SNP rs2414096 polymorphism and polycystic ovary syndrome in Chinese women. BMC. Med. Genet. 2009, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Mehdizadeh, A.; Kalantar, S.M.; Sheikhha, M.H.; Aali, B.S.; Ghanei, A. Association of SNP rs.2414096 CYP19 gene with polycystic ovarian syndrome in Iranian women. Int. J. Reprod. Biomed. 2017, 15, 491–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.M.; Abel, J.; Ko, Y.; Harth, V.; Park, W.Y.; Seo, J.S.; Yoo, K.Y.; Choi, J.Y.; Shin, A.; Ahn, S.H.; et al. Genetic polymorphisms of cytochrome P450 19 and 1B1, alcohol use, and breast cancer risk in Korean women. Br. J. Cancer 2003, 88, 675–678. [Google Scholar] [CrossRef] [Green Version]
- Barone, I.; Giordano, C.; Malivindi, R.; Lanzino, M.; Rizza, P.; Casaburi, I.; Bonofiglio, D.; Catalano, S.; Andò, S. Estrogens and PTP1B function in a novel pathway to regulate aromatase enzymatic activity in breast cancer cells. Endocrinology 2012, 153, 5157–5166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornil, C.A.; Ball, G.F.; Balthazart, J. Functional significance of the rapid regulation of brain estrogen action: Where do the estrogens come from? Brain Res. 2006, 1126, 2–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balthazart, J.; Choleris, E.; Remage-Healey, L. Steroids and the brain: 50 years of research, conceptual shifts and the ascent of non-classical and membrane-initiated actions. Horm. Behav. 2018, 99, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Forlano, P.M.; Deitcher, D.L.; Myers, D.A.; Bass, A.H. Anatomical distribution and cellular basis for high levels of aromatase activity in the brain of teleost fish: Aromatase enzyme and mRNA expression identify glia as source. J. Neurosci. 2001, 21, 8943–8955. [Google Scholar] [CrossRef] [PubMed]
- Diotel, N.; Le Page, Y.; Mouriec, K.; Tong, S.-K.; Pellegrini, E.; Vaillant, C.; Anglade, I.; Brion, F.; Pakdel, F.; Chung, B.-C.; et al. Aromatase in the brain of teleost fish: Expression, regulation and putative functions. Front. Neuroendocrinol. 2010, 31, 172–192. [Google Scholar] [CrossRef] [Green Version]
- The UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [Green Version]
- Ashkenazy, H.; Abadi, S.; Martz, E.; Chay, O.; Mayrose, I.; Pupko, T.; Ben-Tal, N. ConSurf 2016: An improved methodology to estimate and visualize evolutionary conservation in macromolecules. Nucleic Acids Res. 2016, 44, W344–W350. [Google Scholar] [CrossRef] [Green Version]
- Landau, M.; Mayrose, I.; Rosenberg, Y.; Glaser, F.; Martz, E.; Pupko, T.; Ben-Tal, N. ConSurf 2005: The projection of evolutionary conservation scores of residues on protein structures. Nucleic Acids Res. 2005, 33, W299–W302. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Clamp, M.; Cuff, J.; Searle, S.M.; Barton, G.J. The Jalview Java alignment editor. Bioinformatics 2004, 20, 426–427. [Google Scholar] [CrossRef] [Green Version]
- Mayrose, I.; Graur, D.; Ben-Tal, N.; Pupko, T. Comparison of site-specific rate-inference methods for protein sequences: Empirical bayesian methods are superior. Mol. Biol. Evol. 2004, 21, 1781–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoch, G.A.; Yano, J.K.; Sansen, S.; Dansette, P.M.; Stout, C.D.; Johnson, E.F. Determinants of cytochrome p450 2c8 substrate binding: Structures of complexes with montelukast, troglitazone, felodipine, and 9-cis-retinoic acid. J. Biol. Chem. 2008, 283, 17227–17237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera-A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midlik, A.; Navrátilová, V.; Moturu, T.R.; Koča, J.; Svobodová, R.; Berka, K. Uncovering of cytochrome P450 anatomy by SecStrAnnotator. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Gotoh, O. Substrate recognition sites in cytochrome P450 family 2 (CYP2) proteins inferred from comparative analyses of amino acid and coding nucleotide sequences. J. Biol. Chem. 1992, 267, 83–90. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Comparative protein structure modeling using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5.6.1–5.6.37. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-Tasser Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theor. Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [Green Version]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef] [Green Version]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T. Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 1999, 292, 195–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]





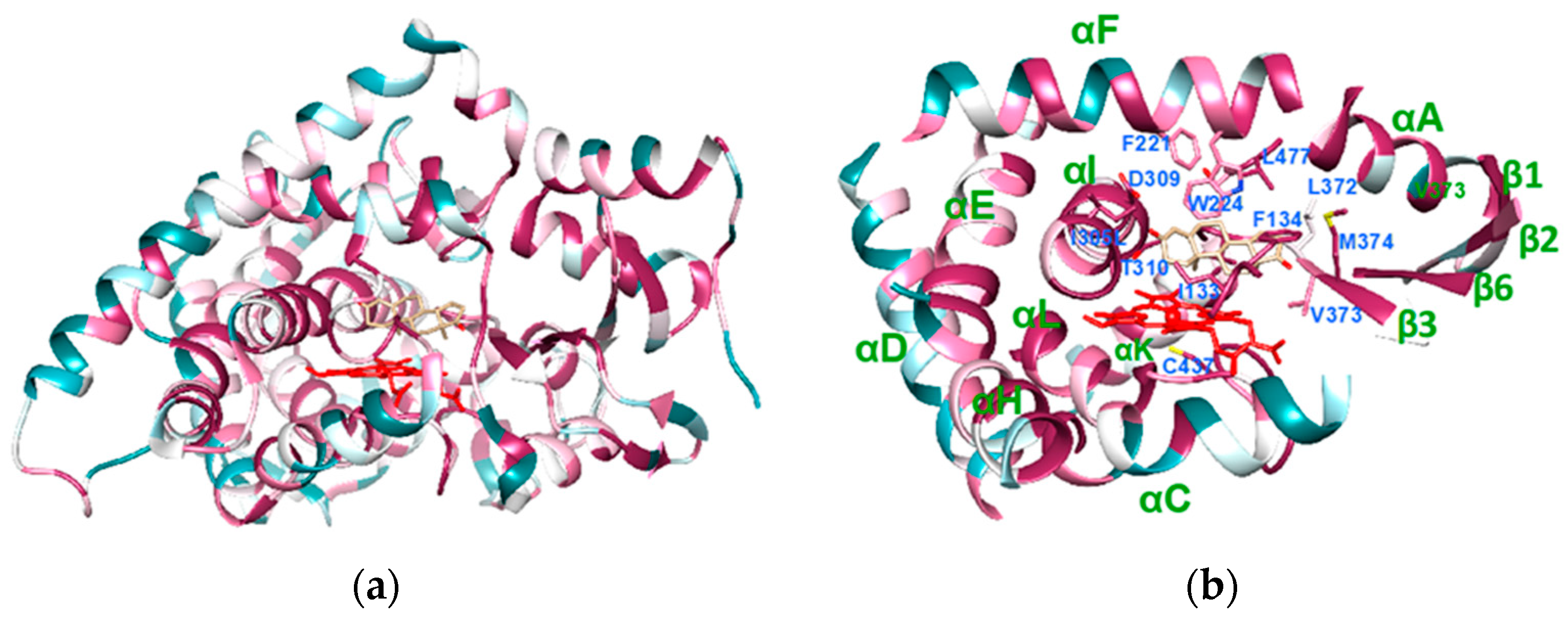{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue | Location | Conservation Score | Notes |
|---|---|---|---|
| C437 | K″-L helix loop | −1.095 | |
| I305 | I-helix | −0.936 | L/V only in invertebrate Branchiostoma |
| A306 | I-helix | −1.002 | T in the mammal Capra hircus |
| D309 | I-helix | −1.058 | Q in CYP19B1 of the fish Halichoeres tenuispinis |
| T310 | I-helix | −1.011 | I in the fish Maylandia zebra |
| F221 | F-helix | −0.805 | |
| W224 | F-helix | −0.896 | |
| I133 | B-C loop | −1.038 | M in pig aromatase isoform 3 |
| F134 | B-C loop | −1.073 | |
| V370 | K-helix—β3 loop | −1.001 | |
| L372 | K-helix—β3 loop | −0.202 | Phe in fishes |
| V373 | K-helix—β3 loop | −0.583 | S/ T in most fishes and in CYP19A1 of zebrafish and goldfish |
| M374 | β3 | −1.031 | |
| L477 | β8–β9 loop | −1.011 | |
| S478 | β8–β9 loop | −0.828 | A in many sequences, starting from mammals to amphibians. S in fishes. |
| R192 | Helix E | −0.974 | C or H in some mammals, birds and fishes, including the two isoforms of zebrafish |
| E483 | β9–β10 loop | −0.761 | Conserved in the two isoforms of zebrafish and goldfish |
| Residue | Conservation Score | Notes |
|---|---|---|
| K99 | 0.287 | R in most fishes and Branchiostoma floridae, not conserved in 1 amphibian, 1 reptile, in 15% of fishes and Branchiostoma belcheri (S) |
| K108 | −0.024 | Always substituted by R |
| R145 | −0.972 | Well conserved |
| K352 | 1.293 | Conserved only in mammals |
| K389 | 0.767 | Not conserved in invertebrates (P) and 70% of fishes (including only isoform CYP19A1 in zebrafish) |
| K390 | −0.231 | K or R |
| K420 | 0.472 | Not conserved in two mammals, 20% of fishes (including CYP19B1 of zebrafish) and E in invertebrates |
| R425 | −0.881 | Well conserved with some exceptions in fishes and the invertebrates (T) |
| K440 | −0.897 | R in invertebrates |
| S153 | −0.533 | T in invertebrates and most fishes |
| Q351 | 0.799 | Conserved in 90% of mammals |
| Y424 | 0.308 | Conserved in mammals |
| Y441 | −0.553 | Conserved in mammals and amphibians, H in 97% of fishes and T in invertebrates |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Nardo, G.; Zhang, C.; Marcelli, A.G.; Gilardi, G. Molecular and Structural Evolution of Cytochrome P450 Aromatase. Int. J. Mol. Sci. 2021, 22, 631. https://doi.org/10.3390/ijms22020631
Di Nardo G, Zhang C, Marcelli AG, Gilardi G. Molecular and Structural Evolution of Cytochrome P450 Aromatase. International Journal of Molecular Sciences. 2021; 22(2):631. https://doi.org/10.3390/ijms22020631
Chicago/Turabian StyleDi Nardo, Giovanna, Chao Zhang, Anna Giulia Marcelli, and Gianfranco Gilardi. 2021. "Molecular and Structural Evolution of Cytochrome P450 Aromatase" International Journal of Molecular Sciences 22, no. 2: 631. https://doi.org/10.3390/ijms22020631
APA StyleDi Nardo, G., Zhang, C., Marcelli, A. G., & Gilardi, G. (2021). Molecular and Structural Evolution of Cytochrome P450 Aromatase. International Journal of Molecular Sciences, 22(2), 631. https://doi.org/10.3390/ijms22020631