Varied Expression of Senescence-Associated and Ethylene-Related Genes during Postharvest Storage of Brassica Vegetables



Abstract
:1. Introduction
2. Results
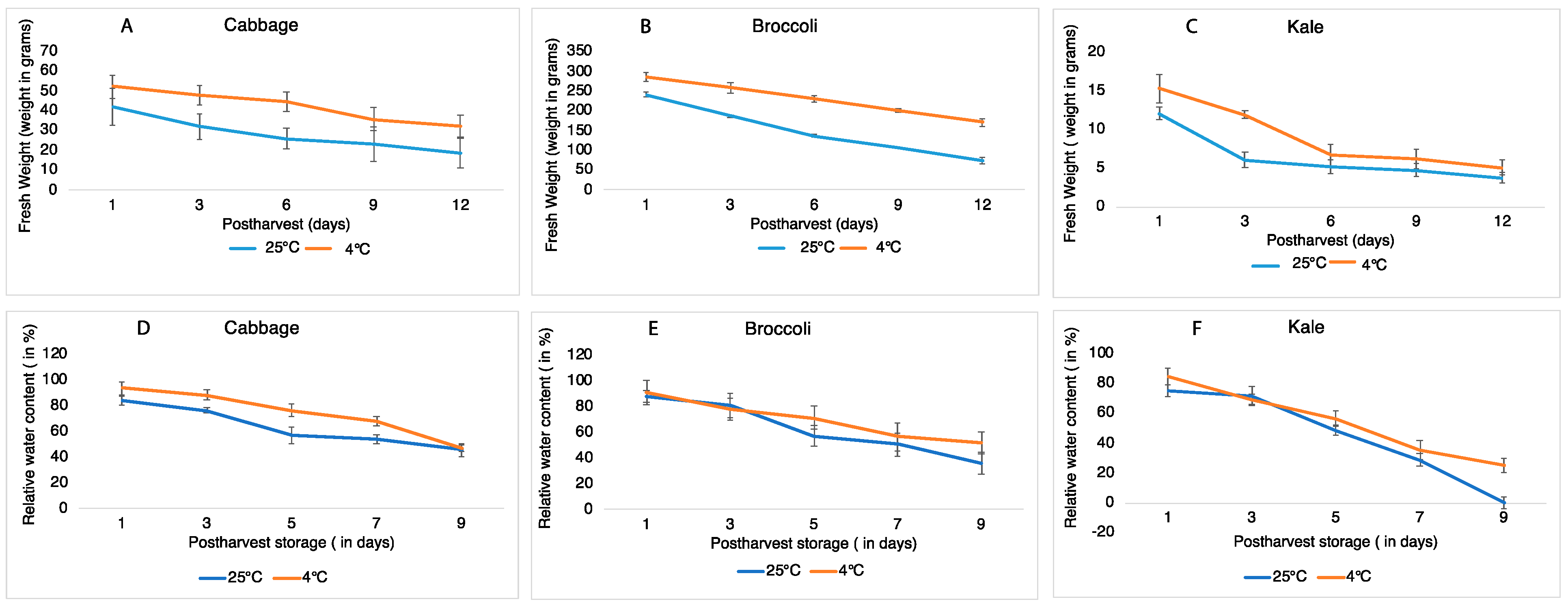
2.1. Effect of Storage Temperatures on the Fresh Weight and Relative Water Content of Brassica Vegetables
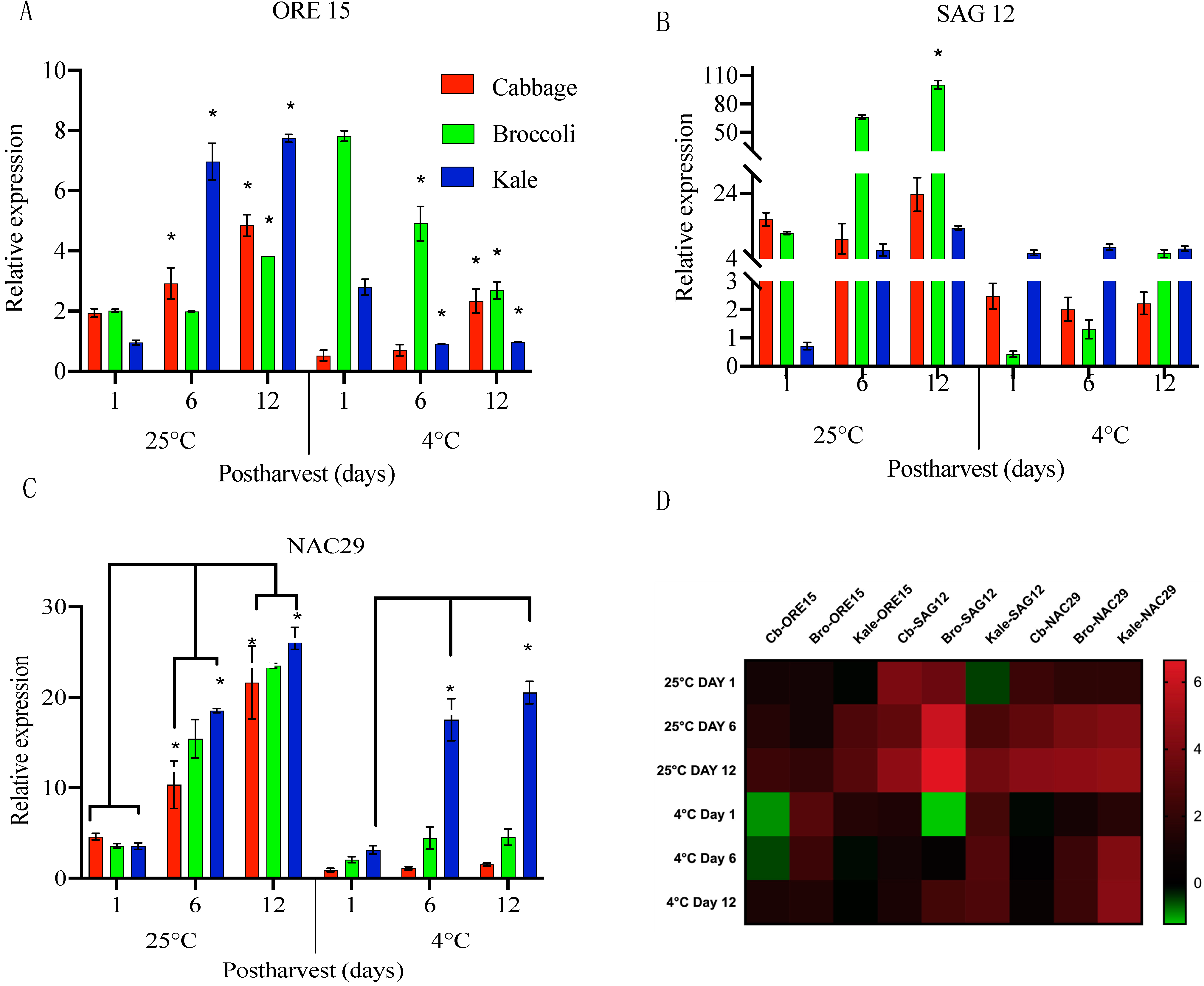
2.2. The Expression Levels of Specific Senescence-Associated Genes (SAGs) Were Highly Induced in Postharvest Cabbage, Broccoli and Kale While Stored at Room Temperature
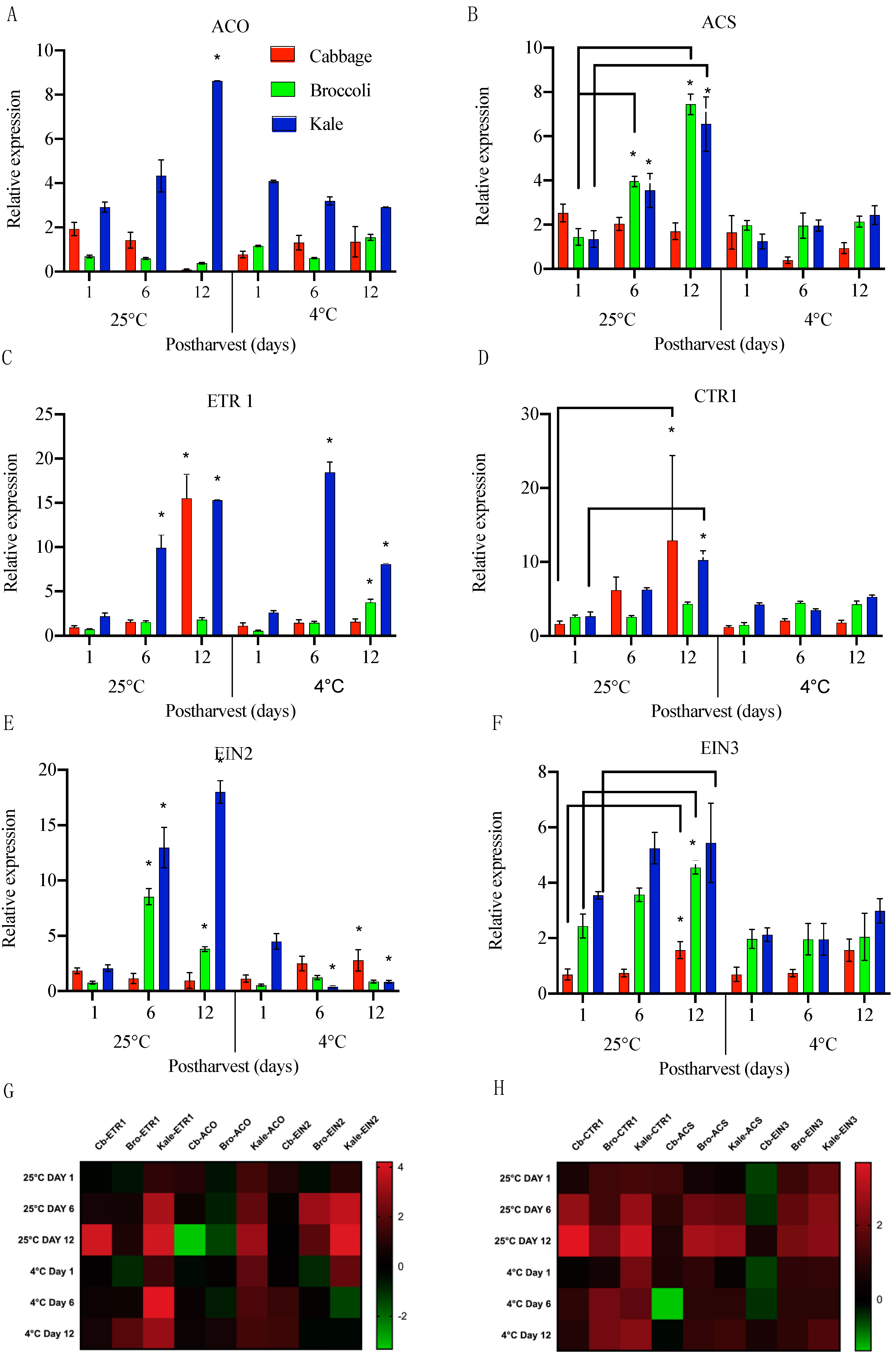
2.3. Expression of Ethylene-Related Genes in Harvested Cabbage, Broccoli and Kale during Storage
3. Discussion
4. Materials and Methods
4.1. Collection of Plant Materials
4.2. Measurement of Moisture Content
4.3. Measurement of Relative Water Content in Cabbage, Broccoli and Kale
4.4. RNA Extraction and cDNA Synthesis
4.5. Gene Expression Studies by RT-qPCR
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Céline, M.; Valérie, G.; Karine, G.; Sandrine, C.; Nathalie, G.; Stéphane, G.; Sébastien, G. Consumer behaviour in the prediction of postharvest losses reduction for fresh strawberries packed in modified atmosphere packaging. Postharvest Biol. Technol. 2020, 163, 111119. [Google Scholar] [CrossRef]
- Agarwal, G.; Choudhary, D.; Singh, V.P.; Arora, A. Role of ethylene receptors during senescence and ripening in horticultural crops. Plant Signal. Behav. 2012, 7, 827–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, M.S.; Chong, C.; Landry, B.S.; Crête, R. Cabbage. In Genetic Improvement of Vegetable Crops; Elsevier: Amsterdam, The Netherlands, 1993; pp. 113–155. ISBN 978-0-08-040826-2. [Google Scholar]
- Ramirez, D.; Abellán-Victorio, A.; Beretta, V.; Camargo, A.; Moreno, D.A. Functional Ingredients From Brassicaceae Species: Overview and Perspectives. IJMS 2020, 21, 1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manchali, S.; Chidambara Murthy, K.N.; Patil, B.S. Crucial facts about health benefits of popular cruciferous vegetables. J. Funct. Foods 2012, 4, 94–106. [Google Scholar] [CrossRef]
- Woo, H.R.; Kim, H.J.; Lim, P.O.; Nam, H.G. Leaf Senescence: Systems and Dynamics Aspects. Annu. Rev. Plant Biol. 2019, 70, 347–376. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Fan, Z.; Shan, W.; Yin, X.; Kuang, J.; Lu, W.; Chen, J. Association of BrERF72 with methyl jasmonate-induced leaf senescence of Chinese flowering cabbage through activating JA biosynthesis-related genes. Hortic. Res. 2018, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Ombra, M.N.; Cozzolino, A.; Nazzaro, F.; d’Acierno, A.; Tremonte, P.; Coppola, R.; Fratianni, F. Biochemical and biological characterization of two Brassicaceae after their commercial expiry date. Food Chem. 2017, 218, 335–340. [Google Scholar] [CrossRef]
- Christiansen, M.W.; Gregersen, P.L. Members of the barley NAC transcription factor gene family show differential co-regulation with senescence-associated genes during senescence of flag leaves. J. Exp. Bot. 2014, 65, 4009–4022. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Ecker, J.R. The ethylene signaling pathway: New insights. Curr. Opin. Plant Biol. 2004, 7, 40–49. [Google Scholar] [CrossRef]
- Kim, H.J.; Hong, S.H.; Kim, Y.W.; Lee, I.H.; Jun, J.H.; Phee, B.-K.; Rupak, T.; Jeong, H.; Lee, Y.; Hong, B.S.; et al. Gene regulatory cascade of senescence-associated NAC transcription factors activated by ETHYLENE-INSENSITIVE2-mediated leaf senescence signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 4023–4036. [Google Scholar] [CrossRef] [Green Version]
- Balazadeh, S.; Kwasniewski, M.; Caldana, C.; Mehrnia, M.; Zanor, M.I.; Xue, G.-P.; Mueller-Roeber, B. ORS1, an H2O2-Responsive NAC Transcription Factor, Controls Senescence in Arabidopsis thaliana. Mol. Plant 2011, 4, 346–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauf, M.; Arif, M.; Dortay, H.; Matallana-Ramírez, L.P.; Waters, M.T.; Gil Nam, H.; Lim, P.; Mueller-Roeber, B.; Balazadeh, S. ORE1 balances leaf senescence against maintenance by antagonizing G2-like-mediated transcription. EMBO Rep. 2013, 14, 382–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Kim, J.; Jun, S.E.; Park, S.; Timilsina, R.; Kwon, D.S.; Kim, Y.; Park, S.-J.; Hwang, J.Y.; Nam, H.G.; et al. ORESARA15, a PLATZ transcription factor, mediates leaf growth and senescence in Arabidopsis. New Phytol. 2018, 220, 609–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, S.E.; Kim, J.H.; Hwang, J.Y.; Huynh Le, T.T.; Kim, G.-T. ORESARA15 Acts Synergistically with ANGUSTIFOLIA3 and Separately from AINTEGUMENTA to Promote Cell Proliferation during Leaf Growth. IJMS 2019, 21, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCabe, M.S.; Garratt, L.C.; Schepers, F.; Jordi, W.J.R.M.; Stoopen, G.M.; Davelaar, E.; van Rhijn, J.H.A.; Power, J.B.; Davey, M.R. Effects of P SAG12-IPT Gene Expression on Development and Senescence in Transgenic Lettuce. Plant Physiol. 2001, 127, 505–516. [Google Scholar] [CrossRef]
- Grbic, V.; Bleecker, A.B. Ethylene regulates the timing of leaf senescence in Arabidopsis. Plant J. 1995, 8, 595–602. [Google Scholar] [CrossRef]
- Rasori, A.; Ruperti, B.; Bonghi, C.; Tonutti, P.; Ramina, A. Characterization of two putative ethylene receptor genes expressed during peach fruit development and abscission. J. Exp. Bot. 2002, 53, 2333–2339. [Google Scholar] [CrossRef] [Green Version]
- Leclercq, J. LeCTR1, a Tomato CTR1-Like Gene, Demonstrates Ethylene Signaling Ability in Arabidopsis and Novel Expression Patterns in Tomato. Plant Physiol. 2002, 130, 1132–1142. [Google Scholar] [CrossRef] [Green Version]
- Trainotti, L.; Pavanello, A.; Casadoro, G. Different ethylene receptors show an increased expression during the ripening of strawberries: Does such an increment imply a role for ethylene in the ripening of these non-climacteric fruits? J. Exp. Bot. 2005, 56, 2037–2046. [Google Scholar] [CrossRef] [Green Version]
- Shakeel, S.N.; Gao, Z.; Amir, M.; Chen, Y.-F.; Rai, M.I.; Haq, N.U.; Schaller, G.E. Ethylene Regulates Levels of Ethylene Receptor/CTR1 Signaling Complexes in Arabidopsis thaliana. J. Biol. Chem. 2015, 290, 12415–12424. [Google Scholar] [CrossRef] [Green Version]
- Lim, P.O.; Kim, H.J.; Gil Nam, H. Leaf Senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, K.M.; Hobson, G.E.; Grierson, D. Differential Effect of Silver Ions on the Accumulation of Ripening-related mRNAs in Tomato Fruit. J. Plant Physiol. 1990, 135, 708–713. [Google Scholar] [CrossRef]
- Jayas, D.S.; Jeyamkondan, S. PH—Postharvest Technology. Biosyst. Eng. 2002, 82, 235–251. [Google Scholar] [CrossRef]
- Miret, J.A.; Munné-Bosch, S.; Dijkwel, P.P. ABA signalling manipulation suppresses senescence of a leafy vegetable stored at room temperature. Plant Biotechnol. J. 2018, 16, 530–544. [Google Scholar] [CrossRef] [Green Version]
- Barrs, H.; Weatherley, P. A Re-Examination of the Relative Turgidity Technique for Estimating Water Deficits in Leaves. Aust. J. Bio. Sci. 1962, 15, 413. [Google Scholar] [CrossRef] [Green Version]
- Houben, M.; Van de Poel, B. 1-Aminocyclopropane-1-Carboxylic Acid Oxidase (ACO): The Enzyme That Makes the Plant Hormone Ethylene. Front. Plant Sci. 2019, 10, 695. [Google Scholar] [CrossRef] [Green Version]
- Zacarias, L.; Reid, M.S. Role of growth regulators in the senescence of Arabidopsis thaliana leaves. Physiol. Plant 1990, 80, 549–554. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SAG12-RT-FP | TGGAATTGAAGGAGGTGGTTT |
| SAG12-RT-RP | GCCGTATCCAATCGCAGTTA |
| EIN2-RT-FP | GTGCGTCTTATGGTCGGTTA |
| EIN2-RT-RP | CTCTGTAGCCTCCTCTTGATTG |
| ACTIN2-RT-FP | CTTGCACCAAGCAGCATGAA |
| ACTIN2-RT-RP | CCGATCCAGACACTGTACTTCCTT |
| EIN3-RT-FP | AAGGGAGTGGTGGAAAGATAAG |
| EIN3-RT-RP | GTCGGTCCAATCGGGTTATT |
| CTR1-RT-FP | AAATCCGGCTCAGGTTGTAG |
| CTR1-RT-FP | GGTCCAACAACCCTCGATTAT |
| ETR1-RT-FP | TGAGTTCAAACGAGGAGTGTC |
| ETR1-RT-FP | CGGAGAGCGATTTGGTAGTT |
| ACO_FP_RT | AGATGATGGAAGTGATGGATGAG |
| ACO_RP_RT | GATGAGGACAAGGAGGGTAATG |
| ACS_FP_RT | CACGAGACGGTTGCTTTCT |
| ACS_RP_RT | CCGGTTCTCCATCTCAAATCTC |
| NAC29_FP | GCAGAGAGAAGAACTGAAGAAGAG |
| NAC29_RP | AGAGACGGGTCCCATGTAAT |
| ORE15_FP | CATCGCTCTCATCCTCTTCTTC |
| ORE15_RP | AGCTTCTCAAGATCACTCAACC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahlawat, Y.; Liu, T. Varied Expression of Senescence-Associated and Ethylene-Related Genes during Postharvest Storage of Brassica Vegetables. Int. J. Mol. Sci. 2021, 22, 839. https://doi.org/10.3390/ijms22020839
Ahlawat Y, Liu T. Varied Expression of Senescence-Associated and Ethylene-Related Genes during Postharvest Storage of Brassica Vegetables. International Journal of Molecular Sciences. 2021; 22(2):839. https://doi.org/10.3390/ijms22020839
Chicago/Turabian StyleAhlawat, Yogesh, and Tie Liu. 2021. "Varied Expression of Senescence-Associated and Ethylene-Related Genes during Postharvest Storage of Brassica Vegetables" International Journal of Molecular Sciences 22, no. 2: 839. https://doi.org/10.3390/ijms22020839
APA StyleAhlawat, Y., & Liu, T. (2021). Varied Expression of Senescence-Associated and Ethylene-Related Genes during Postharvest Storage of Brassica Vegetables. International Journal of Molecular Sciences, 22(2), 839. https://doi.org/10.3390/ijms22020839