
Ultramicronized Palmitoylethanolamide in the Management of Sepsis-Induced Coagulopathy and Disseminated Intravascular Coagulation

 , ,
, ,  ,
,  , ,
, ,  ,
,  ,
,  , ,
, ,  and
and 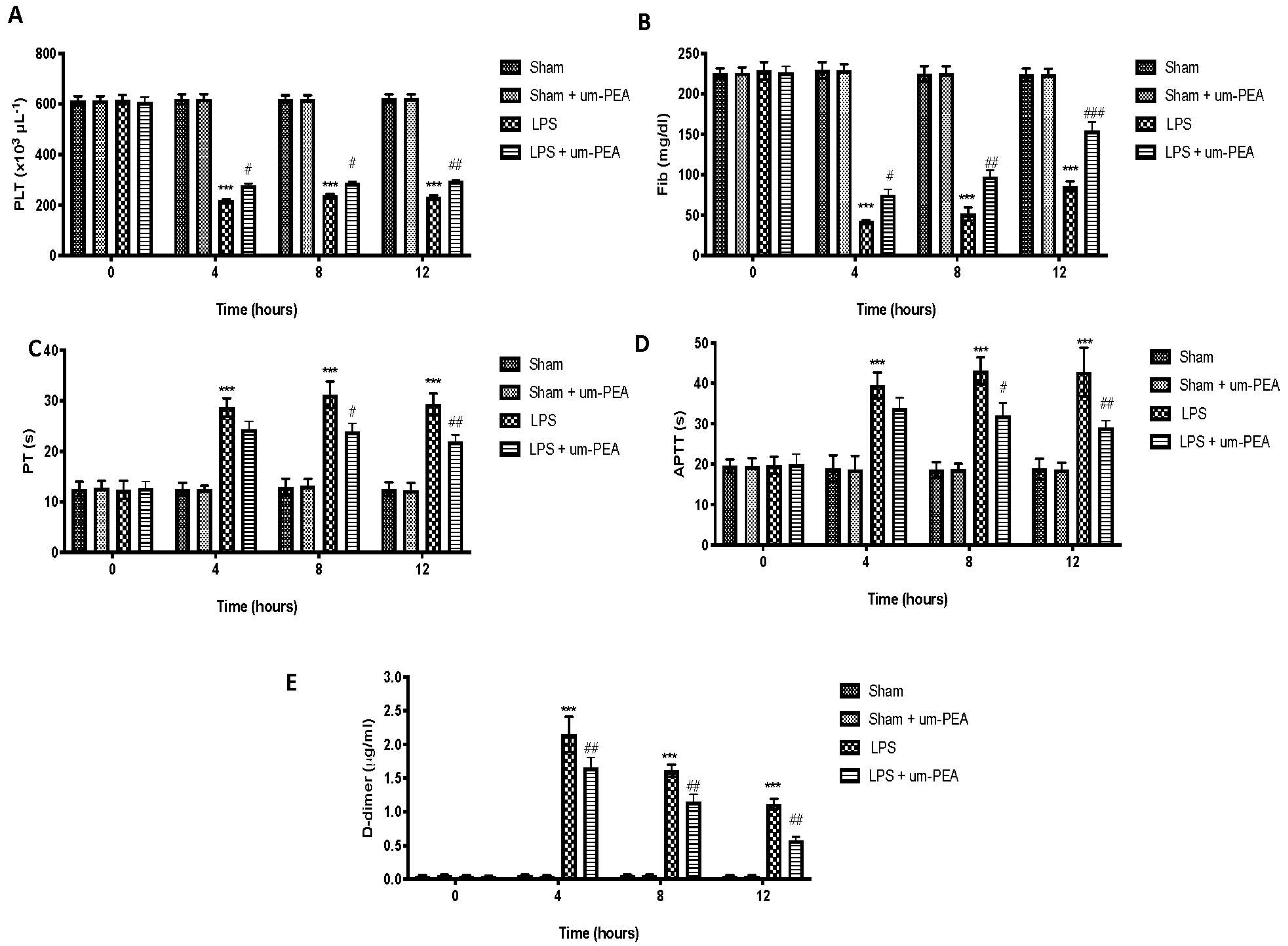{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of um-PEA on Blood Coagulation Parameters
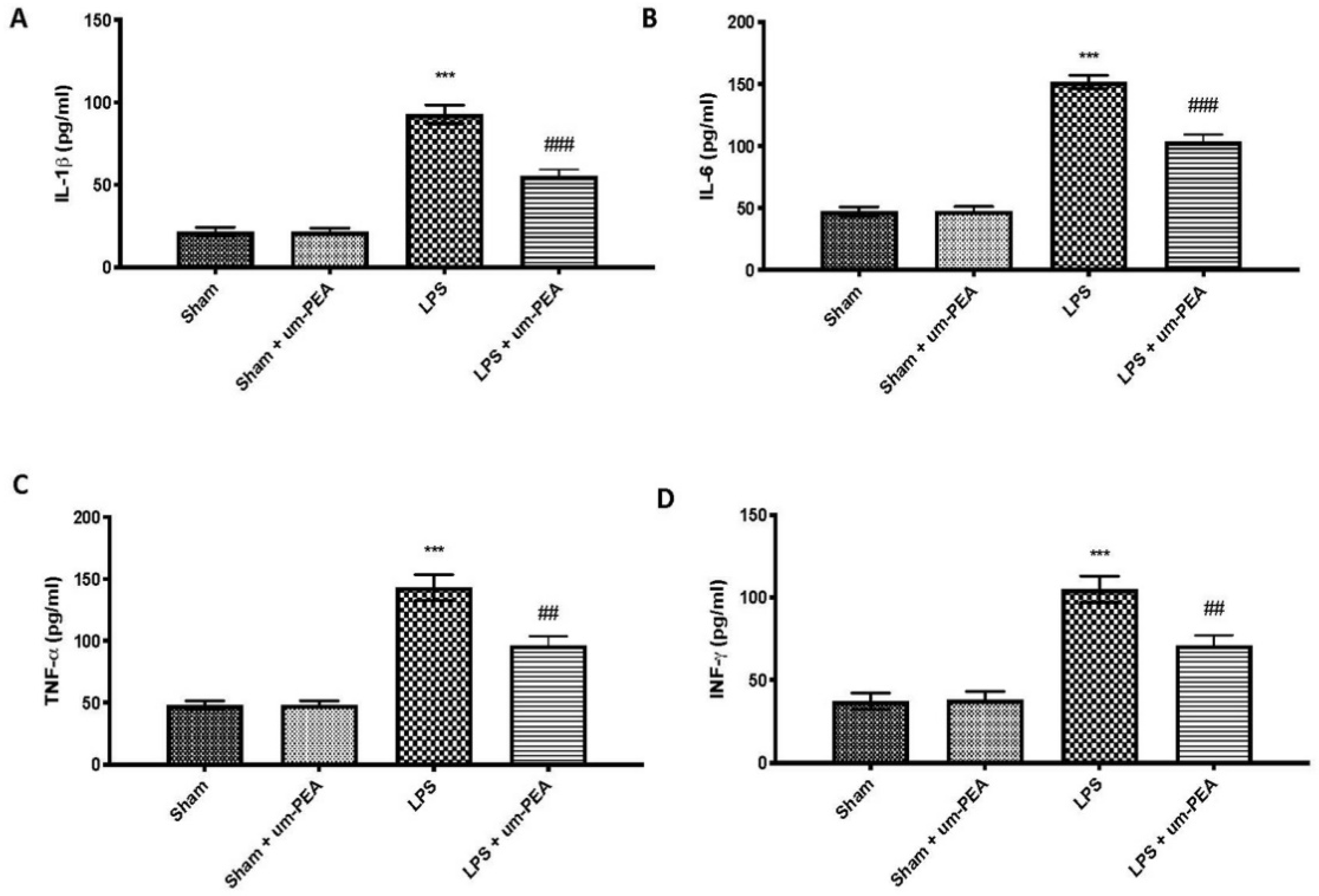
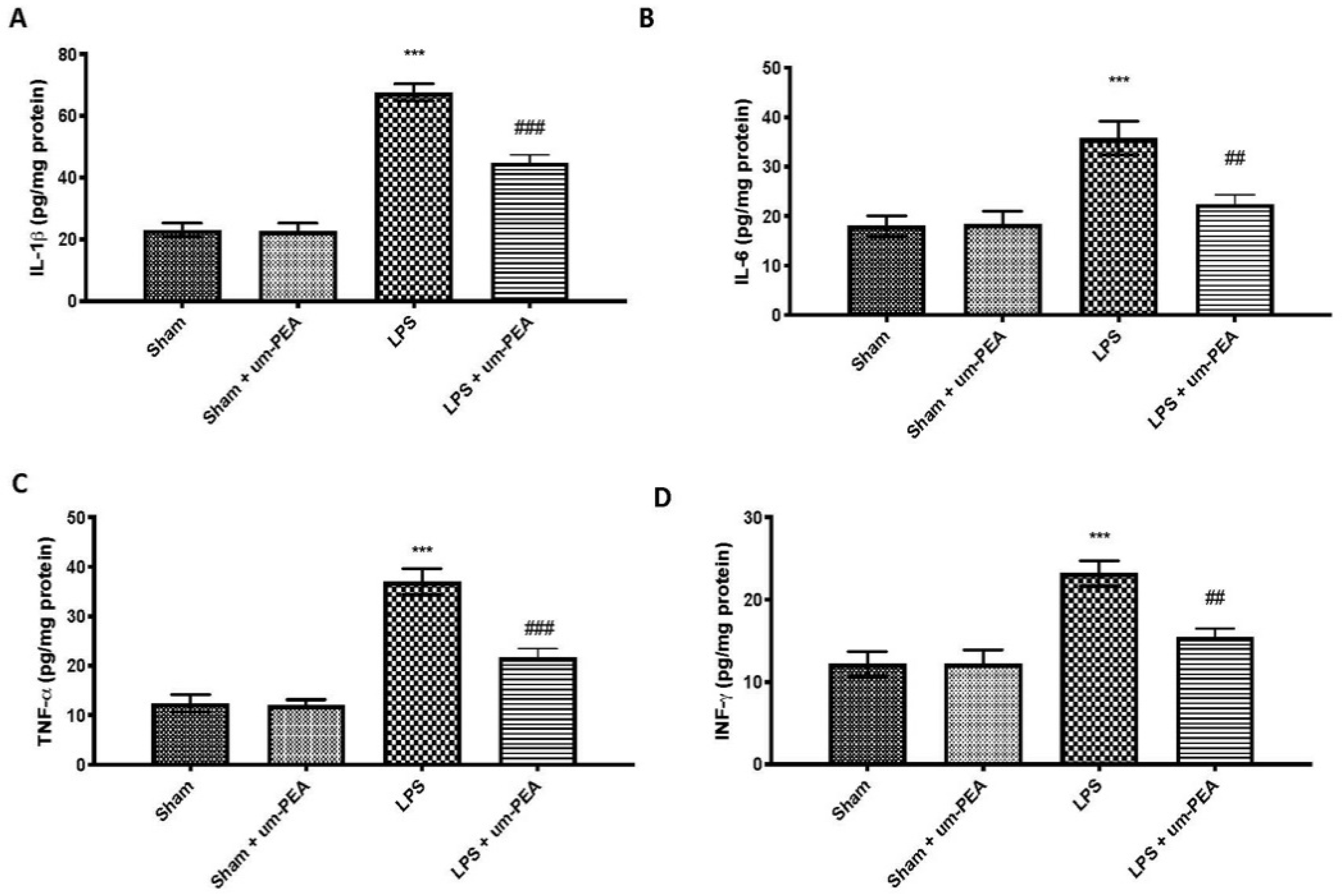
2.2. Effects of um-PEA on Proinflammatory Cytokine Levels
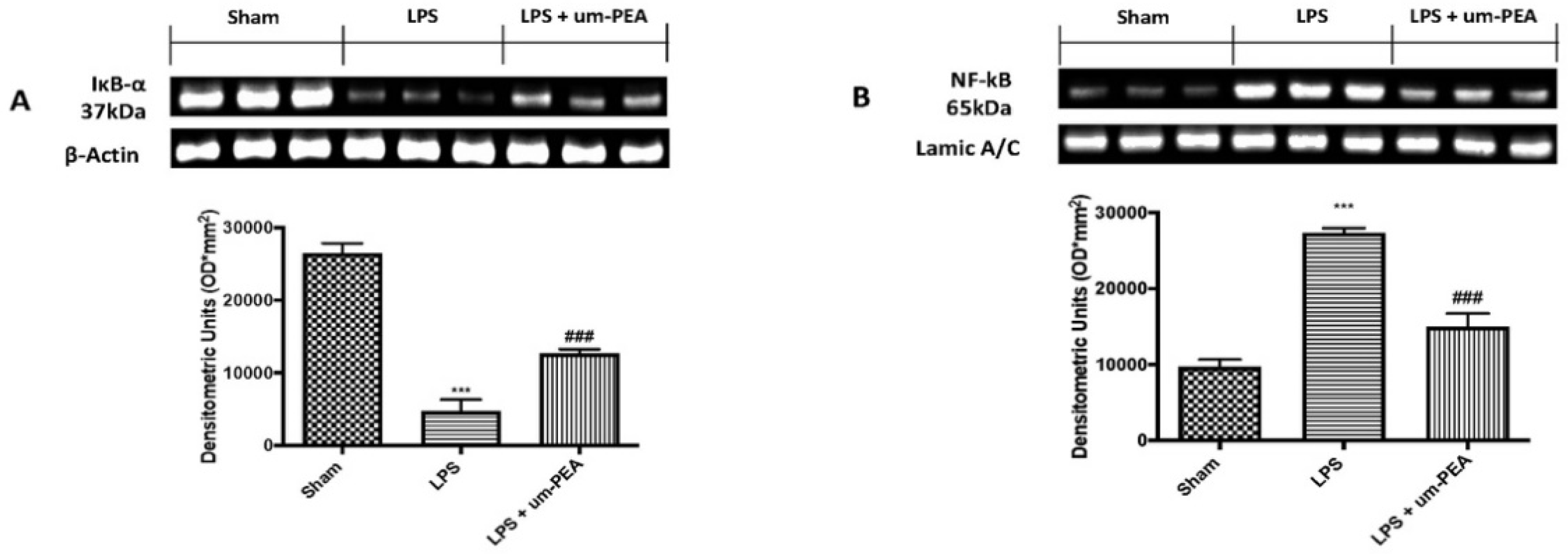
2.3. Effects of um-PEA on Inflammatory Pathway
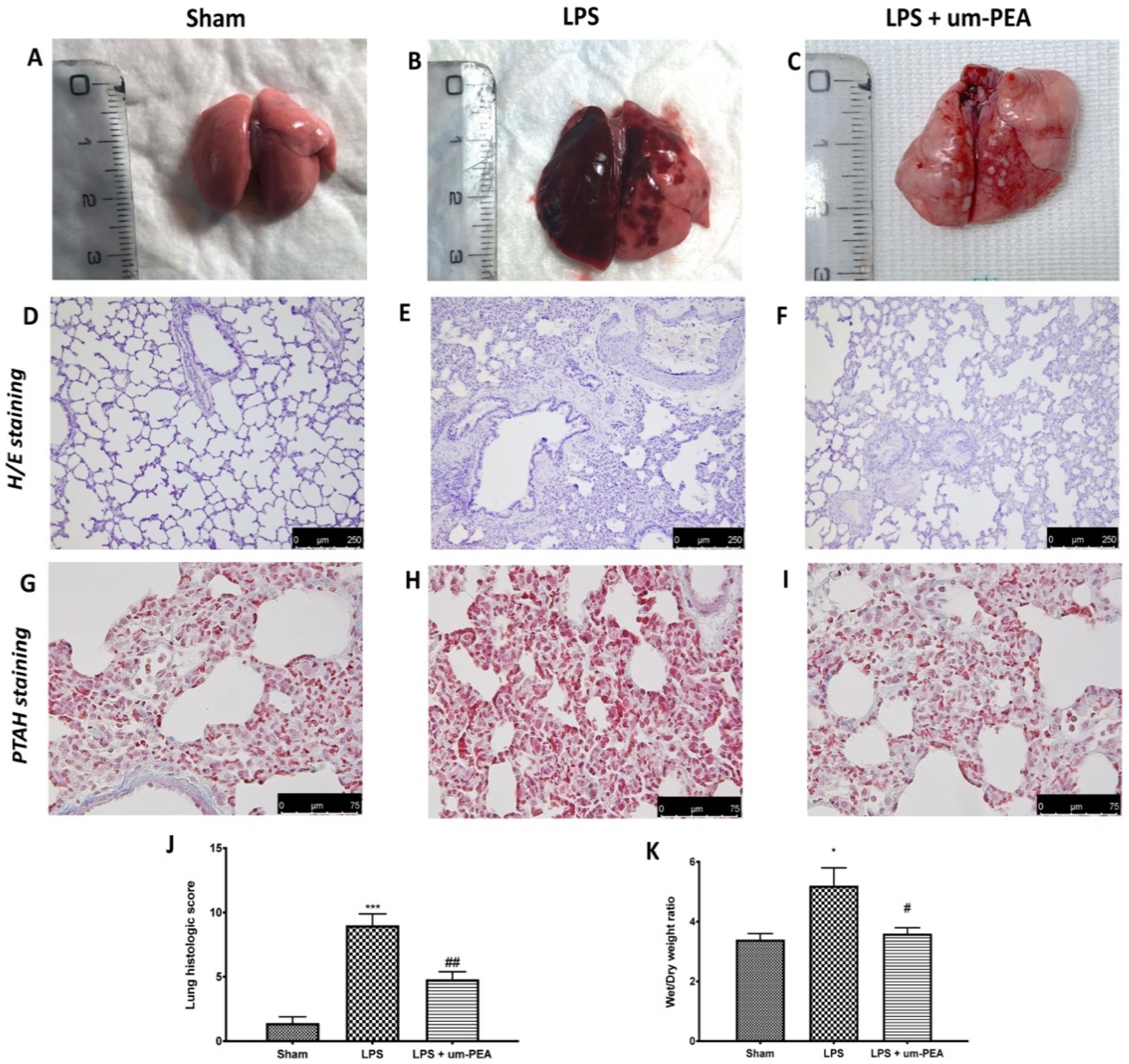
2.4. Effect of um-PEA on the Histological Changes
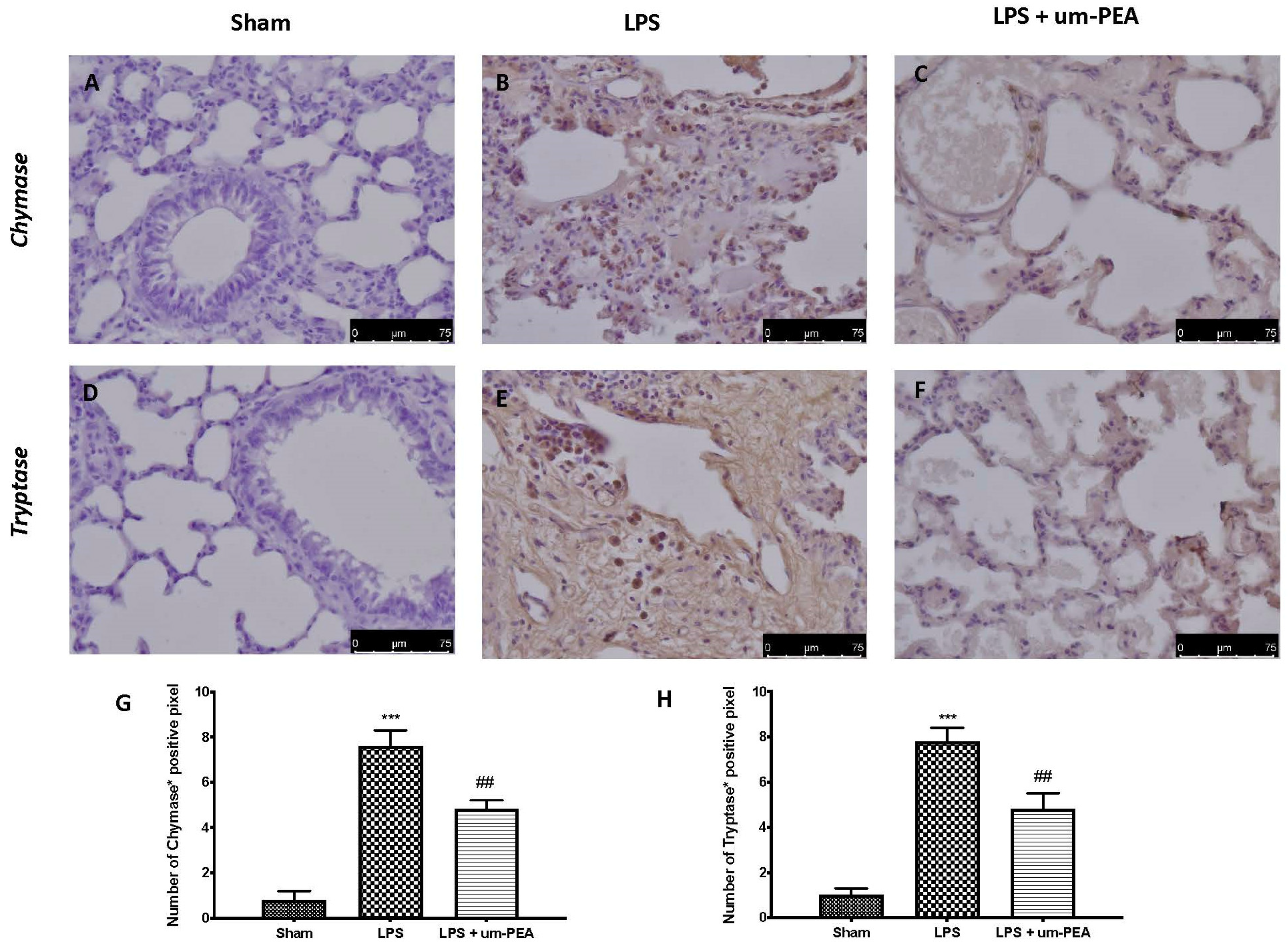
2.5. Effect of um-PEA on MC Activation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Induction of DIC
4.3. Experimental Groups
- LPS group: rats received infusion of LPS, as previously described;
- LPS + um-PEA group: rats received infusion of LPS, as previously described, and were treated with um-PEA (30 mg/kg) by oral gavage 30 min before and 1 h after the start of intravenous infusion of LPS;
- Sham group: identical to the LPS group, but animals received saline (0.9% w/v) instead of LPS;
- Sham + um-PEA group: identical to the sham group, but rats were treated with um-PEA (30 mg/Kg) by oral gavage 30 min before and 1 h after the start of experiment.
4.4. Blood Sampling and Parameter Measurement
4.5. Cytokine Levels
4.6. Western Blot Analysis
4.7. Histopathology
4.8. Measurement of Lung Edema
4.9. Staining of Mast Cells (MCs)
4.10. Immunohistochemical Analysis
4.11. Statistical Evaluation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fang, Y.; Wu, L.C.; Ma, K.; Pan, G.; Yang, S.; Zheng, Y.; Li, Y. Paeoniflorin alleviates lipopolysaccharide-induced disseminated intravascular coagulation by inhibiting inflammation and coagulation activation. Drug Dev. Res. 2020, 81, 517–525. [Google Scholar] [CrossRef]
- Oahin, A.; Ozer, N. The effect of activated protein C in the experimental disseminated intravascular coagulation model formed by lipopolysaccharide infusion. Acta Cir. Bras. 2020, 35, e351102. [Google Scholar] [CrossRef]
- McGonagle, D.; O’Donnell, J.S.; Sharif, K.; Emery, P.; Bridgewood, C. Immune mechanisms of pulmonary intravascular coagulopathy in COVID-19 pneumonia. Lancet Rheumatol. 2020, 2, e437–e445. [Google Scholar] [CrossRef]
- Cucinotta, D.; Vanelli, M. WHO Declares COVID-19 a Pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar] [CrossRef]
- Tang, X.M.; Liao, Z.K.; Huang, Y.W.; Lin, X.; Wu, L.C. Atractylenolide protects against lipopolysaccharide-induced disseminated intravascular coagulation by anti-inflammatory and anticoagulation effect. Asian Pac. J. Trop. Med. 2017, 10, 582–587. [Google Scholar] [CrossRef]
- Levi, M. Diagnosis and treatment of disseminated intravascular coagulation. Int. J. Lab. Hematol. 2014, 36, 228–236. [Google Scholar] [CrossRef]
- Jin, Y.; Yu, G.; Peng, P.; Zhang, Y.; Xin, X. Down-regulated expression of AQP5 on lung in rat DIC model induced by LPS and its effect on the development of pulmonary edema. Pulm. Pharmacol. Ther. 2013, 26, 661–665. [Google Scholar] [CrossRef]
- Asakura, H. Classifying types of disseminated intravascular coagulation: Clinical and animal models. J. Intensive Care 2014, 2, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ware, L.B. Physiological and biological heterogeneity in COVID-19-associated acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 1163–1165. [Google Scholar] [CrossRef]
- Brooks, M.B.; Turk, J.R.; Guerrero, A.; Narayanan, P.K.; Nolan, J.P.; Besteman, E.G.; Wilson, D.W.; Thomas, R.A.; Fishman, C.E.; Thompson, K.L.; et al. Non-Lethal Endotoxin Injection: A Rat Model of Hypercoagulability. PLoS ONE 2017, 12, e0169976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazar, E.; Bulbul, A.; Avci, G.E.; Er, A.; Uney, K.; Elmas, M.; Tras, B. Effects of enrofloxacin, flunixin meglumine and dexamethasone on disseminated intravascular coagulation, cytokine levels and adenosine deaminase activity in endotoxaemia in rats. Acta Vet. Hung. 2010, 58, 357–367. [Google Scholar] [CrossRef]
- Zeerleder, S.; Hack, C.E.; Wuillemin, W.A. Disseminated intravascular coagulation in sepsis. Chest 2005, 128, 2864–2875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cicala, C.; Cirino, G. Linkage between inflammation and coagulation: An update on the molecular basis of the crosstalk. Life Sci. 1998, 62, 1817–1824. [Google Scholar] [CrossRef]
- Levi, M.; Scully, M. How I treat disseminated intravascular coagulation. Blood 2018, 131, 845–854. [Google Scholar] [CrossRef]
- Iba, T.; Yamada, A.; Hashiguchi, N.; Nagaoka, I. New therapeutic options for patients with sepsis and disseminated intravascular coagulation. Pol. Arch. Med. Wewn. 2014, 124, 321–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, H.; Japanese Society of Thrombosis Hemostasis/DIC subcommittee; Okamoto, K.; Iba, T.; Kushimoto, S.; Kawasugi, K.; Gando, S.; Madoiwa, S.; Uchiyama, T.; Mayumi, T.; et al. Addition of recommendations for the use of recombinant human thrombomodulin to the “Expert consensus for the treatment of disseminated intravascular coagulation in Japan”. Thromb. Res. 2014, 134, 924–925. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Thachil, J.; Asakura, H.; Levy, J.H.; Iba, T. Thrombomodulin in disseminated intravascular coagulation and other critical conditions-a multi-faceted anticoagulant protein with therapeutic potential. Crit. Care 2019, 23, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordaro, M.; Scuto, M.; Siracusa, R.; D’Amico, R.; Filippo Peritore, A.; Gugliandolo, E.; Fusco, R.; Crupi, R.; Impellizzeri, D.; Pozzebon, M.; et al. Effect of N-palmitoylethanolamine-oxazoline on comorbid neuropsychiatric disturbance associated with inflammatory bowel disease. FASEB J. 2020, 34, 4085–4106. [Google Scholar] [CrossRef]
- D’Amico, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. ALIAmides Update: Palmitoylethanolamide and Its Formulations on Management of Peripheral Neuropathic Pain. Int. J. Mol. Sci. 2020, 21, 5330. [Google Scholar] [CrossRef] [PubMed]
- Noce, A.; Albanese, M.; Marrone, G.; Di Lauro, M.; Pietroboni Zaitseva, A.; Palazzetti, D.; Guerriero, C.; Paolino, A.; Pizzenti, G.; Di Daniele, F.; et al. Ultramicronized Palmitoylethanolamide (um-PEA): A New Possible Adjuvant Treatment in COVID-19 patients. Pharmaceuticals 2021, 14, 336. [Google Scholar] [CrossRef] [PubMed]
- Pesce, M.; Seguella, L.; Cassarano, S.; Aurino, L.; Sanseverino, W.; Lu, J.; Corpetti, C.; Del Re, A.; Vincenzi, M.; Sarnelli, G.; et al. Phytotherapics in COVID19: Why palmitoylethanolamide? Phytother. Res. 2021, 35, 2514–2522. [Google Scholar] [CrossRef]
- Levi, M.; Sivapalaratnam, S. Disseminated intravascular coagulation: An update on pathogenesis and diagnosis. Expert Rev. Hematol. 2018, 11, 663–672. [Google Scholar] [CrossRef]
- Papageorgiou, C.; Jourdi, G.; Adjambri, E.; Walborn, A.; Patel, P.; Fareed, J.; Elalamy, I.; Hoppensteadt, D.; Gerotziafas, G.T. Disseminated Intravascular Coagulation: An Update on Pathogenesis, Diagnosis, and Therapeutic Strategies. Clin. Appl. Thromb. Hemost. 2018, 24, 8S–28S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi, M.; Keller, T.T.; van Gorp, E.; ten Cate, H. Infection and inflammation and the coagulation system. Cardiovasc. Res. 2003, 60, 26–39. [Google Scholar] [CrossRef]
- van der Poll, T.; Levi, M. Crosstalk between inflammation and coagulation: The lessons of sepsis. Curr. Vasc. Pharmacol. 2012, 10, 632–638. [Google Scholar] [CrossRef]
- Lazzaroni, M.G.; Piantoni, S.; Masneri, S.; Garrafa, E.; Martini, G.; Tincani, A.; Andreoli, L.; Franceschini, F. Coagulation dysfunction in COVID-19: The interplay between inflammation, viral infection and the coagulation system. Blood Rev. 2021, 46, 100745. [Google Scholar] [CrossRef] [PubMed]
- Berthelsen, L.O.; Kristensen, A.T.; Tranholm, M. Animal models of DIC and their relevance to human DIC: A systematic review. Thromb. Res. 2011, 128, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Mangalmurti, N.; Hunter, C.A. Cytokine Storms: Understanding COVID-19. Immunity 2020, 53, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Mannino, F.; Bitto, A.; Irrera, N. Severe Acute Respiratory Syndrome Coronavirus-2 Induces Cytokine Storm and Inflammation During Coronavirus Disease 19: Perspectives and Possible Therapeutic Approaches. Front. Pharmacol. 2020, 11, 592169. [Google Scholar] [CrossRef]
- Tang, N.; Li, D.; Wang, X.; Sun, Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J. Thromb. Haemost. 2020, 18, 844–847. [Google Scholar] [CrossRef] [Green Version]
- Bosmuller, H.; Matter, M.; Fend, F.; Tzankov, A. The pulmonary pathology of COVID-19. Virchows Arch. 2021, 478, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Inata, Y. Should we treat sepsis-induced DIC with anticoagulants? J. Intensive Care 2020, 8, 18. [Google Scholar] [CrossRef]
- Wada, H.; Hasegawa, K.; Watanabe, M. DIC: An update on diagnosis and treatment. Rinsho Ketsueki 2017, 58, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Impellizzeri, D.; Fusco, R.; Cordaro, M.; Siracusa, R.; Crupi, R.; Esposito, E.; Cuzzocrea, S. Ultramicronized palmitoylethanolamide (PEA-um((R))) in the treatment of idiopathic pulmonary fibrosis. Pharmacol. Res. 2016, 111, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Cordaro, M.; Crupi, R.; Siracusa, R.; Campolo, M.; Bruschetta, G.; Fusco, R.; Pugliatti, P.; Esposito, E.; Cuzzocrea, S. Protective Effects of Ultramicronized Palmitoylethanolamide (PEA-um) in Myocardial Ischaemia and Reperfusion Injury In Vivo. Shock 2016, 46, 202–213. [Google Scholar] [CrossRef]
- Peritore, A.F.; Siracusa, R.; Fusco, R.; Gugliandolo, E.; D’Amico, R.; Cordaro, M.; Crupi, R.; Genovese, T.; Impellizzeri, D.; Cuzzocrea, S.; et al. Ultramicronized Palmitoylethanolamide and Paracetamol, a New Association to Relieve Hyperalgesia and Pain in a Sciatic Nerve Injury Model in Rat. Int. J. Mol. Sci. 2020, 21, 3509. [Google Scholar] [CrossRef]
- Wang, B.; Wu, S.; Ma, Z.; Wang, T.; Yang, C. BMSCs pre-treatment ameliorates inflammation-related tissue destruction in LPS-induced rat DIC model. Cell Death Dis. 2018, 9, 1024. [Google Scholar] [CrossRef]
- Siracusa, R.; Fusco, R.; Cordaro, M.; Peritore, A.F.; D’Amico, R.; Gugliandolo, E.; Crupi, R.; Genovese, T.; Evangelista, M.; Di Paola, R.; et al. The Protective Effects of Pre- and Post-Administration of Micronized Palmitoylethanolamide Formulation on Postoperative Pain in Rats. Int. J. Mol. Sci. 2020, 21, 7700. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Siracusa, R.; Cordaro, M.; Crupi, R.; Peritore, A.F.; Gugliandolo, E.; D’Amico, R.; Petrosino, S.; Evangelista, M.; Di Paola, R.; et al. N-Palmitoylethanolamine-oxazoline (PEA-OXA): A new therapeutic strategy to reduce neuroinflammation, oxidative stress associated to vascular dementia in an experimental model of repeated bilateral common carotid arteries occlusion. Neurobiol. Dis. 2019, 125, 77–91. [Google Scholar] [CrossRef]
- Thachil, J. Disseminated intravascular coagulation—New pathophysiological concepts and impact on management. Expert Rev. Hematol. 2016, 9, 803–814. [Google Scholar] [CrossRef]
- Iba, T.; Levy, J.H.; Raj, A.; Warkentin, T.E. Advance in the Management of Sepsis-Induced Coagulopathy and Disseminated Intravascular Coagulation. J. Clin. Med. 2019, 8, 728. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, T.; Wada, H. Evaluation of FDP and D-dimer for the diagnosis of DIC. Rinsho Byori 2011, 147, 79–83. [Google Scholar]
- Chen, X.; Tan, J.; Yang, M.; Liao, Z.K.; Lu, C.; Huang, Y.; Wu, L.C. Genistein has the function of alleviating and treating disseminated intravascular coagulation caused by lipopolysaccharide. J. Nat. Med. 2018, 72, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Asakura, H.; Ogawa, H. COVID-19-associated coagulopathy and disseminated intravascular coagulation. Int. J. Hematol. 2021, 113, 45–57. [Google Scholar] [CrossRef]
- Asakura, H.; Sano, Y.; Omote, M.; Yoshida, T.; Ontachi, Y.; Mizutani, T.; Kaneda, M.; Yamazaki, M.; Morishita, E.; Takami, A.; et al. Significance of decreased plasma D-dimer levels following lipopolysaccharide-induced disseminated intravascular coagulation in rats. Int. J. Hematol. 2004, 79, 394–399. [Google Scholar] [CrossRef]
- Boral, B.M.; Williams, D.J.; Boral, L.I. Disseminated Intravascular Coagulation. Am. J. Clin. Pathol. 2016, 146, 670–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semeraro, N.; Ammollo, C.T.; Semeraro, F.; Colucci, M. Sepsis-associated disseminated intravascular coagulation and thromboembolic disease. Mediterr. J. Hematol. Infect. Dis. 2010, 2, e2010024. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Antonopoulou, S.; Demopoulos, C.A. Coronavirus 2019, Microthromboses, and Platelet Activating Factor. Clin. Ther. 2020, 42, 1850–1852. [Google Scholar] [CrossRef]
- Demopoulos, C.; Antonopoulou, S.; Theoharides, T.C. COVID-19, microthromboses, inflammation, and platelet activating factor. Biofactors 2020, 46, 927–933. [Google Scholar] [CrossRef]
- Theoharides, T.C. Potential association of mast cells with coronavirus disease 2019. Ann. Allergy Asthma Immunol. 2021, 126, 217–218. [Google Scholar] [CrossRef]
- Theoharides, T.C. COVID-19, pulmonary mast cells, cytokine storms, and beneficial actions of luteolin. Biofactors 2020, 46, 306–308. [Google Scholar] [CrossRef]
- Guilarte, M.; Sala-Cunill, A.; Luengo, O.; Labrador-Horrillo, M.; Cardona, V. The Mast Cell, Contact, and Coagulation System Connection in Anaphylaxis. Front. Immunol. 2017, 8, 846. [Google Scholar] [CrossRef] [Green Version]
- Asakura, H.; Sano, Y.; Yoshida, T.; Omote, M.; Ontachi, Y.; Mizutani, T.; Yamazaki, M.; Morishita, E.; Takami, A.; Miyamoto, K.; et al. Beneficial effect of low-molecular-weight heparin against lipopolysaccharide-induced disseminated intravascular coagulation in rats is abolished by coadministration of tranexamic acid. Intensive Care Med. 2004, 30, 1950–1955. [Google Scholar] [CrossRef]
- Peritore, A.F.; D’Amico, R.; Siracusa, R.; Cordaro, M.; Fusco, R.; Gugliandolo, E.; Genovese, T.; Crupi, R.; Di Paola, R.; Cuzzocrea, S.; et al. Management of Acute Lung Injury: Palmitoylethanolamide as a New Approach. Int. J. Mol. Sci. 2021, 22, 5533. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Cordaro, M.; Verde, R.; Schiano Moriello, A.; Marcolongo, G.; Schievano, C.; Siracusa, R.; Piscitelli, F.; Peritore, A.F.; Crupi, R.; et al. Oral Ultramicronized Palmitoylethanolamide: Plasma and Tissue Levels and Spinal Anti-hyperalgesic Effect. Front. Pharmacol. 2018, 9, 249. [Google Scholar] [CrossRef]
- Arfuso, F.; Giannetto, C.; Fazio, F.; Panzera, F.; Piccione, G. Training Program Intensity Induces an Acute Phase Response in Clinically Healthy Horses. J. Equine Vet. Sci. 2020, 88, 102986. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Cheng, P.; Zhang, H.; Li, N.; Qi, Y.; Wang, H.; Wang, Y.; Wang, W. The Protective Role of Tanshinone IIA in Silicosis Rat Model via TGF-beta1/Smad Signaling Suppression, NOX4 Inhibition and Nrf2/ARE Signaling Activation. Drug Des. Dev. Ther. 2019, 13, 4275–4290. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yang, J.; Guo, L.; Uyeminami, D.; Dong, H.; Hammock, B.D.; Pinkerton, K.E. Use of a soluble epoxide hydrolase inhibitor in smoke-induced chronic obstructive pulmonary disease. Am. J. Respir. Cell Mol. Biol. 2012, 46, 614–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusco, R.; Cordaro, M.; Siracusa, R.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Biochemical Evaluation of the Antioxidant Effects of Hydroxytyrosol on Pancreatitis-Associated Gut Injury. Antioxidants 2020, 9, 781. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Fusco, R.; D’Amico, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; Scuto, M.; Crupi, R.; Mandalari, G.; et al. Cashew (Anacardium occidentale L.) Nuts Counteract Oxidative Stress and Inflammation in an Acute Experimental Model of Carrageenan-Induced Paw Edema. Antioxidants 2020, 9, 660. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Cordaro, M.; Genovese, T.; Impellizzeri, D.; Siracusa, R.; Gugliandolo, E.; Peritore, A.F.; D’Amico, R.; Crupi, R.; Cuzzocrea, S.; et al. Adelmidrol: A New Promising Antioxidant and Anti-Inflammatory Therapeutic Tool in Pulmonary Fibrosis. Antioxidants 2020, 9, 601. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhou, C.L.; Zhou, Q.S.; Zou, H.D. Galantamine protects against lipopolysaccharide-induced acute lung injury in rats. Braz. J. Med. Biol. Res. 2016, 49, e5008. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, R.; Fusco, R.; Cordaro, M.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Scuto, M.; Cuzzocrea, S.; Di Paola, R.; et al. Modulation of NLRP3 Inflammasome through Formyl Peptide Receptor 1 (Fpr-1) Pathway as a New Therapeutic Target in Bronchiolitis Obliterans Syndrome. Int. J. Mol. Sci. 2020, 21, 2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siracusa, R.; Fusco, R.; Peritore, A.F.; Cordaro, M.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Crupi, R.; Smeriglio, A.; Mandalari, G.; et al. The Antioxidant and Anti-Inflammatory Properties of Anacardium occidentale L. Cashew Nuts in a Mouse Model of Colitis. Nutrients 2020, 12, 834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusco, R.; Gugliandolo, E.; Siracusa, R.; Scuto, M.; Cordaro, M.; D’Amico, R.; Evangelista, M.; Peli, A.; Peritore, A.F.; Impellizzeri, D.; et al. Formyl Peptide Receptor 1 Signaling in Acute Inflammation and Neural Differentiation Induced by Traumatic Brain Injury. Biology 2020, 9, 238. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amico, R.; Monaco, F.; Siracusa, R.; Cordaro, M.; Fusco, R.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Cuzzocrea, S.; Di Paola, R.; et al. Ultramicronized Palmitoylethanolamide in the Management of Sepsis-Induced Coagulopathy and Disseminated Intravascular Coagulation. Int. J. Mol. Sci. 2021, 22, 11388. https://doi.org/10.3390/ijms222111388
D’Amico R, Monaco F, Siracusa R, Cordaro M, Fusco R, Peritore AF, Gugliandolo E, Crupi R, Cuzzocrea S, Di Paola R, et al. Ultramicronized Palmitoylethanolamide in the Management of Sepsis-Induced Coagulopathy and Disseminated Intravascular Coagulation. International Journal of Molecular Sciences. 2021; 22(21):11388. https://doi.org/10.3390/ijms222111388
Chicago/Turabian StyleD’Amico, Ramona, Francesco Monaco, Rosalba Siracusa, Marika Cordaro, Roberta Fusco, Alessio Filippo Peritore, Enrico Gugliandolo, Rosalia Crupi, Salvatore Cuzzocrea, Rosanna Di Paola, and et al. 2021. "Ultramicronized Palmitoylethanolamide in the Management of Sepsis-Induced Coagulopathy and Disseminated Intravascular Coagulation" International Journal of Molecular Sciences 22, no. 21: 11388. https://doi.org/10.3390/ijms222111388
APA StyleD’Amico, R., Monaco, F., Siracusa, R., Cordaro, M., Fusco, R., Peritore, A. F., Gugliandolo, E., Crupi, R., Cuzzocrea, S., Di Paola, R., Impellizzeri, D., & Genovese, T. (2021). Ultramicronized Palmitoylethanolamide in the Management of Sepsis-Induced Coagulopathy and Disseminated Intravascular Coagulation. International Journal of Molecular Sciences, 22(21), 11388. https://doi.org/10.3390/ijms222111388