
Molecular Mechanisms of Mesenchymal Stem Cell-Based Therapy in Acute Kidney Injury


Abstract
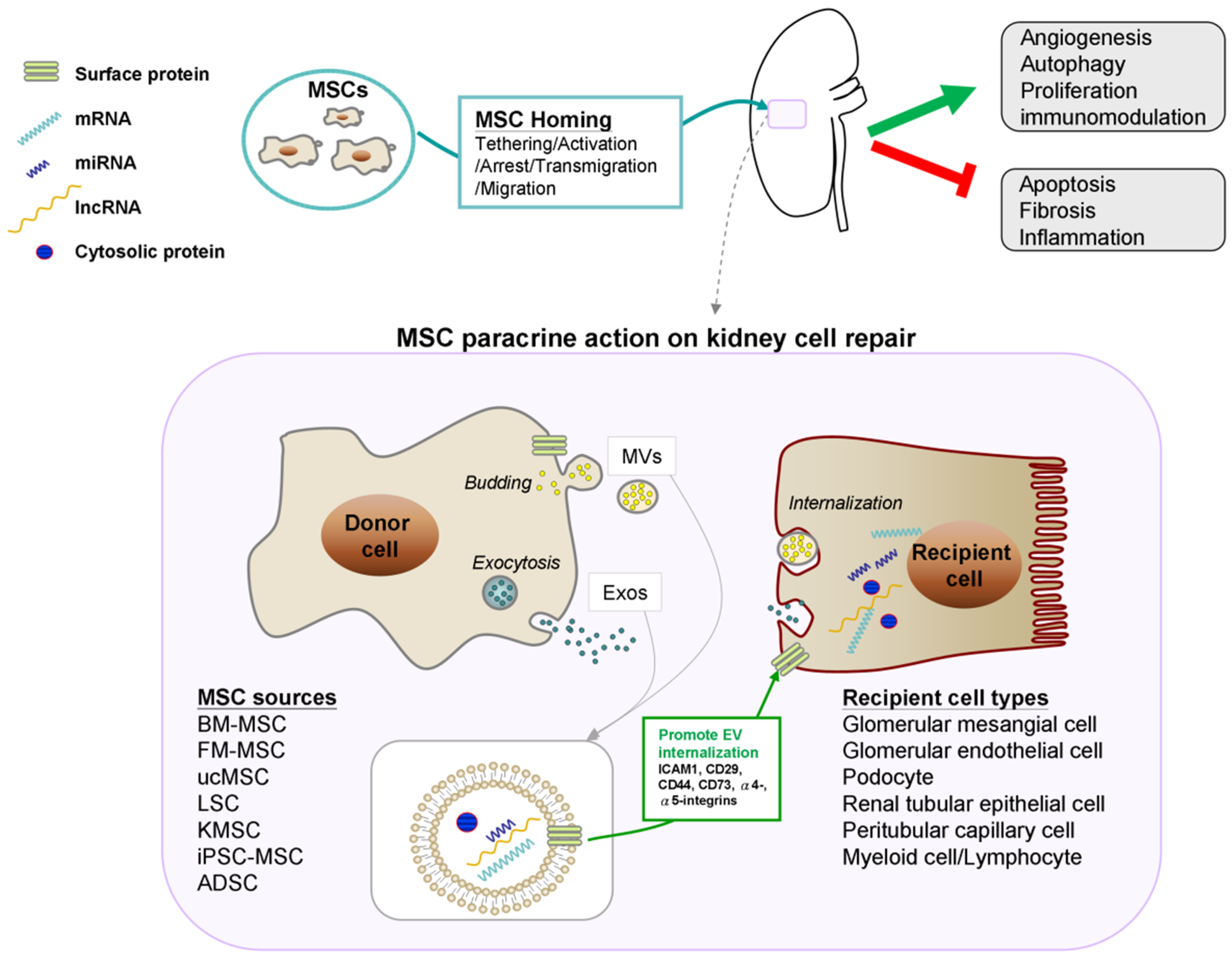
:1. Introduction
2. Mesenchymal Stem Cell-Based Therapy in AKI
2.1. Different Stem Cells Sources in AKI
2.2. MSCs and MSC-Derived EVs Protect from Acute Tubular Injury in Different Models
2.3. MSCs and MSC-Derived EVs Protect from Acute Glomerular Injury in Different Models
3. Delivered Organelles Shuttled from MSC-Derived EVs
3.1. Surface Proteins
3.2. Nucleic Acid (mRNA and miRNA) Trafficking
3.2.1. Delivery of miRNA
3.2.2. Delivery of mRNA
3.2.3. Delivery of lncRNA
3.3. Protein Trafficking
4. Clinical Trials
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Uchino, S.; Kellum, J.A.; Bellomo, R. Acute renal failure in critically ill patients: A multinational, multicenter study. JAMA 2005, 294, 813–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chertow, G.M.; Burdick, E.; Honour, M.; Bonventre, J.V.; Bates, D.W. Acute kidney injury, mortality, length of stay, and costs in hospitalized patients. J. Am. Soc. Nephrol. 2005, 16, 3365–3370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basile, D.P.; Anderson, M.D.; Sutton, T.A. Pathophysiology of acute kidney injury. Compr. Physiol. 2012, 2, 1303–1353. [Google Scholar] [CrossRef] [Green Version]
- Chawla, L.S.; Kimmel, P.L. Acute kidney injury and chronic kidney disease: An integrated clinical syndrome. Kidney Int. 2012, 82, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Makris, K.; Spanou, L. Acute Kidney Injury: Definition, Pathophysiology and Clinical Phenotypes. Clin. Biochem. Rev. 2016, 37, 85–98. [Google Scholar]
- Tetta, C.; Deregibus, M.C.; Camussi, G. Stem cells and stem cell-derived extracellular vesicles in acute and chronic kidney diseases: Mechanisms of repair. Ann. Transl. Med. 2020, 8, 570. [Google Scholar] [CrossRef]
- Lameire, N. The pathophysiology of acute renal failure. Crit. Care Clin. 2005, 21, 197–210. [Google Scholar] [CrossRef]
- Fang, W.; Wang, Z.; Li, Q. Gpr97 is an important mediator of AKI, and pharmacologic targeting of Gpr97mediated Sema3A signaling at multiple levels may provide a novel approach for the treatment of AKI. Gpr97 Exacerbates AKI by Mediating Sema3A Signaling. J. Am. Soc. Nephrol. 2018, 29, 1475–1489. [Google Scholar] [CrossRef] [Green Version]
- Cantley, L.G. Adult stem cells in the repair of the injured renal tubule. Nat. Clin. Pract. Nephrol. 2005, 1, 22–32. [Google Scholar] [CrossRef]
- Galderisi, U.; Peluso, G.; Di Bernardo, G. Clinical Trials Based on Mesenchymal Stromal Cells are Exponentially Increasing: Where are We in Recent Years? Stem Cell Rev. Rep. 2021, 1–14. [Google Scholar] [CrossRef]
- Bianco, P. “Mesenchymal” Stem Cells. Annu. Rev. Cell Dev. Biol. 2014, 30, 677–704. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, E.M.; Le Blanc, K.; Dominici, M.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Deans, R.J.; Krause, D.S.; Keating, A. Clarification of the nomenclature for MSC: The International Society for Cellular Therapy position statement. Cytotherapy 2005, 7, 393–395. [Google Scholar] [CrossRef]
- Lv, F.-J.; Tuan, R.S.; Cheung, K.M.C.; Leung, V.Y.L. Concise Review: The Surface Markers and Identity of Human Mesenchymal Stem Cells. Stem Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Herrera, M.B.; Bussolati, B.; Bruno, S.; Morando, L.; Mauriello-Romanazzi, G.; Sanavio, F. Exogenous mesenchymal stem cells localize to the kidney by means of CD44 following acute tubular injury. Kidney Int. 2007, 72, 430–441. [Google Scholar] [CrossRef] [Green Version]
- Herrera, M.B.; Bussolati, B.; Bruno, S. Mesenchymal stem cells contribute to renal repair on acute tubular epithelial injury. Int. J. Mol. Med. 2004, 14, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Morigi, M.; Imberti, B.; Zoja, C. Mesenchymal stem cells are renotropic, helping to repair the kidney and improve function in acute renal failure. J. Am. Soc. Nephrol. 2004, 15, 1794–1804. [Google Scholar] [CrossRef] [PubMed]
- Kramann, R.; Humphreys, B.D. Kidney pericytes: Roles in regeneration and fibrosis. Semin. Nephrol. 2014, 34, 374–383. [Google Scholar] [CrossRef] [Green Version]
- Griffin, M.D.; Ryan, A.E.; Alagesan, S.; Lohan, P.; Treacy, O.; Ritter, T. Anti-donor immune responses elicited by allogeneic mesenchymal stem cells: What have we learned so far? Immunol. Cell Biol. 2013, 91, 40–51. [Google Scholar] [PubMed]
- Hass, R.; Kasper, C.; Bohm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [PubMed] [Green Version]
- Ragni, E.; Banfi, F.; Barilani, M. Extracellular vesicle-shuttled mRNA in mesenchymal stem cell communication. Stem Cells 2017, 35, 1093–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najar, M.; Raicevic, G.; Fayyad-Kazan, H.; Bron, D.; Toungouz, M.; Lagneaux, L. Mesenchymal stromal cells and immunomodulation: A gathering of regulatory immune cells. Cytotherapy 2016, 18, 160–171. [Google Scholar] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Born, L.J.; Harmon, J.W.; Jay, S.M. Therapeutic potential of extracellular vesicle-associated long noncoding RNA. Bioeng. Transl. Med. 2020, 5, e10172. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaborowski, M.P.; Balaj, L.; Breakefield, X.O.; Lai, C.P. Extracellular Vesicles: Composition, Biological Relevance, and Methods of Study. BioScience 2015, 65, 783–797. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, B.D.; Bonventre, J.V. Mesenchymal stem cells in acute kidney injury. Annu. Rev. Med. 2008, 59, 311–325. [Google Scholar] [CrossRef] [Green Version]
- Brossa, A.; Papadimitriou, E.; Collino, F.; Incarnato, D.; Oliviero, S.; Camussi, G. Role of CD133 molecule in Wnt response and renal repair. Stem Cells Transl. Med. 2018, 7, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Biancone, L.; Cantaluppi, V.; Duò, D.; Deregibus, M.C.; Torre, C.; Camussi, G. Role of L-selectin in the vascular homing of peripheral blood-derived endothelial progenitor cells. J. Immunol. 2004, 173, 5268–5274. [Google Scholar] [CrossRef] [Green Version]
- Cantaluppi, V.; Medica, D.; Mannari, C. Endothelial progenitor cell-derived extracellular vesicles protect from complement-mediated mesangial inury in experimental anti-Thy 1.1 glomerulonephritis. Nephrol. Dial. Transplant. 2015, 30, 410–422. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, M.B.H.; Bruno, S.; Grange, C.; Tapparo, M.; Cantaluppi, V.; Tetta, C. Human liver stem cells and derived extracellular vesicles improve recovery in a murine model of acute kidney injury. Stem Cell Res Ther. 2014, 5, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Z.; Zhang, G.; Wang, F.; Liu, H.; Liu, L.; Han, Y. Protective effects of mesenchymal stem cells with CXCR4 up-regulation in a rat renal transplantation model. PLoS ONE 2013, 8, e82949. [Google Scholar] [CrossRef] [Green Version]
- Bai, M.; Zhang, L.; Fu, B.; Bai, J.; Zhang, Y.; Cai, G. IL-17A improves the efficacy of mesenchymal stem cells in ischemic-reperfusion renal injury by increasing Treg percentages by the COX-2/PGE2 pathway. Kidney Int. 2018, 93, 814–825. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.; Liu, D.D.; Thakor, A.S. Mesenchymal Stromal Cell Homing: Mechanisms and Strategies for Improvement. iScience 2019, 15, 421–438. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, B.D. Kidney injury, stem cells and regeneration. Curr. Opin. Nephrol. Hypertens. 2014, 23, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, R.; Sann, J.; Frueh, J.T.; Ullrich, E.; Geiger, H.; Baer, P.C. Tracking of Adipose-Derived Mesenchymal Stromal/Stem Cells in a Model of Cisplatin-Induced Acute Kidney Injury: Comparison of Bioluminescence Imaging versus qRT-PCR. Int. J. Mol. Sci. 2018, 19, 2564. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Ren, X.; Ren, R.; Wang, Y.; Shang, J. The combination of G-CSF and AMD3100 mobilizes bone marrow-derived stem cells to protect against cisplatin-induced acute kidney injury in mice. Stem Cell Res. Ther. 2021, 12, 209. [Google Scholar] [CrossRef]
- Medica, D.; Dellepiane, S.; Cantaluppi, V. Regenerative Role of Stem Cell-Derived Extracellular Vesicles in Acute Kidney Injury. Nephron 2020, 144, 1–6. [Google Scholar] [CrossRef]
- Cha, J.M.; Shin, E.K.; Sung, J.H.; Moon, G.J.; Kim, E.H.; Cho, Y.H. Efficient scalable production of therapeutic microvesicles derived from human mesenchymal stem cells. Sci. Rep. 2018, 8, 1171. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.M.; Wang, M.Z. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatti, S.; Bruno, S.; Deregibus, M.C.; Sordi, A.; Cantaluppi, V.; Tetta, C. Microvesicles derived from human adult mesenchymal stem cells protect against ischaemia-reperfusioninduced acute and chronic kidney injury. Nephrol. Dial. Transplant. 2011, 26, 1474–1483. [Google Scholar] [CrossRef] [Green Version]
- Ranghino, A.; Bruno, S.; Bussolati, B. The effects of glomerular and tubular renal progenitors and derived extracellular vesicles on recovery from acute kidney injury. Stem Cell Res. Ther. 2017, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Yan, Y.; Song, C.; Ding, Y.; Du, T. Microvesicles derived from human Wharton’s Jellymesenchymal stem cells ameliorate ischemia-reperfusion-induced renal fibrosis by releasing from G2/M cell cycle arrest. Biochem. J. 2017, 474, 4207–4218. [Google Scholar] [CrossRef]
- Bruno, S.; Grange, C.; Collino, F.; Deregibus, M.C.; Cantaluppi, V.; Biancone, L. Microvesicles derived from mesenchymal stem cells enhance survival in a lethal model of acute kidney injury. PLoS ONE 2012, 7, e33115. [Google Scholar] [CrossRef]
- Overath, J.M.; Gauer, S.; Obermüller, N.; Schubert, R.; Schäfer, R.; Geiger, H.; Baer, P.C. Short-term preconditioning enhances the therapeutic potential of adipose-derived stromal/stem cell-conditioned medium in cisplatin-induced acute kidney injury. Exp. Cell Res. 2016, 342, 175–183. [Google Scholar] [CrossRef]
- Finn, W.F. Acute Renal Failurea Companion to Brenner and Rector’s The Kidney; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2001. [Google Scholar]
- Lieberthal, W.; Nigam, S.K. Acute renal failure. I. Relative importance of proximal vs. distal tubular injury. Am. J. Physiol. Ren. Physiol. 1998, 275, F623–F632. [Google Scholar] [CrossRef]
- Tsuda, H.; Yamahara, K.; Ishikane, S.; Otani, K.; Nakamura, A.; Sawai, K.; Ichimaru, N.; Sada, M.; Taguchi, A.; Hosoda, H.; et al. Allogenic fetal membrane-derived mesenchymal stem cells contribute to renal repair in experimental glomerulonephritis. Am. J. Physiol. Ren. Physiol. 2010, 299, F1004–F1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoja, C.; Garcia, P.B.; Rota, C.; Conti, S.; Gagliardini, E.; Corna, D.; Zanchi, C.; Bigini, P.; Benigni, A.; Remuzzi, G.; et al. Mesenchymal stem cell therapy promotes renal repair by limiting glomerular podocyte and progenitor cell dysfunction in adriamycin-induced nephropathy. Am. J. Physiol. Ren. Physiol. 2012, 303, F1370–F1381. [Google Scholar] [CrossRef] [Green Version]
- Iseri, K.; Iyoda, M.; Ohtaki, H.; Matsumoto, K.; Wada, Y.; Suzuki, T.; Yamamoto, Y.; Saito, T.; Hihara, K.; Tachibana, S.; et al. Therapeutic effects and mechanism of conditioned media from human mesenchymal stem cells on anti-GBM glomerulonephritis in WKY rats. Am. J. Physiol. Ren. Physiol. 2016, 310, F1182–F1191. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wei, W.; Liu, M.L. Extracellular vesicles and lupus nephritis—New insights into pathophysiology and clinical implications. J. Autoimmun. 2020, 115, 102540. [Google Scholar] [CrossRef]
- Record, M.; Carayon, K.; Poirot, M.; Silvente-Poirot, S. Exosomes as new vesicular lipid transporters involved in cell–cell communication and various pathophysiologies. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2014, 1841, 108–120. [Google Scholar] [CrossRef]
- Christianson, H.C.; Svensson, K.J.; van Kuppevelt, T.H.; Li, J.P.; Belting, M. Cancer cell exosomes depend on cell-surface heparan sulfate proteoglycans for their internalization and functional activity. Proc. Natl. Acad. Sci. USA 2013, 110, 17380–17385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szatmári, T.; Hargitai, R.; Sáfrány, G.; Lumniczky, K. Extracellular Vesicles in Modifying the Effects of Ionizing Radiation. Int. J. Mol. Sci. 2019, 20, 5527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R.F. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, J.H.; Liu, Y.; Gao, H.; Dominguez, J.M.; Xie, D.; Kelly, K.J. Renal tubular cell-derived extracellular vesicles accelerate the recovery of established renal ischemia reperfusion injury. J. Am. Soc. Nephrol. 2017, 28, 3533–3544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.M.; Choi, E.-J.; Kim, J.H.; Kim, T.D.; Kim, Y.-K.; Kang, C.; Gho, Y.S. A membranous form of ICAM-1 on exosomes efficiently blocks leukocyte adhesion to activated endothelial cells. Biochem. Biophys. Res. Commun. 2010, 397, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, J.; Miekus, K.; Kucia, M.; Zhang, J.; Reca, R.; Dvorak, P.; Ratajczak, M. Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: Evidence for horizontal transfer of mRNA and protein delivery. Leukemia 2006, 20, 847–856. [Google Scholar] [CrossRef] [Green Version]
- Schorey, J.S.; Bhatnagar, S. Exosome function: From tumor immunology to pathogen biology. Traffic 2008, 9, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Bruno, S.; Grange, C.; Deregibus, M.C. Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J. Am. Soc. Nephrol. 2009, 20, 1053–1067. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.Y.; Moon, S.J.; Ratliff, B.B. Microparticles from kidney-derived mesenchymal stem cells act as carriers of proangiogenic signals and contribute to recovery from acute kidney injury. PLoS ONE 2014, 9, e87853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camussi, G.; Deregibus, M.C.; Tetta, C. Paracrine/endocrine mechanism of stem cells on kidney repair: Role of microvesicle-mediated transfer of genetic information. Curr. Opin. Nephrol. Hypertens. 2010, 19, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Lyon, C.J.; Fletcher, J.K.; Tang, W.; Wan, M.; Hu, T.Y. Extracellular vesicle activities regulating macrophage- and tissue-mediated injury and repair responses. Acta Pharm. Sin. B 2021, 11, 1493–1512. [Google Scholar] [CrossRef] [PubMed]
- Lindoso, R.S.; Collino, F.; Bruno, S. Extracellular vesicles released from mesenchymal stromal cells modulate miRNA in renal tubular cells and inhibit ATP depletion injury. Stem Cells Dev. 2014, 23, 1809–1819. [Google Scholar] [CrossRef] [Green Version]
- Collino, F.; Bruno, S.; Incarnato, D. AKI recovery induced by mesenchymal stromal cell-derived extracellular vesicles carrying microRNAs. J. Am. Soc. Nephrol. 2015, 26, 2349–2360. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Zou, X.; Ju, G.; Zhang, G.; Bao, E.; Zhu, Y. Mesenchymal stromal cells derived extracellular vesicles ameliorate acute renal ischemia reperfusion injury by inhibition of mitochondrial fission through miR-30. Stem Cells Int. 2016, 2016, 2093940. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Pei, L.; Lin, F. Exosomes from human-bone-marrow-derived mesenchymal stem cells protect against renal ischemia/ reperfusion injury via transferring miR-199a-3p. J. Cell Physiol. 2019, 234, 23736–23749. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, Y.; Li, Y.; Liu, W.; Yin, L.; Yin, S.; Ji, C.; Hu, Y.; Wang, Q.; Zhou, X.; et al. Human umbilical cord mesenchymal stem cell exosomes alleviate sepsis-associated acute kidney injury via regulating microRNA-146b expression. Biotechnol. Lett. 2020, 42, 669–679. [Google Scholar] [CrossRef]
- Cao, J.Y.; Wang, B.; Tang, T.T.; Wen, Y.; Li, Z.L.; Feng, S.T.; Wu, M.; Liu, D.; Yin, D.; Ma, K.L.; et al. Exosomal miR-125b-5p deriving from mesenchymal stem cells promotes tubular repair by suppression of p53 in ischemic acute kidney injury. Theranostics 2021, 11, 5248–5266. [Google Scholar] [CrossRef]
- Tomasoni, S.; Longaretti, L.; Rota, C. Transfer of growth factor receptor mRNA via exosomes unravels the regenerative effect of mesenchymal stem cells. Stem Cells Dev. 2013, 22, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Zou, X.; Cheng, J. Human Wharton’s jelly-derived mesenchymal stromal cells reduce renal fibrosis through induction of native and foreign hepatocyte growth factor synthesis in injured tubular epithelial cells. Stem Cell Res. Ther. 2013, 4, 59. [Google Scholar] [CrossRef] [Green Version]
- Ju, G.Q.; Cheng, J.; Zhong, L. Microvesicles derived from human umbilical cord mesenchymal stem cells facilitate tubular epithelial cell dedifferentiation and growth via hepatocyte growth factor induction. PLoS ONE 2015, 10, e0121534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.Y.; Hou, Y.P.; Zou, X.Y.; Xing, X.Y.; Ju, G.Q.; Zhong, L.; Sun, J. Oct-4 Enhanced the Therapeutic Effects of Mesenchymal Stem Cell-Derived Extracellular Vesicles in Acute Kidney Injury. Kidney Blood Press. Res. 2020, 45, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Li, D.; Chen, X. Extracellular vesicles from human-induced pluripotent stem cell-derived mesenchymal stromal cells (hiPSC-MSCs) protect against renal ischemia/reperfusion injury via delivering specificity protein (SP1) and transcriptional activating of sphingosine kinase 1 and inhibiting necroptosis. Cell Death Dis. 2017, 8, 3200. [Google Scholar] [PubMed] [Green Version]
- Wang, J.; Jia, H.; Zhang, B. HucMSC exosome-transported 14-33ζ prevents the injury of cisplatin to HK-2 cells by inducing autophagy in vitro. Cytotherapy 2018, 20, 29–44. [Google Scholar] [CrossRef]
- Tseng, W.C.; Lee, P.Y.; Tsai, M.T.; Chang, F.P.; Chen, N.J.; Chien, C.T.; Hung, S.C.; Tarng, D.C. Hypoxic mesenchymal stem cells ameliorate acute kidney ischemia-reperfusion injury via enhancing renal tubular autophagy. Stem Cell Res. Ther. 2021, 12, 367. [Google Scholar] [CrossRef]
- Zhao, L.; Hu, C.; Han, F.; Wang, J.; Chen, J. Regenerative abilities of mesenchymal stem cells via acting as an ideal vehicle for subcellular component delivery in acute kidney injury. J. Cell. Mol. Med. 2020, 24, 4882–4891. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; McIntyre, R.L.; Janssens, G.E.; Houtkooper, R.H. Mitochondrial fission and fusion: A dynamic role in aging and potential target for age-related disease. Mech. Ageing Dev. 2020, 186, 111212. [Google Scholar] [CrossRef]
- Westermann, B. Mitochondrial fusion and fission in cell life and death. Nat. Rev. Mol. Cell Biol. 2010, 11, 872–884. [Google Scholar] [CrossRef]
- Wu, L.; Rong, C.; Zhou, Q.; Zhao, X.; Zhuansun, X.M.; Wan, S.; Sun, M.M.; Wang, S.L. Bone Marrow Mesenchymal Stem Cells Ameliorate Cisplatin-Induced Renal Fibrosis via miR-146a-5p/Tfdp2 Axis in Renal Tubular Epithelial Cells. Front. Immunol. 2020, 11, 623693. [Google Scholar] [CrossRef]
- Zhou, D.; Tan, R.J.; Lin, L.; Zhou, L.; Liu, Y. Activation of hepatocyte growth factor receptor, c-met, in renal tubules is required for renoprotection after acute kidney injury. Kidney Int. 2013, 84, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Liang, J.; Ni, S.; Zhou, T.; Qing, X.; Li, H.; He, W.; Chen, J.; Li, F.; Zhuang, Q. A mesenchymal-to-epithelial transition initiates and is required for the nuclear reprogramming of mouse fibroblasts. Cell Stem Cell 2010, 7, 51–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xuan, Z.; Liu, C. Long non-coding RNAs and complex human diseases. Int. J. Mol. Sci. 2013, 14, 18790–18808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [Green Version]
- Kung, J.T.; Colognori, D.; Lee, J.T. Long noncoding RNAs: Past, present, and future. Genetics 2013, 193, 651–669. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhu, X.; Huang, S. Clinical applications of extracellular vesicle long RNAs. Crit. Rev. Clin. Lab. Sci. 2020, 57, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, P.J.; Chang, H.Y. Long noncoding RNAs: Cellular address codes in development and disease. Cell 2013, 152, 1298–1307. [Google Scholar] [CrossRef] [Green Version]
- Mercer, T.R.; Mattick, J.S. Structure and function of long noncoding RNAs in epigenetic regulation. Nat. Struct. Mol. Biol. 2013, 20, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wapinski, O.; Chang, H.Y. Long noncoding RNAs and human disease. Trends Cell Biol. 2011, 21, 354–361. [Google Scholar] [CrossRef]
- Cooper, D.R.; Wang, C.; Patel, R.; Trujillo, A.; Patel, N.A.; Prather, J.; Gould, L.J.; Wu, M.H. Human adipose-derived stem cell conditioned media and exosomes containing MALAT1 promote human dermal fibroblast migration and ischemic wound healing. Adv. Wound Care 2018, 7, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.; Zhang, L.; Liang, C.; Liu, B.; Pan, X.; Wang, Y.; Zhang, Y.; Zhang, Y.; Xie, W.; Yan, B. Stem cell-derived exosomes prevent aging-induced cardiac dysfunction through a novel exosome/lncRNA MALAT1/NF-κB/TNF-α signaling pathway. Oxid. Med. Cell. Longev. 2019, 2019, 9739258. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zou, R.; Wang, Z.; Wen, C.; Zhang, F.; Lin, F. Exosomal KLF3-AS1 from hMSCs promoted cartilage repair and chondrocyte proliferation in osteoarthritis. Biochem. J. 2018, 475, 3629–3638. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wang, L.; Wu, Q.; Zheng, G.; Long, H.; Wu, H.; Zhou, C.; Guo, T.; Zhong, T.; Wang, L. Long noncoding RNA H19 upregulates vascular endothelial growth factor A to enhance mesenchymal stem cells survival and angiogenic capacity by inhibiting miR-199a-5p. Stem Cell Res. Ther. 2018, 9, 109. [Google Scholar] [CrossRef]
- Jin, Y.; Wang, J.; Li, H.; Gao, S.; Shi, R.; Yang, D.; Wang, X.; Wang, X.; Zhu, L.; Wang, X. Extracellular vesicles secreted by human adipose-derived stem cells (hASCs) improve survival rate of rats with acute liver failure by releasing lncRNA H19. EBioMedicine 2018, 34, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Lorenzen, J.M.; Schauerte, C.; Kielstein, J.T.; Hübner, A.; Martino, F.; Fiedler, J.; Gupta, S.K.; Faulhaber-Walter, R.; Kumarswamy, R.; Hafer, C.; et al. Circulating long noncoding RNATapSaki is a predictor of mortality in critically ill patients with acute kidney injury. Clin. Chem. 2015, 61, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Liu, L.; Zhang, F.; Gu, J.; Pan, G. LncRNA TapSAKI promotes inflammation injury in HK-2 cells and urine derived sepsis-induced kidney injury. J. Pharm. Pharmacol. 2019, 71, 839–848. [Google Scholar] [CrossRef]
- Han, X.; Yuan, Z.; Jing, Y.; Zhou, W.; Sun, Y.; Xing, J. Knockdown of lncRNA TapSAKI alleviates LPS-induced injury in HK-2 cells through the miR-205/IRF3 pathway. Open Med. 2021, 16, 581–590. [Google Scholar] [CrossRef]
- Pozuelo-Rubio, M. 14-3-3 Proteins are regulators of autophagy. Cells 2012, 1, 754–773. [Google Scholar] [CrossRef] [PubMed]
- Ezquer, M.E.; Ezquer, F.E.; Arango-Rodríguez, M.L.; Conget, P.A. MSC transplantation: A promising therapeutic strategy to manage the onset and progression of diabetic nephropathy. Biol. Res. 2012, 45, 289–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peired, A.J.; Sisti, A.; Romagnani, P. Mesenchymal stem cell-based therapy for kidney disease: A review of clinical evidence. Stem Cells Int. 2016, 2016, 4798639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezquer, F.; Giraud-Billoud, M.; Carpio, D.; Cabezas, F.; Conget, P.; Ezquer, M. Proregenerative microenvironment triggered by donor mesenchymal stem cells preserves renal function and structure in mice with severe diabetes mellitus. BioMed Res. Int. 2015, 2015, 164703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.; Shad, F.; Smith, M.C. Acute kidney injury: A guide to diagnosis and management. Am. Fam. Physician 2012, 86, 631–639. [Google Scholar] [PubMed]
- Tögel, F.E.; Westenfelder, C. Kidney protection and regeneration following acute injury: Progress through stem cell therapy. Am. J. Kidney Dis. 2012, 60, 1012–1022. [Google Scholar] [CrossRef]
- Swaminathan, M.; Stafford-Smith, M.; Chertow, G.M.; Warnock, D.G.; Paragamian, V.; Brenner, R.M. Allogeneic mesenchymal stem cells for treatment of AKI after cardiac surgery. J. Am. Soc. Nephrol. 2018, 29, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Ilic, D.; Liovic, M. Industry updates from the field of stem cell research and regenerative medicine in August 2020. Regen. Med. 2020, 15, 2329–2334. [Google Scholar] [CrossRef]
- Mahendiratta, S.; Bansal, S.; Sarma, P.; Kumar, H.; Choudhary, G.; Kumar, S.; Prakash, A.; Sehgal, R.; Medhi, B. Stem cell therapy in COVID-19: Pooled evidence from SARS-CoV-2, SARS-CoV, MERS-CoV and ARDS: A systematic review. Biomed. Pharmacother. 2021, 137, 111300. [Google Scholar] [CrossRef]
- Nargesi, A.A.; Lerman, L.O.; Eirin, A. Mesenchymal stem cell-derived extracellular vesicles for kidney repair: Current status and looming challenges. Stem Cell Res. Ther. 2017, 8, 273. [Google Scholar] [CrossRef]
- Kumar, S. Cellular and molecular pathways of renal repair after acute kidney injury. Kidney Int. 2018, 93, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thadhani, R.; Pascual, M.; Bonventre, J.V. Acute renal failure. N. Engl. J. Med. 1996, 334, 1448–1460. [Google Scholar] [CrossRef] [PubMed]
- Yuana, Y.; Böing, A.N.; Grootemaat, A.E.; van der Pol, E.; Hau, C.M.; Cizmar, P.; Buhr, E.; Sturk, A.; Nieuwland, R. Handling and storage of human body fluids for analysis of extracellular vesicles. J. Extracell. Vesicles 2015, 4, 29260. [Google Scholar] [CrossRef] [PubMed]
- Eldh, M.; Lötvall, J.; Malmhäll, C.; Ekström, K. Importance of RNA isolation methods for analysis of exosomal RNA: Evaluation of different methods. Mol. Immunol. 2012, 50, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Ramasubramanian, L.; Kumar, P.; Wang, A. Engineering Extracellular Vesicles as Nanotherapeutics for Regenerative Medicine. Biomolecules 2019, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-W.; Rocco, P.R.; Pelosi, P. Mesenchymal stem cell therapy for acute respiratory distress syndrome: A light at the end of the tunnel? Anesthesiology 2015, 122, 238–240. [Google Scholar] [CrossRef] [Green Version]
- Park, S.E.; Lee, N.K.; Lee, J.; Hwang, J.W.; Choi, S.J.; Hwang, H.; Hyung, B.; Chang, J.W.; Na, D.L. Distribution of human umbilical cord blood-derived mesenchymal stem cells in the Alzheimer’s disease transgenic mouse after a single intravenous injection. Neuroreport 2016, 27, 235–241. [Google Scholar] [CrossRef]
- Harting, M.T.; Jimenez, F.; Xue, H.; Fischer, U.M.; Baumgartner, J.; Dash, P.K.; Cox, C.S. Intravenous mesenchymal stem cell therapy for traumatic brain injury. J. Neurosurg. 2009, 110, 1189–1197. [Google Scholar] [CrossRef] [Green Version]
- Argibay, B.; Trekker, J.; Himmelreich, U.; Beiras, A.; Topete, A.; Taboada, P.; Pérez-Mato, M.; Vieites-Prado, A.; Iglesias-Rey, R.; Rivas, J. Intraarterial route increases the risk of cerebral lesions after mesenchymal cell administration in animal model of ischemia. Sci. Rep. 2017, 7, 40758. [Google Scholar] [CrossRef]


{kind=link}
| Histology | Authors/Year Reference | EV Sources | EV Types | Experimental Model | Species | EV Factors | Molecular Response | Functional Modulation |
|---|---|---|---|---|---|---|---|---|
| Acute Tubular Injury | Herrera et al., 2007 [17] | BM-MSCs | NM | In vitro/in vivo, glycerol-induced AKI | Mouse | NM | ↑CD44 and hyaluronic acid (major ligand of CD44) interactions | ↑exogenous MSC migration and homing |
| Gatti et al., 2011 [43] | BM-MSCs | MVs | In vivo, I/R induced acute tubular injury | Rat | NM | NM | ↓tubular cell apoptosis, ↑TEC proliferation | |
| Bruno et al., 2012 [46] | BM-MSCs | MVs | In vitro/in vivo, cisplatin-induced acute tubular injury | Mouse | Human POLR2E mRNA | ↑anti-apoptotic genes, Bcl-xL, Bcl2, and BIRC8, ↓apoptosis genes, Casp1, Casp8, and LTA | ↑renal function, morphology, and survival | |
| Mb et al., 2014 [33] | hLSCs | NM | In vitro/in vivo, intra-muscle glycerol induced AKI | Mouse | NM | ↑PCNA expression | ↑tubular cell proliferation, ↑renal function, ↑morphology | |
| Chen et al., 2017 [45] | hWJMSCs | MVs | In vitro/in vivo, I/R-induced renal fibrosis | Rat | NM | ↑ERK1/2 signaling ↓EMT–related protein, TGF-β1 ↑cell cycle-related proteins, CDK 1 and CyclinB1 | ↑proliferation, ↓apoptosis, ↓collagen deposition, ↑cells in G2/M cell cycle, ↓fibrosis, ↓EMT | |
| Ranghino et al., 2017 [44] | Gl-MSCs T-CD133+ cells | Gl-MSC-EVs T-CD133+-EVs | In vivo, I/R induced acute tubular injury | Mouse | 62 group of miRNAs | NM | ↑TEC proliferation | |
| Overath et al., 2016 [47] | ADSC-pCM | pCM | In vitro/in vivo, cisplatin-induced acute tubular injury | Mouse | 64 expressed proteins | ↓inflammatory cytokines, IL-1β, IL-6 | ↑ survival ↓ serum Cr and N-GAL | |
| Acute Glomerular Injury | Tsuda et al., 2010 [50] | FM-MSCs | NM | In vitro/in vivo, anti-Thy1 nephritis | rats | NM | ↓TNF and MCP-1 through a PGE2-dependent mechanism. | ↓Proteinuria ↓mesangial matrix/cell proliferation, ↓glomerular monocyte/macrophage infiltration, |
| Zoja et al., 2012 [51] | BM-MSCs | NM | In vitro/in vivo, Adriamycin-induced crescentic nephritis | rats | NM | ↑VEGF expression ↑nephrin and CD2AP | ↓monocyte infiltration, ↓podocyte apoptosis, ↓microvascular rarefaction | |
| Iseri et al., 2016 [52] | hMSC-CM | CM | In vitro/in vivo, anti-glomerular basement membrane nephritis | rats | NM | ↓proinflammatory cytokines TNF-α, IL-1-β, MCP-1, and IL-6 | ↑M2 macrophage polarization, ↓proteinuria and crescent formation |
| Substances Delivered | Authors/Year, Reference | EV Sources | EV Types | Experimental AKI Type | Species | EV Factors | Molecular Response | Functional Modulation |
|---|---|---|---|---|---|---|---|---|
| Delivery of miRNA | Lindoso et al., 2014 [66] | hMSCs | NM | H/R of PTECs in ATP depletion model | PTECs (HK-2) | 20 miRNAs ( such as miR-222, miR-145, etc) | ↓coding-mRNAs: CASP-3, CASP-7, SHC1 and SMAD4 | ↓cell death by apoptosis or hypoxia |
| Collino et al., 2015 [67] | BM-MSCs | NM | Glycerol | Mouse | 8 miRNA families (miR-483-5p, miR-191, miR-283p, miR-744, miR-423-5p, miR-24, miR-129-3p, miR-148a) | ↑genes with fatty acid metabolism, complement, and coagulation cascades ↓genes with inflammation, and adhesion molecules | ↑Proregeneration, ↓hyaline casts, ↓tubular necrosis, ↓tubular damage markers: lipocalin2 and fibrinogen subunits | |
| Gu et al., 2016 [68] | hWJMSCs | NM | I/R by unilateral nephrectomy | Rat | miR-30 | ↓DRP1 expression | ↓mitochondrial fission | |
| Zhu et al., 2019 [69] | BM-MSCs | Exos | I/R | Mouse | miR-199a-3p | ↓semaphorin 3A, ↑AKT and ERK pathways | ↓cell apoptosis | |
| Zhang et al., 2020 [70] | hucMSCs | Exos | Sepsis model through cecal ligation | Mouse | miRNA-146b | ↓IRAK1 expression, ↑NF-κB activity | ↑survival and kidney function | |
| Cao et al., 2021 [71] | hucMSCs | Exos | I/R | Mouse | miR-125b-5p | ↓p53 protein, ↑CDK1 and Cyclin B1, ↓apoptosis-related proteins, Bax and cleaved-caspase-3, ↑anti-apoptosis protein, Bcl-2 | ↑proliferative TECs, ↓G2/M cell cycle arrest and apoptosis of TECs | |
| Delivery of mRNA | Bruno et al., 2009 [62] | BM-MSCs | MVs | Glycerol | Mouse | Human POLR2E mRNA | ↑cytoplasmic POLR2E protein, ↑cytoplasmic and nuclear SUMO-1 protein | ↑proliferative and anti-apoptotic effects |
| Bruno et al., 2012 [46] | BM-MSCs | MVs | Cisplatin | Mouse | Human POLR2E mRNA | ↑anti-apoptotic genes, Bcl-xL, Bcl2, and BIRC8, ↓apoptosis genes, Casp1, Casp8, and LTA | ↑renal function, morphology, and survival | |
| Tomasoni et al., 2013 [72] | BM-MSCs | Exos | Cisplatin | PTECs (HK2) | IGF-1R mRNA | ↑IGF-1R-corresponding protein, IGF-1R | ↑sensitivity to IGF-1, ↑PTEC proliferation | |
| Du et al., 2013 [73] | WJ-MSCs | NM | I/R by renal pedicle ligation | Rat | Human HGF mRNA | ↑HGF protein expression, ↑TGF-β1, ↓α-SMA/E-cadherin | ↓renal fibrosis ↓tubular EMT ↓renal fibrosis | |
| Choi et al., 2014 [63] | KMSCs | MVs | I/R | Mouse | VEGF-A, IGF-1, and FGF mRNA | ↑PCNA, ↓CD 31 | ↑cell proliferation, ↑angiogenesis | |
| Ju et al., 2015 [74] | hucMSCs | MVs | I/R by renal pedicle ligation | Rat | Human HGF mRNA | ↑HGF protein expression, ↑ERK1/2 signaling activation | ↑TEC de-differentiation ↓apoptosis | |
| Ragni et al., 2017 [23] | BM-MSCs and hucMSCs | EVs | Cisplatin | PTECs (HKC8) | IL-10 mRNA | ↑IL-10-corresponding protein, IL-10, in PTECs | ↑rescue AKI | |
| Zhang et al., 2020 [75] | hucMSCs | NM | I/R by unilateral nephrectomy | Mouse | Oct-4 mRNA | ↓Snail expression, ↓α-SMA | ↓EMT, ↓apoptosis ↑proliferation | |
| Delivery of proteins | Yuan et al., 2017 [76] | iPSC-MSCs | EVs | I/R by renal pedicle ligation | Rat | SP1 | ↑SP1–SK1–S1P signaling pathway, ↑SK1, ↑S1P | ↓necroptosis |
| Wang et al., 2018 [77] | hucMSCs | Exos | Cisplatin | PTECs (HK2) | 14-3-3ζ | ↑PCNA | ↑autophagy | |
| Tseng et al., 2021 [78] | BM-MSCs | NM | I/R by unilateral nephrectomy | Rat | LC3B, Atg5, and Beclin 1 | ↓proinflammatory IL-1β, pro-apoptotic Bax, caspase 3 ↑autophagy-related LC3B, Atg5, and Beclin 1 | ↓macrophage infiltration ↓Tubular apoptosis, ↑tubular proliferation, |
| ID/Reference | Study Title | Conditions | Interventions | Status | Start and Complete Date | Link |
|---|---|---|---|---|---|---|
| NCT00733876 [105,106,107] | Allogeneic Multipotent Stromal Cell Treatment for Acute Kidney Injury Following Cardiac Surgery | Acute renal tubular necrosis | Biological: MSC administration | Completed | August 2008~October 2013 | https://clinicaltrials.gov/ct2/show/NCT00733876 (accessed on 1 June 2021) |
| NCT01275612 | Mesenchymal Stem Cells in Cisplatin-Induced Acute Renal Failure In Patients With Solid Organ Cancers | Solid tumors AKI | Biological: MSC infusion | Withdrawn | November 2010~19 March 2018 | https://clinicaltrials.gov/ct2/show/NCT01275612 (accessed on 1 June 2021) |
| NCT01602328 | A Study to Evaluate the Safety and Efficacy of AC607 for the Treatment of Kidney Injury in Cardiac Surgery Subjects | AKI | Biological: AC607 Biological: Vehicle Only | Terminated | June 2012~August 2014 | https://clinicaltrials.gov/ct2/show/NCT01602328 (accessed on 1 June 2021) |
| NCT03015623 | A Study of Cell Therapy for Subjects With Acute Kidney Injury Who Are Receiving Continuous Renal Replacement Therapy | AKI | Biological: SBI-101 Device: Sham | Active, not recruiting | June 2017~December 2021 | https://clinicaltrials.gov/ct2/show/NCT03015623 (accessed on 1 June 2021) |
| NCT04445220 | A Study of Cell Therapy in COVID-19 Subjects With Acute Kidney Injury Who Are Receiving Renal Replacement Therapy | COVID-19 AKI | Biological: SBI-101 | Recruiting | November 2020~December 2021 | https://clinicaltrials.gov/ct2/show/NCT04445220 (accessed on 1 June 2021) |
| NCT04194671 | Clinical Trial of Mesenchymal Stem Cells in the Treatment of Severe Acute Kidney Injury | AKI | Biological: MSC Other: Saline | Not yet recruiting | 31 January 2020~31 December 2022 | https://clinicaltrials.gov/ct2/show/NCT04194671 (accessed on 1 June 2021) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, P.-W.; Wu, B.-S.; Yang, C.-Y.; Lee, O.K.-S. Molecular Mechanisms of Mesenchymal Stem Cell-Based Therapy in Acute Kidney Injury. Int. J. Mol. Sci. 2021, 22, 11406. https://doi.org/10.3390/ijms222111406
Lee P-W, Wu B-S, Yang C-Y, Lee OK-S. Molecular Mechanisms of Mesenchymal Stem Cell-Based Therapy in Acute Kidney Injury. International Journal of Molecular Sciences. 2021; 22(21):11406. https://doi.org/10.3390/ijms222111406
Chicago/Turabian StyleLee, Pei-Wen, Bo-Sheng Wu, Chih-Yu Yang, and Oscar Kuang-Sheng Lee. 2021. "Molecular Mechanisms of Mesenchymal Stem Cell-Based Therapy in Acute Kidney Injury" International Journal of Molecular Sciences 22, no. 21: 11406. https://doi.org/10.3390/ijms222111406
APA StyleLee, P. -W., Wu, B. -S., Yang, C. -Y., & Lee, O. K. -S. (2021). Molecular Mechanisms of Mesenchymal Stem Cell-Based Therapy in Acute Kidney Injury. International Journal of Molecular Sciences, 22(21), 11406. https://doi.org/10.3390/ijms222111406