
Study on the bZIP-Type Transcription Factors NapA and RsmA in the Regulation of Intracellular Reactive Species Levels and Sterigmatocystin Production of Aspergillus nidulans


 , ,
, ,  , ,
, ,  and
and 
Abstract
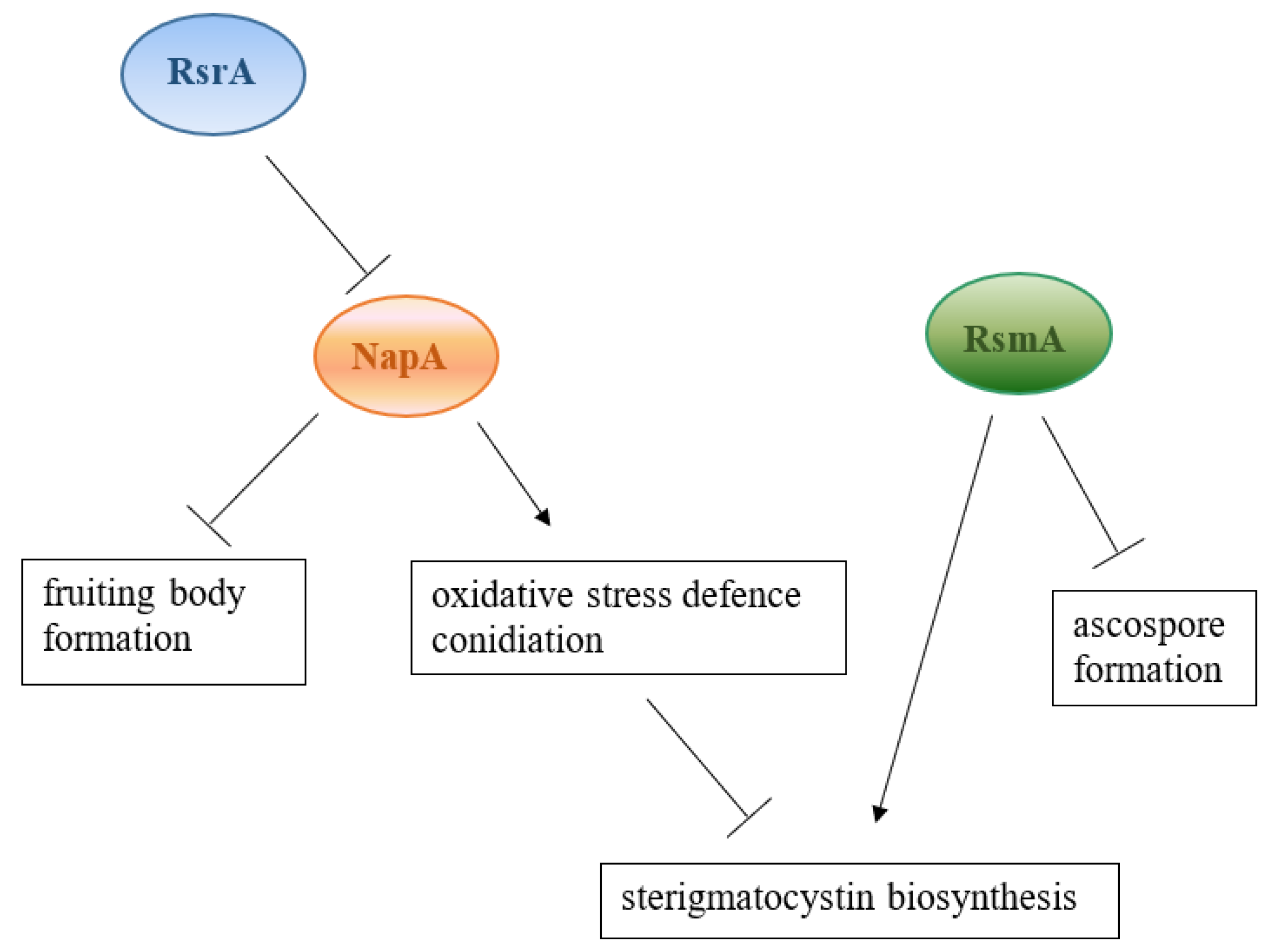
:1. Introduction
2. Results
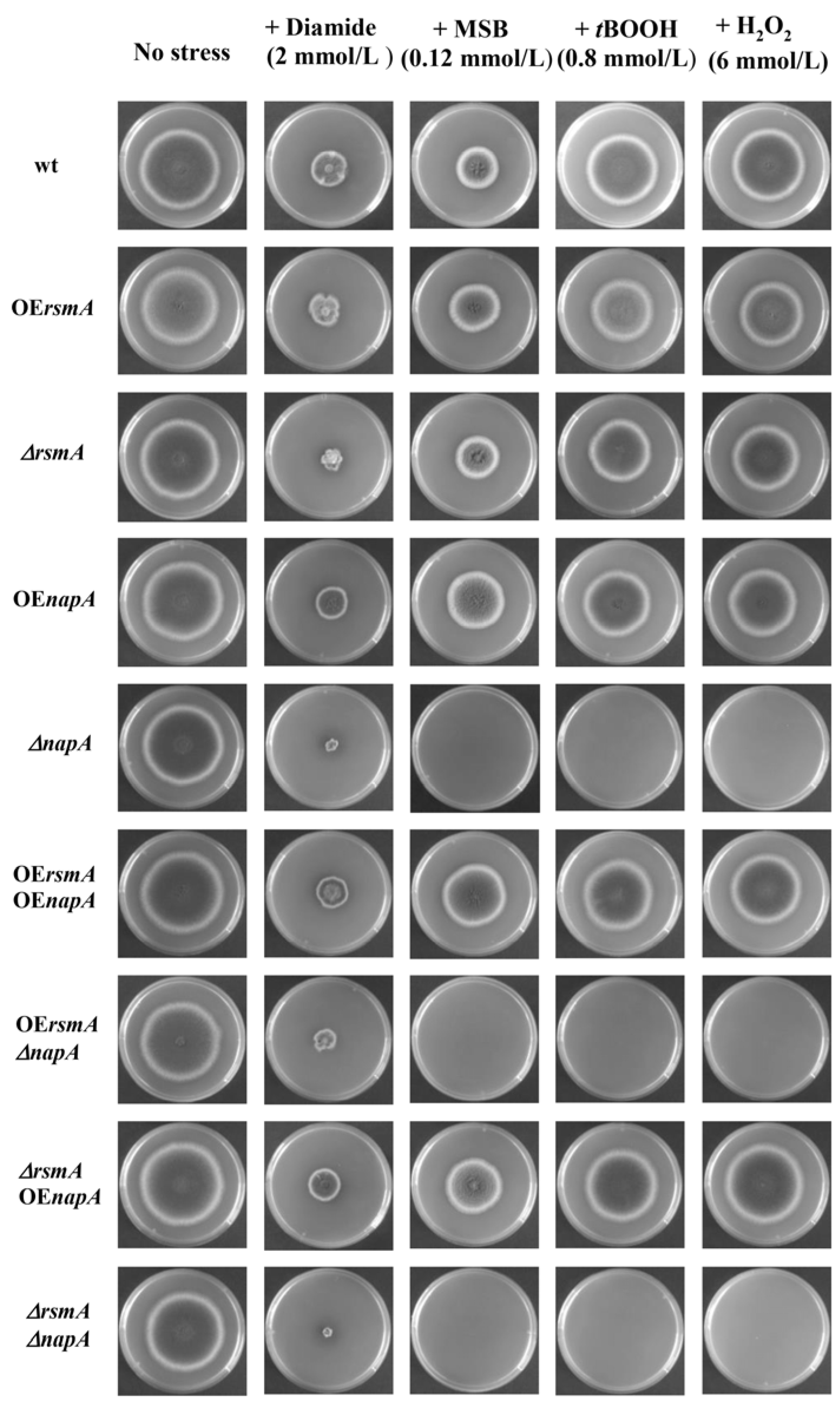
2.1. Stress Sensitivity Phenotypes of the RsmA and NapA Mutants
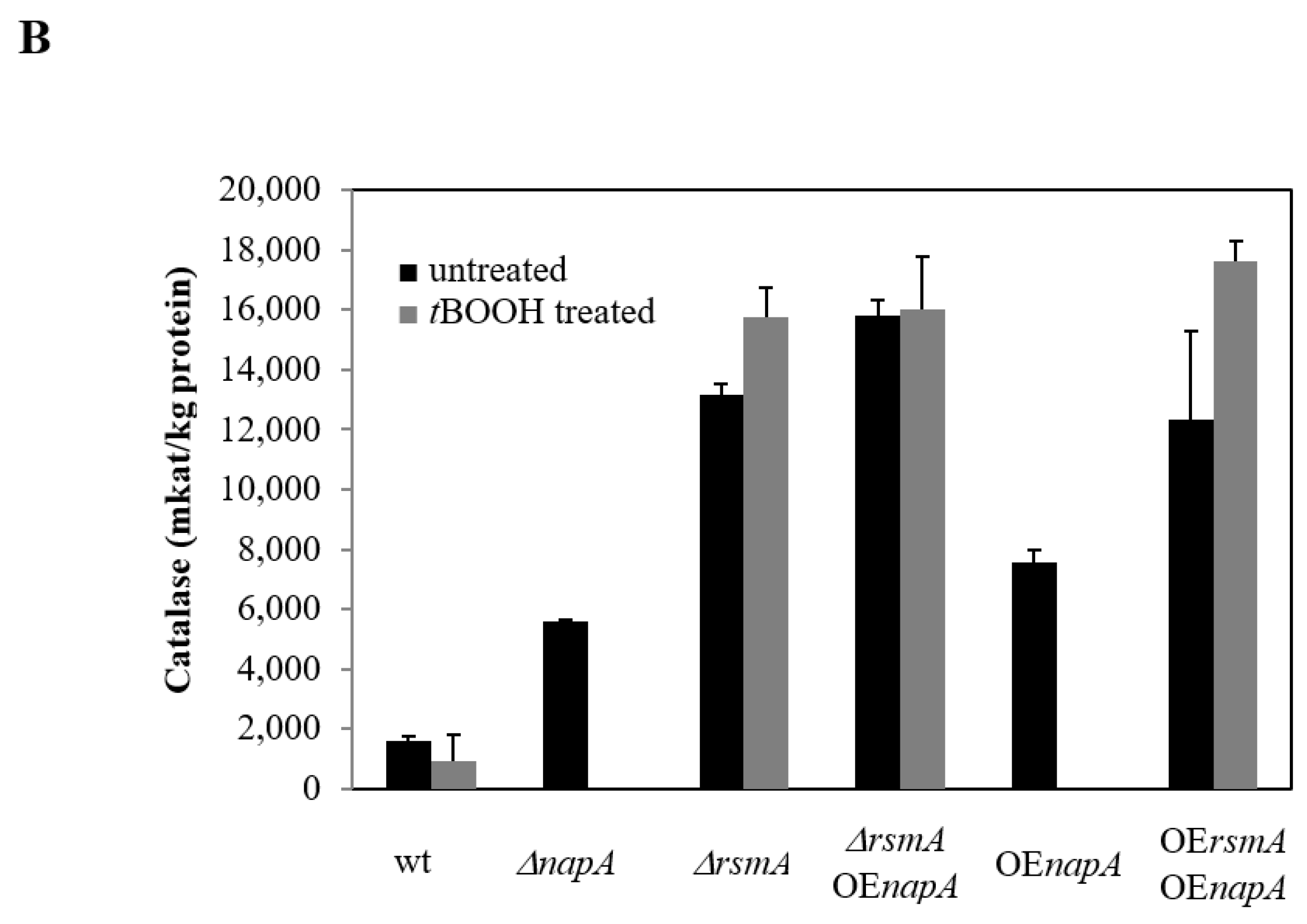
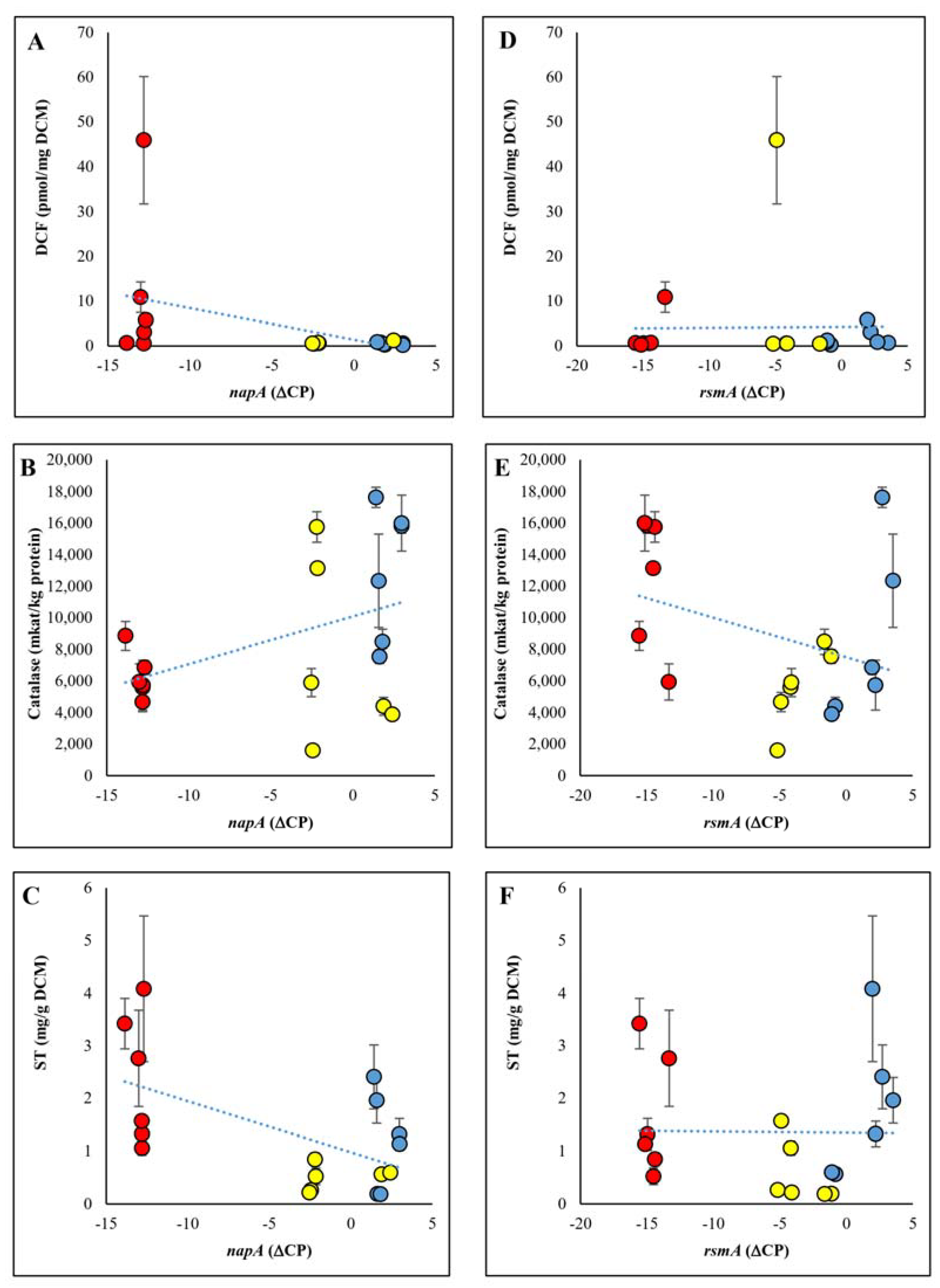
2.2. Biomass, Specific RS and Specific Catalase Enzyme Productions
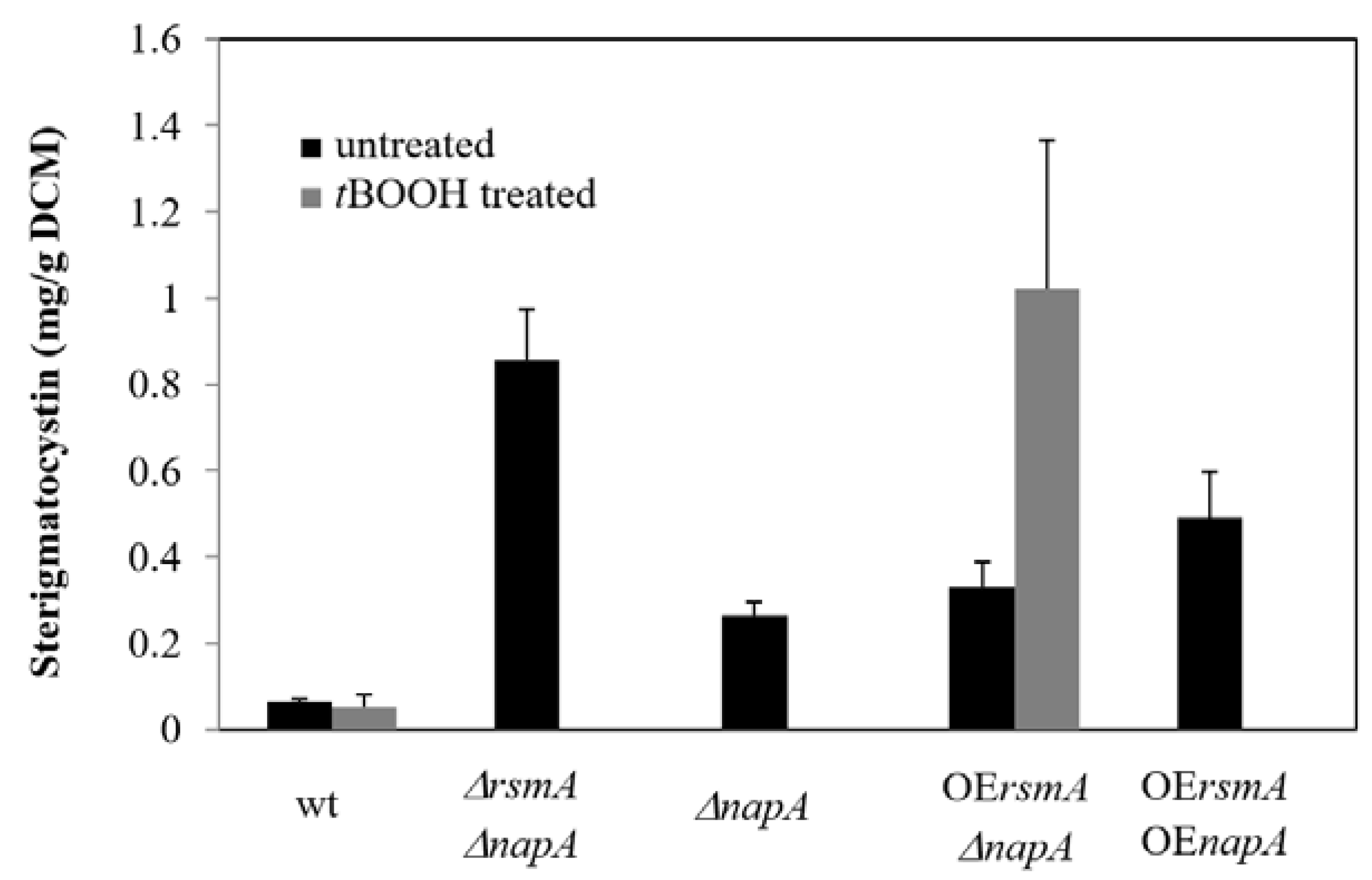
2.3. ST Production
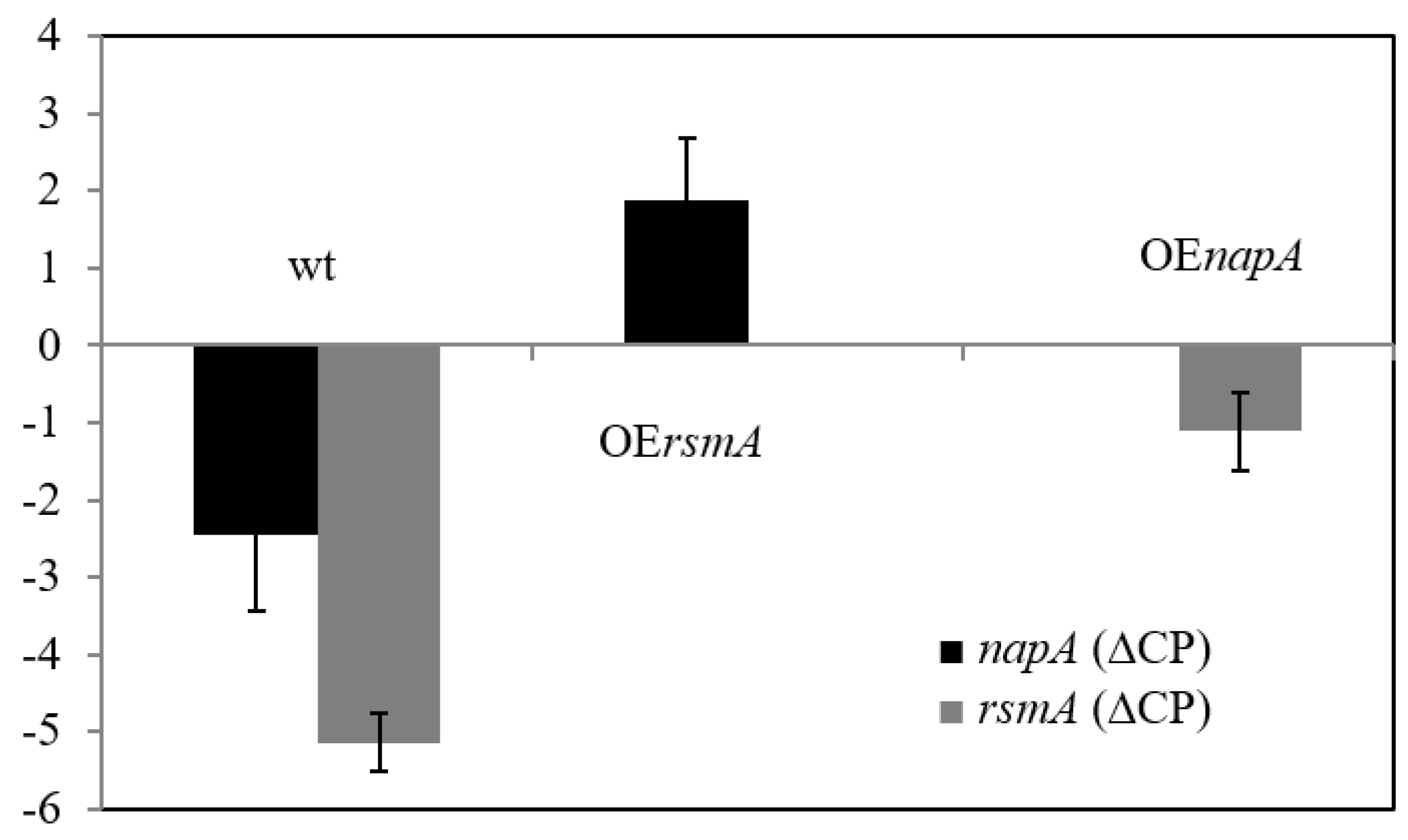
2.4. Expression Patterns
2.5. Interaction between NapA or RsmA Expressions and Specific Catalase Activities, DCF Formation, or ST Production
3. Discussion
4. Materials and Methods
4.1. Strains, Culture Media, and Growth Conditions
4.2. Oxidative Stress Sensitivity Experiments
4.3. Reactive Species Production and Catalase Activity
4.4. Sterigmatocystin Determination
4.5. rRT-PCR Assays
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rodrigues-Pousada, C.; Menezes, R.A.; Pimentel, C. The Yap family and its role in stress response. Yeast 2010, 27, 245–258. [Google Scholar] [CrossRef]
- Asano, Y.; Hagiwara, D.; Yamashino, T.; Mizuno, T. Characterization of the bZip-type transcription factor NapA with reference to oxidative stress response in Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2007, 71, 1800–1803. [Google Scholar] [CrossRef] [Green Version]
- Balázs, A.; Pócsi, I.; Hamari, Z.S.; Leiter, É.; Emri, T.; Miskei, M.; Oláh, J.; Tóth, V.; Hegedűs, N.; Prade, R.A.; et al. AtfA bZIP-type transcription factor regulates oxidative and osmotic stress responses in Aspergillus nidulans. Mol. Genet. Genom. 2010, 283, 289–303. [Google Scholar] [CrossRef]
- Hagiwara, D.; Asano, Y.; Yamashino, T.; Mizuno, T. Characterization of bZip-type transcription factor AtfA with reference to stress responses of conidia of Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2008, 72, 2756–2760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, J.; Kontoyiannis, D.P.; Calderone, R.; Li, D.; Ma, Y.; Wan, Z.; Li, R.; Liu, W. Afyap1, encoding a bZip transcriptional factor of Aspergillus fumigatus, contributes to oxidative stress response but is not essential to the virulence of this pathogen in mice immunosuppressed by cyclophosphamide and triamcinolone. Med. Mycol. 2008, 46, 773–782. [Google Scholar] [CrossRef] [Green Version]
- Reverberi, M.; Zjalic, S.; Punelli, F.; Ricelli, A.; Fabbri, A.A.; Fanelli, C. Apyap1 affects aflatoxin biosynthesis during Aspergillus parasiticus growth in maize seeds. Food Addit. Contam. 2007, 24, 1070–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reverberi, M.; Gazzetti, K.; Punelli, F.; Scarpari, M.; Zjalic, S.; Ricelli, A.; Fabbri, A.A.; Fanelli, C. Aoyap1 regulates OTA synthesis by controlling cell redox balance in Aspergillus ochraceus. Appl. Microbiol. Biotechnol. 2012, 95, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Roze, L.V.; Chanda, A.; Wee, J.; Awad, D.; Linz, J.E. Stress-related transcription factor AtfB integrates secondary metabolism with oxidative stress response in aspergilli. J. Biol. Chem. 2011, 286, 35137–35148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, C.; Li, J.; Glass, N.L. Exploring the bZIP transcription factor regulatory network in Neurospora crassa. Microbiology 2011, 157, 747–759. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Chen, Y.; Du, Y.; Dong, Y.; Guo, W.; Zhai, S.; Zhang, H.; Dong, S.; Zhang, Z.; Wang, Y.; et al. The bZIP transcription factor MoAP1 mediates the oxidative stress response and is critical for pathogenicity of the rice blast fungus Magnaporthe oryzae. PLoS Pathog. 2011, 7, e1001302. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Ru, Y.; Hong, L.; Zhu, Q.; Zuo, R.; Guo, X.; Wang, J.; Zhang, H.; Zheng, X.; Wang, P.; et al. System-wide characterization of bZIP transcription factor proteins involved in infection-related morphogenesis of Magnaporthe oryzae. Environ. Microbiol. 2014, 17, 1377–1396. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wu, F.; Liu, L.; Liu, X.; Chec, Y.; Keller, N.P.; Guo, L.; Yin, W.-B. The bZIP transcription factor PfZipA regulates secondary metabolism and oxidative stress response in the plant endophytic fungus Pestalotiopsis fici. Fungal Genet. Biol. 2015, 81, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.B.; Reinke, A.W.; Szilágyi, M.; Emri, T.; Chiang, Y.M.; Keating, A.E.; Pócsi, I.; Wang, C.C.C.; Keller, N.P. bZIP transcription factors affecting secondary metabolism, sexual development and stress responses in Aspergillus nidulans. Microbiology 2013, 159, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza-Martínez, A.E.; Lara-Rojas, F.; Sánchez, O.; Aguirre, J. NapA mediates a redox regulation of the antioxidant response, carbon utilization and development in Aspergillus nidulans. Front. Microbiol. 2017, 8, 516. [Google Scholar] [CrossRef]
- Thön, M.; Abdallah, Q.A.; Hortschansky, P.; Scharf, D.H.; Eisendle, M.; Haas, H.; Brakhage, A.A. The CCAAT-binding complex coordinates the oxidative stress response in eukaryotes. Nucleic Acids Res. 2010, 38, 1098–1113. [Google Scholar] [CrossRef]
- Zheng, H.; Kim, J.; Liew, M.; Yan, J.K.; Herrera, O.; Bok, J.-W.; Kelleher, N.L.; Keller, N.P.; Wang, Y. Redox metabolites signal polymicrobial biofilm development via the NapA oxidative stress cascade in Aspergillus. Curr. Biol. 2015, 25, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bok, J.-W.; Wiemann, P.; Garvey, G.S.; Lim, F.Y.; Haas, B.; Wortman, J.; Keller, N.P. Illumina identification of RsrA, a conserved C2H2 transcription factor coordinating the NapA mediated oxidative stress signaling pathway in Aspergillus. BMC Genom. 2014, 15, 1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaaban, M.I.; Bok, J.W.; Lauer, C.; Keller, N.P. Suppressor mutagenesis identifies a Velvet Complex Remediator of Aspergillus nidulans secondary metabolism. Eukaryot. Cell 2010, 9, 1816–1824. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.-B.; Amaike, S.; Wohlbach, D.J.; Gasch, A.P.; Chiang, Y.-M.; Wang, C.C.C.; Bok, J.-W.; Rohlfs, M.; Keller, N.P. An Aspergillus nidulans bZIP response pathway hardwired for defensive secondary metabolism operates through aflR. Mol. Microbiol. 2012, 83, 1024–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekonyela, R.; Palmer, J.M.; Bok, J.-W.; Jain, S.; Berthier, E.; Forseth, R.; Schroeder, F.; Keller, N.P. RsmA regulates Aspergillus fumigatus gliotoxin cluster metabolites including cyclo(L-Phe-L-Ser), a potential new diagnostic marker for invasive aspergillosis. PLoS ONE 2013, 8, e62591. [Google Scholar] [CrossRef]
- Wang, X.; Zha, W.; Liang, L.; Fasoyin, O.E.; Wu, L.; Wang, S. The bZIP transcription factor AflRsmA regulates aflatoxin B1 biosynthesis, oxidative stress response and sclerotium formation in Aspergillus flavus. Toxins 2020, 12, 271. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.Y.; Roze, L.V.; Linz, J.E. Oxidative stress-related transcription factors in the regulation of secondarymetabolism. Toxins 2013, 5, 683–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.Y.; Roze, L.V.; Wee, J.; Linz, J.E. Evidence that a transcription factor regulatory network coordinates oxidative stress response and secondary metabolism in aspergilli. Microbiologyopen 2013, 2, 144–160. [Google Scholar] [CrossRef]
- Reverberi, M.; Zjalic, S.; Ricelli, A.; Punelli, F.; Camera, E.; Fabbri, C.; Picardo, M.; Fanelli, C.; Fabbri, A.A. Modulation of antioxidant defense in Aspergillus parasiticus is involved in aflatoxin biosynthesis: A role for the ApyapA gene. Eukaryot. Cell 2008, 7, 988–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reverberi, M.; Ricelli, A.; Zjalic, S.; Fabbri, A.A.; Fanelli, C. Natural functions of mycotoxins and control of their biosynthesis in fungi. Appl. Microbiol. Biotechnol. 2010, 87, 899–911. [Google Scholar] [CrossRef]
- Jindrich, K.; Degnan, B.M. The diversification of the basic leucine zipper family in eukaryotes correlates with the evolution of multicellularity. BMC Evol. Biol. 2016, 16, 28. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.P.; Horta, M.A.C.; Goldman, G.H. Genetic interactions between Aspergillus fumigatus basic Leucine Zipper (bZIP) transcription factors AtfA, AtfB, AtfC, and AtfD. Front. Fungal Biol. 2021, 2, 632048. [Google Scholar] [CrossRef]
- Pontecorvo, G.; Roper, J.A.; Hemmons, L.M.; Macdonald, K.D.; Bufton, A.W.J. The genetics of Aspergillus nidulans. In Advances in Genetics; Elsevier: Amsterdam, The Netherlands, 1953; Volume 5, pp. 141–238. [Google Scholar]
- Barratt, R.W.; Johnson, G.B.; Ogata, W.N. Wild-type and mutant stocks of Aspergillus nidulans. Genetics 1965, 52, 233–246. [Google Scholar] [CrossRef]
- Tsitsigiannis, D.I.; Zarnowski, R.; Keller, N.P. The lipid body protein, PpoA, coordinates sexual and asexual sporulation in Aspergillus nidulans. J. Biol. Chem. 2004, 279, 11344–11353. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Chapter 5. Measurement of Reactive Species. In Free Radicals in Biology and Medicine, 4th ed.; Oxford University Press: Oxford, UK, 2007; pp. 268–330. [Google Scholar]
- Emri, T.; Pócsi, I.; Szentirmai, A. Glutathione metabolism and protection against oxidative stress caused by peroxides in Penicillium chrysogenum. Free Rad. Biol. Med. 1997, 23, 809–814. [Google Scholar] [CrossRef]
- Emri, T.; Pócsi, I.; Szentirmai, A. Analysis of the oxidative stress response of Penicillium chrysogenum to menadione. Free Rad. Res. 1999, 30, 125–132. [Google Scholar] [CrossRef]
- Roggenkamp, R.; Sahm, H.; Wagner, F. Microbial assimilation of methanol induction and function of catalase in Candida boidinii. FEBS Lett. 1974, 41, 283–286. [Google Scholar] [CrossRef] [Green Version]
- Peterson, G.L. Determination of total protein. Meth. Enzymol. 1983, 91, 86–105. [Google Scholar]
- Shwab, E.K.; Bok, J.W.; Tribus, M.; Galehr, J.; Graessle, S.; Keller, N.P. Histone deacetylase activity regulates chemical diversity in Aspergillus. Eukaryot. Cell 2007, 6, 1656–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomczynski, P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. BioTechniques 1993, 15, 532–534, 536–537. [Google Scholar] [PubMed]
- Emri, T.; Szarvas, V.; Orosz, E.; Antal, K.; Park, H.; Han, K.H.; Yu, J.-H.; Pócsi, I. Core oxidative stress response in Aspergillus nidulans. BMC Genom. 2015, 16, 478. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Genotype | Reference |
|---|---|---|
| RDIT 9.32 | wild type | [30] |
| RWY 2.12 | gpdA(p)::rsmA::A. fumigatus pyrG | [19] |
| RWY 8.5 | ΔrsmA::pyrG A. parasiticus | [13] |
| RWY 17.3 | A. fumigatus pyroA::gpdA(p)::napA, pyroA4 | [13] |
| RWY 10.3 | ΔnapA::pyroA A. fumigatus | [13] |
| TWY7.3 | pyrG89; ΔnapA::pyroA A. fumigatus, pyroA4, ΔnkuA::argB | [13] |
| TMS6.30 | pyrG89; ∆rsmA::pyrG A. parasiticus, pyroA4 | [18] |
| TWY13.15 | pyrG89; gpdA(p)::napA:: pyroA A. fumigatus, pyroA4, ΔnkuA::argB | [13] |
| RWY16.47 | AfumpyrG::gpdA(p)::rsmA, ∆aflR::argB, pyroA4, TrpC801 | [19] |
| RWY33.2 | gpdA(p)::napA:: pyroA A. fumigatus, AfumpyrG::gpdA(p)::rsmA From cross of RWY16.47 X TWY13.15 | This study |
| RWY34.30 | A. fumigatus pyrG::gpdA(p)::rsmA,∆napA:: A. fumigatus pyroA From cross of RWY16.47 X TWY7.3 | This study |
| RWY35.5 | ΔrsmA::A. parasiticus pyrG, A. fumigatus pyroA::gpdA(p)::napA From cross of TMS6.30 X TWY13.15 | This study |
| RWY6.2 | ∆napA::A. fumigatus pyroA, pyroA4, pyrG89,ΔrsmA::A. parasiticus pyrG From cross of TWY7.3 X TMS6.30 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bákány, B.; Yin, W.-B.; Dienes, B.; Nagy, T.; Leiter, É.; Emri, T.; Keller, N.P.; Pócsi, I. Study on the bZIP-Type Transcription Factors NapA and RsmA in the Regulation of Intracellular Reactive Species Levels and Sterigmatocystin Production of Aspergillus nidulans. Int. J. Mol. Sci. 2021, 22, 11577. https://doi.org/10.3390/ijms222111577
Bákány B, Yin W-B, Dienes B, Nagy T, Leiter É, Emri T, Keller NP, Pócsi I. Study on the bZIP-Type Transcription Factors NapA and RsmA in the Regulation of Intracellular Reactive Species Levels and Sterigmatocystin Production of Aspergillus nidulans. International Journal of Molecular Sciences. 2021; 22(21):11577. https://doi.org/10.3390/ijms222111577
Chicago/Turabian StyleBákány, Bernadett, Wen-Bing Yin, Beatrix Dienes, Tibor Nagy, Éva Leiter, Tamás Emri, Nancy P. Keller, and István Pócsi. 2021. "Study on the bZIP-Type Transcription Factors NapA and RsmA in the Regulation of Intracellular Reactive Species Levels and Sterigmatocystin Production of Aspergillus nidulans" International Journal of Molecular Sciences 22, no. 21: 11577. https://doi.org/10.3390/ijms222111577
APA StyleBákány, B., Yin, W. -B., Dienes, B., Nagy, T., Leiter, É., Emri, T., Keller, N. P., & Pócsi, I. (2021). Study on the bZIP-Type Transcription Factors NapA and RsmA in the Regulation of Intracellular Reactive Species Levels and Sterigmatocystin Production of Aspergillus nidulans. International Journal of Molecular Sciences, 22(21), 11577. https://doi.org/10.3390/ijms222111577