
Improved Production of Recombinant Myrosinase in Pichia pastoris

 and
and
Abstract
:1. Introduction
2. Results and Discussion
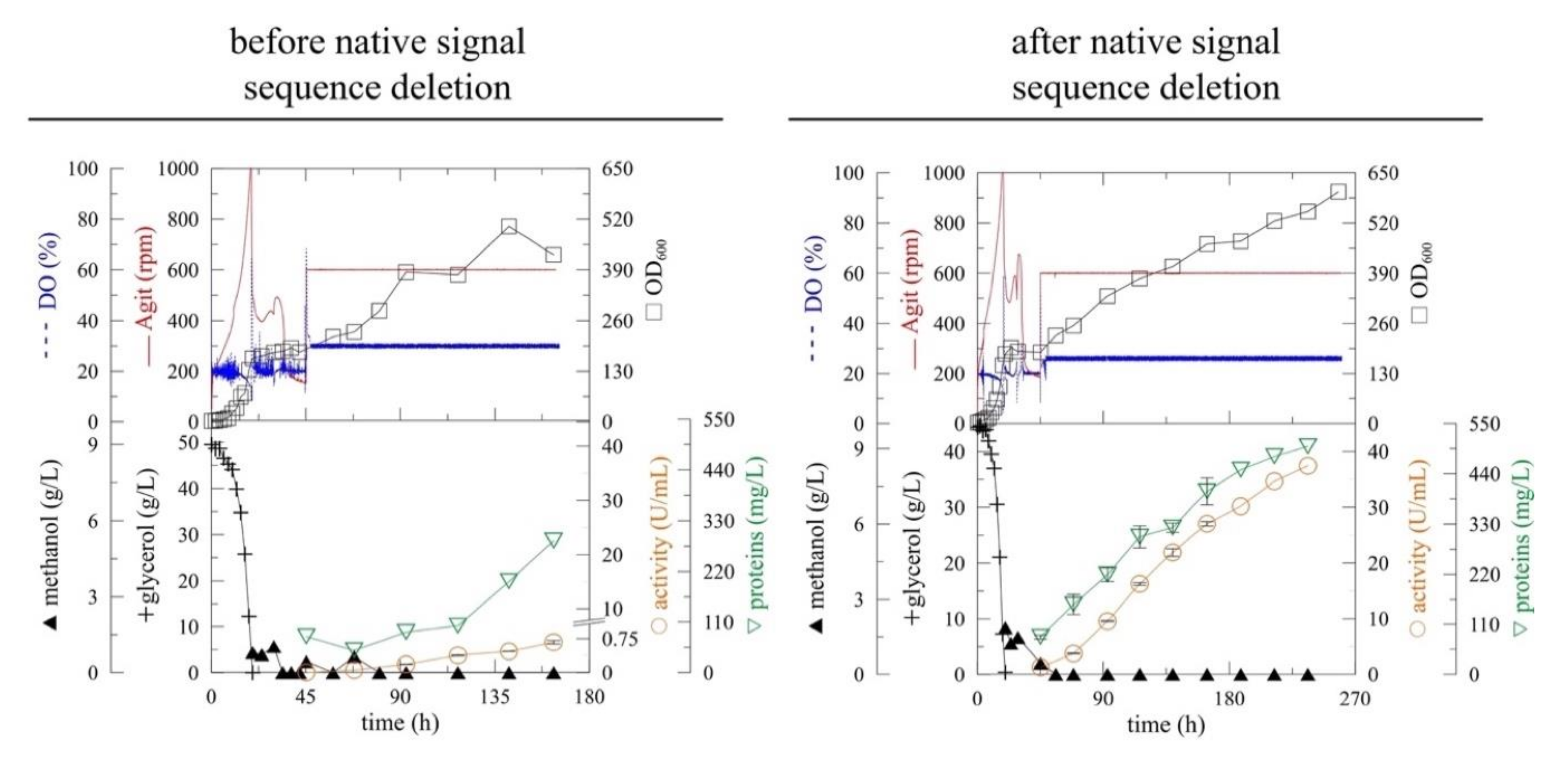
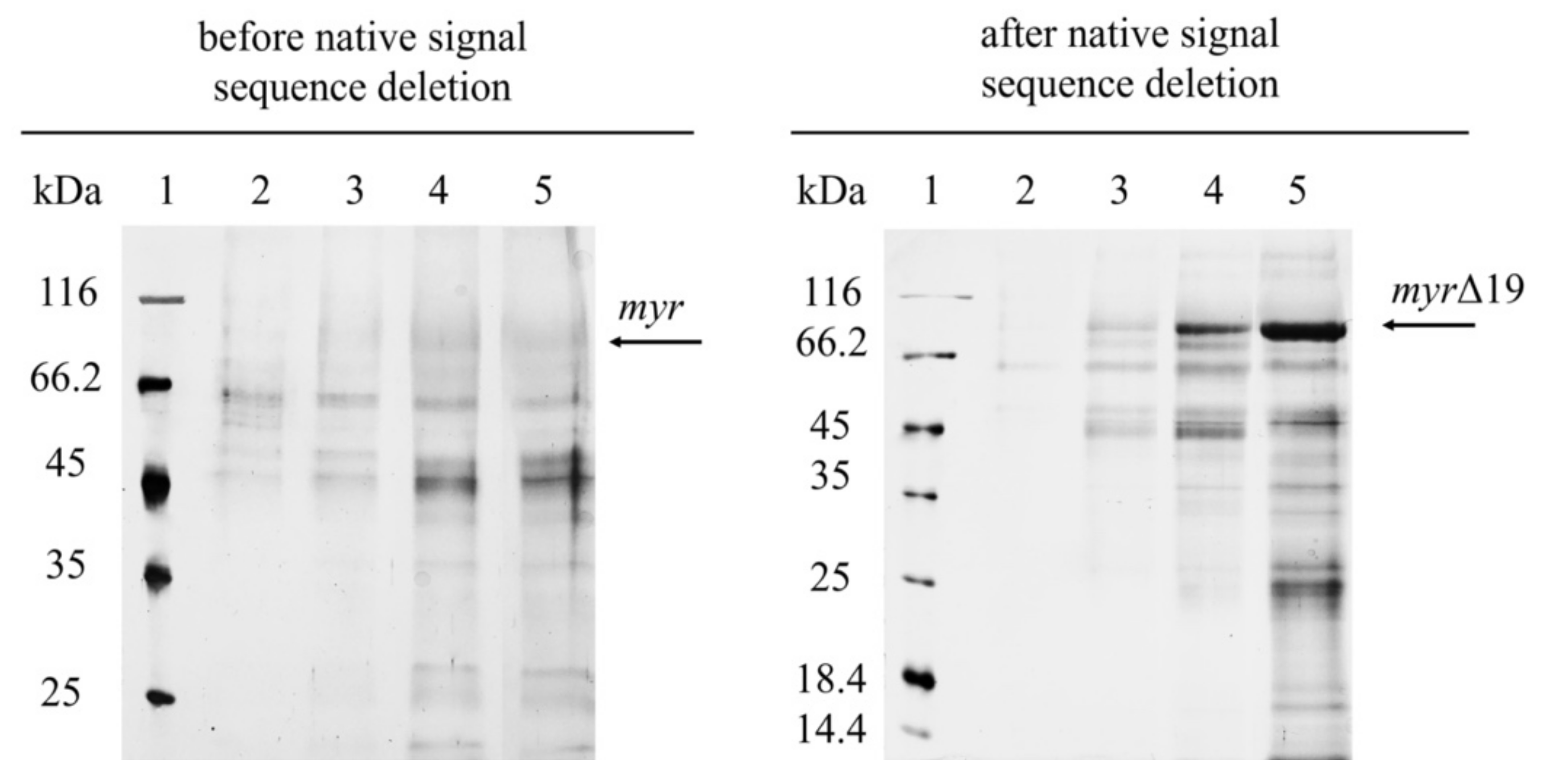
2.1. The Overexpression of Myrosinase without Native Signal Peptide
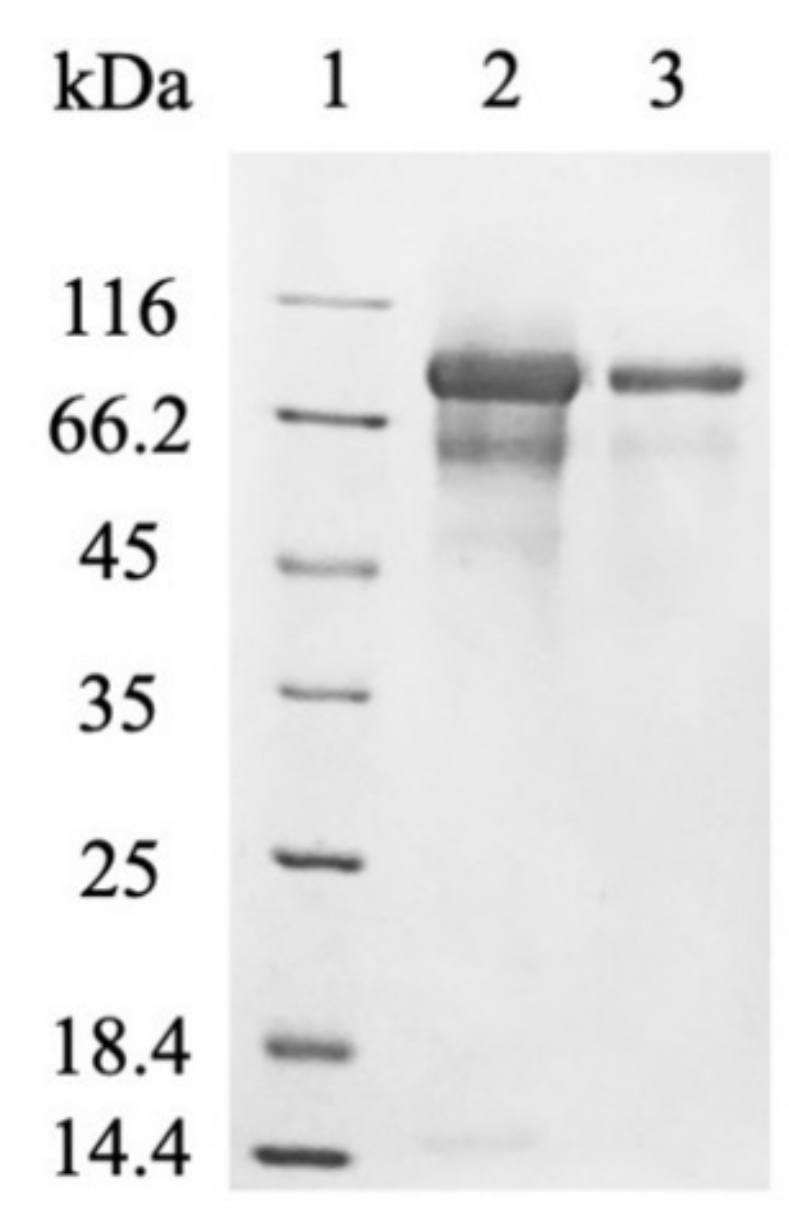
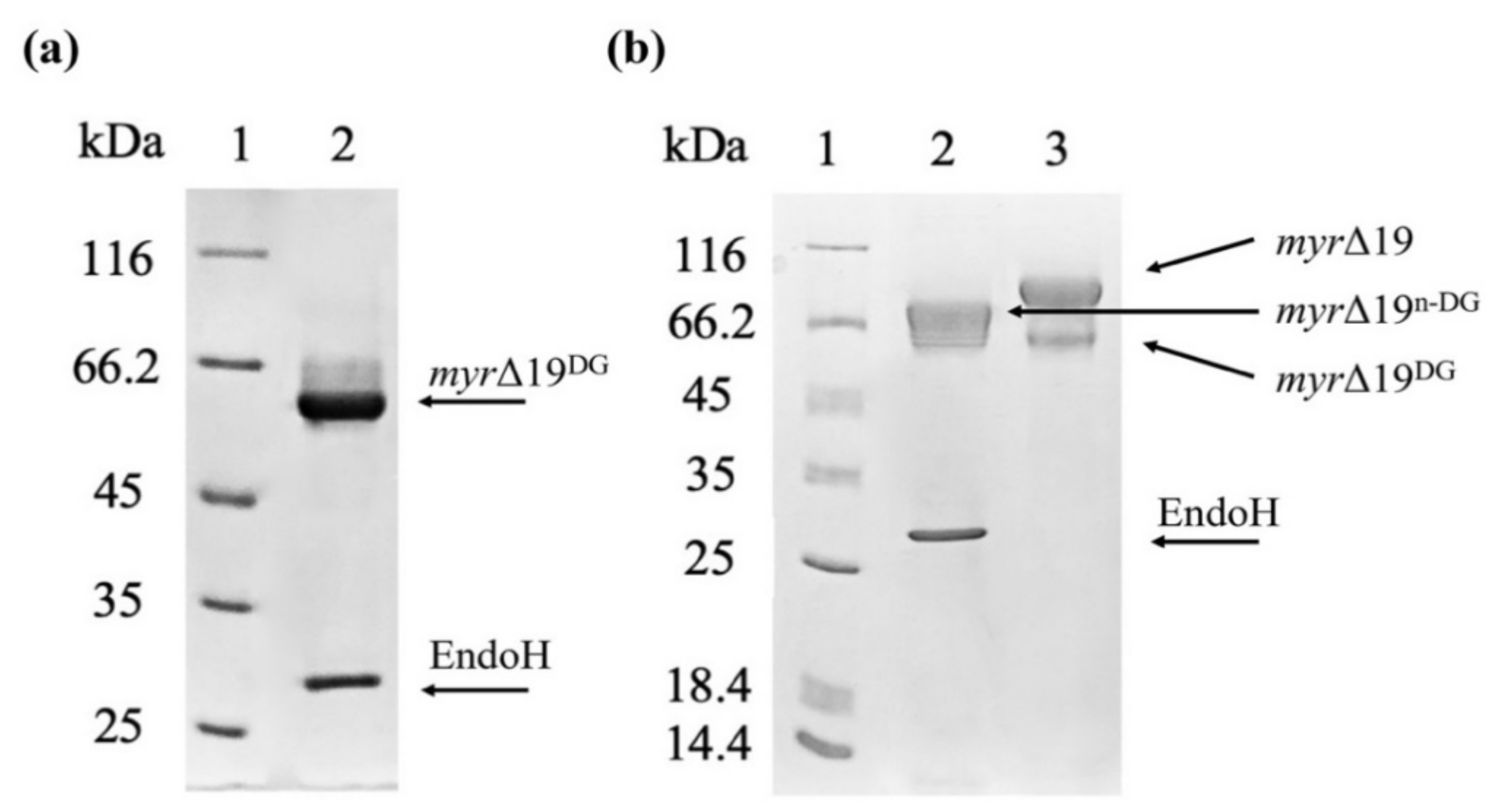
2.2. The Purification of Myrosinase
2.3. The Hyper-Glycosylation of Recombinant Myrosinase
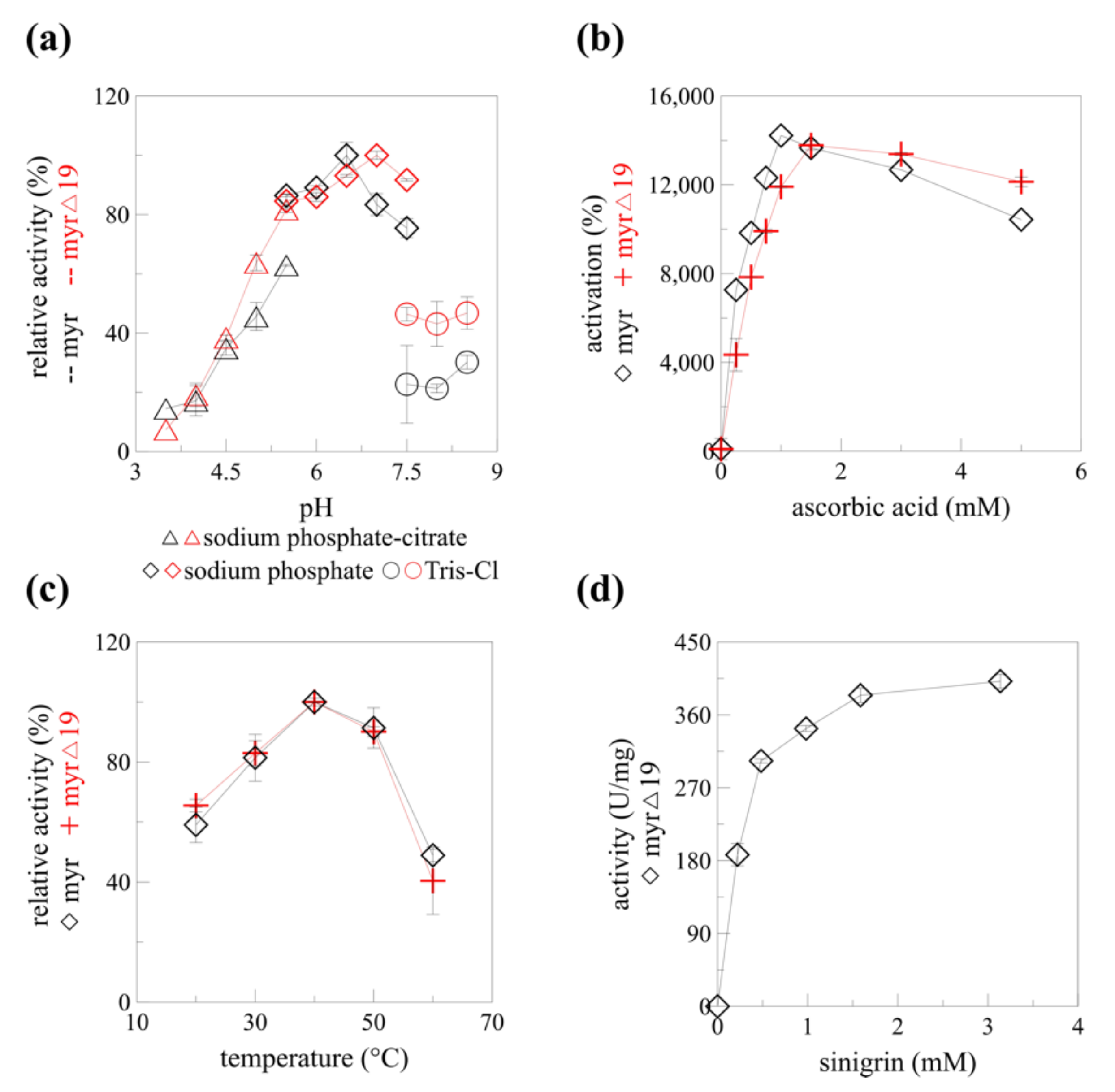
2.4. The Catalytic Properties of Produced Myrosinase
3. Materials and Methods
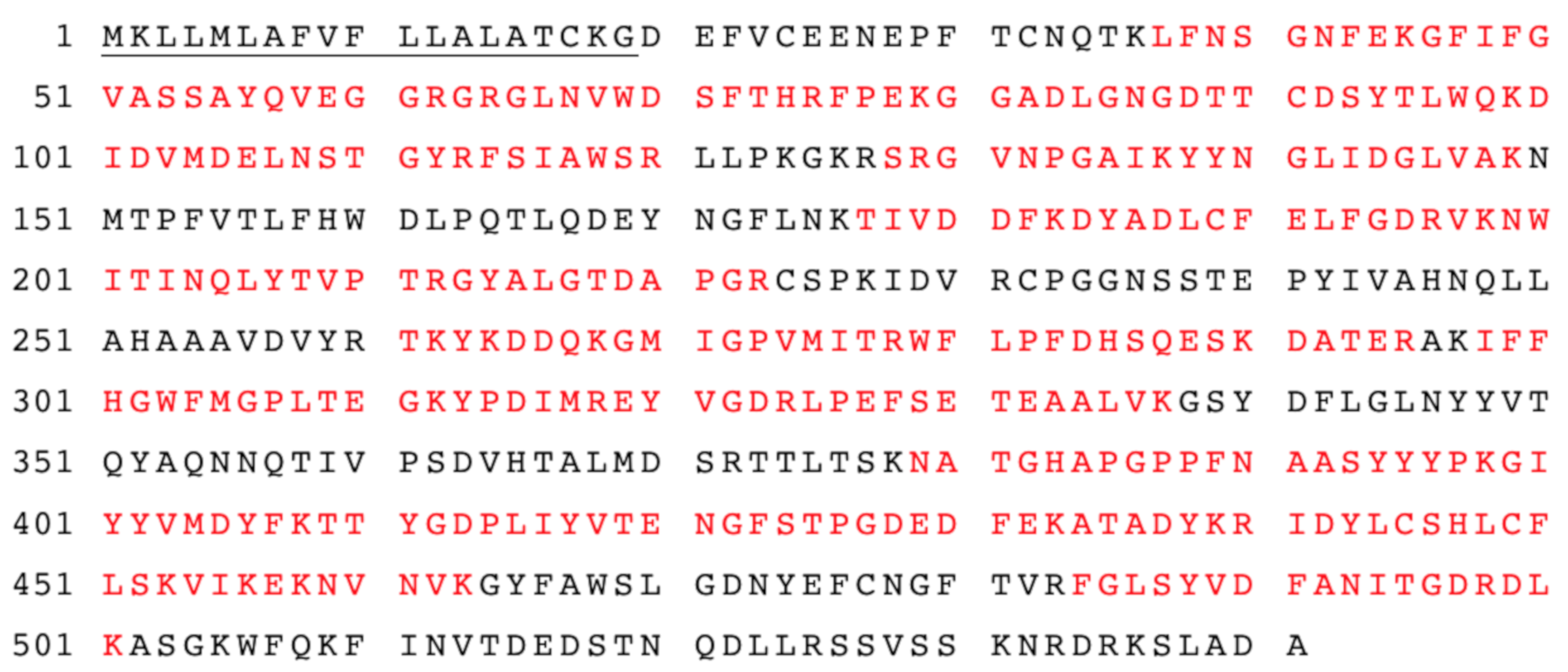
3.1. Signal Peptide Deletion and Recombinant Strain Preparation
3.2. Overexpression of Recombinant Myrosinase (myr-Δ19)
3.3. Purification of Recombinant Myrosinase (myr-Δ19)
3.4. Deglycosylation of Myrosinase (myr-Δ19)
3.5. Myrosinase Activity Assay
3.6. Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia pastoris: A Highly Successful Expression System for Optimal Synthesis of Heterologous Proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Miralles, N.; Domingo-Espín, J.; Corchero, J.; Vázquez, E.; Villaverde, A. Microbial Factories for Recombinant Pharmaceuticals. Microb. Cell Factories 2009, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Rong, Y.; Wang, Y.; Kong, D.; Wang, P.G.; Chen, M.; Kong, Y. Homogeneous Production and Characterization of Recombinant N-GlcNAc-Protein in Pichia pastoris. Microb. Cell Factories 2020, 19, 7. [Google Scholar] [CrossRef]
- Demain, A.L.; Vaishnav, P. Production of Recombinant Proteins by Microbes and Higher Organisms. Biotechnol. Adv. 2009, 27, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Cereghino, J.L.; Cregg, J.M. Heterologous Protein Expression in the Methylotrophic Yeast. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef]
- Radoman, B.; Grünwald-Gruber, C.; Schmelzer, B.; Zavec, D.; Gasser, B.; Altmann, F.; Mattanovich, D. The Degree and Length of O-Glycosylation of Recombinant Proteins Produced in Pichia Pastoris Depends on the Nature of the Protein and the Process Type. Biotechnol. J. 2021, 16, 2000266. [Google Scholar] [CrossRef] [PubMed]
- Baeshen, M.N.; Bouback, T.A.F.; Alzubaidi, M.A.; Bora, R.S.; Alotaibi, M.A.T.; Alabbas, O.T.O.; Alshahrani, S.M.; Aljohani, A.A.M.; Munshi, R.A.A.; Al-Hejin, A.; et al. Expression and Purification of C-Peptide Containing Insulin Using Pichia pastoris Expression System. BioMed Res. Int. 2016, 2016, 3423685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eissazadeh, S.; Moeini, H.; Dezfouli, M.G.; Heidary, S.; Nelofer, R.; Abdullah, M.P. Production of Recombinant Human Epidermal Growth Factor in Pichia pastoris. Braz. J. Microbiol. 2017, 48, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, J.; Sun, H.; Li, S.; Chen, T.; Liu, G.; Dyson, P. High-Level Heterologous Production and Functional Secretion by Recombinant Pichia pastoris of the Shortest Proline-Rich Antibacterial Honeybee Peptide Apidaecin. Sci. Rep. 2017, 7, 14543. [Google Scholar] [CrossRef] [Green Version]
- Navarro, S.L.; Li, F.; Lampe, J.W. Mechanisms of Action of Isothiocyanates in Cancer Chemoprevention: An Update. Food Funct. 2011, 2, 579. [Google Scholar] [CrossRef] [PubMed]
- Mitsiogianni, M.; Koutsidis, G.; Mavroudis, N.; Trafalis, D.T.; Botaitis, S.; Franco, R.; Zoumpourlis, V.; Amery, T.; Galanis, A.; Pappa, A.; et al. The Role of Isothiocyanates as Cancer Chemo-Preventive, Chemo-Therapeutic and Anti-Melanoma Agents. Antioxidants 2019, 8, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, R.; Vyas, D. Myrosinase: Insights on Structural, Catalytic, Regulatory, and Environmental Interactions. Crit. Rev. Biotechnol. 2019, 39, 508–523. [Google Scholar] [CrossRef]
- Rosenbergová, Z.; Kántorová, K.; Šimkovič, M.; Breier, A.; Rebroš, M. Optimisation of Recombinant Myrosinase Production in Pichia pastoris. Int. J. Mol. Sci. 2021, 22, 3677. [Google Scholar] [CrossRef] [PubMed]
- Thangstad, O.P.; Winge, P.; Husebye, H.; Bones, A. The Myrosinase (Thioglucoside Glucohydrolase) Gene Family in Brassicaceae. Plant Mol. Biol. 1993, 23, 511–524. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Nielsen, H.; Engelbrecht, J.; Brunak, S.; von Heijne, G. Identification of Prokaryotic and Eukaryotic Signal Peptides and Prediction of Their Cleavage Sites. Protein Eng. Des. Sel. 1997, 10, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, D.; Chakrabarty, R.; Bejai, S.; Zhang, J.; Rask, L.; Meijer, J. Myrosinases from Root and Leaves of Arabidopsis Thaliana Have Different Catalytic Properties. Phytochemistry 2009, 70, 1345–1354. [Google Scholar] [CrossRef]
- Wang, M.; Li, D.; Sun, X.; Zhu, Y.J.; Nong, H.; Zhang, J. Characterization of a Root-Specific β-Thioglucoside Glucohydrolase Gene in Carica Papaya and Its Recombinant Protein Expressed in Pichia pastoris. Plant Sci. 2009, 177, 716–723. [Google Scholar] [CrossRef]
- Yang, M.; Teymorian, S.; Olivares, P.; Murthy, P.P.N. Extracellular Expression of Alkaline Phytase in Pichia pastoris: Influence of Signal Peptides, Promoters and Growth Medium. Biotechnol. Rep. 2015, 6, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Massahi, A.; Çalık, P. Endogenous Signal Peptides in Recombinant Protein Production by Pichia pastoris: From in-Silico Analysis to Fermentation. J. Theor. Biol. 2016, 408, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Neiers, F.; Belloir, C.; Poirier, N.; Naumer, C.; Krohn, M.; Briand, L. Comparison of Different Signal Peptides for the Efficient Secretion of the Sweet-Tasting Plant Protein Brazzein in Pichia pastoris. Life 2021, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Härtel, F.V.; Brandt, A. Characterization of a Brassica Napus Myrosinase Expressed and Secreted by Pichia pastoris. Protein Expr. Purif. 2002, 24, 221–226. [Google Scholar] [CrossRef]
- Andelova, N.; Waczulikova, I.; Talian, I.; Sykora, M.; Ferko, M. MPTP Proteins Regulated by Streptozotocin-Induced Diabetes Mellitus Are Effectively Involved in the Processes of Maintaining Myocardial Metabolic Adaptation. Int. J. Mol. Sci. 2020, 21, 2622. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Tokuhisa, J.G.; Bevan, D.R.; Esen, A. Properties of β-Thioglucoside Hydrolases (TGG1 and TGG2) from Leaves of Arabidopsis Thaliana. Plant Sci. 2012, 191–192, 82–92. [Google Scholar] [CrossRef]
- Blanchard, V.; Gadkari, R.A.; George, A.V.E.; Roy, S.; Gerwig, G.J.; Leeflang, B.R.; Dighe, R.R.; Boelens, R.; Kamerling, J.P. High-Level Expression of Biologically Active Glycoprotein Hormones in Pichia pastoris Strains—Selection of Strain GS115, and Not X-33, for the Production of Biologically Active N-Glycosylated 15N-Labeled PhCG. Glycoconj. J. 2008, 25, 245–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmeister, W.P.; Cottaz, S.; Driguez, H.; Iori, R.; Palmieri, S.; Henrissat, B. The Crystal Structures of Sinapis Alba Myrosinase and a Covalent Glycosyl–Enzyme Intermediate Provide Insights into the Substrate Recognition and Active-Site Machinery of an S-Glycosidase. Structure 1997, 5, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, S.; Thamilarasan, S.K.; Park, J.-I.; Chung, M.-Y.; Nou, I.-S. Molecular Modeling of Myrosinase from Brassica Oleracea: A Structural Investigation of Sinigrin Interaction. Genes 2015, 6, 1315–1329. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Kumar, S.; Sangwan, S.; Yadav, I.S.; Yadav, R. Protein Modeling and Active Site Binding Mode Interactions of Myrosinase–Sinigrin in Brassica Juncea—An in Silico Approach. J. Mol. Graph. Model. 2011, 29, 740–746. [Google Scholar] [CrossRef]
- Liebminger, E.; Grass, J.; Jez, J.; Neumann, L.; Altmann, F.; Strasser, R. Myrosinases TGG1 and TGG2 from Arabidopsis Thaliana Contain Exclusively Oligomannosidic N-Glycans. Phytochemistry 2012, 84, 24–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretthauer, R.K.; Castellino, F.J. Glycosylation of Pichia pastoris Derived Proteins. Biotechnol. Appl. Biochem. 1999, 30, 3–193. [Google Scholar]
- Pérez de los Santos, A.I.; Cayetano-Cruz, M.; Gutiérrez-Antón, M.; Santiago-Hernández, A.; Plascencia-Espinosa, M.; Farrés, A.; Hidalgo-Lara, M.E. Improvement of Catalytical Properties of Two Invertases Highly Tolerant to Sucrose after Expression in Pichia pastoris. Effect of Glycosylation on Enzyme Properties. Enzym. Microb. Technol. 2016, 83, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Dotsenko, A.S.; Gusakov, A.V.; Rozhkova, A.M.; Sinitsyna, O.A.; Nemashkalov, V.A.; Sinitsyn, A.P. Effect of N-Linked Glycosylation on the Activity and Other Properties of Recombinant Endoglucanase IIa (Cel5A) from Penicillium verruculosum. Protein Eng. Des. Sel. 2016, 29, 495–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonzom, C.; Hüttner, S.; Mirgorodskaya, E.; Chong, S.-L.; Uthoff, S.; Steinbüchel, A.; Verhaert, R.M.D.; Olsson, L. Glycosylation Influences Activity, Stability and Immobilization of the Feruloyl Esterase 1a from Myceliophthora thermophila. AMB Express 2019, 9, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerstorferová, D.; Fliedrová, B.; Halada, P.; Marhol, P.; Křen, V.; Weignerová, L. Recombinant α-l-Rhamnosidase from Aspergillus Terreus in Selective Trimming of Rutin. Process Biochem. 2012, 47, 828–835. [Google Scholar] [CrossRef]
- Burmeister, W.P.; Cottaz, S.; Rollin, P.; Vasella, A.; Henrissat, B. High Resolution X-Ray Crystallography Shows That Ascorbate Is a Cofactor for Myrosinase and Substitutes for the Function of the Catalytic Base. J. Biol. Chem. 2000, 275, 39385–39393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikita, M.; Fahey, J.W.; Golden, T.R.; David Holtzclaw, W.; Talalay, P. An Unusual Case of ‘Uncompetitive Activation’ by Ascorbic Acid: Purification and Kinetic Properties of a Myrosinase from Raphanus Sativus Seedlings. Biochem. J. 1999, 341, 725–732. [Google Scholar] [CrossRef]
- Lin-Cereghino, J.; Wong, W.W.; Xiong, S.; Giang, W.; Luong, L.T.; Vu, J.; Johnson, S.D.; Lin-Cereghino, G.P. Condensed Protocol for Competent Cell Preparation and Transformation of the Methylotrophic Yeast Pichia pastoris. BioTechniques 2005, 38, 44–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markošová, K.; Weignerová, L.; Rosenberg, M.; Křen, V.; Rebroš, M. Upscale of Recombinant α-L-Rhamnosidase Production by Pichia pastoris MutS Strain. Front. Microbiol. 2015, 6, 1140. [Google Scholar] [CrossRef]
- Tsao, R.; Yu, Q.; Potter, J.; Chiba, M. Direct and Simultaneous Analysis of Sinigrin and Allyl Isothiocyanate in Mustard Samples by High-Performance Liquid Chromatography. J. Agric. Food Chem. 2002, 50, 4749–4753. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Maximal Specific Productivity 1 (U/L/h) | Activity 1 (U/mL) | Maximal Activity (U/mL) | Total Proteins 1 (mg/L) |
|---|---|---|---|---|
| myr | 4.1 ± 0.2 | 0.67 ± 0.04 | 0.67 1 ± 0.04 | 286.5 ± 21.8 |
| myr-Δ19 | 164.8 ± 3.0 | 27 ± 0.4 | 37. 4 2 ± 0.55 | 422.2 ± 18.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosenbergová, Z.; Hegyi, Z.; Ferko, M.; Andelová, N.; Rebroš, M. Improved Production of Recombinant Myrosinase in Pichia pastoris. Int. J. Mol. Sci. 2021, 22, 11889. https://doi.org/10.3390/ijms222111889
Rosenbergová Z, Hegyi Z, Ferko M, Andelová N, Rebroš M. Improved Production of Recombinant Myrosinase in Pichia pastoris. International Journal of Molecular Sciences. 2021; 22(21):11889. https://doi.org/10.3390/ijms222111889
Chicago/Turabian StyleRosenbergová, Zuzana, Zuzana Hegyi, Miroslav Ferko, Natália Andelová, and Martin Rebroš. 2021. "Improved Production of Recombinant Myrosinase in Pichia pastoris" International Journal of Molecular Sciences 22, no. 21: 11889. https://doi.org/10.3390/ijms222111889
APA StyleRosenbergová, Z., Hegyi, Z., Ferko, M., Andelová, N., & Rebroš, M. (2021). Improved Production of Recombinant Myrosinase in Pichia pastoris. International Journal of Molecular Sciences, 22(21), 11889. https://doi.org/10.3390/ijms222111889