
Cold-Inducible RNA-Binding Protein but Not Its Antisense lncRNA Is a Direct Negative Regulator of Angiogenesis In Vitro and In Vivo via Regulation of the 14q32 angiomiRs—microRNA-329-3p and microRNA-495-3p

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
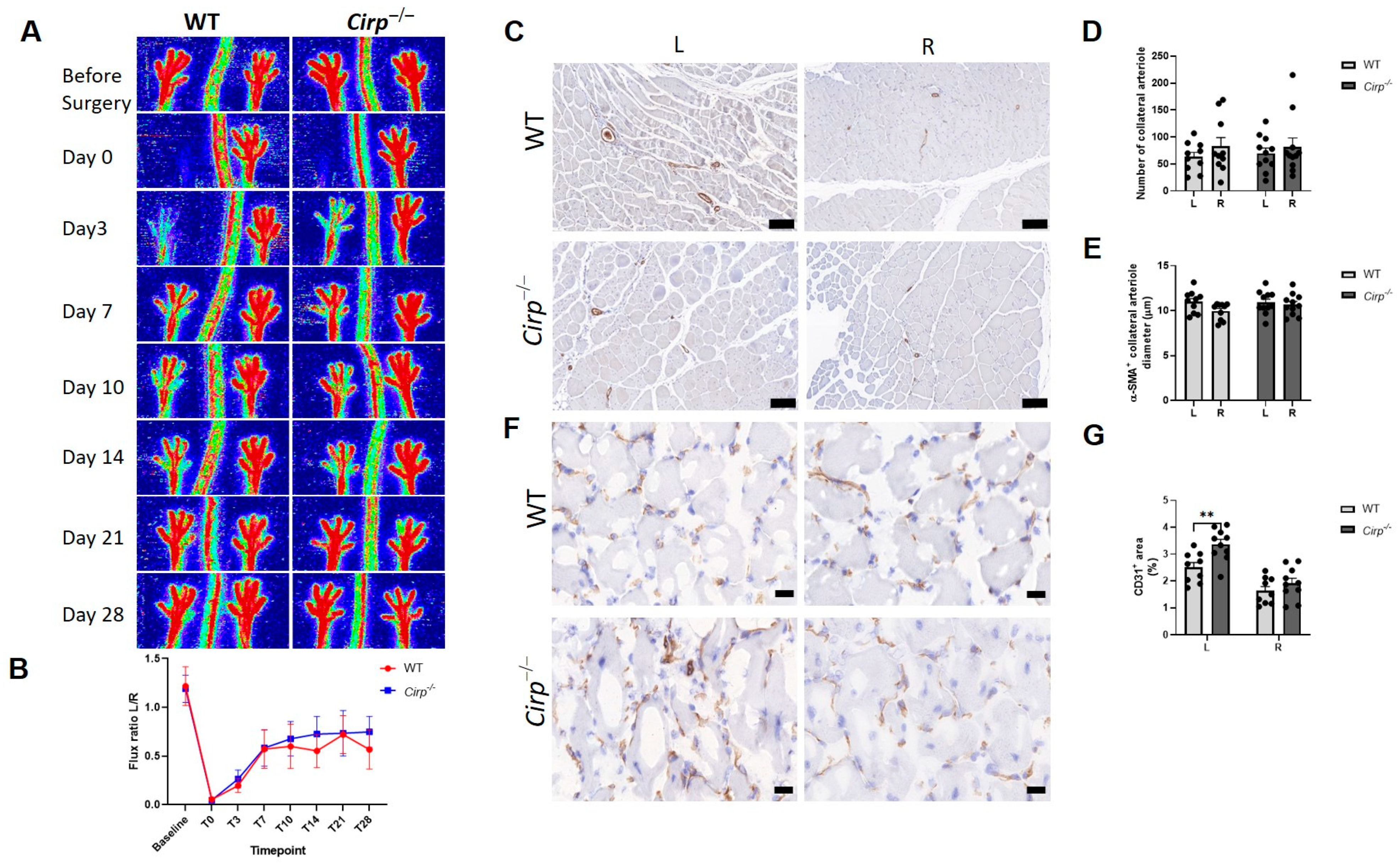
2.1. Blood Flow Recovery and Neovascularization after Hindlimb Ischemia (HLI) Surgery in Cirp−/− Mice
2.2. Ex Vivo Angiogenic Sprouting in Aorta Rings
2.3. MicroRNA Expression in Cirp−/− Mice
2.4. Total CIRBP, CIRBP Splice Variants, and lncRNA-CIRBP-AS1 Expression in Hypothermia
2.5. Angiogenesis Assays and RNA Expression after CIRBP Knockdown
2.6. CIRBP and miRNA Expression in lncRNA-CIRBP-AS1 Knockdown
2.7. Scratch-Wound Healing in CIRBP Knockdown or lncRNA-CIRBP-AS1 Knockdown HUVECs after 4 Hours of siRNA Treatment
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. HLI Model
4.3. Laser Doppler Perfusion Measurements
4.4. Immunohistochemical Staining
4.5. Isolation of Venous Endothelial Cells (HUVECs)
4.6. Primary HUVEC Cell Culture
4.7. Hypothermic HUVEC Cell Culture
4.8. CIRBP and lncRNA-CIRBP-AS1 Knockdown with siRNA Transfection In Vitro
4.9. Migration Assay–Scratch Wound Healing
4.10. Tube-Formation Assay
4.11. Aorta Ring Assay
4.12. RNA Isolation
4.13. MicroRNA Quantification
4.14. mRNA, pri-microRNA, and pre-microRNA Quantification
4.15. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McDermott, M.M. Lower extremity manifestations of peripheral artery disease: The pathophysiologic and functional implications of leg ischemia. Circ. Res. 2015, 116, 1540–1550. [Google Scholar] [CrossRef]
- Vartanian, S.M.; Conte, M.S. Surgical intervention for peripheral arterial disease. Circ. Res. 2015, 116, 1614–1628. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Zhang, Z.; Liang, K.; Shi, S.; Wang, G.; Zhang, K.; Li, K.; Li, W.; Li, T.; Zhai, S. Angioplasty versus bypass surgery in patients with critical limb ischemia-a meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 10595–10602. [Google Scholar] [PubMed]
- Parvar, S.L.; Ngo, L.; Dawson, J.; Nicholls, S.J.; Fitridge, R.; Psaltis, P.J.; Ranasinghe, I. Long-term outcomes following endovascular and surgical revascularization for peripheral artery disease: A propensity score-matched analysis. Eur. Heart J. 2021, ehab116. [Google Scholar] [CrossRef] [PubMed]
- Kubler, M.; Beck, S.; Fischer, S.; Götz, P.; Kumaraswami, K.; Ishikawa-Ankerhold, H.; Lasch, M.; Deindl, E. Absence of Cold-Inducible RNA-Binding Protein (CIRP) Promotes Angiogenesis and Regeneration of Ischemic Tissue by Inducing M2-Like Macrophage Polarization. Biomedicines 2021, 9, 395. [Google Scholar] [CrossRef]
- Kubler, M.; Beck, S.; Peffenköver, L.L.; Götz, P.; Ishikawa-Ankerhold, H.; Preissner, K.T.; Fischer, S.; Lasch, M.; Deindl, E. The Absence of Extracellular Cold-Inducible RNA-Binding Protein (eCIRP) Promotes Pro-Angiogenic Microenvironmental Conditions and Angiogenesis in Muscle Tissue Ischemia. Int. J. Mol. Sci. 2021, 22, 9484. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Zhang, Z.; Li, A.; Xue, J. Effects of hypothermia and cerebral ischemia on cold-inducible RNA-binding protein mRNA expression in rat brain. Brain Res. 2010, 134, 104–110. [Google Scholar] [CrossRef]
- Wellmann, S.; Bührer, C.; Moderegger, E.; Zelmer, A.; Kirschner-Schwabe, R.; Koehne, P.; Fujita, J.; Seeger, K. Oxygen-regulated expression of the RNA-binding proteins RBM3 and CIRP by a HIF-1-independent mechanism. J. Cell Sci. 2004, 117, 1785–1794. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Tong, L.; Tang, L.; Wu, S. The role of cold-inducible RNA binding protein in cell stress response. Int. J. Cancer 2017, 141, 2164–2173. [Google Scholar] [CrossRef] [Green Version]
- Al-Fageeh, M.B.; Smales, C.M. Cold-inducible RNA binding protein (CIRP) expression is modulated by alternative mRNAs. RNA 2009, 15, 1164–1176. [Google Scholar] [CrossRef] [Green Version]
- Lleonart, M.E. A new generation of proto-oncogenes: Cold-inducible RNA binding proteins. Biochim. Biophys. Acta 2010, 1805, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Fujita, J. Cold shock response in mammalian cells. J. Mol. Microbiol. Biotechnol. 1999, 1, 243–255. [Google Scholar] [PubMed]
- Nishiyama, H.; Itoh, K.; Kaneko, Y.; Kishishita, M.; Yoshida, O.; Fujita, J. A glycine-rich RNA-binding protein mediating cold-inducible suppression of mammalian cell growth. J. Cell Biol. 1997, 137, 899–908. [Google Scholar] [CrossRef]
- Nishiyama, H.; Higashitsuji, H.; Yokoi, H.; Itoh, K.; Danno, S.; Matsuda, T.; Fujita, J. Cloning and characterization of human CIRP (cold-inducible RNA-binding protein) cDNA and chromosomal assignment of the gene. Gene 1997, 204, 115–120. [Google Scholar] [CrossRef]
- Zhu, X.; Buhrer, C.; Wellmann, S. Cold-inducible proteins CIRP and RBM3, a unique couple with activities far beyond the cold. Cell. Mol. Life Sci. 2016, 73, 3839–3859. [Google Scholar] [CrossRef] [Green Version]
- Zhong, P.; Huang, H. Recent progress in the research of cold-inducible RNA-binding protein. Future Sci. OA 2017, 3, FSO246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horii, Y.; Shiina, T.; Uehara, S.; Nomura, K.; Shimaoka, H.; Horii, K.; Shimizu, Y. Hypothermia induces changes in the alternative splicing pattern of cold-inducible RNA-binding protein transcripts in a non-hibernator, the mouse. Biomed Res. 2019, 40, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelechano, V.; Steinmetz, L.M. Gene regulation by antisense transcription. Nat. Rev. Genet. 2013, 14, 880–893. [Google Scholar] [CrossRef]
- Gomes, C.P.; Nóbrega-Pereira, S.; Silva, A.B.D.; Rebelo, K.; Alves-Vale, C.; Marinho, S.P.; Carvalho, T.; Dias, S.; De Jesus, B.B. An antisense transcript mediates MALAT1 response in human breast cancer. BMC Cancer 2019, 19, 771. [Google Scholar] [CrossRef] [Green Version]
- Zong, X.; Nakagawa, S.; Freier, S.M.; Fei, J.; Ha, T.; Prasanth, S.G.; Prasanth, K.V. Natural antisense RNA promotes 3’ end processing and maturation of MALAT1 lncRNA. Nucleic Acids Res. 2016, 44, 2898–2908. [Google Scholar] [CrossRef] [Green Version]
- Welten, S.M.; Bastiaansen, A.J.; de Jong, R.C.; de Vries, M.R.; Peters, E.A.; Boonstra, M.C.; Sheikh, S.P.; La Monica, N.; Kandimalla, E.R.; Quax, P.H.; et al. Inhibition of 14q32 MicroRNAs miR-329, miR-487b, miR-494, and miR-495 increases neovascularization and blood flow recovery after ischemia. Circ. Res. 2014, 115, 696–708. [Google Scholar] [CrossRef] [Green Version]
- Welten, S.M.J.; de Jong, R.C.; Wezel, A.; de Vries, M.R.; Boonstra, M.C.; Parma, L.; Jukema, J.W.; van der Sluis, T.C.; Arens, R.; Bot, I.; et al. Inhibition of 14q32 microRNA miR-495 reduces lesion formation, intimal hyperplasia and plasma cholesterol levels in experimental restenosis. Atherosclerosis 2017, 261, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Wezel, A.; Welten, S.M.J.; Razawy, W.; Lagraauw, H.M.; de Vries, M.R.; Goossens, E.A.C.; Boonstra, M.C.; Hamming, J.F.; Kandimalla, E.R.; Kuiper, J.; et al. Inhibition of MicroRNA-494 Reduces Carotid Artery Atherosclerotic Lesion Development and Increases Plaque Stability. Ann. Surg. 2015, 262, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Welten, S.M.; Goossens, E.; Quax, P.; Nossent, A. The multifactorial nature of microRNAs in vascular remodelling. Cardiovasc. Res. 2016, 110, 6–22. [Google Scholar] [CrossRef]
- Downie Ruiz Velasco, A.; Welten, S.M.; Goossens, E.; Quax, P.; Rappsilber, J.; Michlewski, G.; Nossent, A.Y. Posttranscriptional Regulation of 14q32 MicroRNAs by the CIRBP and HADHB during Vascular Regeneration after Ischemia. Mol. Ther. Nucleic Acids 2019, 14, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishna, S.M.; Omer, S.M.; Golledge, J. Evaluation of the clinical relevance and limitations of current pre-clinical models of peripheral artery disease. Clin. Sci. 2016, 130, 127–150. [Google Scholar] [CrossRef]
- Tressel, S.L.; Kim, H.; Ni, C.-W.; Chang, K.; Velasquez-Castano, J.C.; Taylor, W.R.; Yoon, Y.-S.; Jo, H. Angiopoietin-2 stimulates blood flow recovery after femoral artery occlusion by inducing inflammation and arteriogenesis. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1989–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivieri, B.; Vianna, S.; Adenikinju, O.; Beasley, R.E.; Houseworth, J.; Olivieri, B. On the Cutting Edge: Wound Care for the Endovascular Specialist. Semin. Intervent. Radiol. 2018, 35, 406–426. [Google Scholar]
- Idrovo, J.P.; Jacob, A.; Yang, W.L.; Wang, Z.; Yen, H.T.; Nicastro, J.; Coppa, G.F.; Wang, P. A deficiency in cold-inducible RNA-binding protein accelerates the inflammation phase and improves wound healing. Int. J. Mol. Med. 2016, 37, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Qiang, X.; Yang, W.-L.; Wu, R.; Zhou, M.; Jacob, A.; Dong, W.; Kuncewitch, M.; Ji, Y.; Yang, H.; Wang, H.; et al. Cold-inducible RNA-binding protein (CIRP) triggers inflammatory responses in hemorrhagic shock and sepsis. Nat. Med. 2013, 19, 1489–1495. [Google Scholar] [CrossRef] [Green Version]
- de Groot, D.; Hoefer, I.E.; Grundmann, S.; Schoneveld, A.; Haverslag, R.T.; van Keulen, J.K.; Bot, P.T.; Timmers, L.; Piek, J.J.; Pasterkamp, G.; et al. Arteriogenesis requires toll-like receptor 2 and 4 expression in bone-marrow derived cells. J. Mol. Cell. Cardiol. 2011, 50, 25–32. [Google Scholar] [CrossRef]
- Nishiyama, H.; Danno, S.; Kaneko, Y.; Itoh, K.; Yokoi, H.; Fukumoto, M.; Okuno, H.; Millán, J.L.; Matsuda, T.; Yoshida, O.; et al. Decreased expression of cold-inducible RNA-binding protein (CIRP) in male germ cells at elevated temperature. Am. J. Pathol. 1998, 152, 289–296. [Google Scholar] [PubMed]
- Brunt, V.E.; Weidenfeld-Needham, K.M.; Comrada, L.N.; Francisco, M.A.; Eymann, T.M.; Minson, C.T. Serum from young, sedentary adults who underwent passive heat therapy improves endothelial cell angiogenesis via improved nitric oxide bioavailability. Temperature 2019, 6, 169–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neff, D.; Kuhlenhoelter, A.M.; Lin, C.; Wong, B.J.; Motaganahalli, R.L.; Roseguini, B.T. Thermotherapy reduces blood pressure and circulating endothelin-1 concentration and enhances leg blood flow in patients with symptomatic peripheral artery disease. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R392–R400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, T.; Itoh, K.; Higashitsuji, H.; Nakazawa, N.; Sakurai, T.; Liu, Y.; Tokuchi, H.; Fujita, T.; Zhao, Y.; Nishiyama, H.; et al. Cold-inducible RNA-binding protein (Cirp) interacts with Dyrk1b/Mirk and promotes proliferation of immature male germ cells in mice. Proc. Natl. Acad. Sci. USA 2012, 109, 10885–10890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aref, Z.; de Vries, M.R.; Quax, P.H.A. Variations in Surgical Procedures for Inducing Hind Limb Ischemia in Mice and the Impact of These Variations on Neovascularization Assessment. Int. J. Mol. Sci. 2019, 20, 3704. [Google Scholar] [CrossRef] [Green Version]
- Bastiaansen, A.J.; Ewing, M.M.; de Boer, H.C.; van der Pouw Kraan, T.C.; de Vries, M.R.; Peters, E.A.; Welten, S.M.; Arens, R.; Moore, S.M.; Faber, J.E.; et al. Lysine acetyltransferase PCAF is a key regulator of arteriogenesis. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1902–1910. [Google Scholar] [CrossRef] [Green Version]
- Nossent, A.Y.; Bastiaansen, A.J.; Peters, E.A.; de Vries, M.R.; Aref, Z.; Welten, S.M.; de Jager, S.C.A.; van der Pouw Kraan, T.C.T.M.; Quax, P.H.A. CCR7-CCL19/CCL21 Axis is Essential for Effective Arteriogenesis in a Murine Model of Hindlimb Ischemia. J. Am. Heart Assoc. 2017, 6, e005281. [Google Scholar] [CrossRef] [Green Version]
- Parma, L.; Peters, H.A.; Johansson, M.E.; Gutiérrez, S.; Meijerink, H.; De Kimpe, S.; De Vries, M.R.; Quax, P. Bis(maltolato)oxovanadium(IV) Induces Angiogenesis via Phosphorylation of VEGFR2. Int. J. Mol. Sci. 2020, 21, 4643. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Forward Sequence | Reverse Sequence |
|---|---|---|
| HSA-CIRBP | TTGACACCAATGAGCAGTCG | GGCATCCTTAGCGTCGTCAA |
| HSA-splice variant 1 | CGTGGGTTCTCTAGAGGAGGA | CTCGTTGTGTGTAGCGTAACTG |
| HSA-splice variant 2 | CGTGGGTTCTCTAGAGGAGGA | CGCCCTCGGAGTGTGACTTA |
| HSA-splice variant 3 | CGTGGGTTCTCTAGAGGAGGA | TCAACCGTAACTGTCATAACTG |
| HSA-splice variant 4 | GTAGACCAGGCAGGAGGAG | CGCCCTCGGAGTGTGACTTA |
| HSA-lncRNA-CIRBP-AS1 | CAATGGGAAAAGGAGGAAACT | CCTTGTAAAGCTGGTTCTCCA |
| GAPDH | CACCACCATGGAGAAGGC | AGCAGTTGGTGGTGCAGGA |
| HSA-pri-miR-329-1 | TGGGGAAGAATCAGTGGTGT | GACCAGAAGGCCTCCAAGAT |
| HSA-pri-miR-329-2 | TGTCAAGTTTGGGGAAGGAA | GACCAGAAGGCCTCCAAGAT |
| HSA-pre-miR-329 | TGAAGAGAGGTTTTCTGGGTTT | ACCAGGTGTGTTTCGTCCTC |
| HSA-pri-miR-495 | CTGACCCTCAGTGTCCCTTC | ATGGAGGCACTTCAAGGAGA |
| HSA-pre-miR-495 | GCCCATGTTATTTTCGCTTT | CCGAAAAAGAAGTGCACCAT |
| U6 | AGAAGATTAGCATGGCCCCT | ATTTGCGTGTCATCCTTGCG |
| siRNA CIRPB | GAGUCAGAGUGGUGGCUAC | |
| siRNA lncRNA-CIRBP-AS1 | CAGGACCCUCACUCACUA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goossens, E.A.C.; Zhang, L.; de Vries, M.R.; Jukema, J.W.; Quax, P.H.A.; Nossent, A.Y. Cold-Inducible RNA-Binding Protein but Not Its Antisense lncRNA Is a Direct Negative Regulator of Angiogenesis In Vitro and In Vivo via Regulation of the 14q32 angiomiRs—microRNA-329-3p and microRNA-495-3p. Int. J. Mol. Sci. 2021, 22, 12678. https://doi.org/10.3390/ijms222312678
Goossens EAC, Zhang L, de Vries MR, Jukema JW, Quax PHA, Nossent AY. Cold-Inducible RNA-Binding Protein but Not Its Antisense lncRNA Is a Direct Negative Regulator of Angiogenesis In Vitro and In Vivo via Regulation of the 14q32 angiomiRs—microRNA-329-3p and microRNA-495-3p. International Journal of Molecular Sciences. 2021; 22(23):12678. https://doi.org/10.3390/ijms222312678
Chicago/Turabian StyleGoossens, Eveline A. C., Licheng Zhang, Margreet R. de Vries, J. Wouter Jukema, Paul H. A. Quax, and A. Yaël Nossent. 2021. "Cold-Inducible RNA-Binding Protein but Not Its Antisense lncRNA Is a Direct Negative Regulator of Angiogenesis In Vitro and In Vivo via Regulation of the 14q32 angiomiRs—microRNA-329-3p and microRNA-495-3p" International Journal of Molecular Sciences 22, no. 23: 12678. https://doi.org/10.3390/ijms222312678
APA StyleGoossens, E. A. C., Zhang, L., de Vries, M. R., Jukema, J. W., Quax, P. H. A., & Nossent, A. Y. (2021). Cold-Inducible RNA-Binding Protein but Not Its Antisense lncRNA Is a Direct Negative Regulator of Angiogenesis In Vitro and In Vivo via Regulation of the 14q32 angiomiRs—microRNA-329-3p and microRNA-495-3p. International Journal of Molecular Sciences, 22(23), 12678. https://doi.org/10.3390/ijms222312678