
Telmisartan Attenuates Kanamycin-Induced Ototoxicity in Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
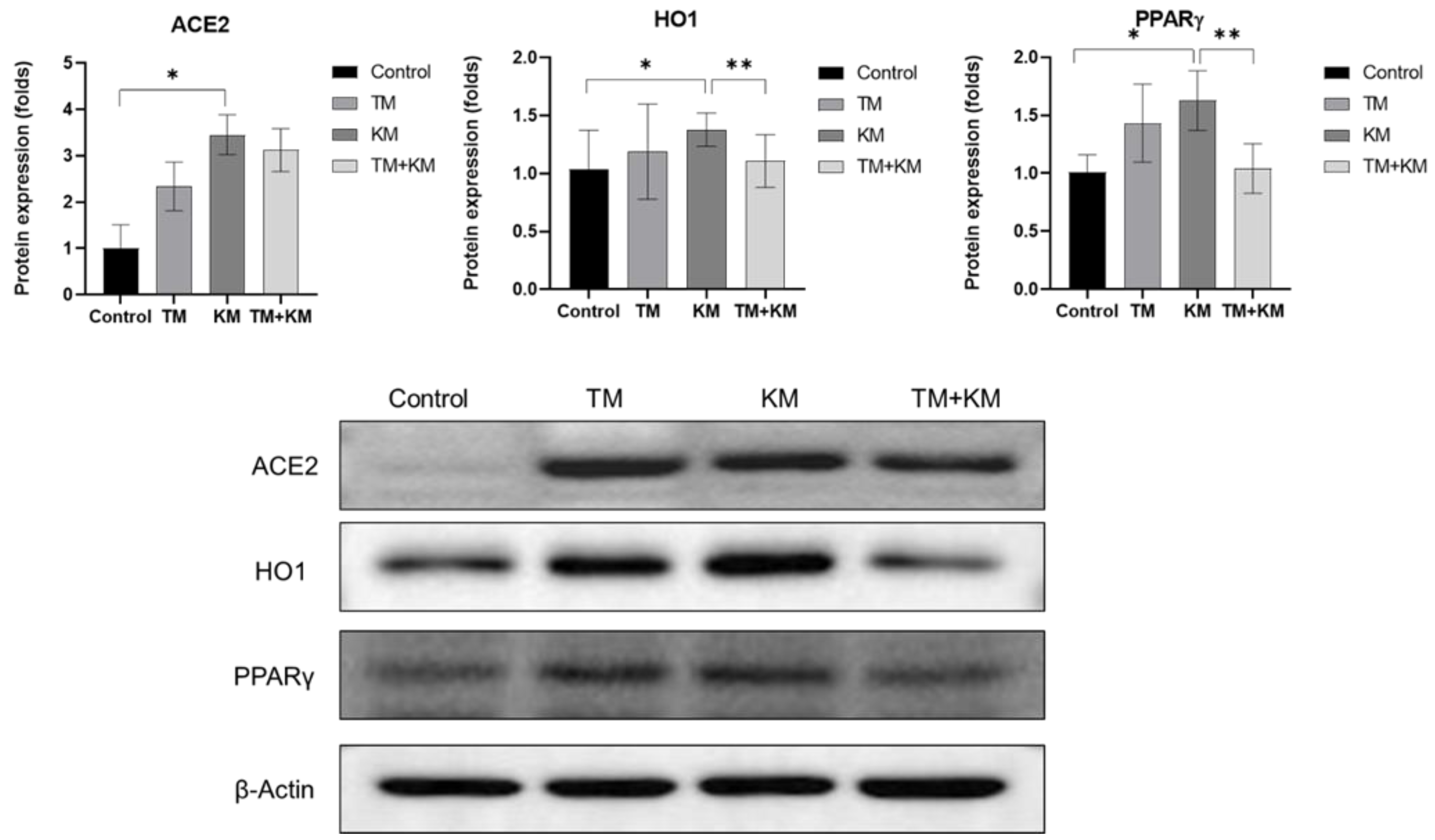
2. Results
3. Discussion
4. Materials and Methods
4.1. Auditory Function Tests
4.2. Cochlear Histologic Examinations
4.3. Protein Expression Levels of ACE2, HO1, and PPARγ
4.4. Statistical Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Michel, M.C.; Foster, C.; Brunner, H.R.; Liu, L. A systematic comparison of the properties of clinically used angiotensin II type 1 receptor antagonists. Pharmacol. Rev. 2013, 65, 809–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, S.; Karnik, S.S.; Saku, K. Review: Angiotensin II type 1 receptor blockers: Class effects versus molecular effects. J. Renin-Angiotensin-Aldosterone Syst. 2011, 12, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, S.; Ohtsu, H.; Suzuki, H.; Shirai, H.; Frank, G.D.; Eguchi, S. Angiotensin II signal transduction through the AT1 receptor: Novel insights into mechanisms and pathophysiology. Clin. Sci. 2007, 112, 417–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benigni, A.; Cassis, P.; Remuzzi, G. Angiotensin II revisited: New roles in inflammation, immunology and aging. EMBO Mol. Med. 2010, 2, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Borem, L.M.A.; Neto, J.F.R.; Brandi, I.V.; Lelis, D.F.; Santos, S.H.S. The role of the angiotensin II type I receptor blocker telmisartan in the treatment of non-alcoholic fatty liver disease: A brief review. Hypertens. Res. 2018, 41, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.A.; Simoes e Silva, A.C.; Maric, C.; Silva, D.M.; Machado, R.P.; de Buhr, I.; Heringer-Walther, S.; Pinheiro, S.V.; Lopes, M.T.; Bader, M.; et al. Angiotensin-(1-7) is an endogenous ligand for the G protein-coupled receptor Mas. Proc. Natl. Acad. Sci. USA 2003, 100, 8258–8263. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.A.; Ferreira, A.J.; Simoes, E.S.A.C. Recent advances in the angiotensin-converting enzyme 2-angiotensin(1-7)-Mas axis. Exp. Physiol. 2008, 93, 519–527. [Google Scholar] [CrossRef]
- Prasad, A.; Quyyumi, A.A. Renin-angiotensin system and angiotensin receptor blockers in the metabolic syndrome. Circulation 2004, 110, 1507–1512. [Google Scholar] [CrossRef]
- Duarte, M.; Pelorosso, F.; Nicolosi, L.N.; Salgado, M.V.; Vetulli, H.; Aquieri, A.; Azzato, F.; Castro, M.; Coyle, J.; Davolos, I.; et al. Telmisartan for treatment of Covid-19 patients: An open multicenter randomized clinical trial. EClinicalMedicine 2021, 37, 100962. [Google Scholar] [CrossRef]
- Benson, S.C.; Pershadsingh, H.A.; Ho, C.I.; Chittiboyina, A.; Desai, P.; Pravenec, M.; Qi, N.; Wang, J.; Avery, M.A.; Kurtz, T.W. Identification of telmisartan as a unique angiotensin II receptor antagonist with selective PPARgamma-modulating activity. Hypertension 2004, 43, 993–1002. [Google Scholar] [CrossRef] [Green Version]
- Bright, J.J.; Kanakasabai, S.; Chearwae, W.; Chakraborty, S. PPAR Regulation of Inflammatory Signaling in CNS Diseases. PPAR Res. 2008, 2008, 658520. [Google Scholar] [CrossRef] [Green Version]
- Oyekan, A. PPARs and their effects on the cardiovascular system. Clin. Exp. Hypertens 2011, 33, 287–293. [Google Scholar] [CrossRef]
- Poka-Mayap, V.; Balkissou Adamou, D.; Pefura-Yone, E.W.; Kuaban, C. Kanamycin-induced ototoxicity during treatment of multidrug-resistant tuberculosis. Rev. Mal. Respir. 2020, 37, 369–375. [Google Scholar] [CrossRef]
- Tabuchi, K.; Nishimura, B.; Nakamagoe, M.; Hayashi, K.; Nakayama, M.; Hara, A. Ototoxicity: Mechanisms of cochlear impairment and its prevention. Curr. Med. Chem. 2011, 18, 4866–4871. [Google Scholar] [CrossRef]
- Esterberg, R.; Linbo, T.; Pickett, S.B.; Wu, P.; Ou, H.C.; Rubel, E.W.; Raible, D.W. Mitochondrial calcium uptake underlies ROS generation during aminoglycoside-induced hair cell death. J. Clin. Investig. 2016, 126, 3556–3566. [Google Scholar] [CrossRef] [Green Version]
- Chun, K.J.; Lee, C.H.; Kim, K.W.; Lee, S.M.; Kim, S.Y. Effects of Androgen Receptor Inhibition on Kanamycin-Induced Hearing Loss in Rats. Int. J. Mol. Sci. 2021, 22, 5307. [Google Scholar] [CrossRef]
- Cortada, M.; Wei, E.; Jain, N.; Levano, S.; Bodmer, D. Telmisartan Protects Auditory Hair Cells from Gentamicin-Induced Toxicity in vitro. Audiol. Neurootol. 2020, 25, 297–308. [Google Scholar] [CrossRef]
- Kros, C.J.; Steyger, P.S. Aminoglycoside- and Cisplatin-Induced Ototoxicity: Mechanisms and Otoprotective Strategies. Cold Spring Harb. Perspect Med. 2019, 9. [Google Scholar] [CrossRef]
- Liu, K.; Jiang, X.; Shi, C.; Shi, L.; Yang, B.; Shi, L.; Xu, Y.; Yang, W.; Yang, S. Cochlear inner hair cell ribbon synapse is the primary target of ototoxic aminoglycoside stimuli. Mol. Neurobiol. 2013, 48, 647–654. [Google Scholar] [CrossRef]
- Koo, J.W.; Quintanilla-Dieck, L.; Jiang, M.; Liu, J.; Urdang, Z.D.; Allensworth, J.J.; Cross, C.P.; Li, H.; Steyger, P.S. Endotoxemia-mediated inflammation potentiates aminoglycoside-induced ototoxicity. Sci. Transl. Med. 2015, 7, 298ra118. [Google Scholar] [CrossRef] [Green Version]
- Prasad, K.N.; Bondy, S.C. Increased oxidative stress, inflammation, and glutamate: Potential preventive and therapeutic targets for hearing disorders. Mech. Ageing Dev. 2020, 185, 111191. [Google Scholar] [CrossRef]
- Han, H.; Dong, Y.; Ma, X. Dihydromyricetin Protects Against Gentamicin-Induced Ototoxicity via PGC-1alpha/SIRT3 Signaling in vitro. Front. Cell Dev. Biol. 2020, 8, 702. [Google Scholar] [CrossRef]
- Hoshino, T.; Tabuchi, K.; Nishimura, B.; Tanaka, S.; Nakayama, M.; Ishii, T.; Warabi, E.; Yanagawa, T.; Shimizu, R.; Yamamoto, M.; et al. Protective role of Nrf2 in age-related hearing loss and gentamicin ototoxicity. Biochem. Biophys. Res. Commun. 2011, 415, 94–98. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.N.M.; Jeong, S.Y.; Kim, D.H.; Park, Y.H.; Lee, J.S.; Lee, K.W.; Moon, I.S.; Choung, S.Y.; Kim, S.H.; Kang, T.H.; et al. Protective Mechanisms of Avocado Oil Extract Against Ototoxicity. Nutrients 2020, 12, 947. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.T.; Lee, J.H.; Carpena, N.T.; Lee, M.Y.; Chung, P.S.; Jung, J.Y. Estrogen Replacement Reduces Hearing Threshold Shifts and Cochlear Hair Cell Loss After Acoustic Overexposure in Ovariectomized Rats. Clin. Exp. Otorhinolaryngol. 2021, 14, 61–68. [Google Scholar] [CrossRef]
- Lee, S.Y.; Han, S.Y.; Shim, Y.J.; Han, J.J.; Cho, D.; Kim, J.E.; Kim, Y.H. Effect of Ginkgo Biloba Extract on N-Methyl-D-Aspartic Acid Receptor Subunit 2B Expression in a Salicylate-Induced Ototoxicity Model. Clin. Exp. Otorhinolaryngol. 2019, 12, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Fusco, R.; Siracusa, R.; D’Amico, R.; Peritore, A.F.; Cordaro, M.; Gugliandolo, E.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. Melatonin Plus Folic Acid Treatment Ameliorates Reserpine-Induced Fibromyalgia: An Evaluation of Pain, Oxidative Stress, and Inflammation. Antioxidants 2019, 8, 628. [Google Scholar] [CrossRef] [Green Version]
- Sekulic-Jablanovic, M.; Petkovic, V.; Wright, M.B.; Kucharava, K.; Huerzeler, N.; Levano, S.; Brand, Y.; Leitmeyer, K.; Glutz, A.; Bausch, A.; et al. Effects of peroxisome proliferator activated receptors (PPAR)-gamma and -alpha agonists on cochlear protection from oxidative stress. PLoS ONE 2017, 12, e0188596. [Google Scholar] [CrossRef]
- Park, C.; Ji, H.M.; Kim, S.J.; Kil, S.H.; Lee, J.N.; Kwak, S.; Choe, S.K.; Park, R. Fenofibrate exerts protective effects against gentamicin-induced toxicity in cochlear hair cells by activating antioxidant enzymes. Int. J. Mol. Med. 2017, 39, 960–968. [Google Scholar] [CrossRef] [Green Version]
- Paciello, F.; Fetoni, A.R.; Rolesi, R.; Wright, M.B.; Grassi, C.; Troiani, D.; Paludetti, G. Pioglitazone Represents an Effective Therapeutic Target in Preventing Oxidative/Inflammatory Cochlear Damage Induced by Noise Exposure. Front. Pharmacol. 2018, 9, 1103. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, H.B.; Napimoga, M.H.; Lopes, A.H.; de Macedo Maganin, A.G.; Cunha, T.M.; Van Dyke, T.E.; Clemente Napimoga, J.T. Activation of PPAR-gamma induces macrophage polarization and reduces neutrophil migration mediated by heme oxygenase 1. Int. Immunopharmacol. 2020, 84, 106565. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Park, S.S.; Lee, D.H.; Lee, S.M.; Kim, M.Y.; Choi, B.Y.; Kim, S.Y. Tauroursodeoxycholic acid attenuates cisplatin-induced hearing loss in rats. Neurosci. Lett. 2020, 722, 134838. [Google Scholar] [CrossRef] [PubMed]
- Abdelkader, N.F.; Abd El-Latif, A.M.; Khattab, M.M. Telmisartan/17beta-estradiol mitigated cognitive deficit in an ovariectomized rat model of Alzheimer’s disease: Modulation of ACE1/ACE2 and AT1/AT2 ratio. Life Sci. 2020, 245, 117388. [Google Scholar] [CrossRef] [PubMed]
- Silva de Almeida, T.C.; Lanza, K.; da Silva Filha, R.; LMC, C.C.; Fonseca, E.G.; Chagas, M.W.; Rocha, N.P.; de Sa, M.A.; Vieira, M.A.R.; Caliari, M.V.; et al. ACE2 activator diminazene aceturate exerts renoprotective effects in gentamicin-induced acute renal injury in rats. Clin. Sci. 2020, 134, 3093–3106. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fattah, M.M.; Elgendy, A.; Mohamed, W.R. Xanthenone, ACE2 activator, counteracted gentamicin-induced nephrotoxicity in rats: Impact on oxidative stress and ACE2/Ang-(1-7) signaling. Life Sci. 2021, 275, 119387. [Google Scholar] [CrossRef]
- Lee, C.H.; Kim, K.W.; Lee, S.M.; Kim, S.Y. Dose-Dependent Effects of Resveratrol on Cisplatin-Induced Hearing Loss. Int. J. Mol. Sci. 2020, 22, 113. [Google Scholar] [CrossRef]
- Montgomery, S.C.; Cox, B.C. Whole Mount Dissection and Immunofluorescence of the Adult Mouse Cochlea. J. Vis. Exp. 2016, 107, e53561. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.H.; Lee, S.M.; Kim, S.Y. Telmisartan Attenuates Kanamycin-Induced Ototoxicity in Rats. Int. J. Mol. Sci. 2021, 22, 12716. https://doi.org/10.3390/ijms222312716
Lee CH, Lee SM, Kim SY. Telmisartan Attenuates Kanamycin-Induced Ototoxicity in Rats. International Journal of Molecular Sciences. 2021; 22(23):12716. https://doi.org/10.3390/ijms222312716
Chicago/Turabian StyleLee, Chang Ho, So Min Lee, and So Young Kim. 2021. "Telmisartan Attenuates Kanamycin-Induced Ototoxicity in Rats" International Journal of Molecular Sciences 22, no. 23: 12716. https://doi.org/10.3390/ijms222312716
APA StyleLee, C. H., Lee, S. M., & Kim, S. Y. (2021). Telmisartan Attenuates Kanamycin-Induced Ototoxicity in Rats. International Journal of Molecular Sciences, 22(23), 12716. https://doi.org/10.3390/ijms222312716