
Ultrastructural Study of Platelet Behavior and Interrelationship in Sprouting and Intussusceptive Angiogenesis during Arterial Intimal Thickening Formation

 , and
, and 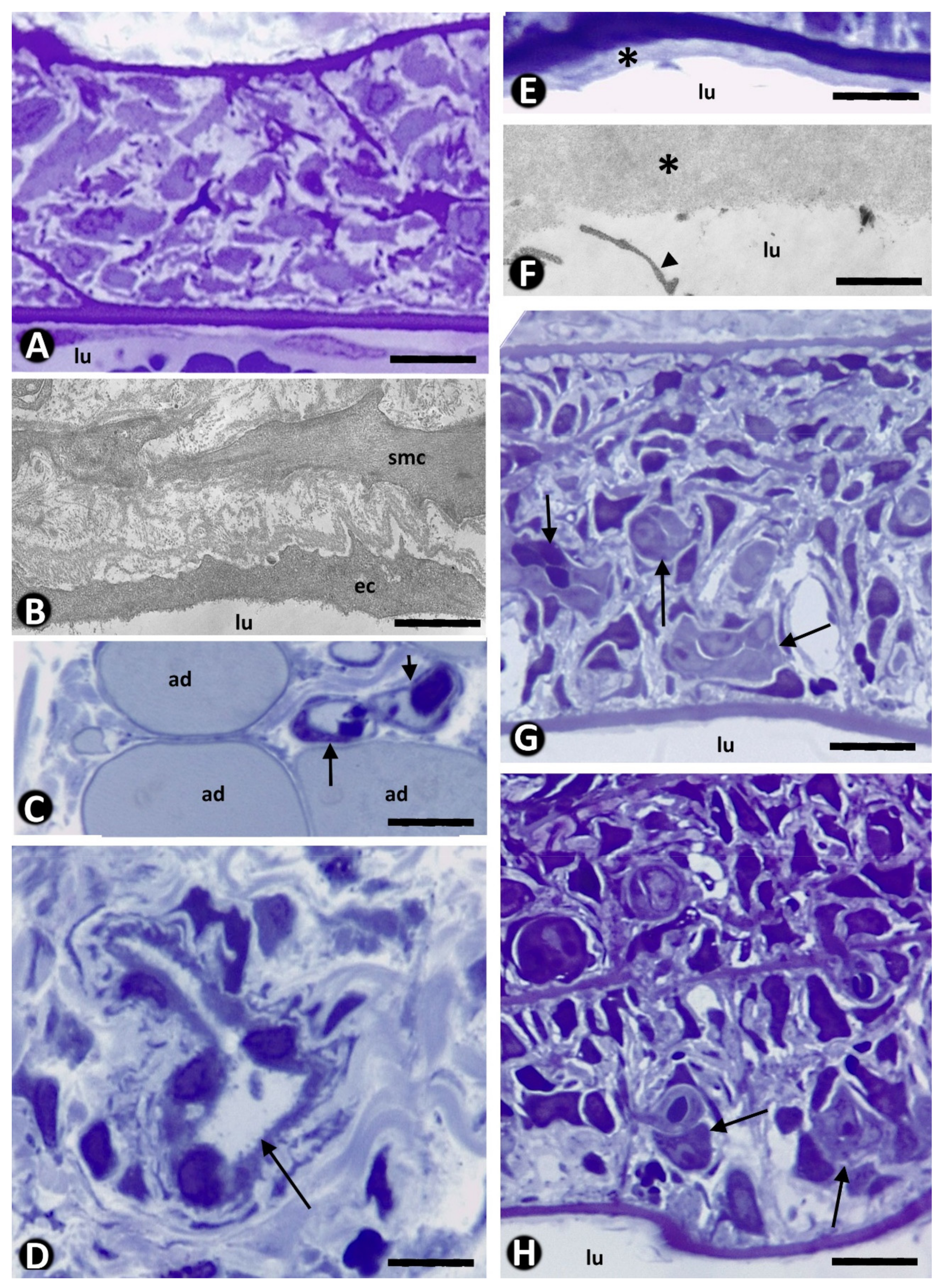{kind=link}
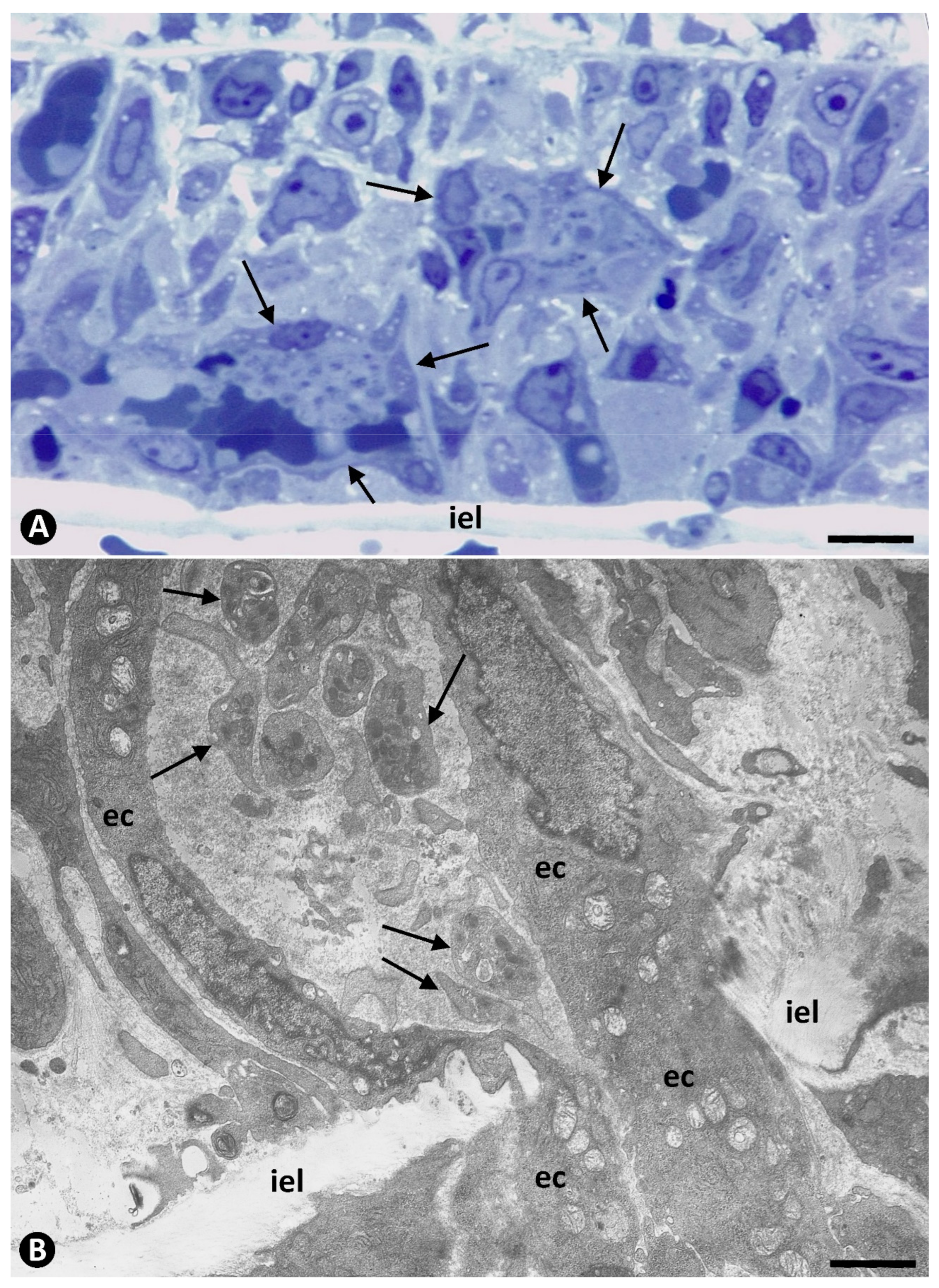{kind=link}
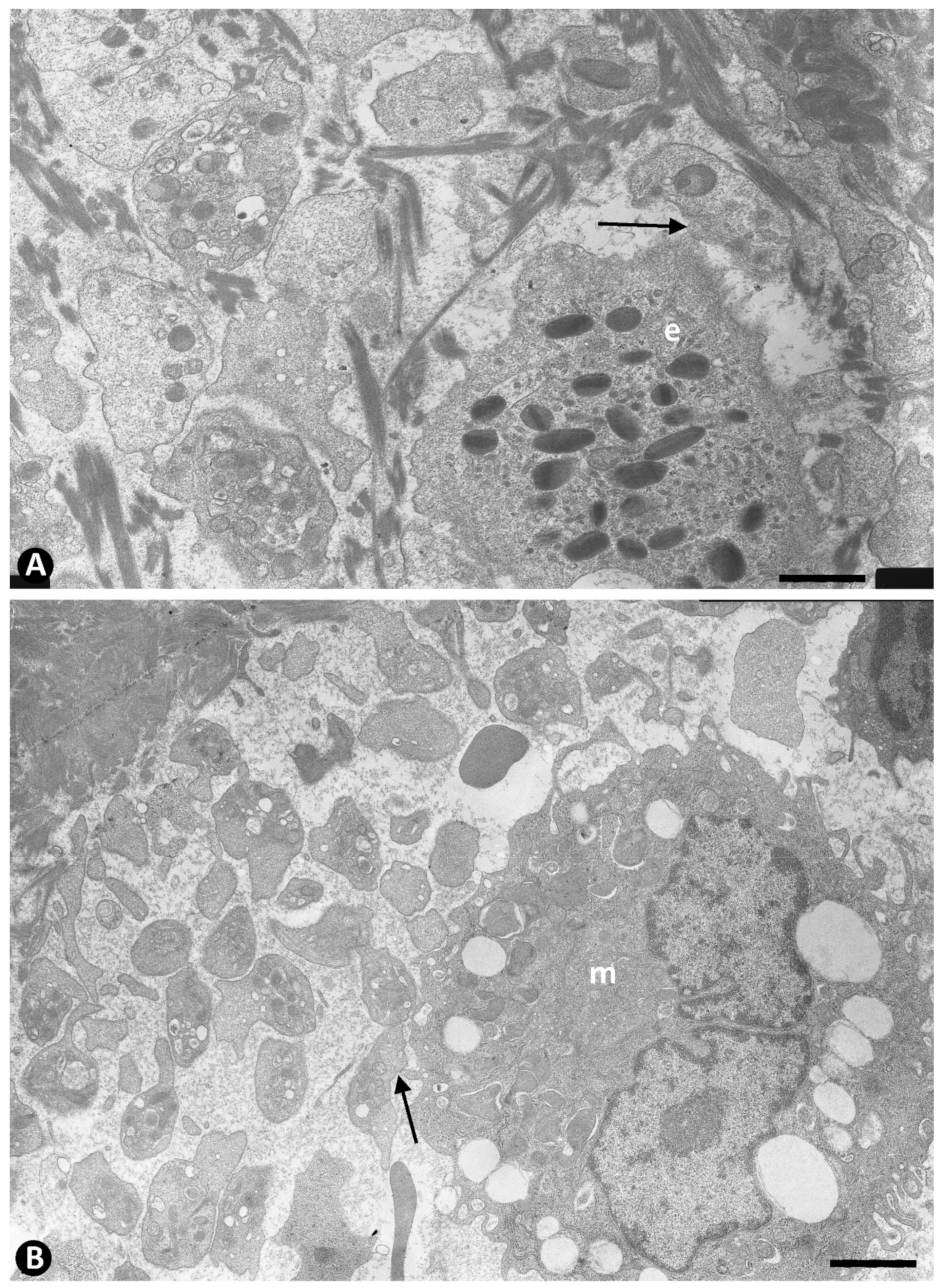{kind=link}
{kind=link}
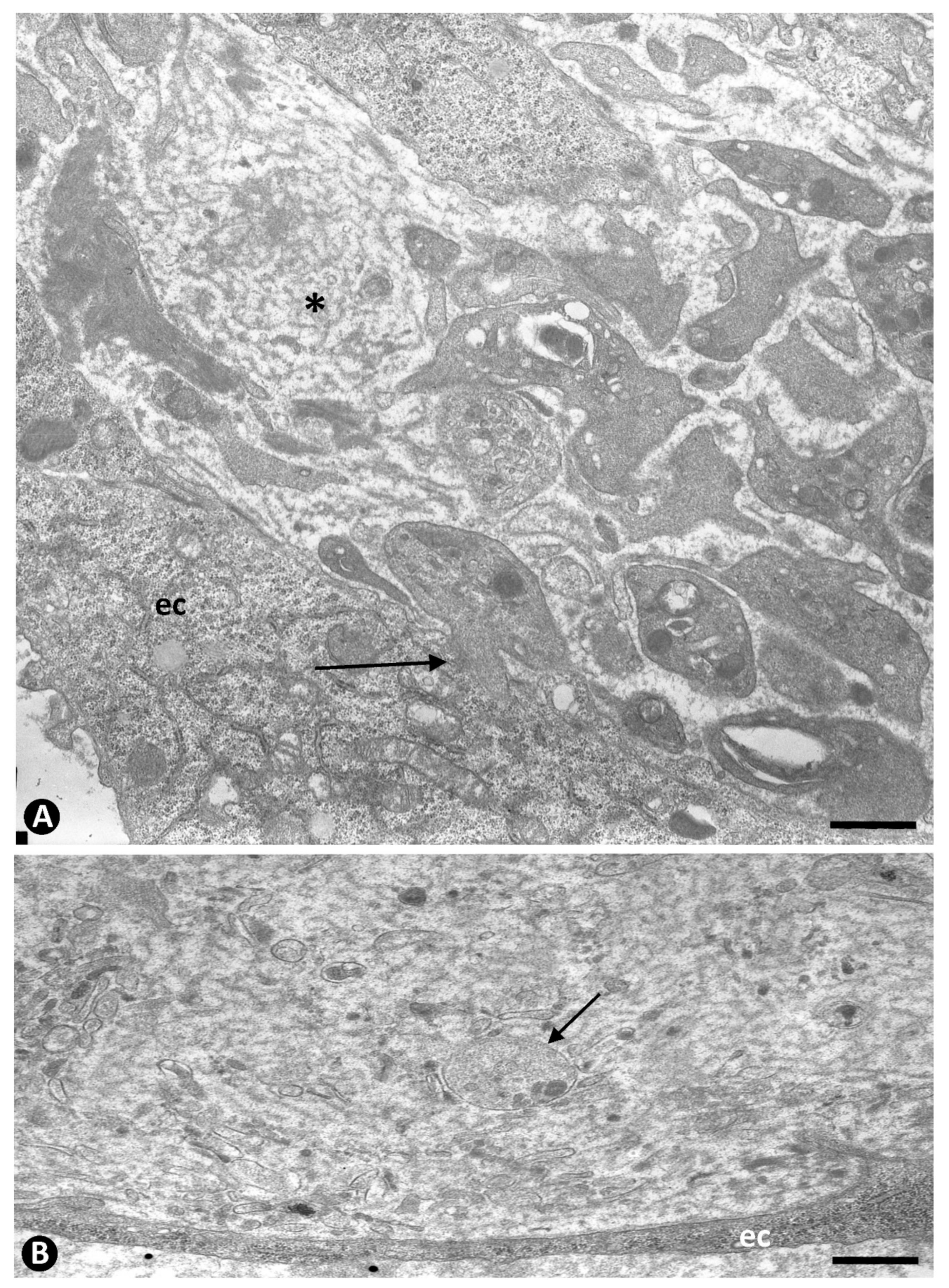{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Controls
2.2. Platelets in Angiogenesis and Neovessel Regression during Arterial IT Formation
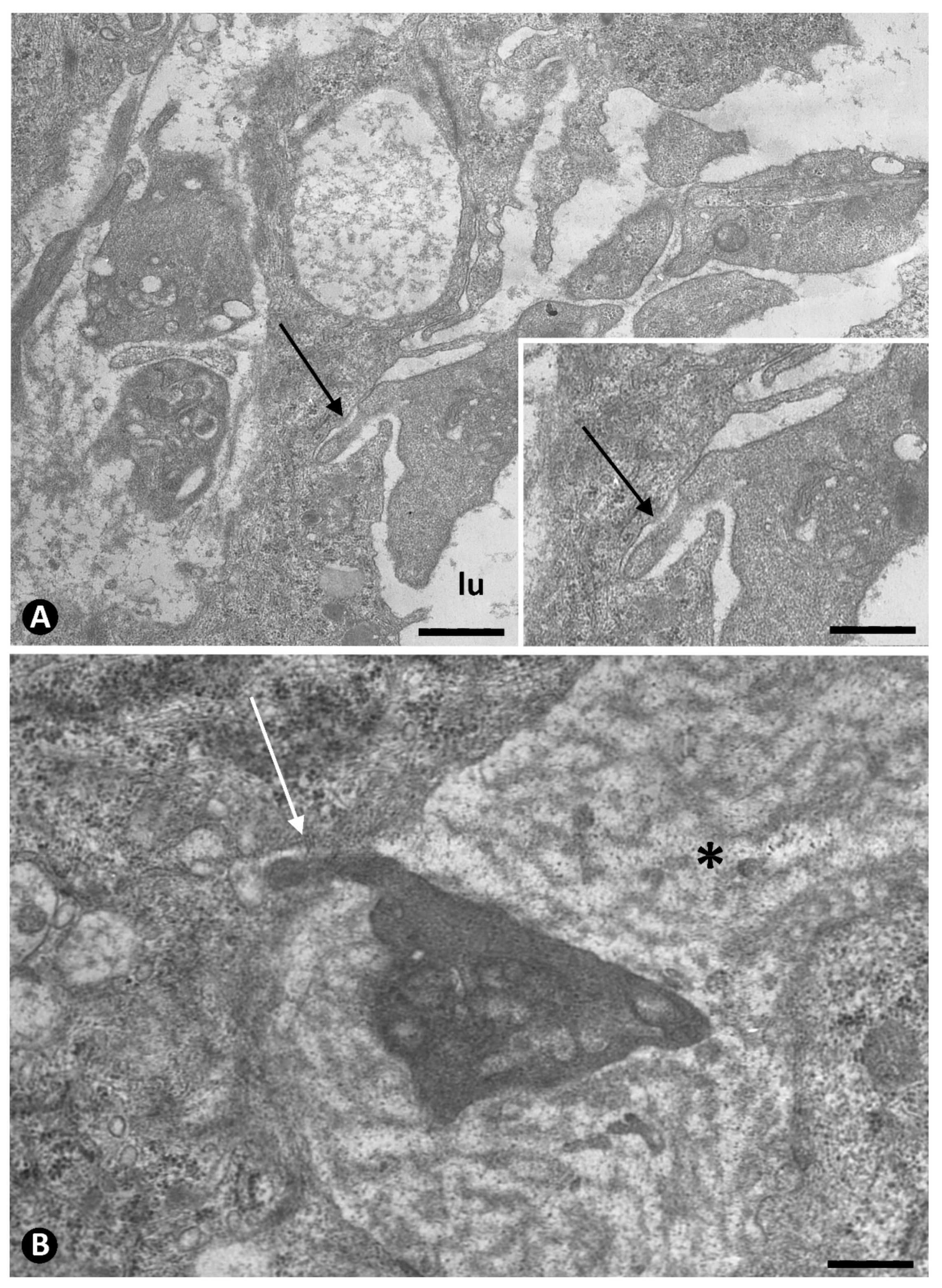
2.2.1. Initial Phase. Platelets during Sprouting and Intussusceptive Angiogenesis
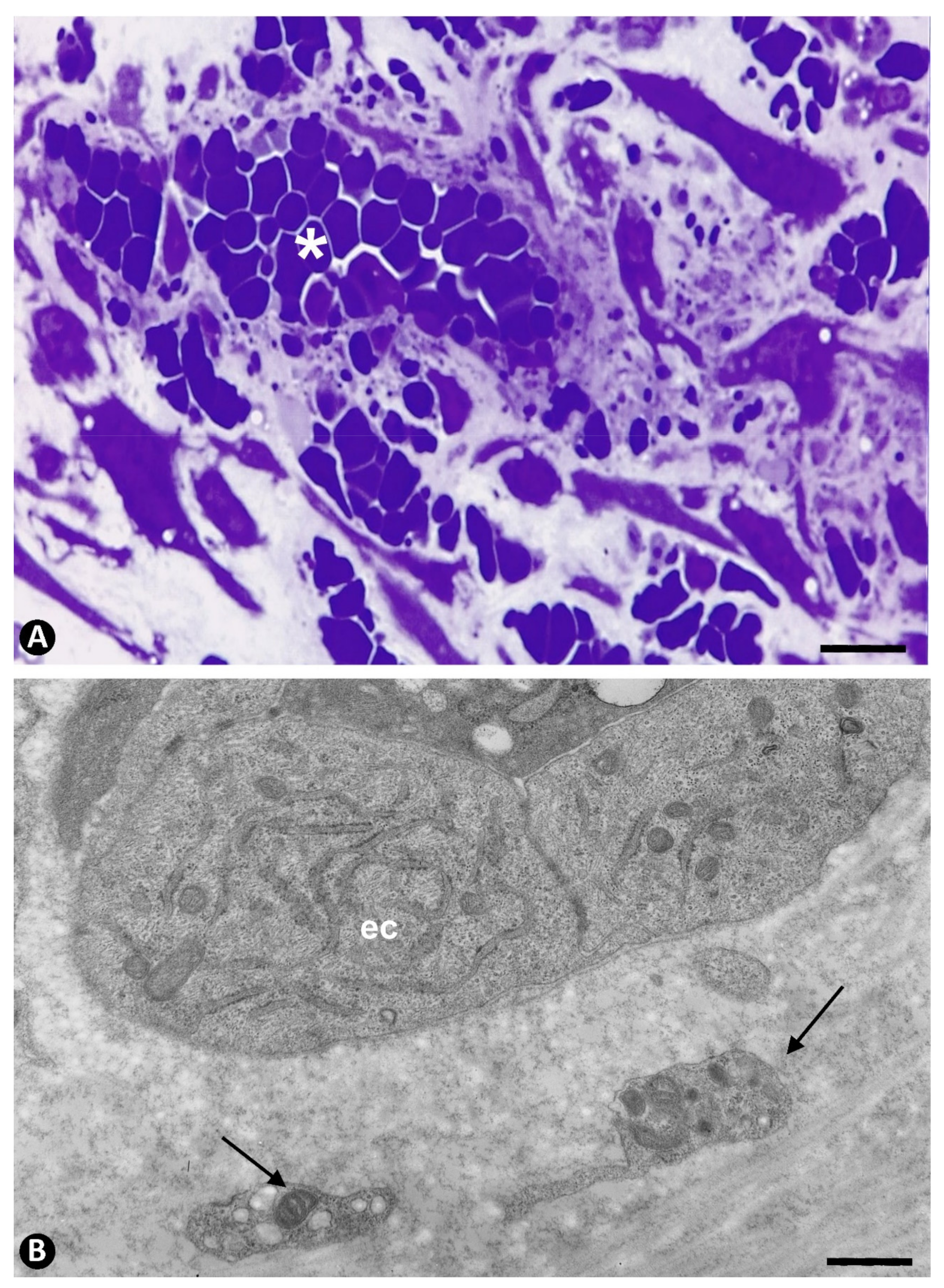
2.2.2. Secondary Phase. Platelets during Regression of the Newly Formed Vessels
3. Discussion
4. Material and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Badimon, J.J.; Ortiz, A.F.; Meyer, B.; Mailhac, A.; Fallon, J.T.; Falk, E.; Badimon, L.; Chesebro, J.H.; Fuster, V. Different response to balloon angioplasty of carotid and coronary arteries: Effects on acute platelet deposition and intimal thickening. Atherosclerosis 1998, 140, 307–314. [Google Scholar] [CrossRef]
- Chandrasekar, B.; Tanguay, J.F. Platelets and restenosis. J. Am. Coll. Cardiol. 2000, 35, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Weyrich, A.S.; Prescott, S.M.; Zimmerman, G.A. Platelets, endothelial cells, inflammatory chemokines, and restenosis: Complex signaling in the vascular play book. Circulation 2002, 106, 1433–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, C.; Massberg, S. Platelets in atherosclerosis and thrombosis. Handb. Exp. Pharmacol. 2012, 210, 111–133. [Google Scholar]
- Lehle, K.; Li, J.; Zimmermann, H.; Hartmann, B.; Wehner, D.; Schmid, T.; Schmid, C. In vitro endothelialization and platelet adhesion on titaniferous upgraded polyether and polycarbonate polyurethanes. Materials 2014, 7, 623–636. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.; Gao, J.; Li, S.; Li, R.; Chen, Z.; Liang, M.; Liu, X.; Xu, G. Mean platelet volume as a predictor for restenosis after carotid angioplasty and stenting. Stroke 2018, 49, 872–876. [Google Scholar] [CrossRef] [PubMed]
- Lebas, H.; Yahiaoui, K.; Martos, R.; Boulaftali, Y. Platelets are at the nexus of vascular diseases. Front. Cardiovasc. Med. 2019, 6, 132. [Google Scholar] [CrossRef]
- Alexandru, N.; Safciuc, F.; Constantin, A.; Nemecz, M.; Tanko, G.; Filippi, A.; Dragan, E.; Bãdilã, E.; Georgescu, A. Platelets of healthy origins promote functional improvement of atherosclerotic endothelial progenitor cells. Front. Pharmacol. 2019, 10, 424. [Google Scholar] [CrossRef] [PubMed]
- Berezin, A.E.; Berezin, A. Platelet-derived vesicles: Diagnostic and predictive value in cardiovascular diseases. J. Unexplored Med. Data 2019, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Tang, C. Targeting platelet in atherosclerosis plaque formation: Current knowledge and future perspectives. Int. J. Mol. Sci. 2020, 21, 9760. [Google Scholar] [CrossRef] [PubMed]
- Haybar, H.; Pezeshki, S.M.S.; Saki, N. Platelets in in-stent restenosis: From fundamental role to possible prognostic application. Curr. Cardiol. Rev. 2020, 16, 285–291. [Google Scholar] [CrossRef]
- Barale, C.; Melchionda, E.; Morotti, A.; Russo, I. PCSK9 biology and its role in atherothrombosis. Int. J. Mol. Sci. 2021, 22, 5880. [Google Scholar] [CrossRef]
- Liu, J.T.; Bao, H.; Fan, Y.J.; Li, Z.T.; Yao, Q.P.; Han, Y.; Zhang, M.L.; Jiang, Z.L.; Qi, Y.X. Platelet-derived microvesicles promote VSMC dedifferentiation after intimal injury via Src/Lamtor1/mTORC1 Signaling. Front. Cell Dev. Biol. 2021, 9, 744320. [Google Scholar] [CrossRef] [PubMed]
- Kaya, U.; Koza, Y. Predictive value of mean platelet volume in saphenous vein graft disease. Braz. J. Cardiovasc. Surg. 2018, 33, 317–322. [Google Scholar] [CrossRef]
- Kisucka, J.; Butterfield, C.E.; Duda, D.G.; Eichenberger, S.C.; Saffaripour, S.; Ware, J.; Ruggeri, Z.M.; Jain, R.K.; Folkman, J.; Wagner, D.D. Platelets and platelet adhesion support angiogenesis while preventing excessive hemorrhage. Proc. Natl. Acad. Sci. USA 2006, 103, 855–860. [Google Scholar] [PubMed] [Green Version]
- Patzelt, J.; Langer, H.F. Platelets in angiogenesis. Curr. Vasc. Pharmacol. 2012, 10, 570–577. [Google Scholar] [CrossRef]
- Etulain, J.; Negrotto, S.; Schattner, M. Role of platelets in angiogenesis in health and disease. Curr. Angiogenesis 2014, 3, 1. [Google Scholar] [CrossRef]
- Martínez, C.E.; Smith, P.C.; Palma Alvarado, V.A. The influence of platelet-derived products on angiogenesis and tissue repair: A concise update. Front. Physiol. 2015, 6, 290. [Google Scholar] [CrossRef] [Green Version]
- Cramer, E.M.; Norol, F.; Guichard, J.; Breton-Gorius, J.; Vainchenker, W.; Massé, J.M.; Debili, N. Ultrastructure of platelet formation by human megakaryocytes cultured with the Mpl ligand. Blood 1997, 89, 2336–2346. [Google Scholar] [PubMed]
- Thon, J.N.; Italiano, J.E. Platelets: Production, morphology and ultrastructure. Handb. Exp. Pharmacol. 2012, 210, 3–22. [Google Scholar]
- Du Plooy, J.N.; Buys, A.; Duim, W.; Pretorius, E. Comparison of platelet ultrastructure and elastic properties in thrombo-embolic ischemic stroke and smoking using atomic force and scanning electron microscopy. PLoS ONE 2013, 8, e69774. [Google Scholar]
- Heijnen, H.F.G.; Korporaal, S.J.A. Platelet morphology and ultrastructure. In Thrombotic and Non-Thrombotic Disorders. Pathophysiology, Pharmacology and Therapeutics: An Update, 1st ed.; Gresele, P., Kleiman, N.S., López, J.A., Page, C.P., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; Chapter 3; pp. 21–37. [Google Scholar]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; Gayoso, S.; Carrasco, J.L.; Díaz-Flores, L., Jr.; González-Gómez, M.; Madrid, J.F. Intussusceptive angiogenesis and peg-socket junctions between endothelial cells and smooth muscle cells in early arterial intimal thickening. Int. J. Mol. Sci. 2020, 21, 8049. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gallant, R.C.; Ni, H. Extracellular matrix proteins in the regulation of thrombus formation. Curr. Opin. Hematol. 2016, 23, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.R.; Carrim, N.; Neves, M.A.; McKeown, T.; Stratton, T.W.; Coelho, R.M.; Lei, X.; Chen, P.; Xu, J.; Dai, X.; et al. Platelets and platelet adhesion molecules: Novel mechanisms of thrombosis and anti-thrombotic therapies. Thromb. J. 2016, 14, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Periayah, M.H.; Halim, A.S.; Mat Saad, A.Z. Mechanism action of platelets and crucial blood coagulation pathways in hemostasis. Int. J. Hematol. Oncol. Stem Cell Res. 2017, 11, 319–327. [Google Scholar] [PubMed]
- Barale, C.; Russo, I. Influence of cardiometabolic risk factors on platelet function. Int. J. Mol. Sci. 2020, 21, 623. [Google Scholar] [CrossRef] [Green Version]
- Scavone, M.; Bozzi, S.; Mencarini, T.; Podda, G.; Cattaneo, M.; Redaelli, A. Platelet adhesion and thrombus formation in microchannels: The effect of assay-dependent variables. Int. J. Mol. Sci. 2020, 21, 750. [Google Scholar] [CrossRef] [Green Version]
- Humar, R.; Kiefer, F.N.; Berns, H.; Resink, T.J.; Battegay, E.J. Hypoxia enhances vascular cell proliferation and angiogenesis in vitro via rapamycin (mTOR)-dependent signaling. FASEB J. 2002, 16, 771–780. [Google Scholar] [CrossRef] [Green Version]
- Narula, N.; Dannenberg, A.J.; Olin, J.W.; Bhatt, D.L.; Johnson, K.W.; Nadkarni, G.; Min, J.; Torii, S.; Poojary, P.; Anand, S.S.; et al. Pathology of peripheral artery disease in patients with critical limb ischemia. J. Am. Coll. Cardiol. 2018, 72, 2152–2163. [Google Scholar] [CrossRef]
- Vasina, E.M.; Cauwenberghs, S.; Feijge, M.A.; Heemskerk, J.W.; Weber, C.; Koenen, R.R. Microparticles from apoptotic platelets promote resident macrophage differentiation. Cell Death Dis. 2011, 2, e211. [Google Scholar] [CrossRef] [Green Version]
- Gros, A.; Ollivier, V.; Ho-Tin-Noé, B. Platelets in inflammation: Regulation of leukocyte activities and vascular repair. Front. Immunol 2015, 5, 678. [Google Scholar] [CrossRef]
- Lam, F.W.; Vijayan, K.V.; Rumbaut, R.E. Platelets and their interactions with other immune cells. Compr. Physiol. 2015, 5, 1265–1280. [Google Scholar] [PubMed] [Green Version]
- Linke, B.; Schreiber, Y.; Picard-Willems, B.; Slattery, P.; Nüsing, R.M.; Harder, S.; Geisslinger, G.; Scholich, K. Activated Platelets induce an anti-inflammatory response of monocytes/macrophages through cross-regulation of PGE2 and cytokines. Mediat. Inflamm. 2017, 2017, 1463216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, L.S.; Migliari Branco, L.; Franklin, B.S. Regulation of innate immune responses by platelets. Front. Immunol. 2019, 10, 1320. [Google Scholar] [CrossRef] [PubMed]
- Rolfes, V.; Ribeiro, L.S.; Hawwari, I.; Böttcher, L.; Rosero, N.; Maasewerd, S.; Santos, M.L.S.; Próchnicki, T.; Silva, C.M.S.; Wanderley, C.W.S.; et al. Platelets fuel the inflammasome activation of innate immune cells. Cell Rep. 2020, 31, 107615. [Google Scholar] [CrossRef] [PubMed]
- Etulain, J.; Schattner, M. Glycobiology of platelet-endothelial cell interactions. Glycobiology 2014, 24, 1252–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nhek, S.; Clancy, R.; Lee, K.A.; Allen, N.M.; Barrett, T.J.; Marcantoni, E.; Nwaukoni, J.; Rasmussen, S.; Rubin, M.; Newman, J.D.; et al. Activated platelets induce endothelial cell activation via an interleukin-1β pathway in systemic lupus erythematosus. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 707–716. [Google Scholar] [PubMed] [Green Version]
- Hamilos, M.; Petousis, S.; Parthenakis, F. Interaction between platelets and endothelium: From pathophysiology to new therapeutic options. Cardiovasc. Diagn. Ther. 2018, 8, 568–580. [Google Scholar] [PubMed]
- Heiss, M.; Hellström, M.; Kalén, M.; May, T.; Weber, H.; Hecker, M.; Augustin, H.G.; Korff, T. Endothelial cell spheroids as a versatile tool to study angiogenesis in vitro. FASEB J. 2015, 29, 3076–3084. [Google Scholar] [CrossRef]
- De Moor, L.; Merovci, I.; Baetens, S.; Verstraeten, J.; Kowalska, P.; Krysko, D.V.; De Vos, W.H.; Declercq, H. High-throughput fabrication of vascularized spheroids for bioprinting. Biofabrication 2018, 10, 035009. [Google Scholar] [CrossRef] [PubMed]
- Laschke, M.W.; Menger, M.D. Spheroids as vascularization units: From angiogenesis research to tissue engineering applications. Biotechnol. Adv. 2017, 35, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, G.; Gamble, H.J. An electron microscopic study of the pericytes of the developing capillaries in human fetal brain and muscle. J. Anat. 1979, 128, 155–168. [Google Scholar]
- Díaz-Flores, L.; Gutiérrez, R.; Varela, H.; Rancel, N.; Valladares, F. Microvascular pericytes: A review of their morphological and functional characteristics. Histol. Histopathol. 1991, 6, 269–286. [Google Scholar] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; Díaz-Flores, L., Jr.; Valladares, F.; Madrid, J.F. Ultrastructure of myopericytoma: A continuum of transitional phenotypes of myopericytes. Ultrastruct. Pathol. 2012, 36, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Bigler, M.; Koutsantonis, D.; Odriozola, A.; Halm, S.; Tschanz, S.A.; Zakrzewicz, A.; Weichert, A.; Baum, O. Morphometry of skeletal muscle capillaries: The relationship between capillary ultrastructure and ageing in humans. Acta Physiol. 2016, 218, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Morgenstern, E.; Neumann, K.; Patscheke, H. The exocytosis of human blood platelets. A fast freezing and freeze-substitution analysis. Eur. J. Cell Biol. 1987, 43, 273–282. [Google Scholar] [PubMed]
- Russo, I. The prothrombotic tendency in metabolic syndrome: Focus on the potential mechanisms involved in impaired haemostasis and fibrinolytic balance. Scientifica 2012, 2012, 525374. [Google Scholar] [CrossRef]
- Fitch-Tewfik, J.L.; Flaumenhaft, R. Platelet granule exocytosis: A comparison with chromaffin cells. Front. Endocrinol. 2013, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Heijnen, H.; van der Sluijs, P. Platelet secretory behaviour: As diverse as the granules… or not? J. Thromb. Haemost. 2015, 13, 2141–2151. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.D.P.; Sáez, F.J.; Díaz-Flores, L., Jr.; Madrid, J.F. Piecemeal mechanism combining sprouting and intussusceptive angiogenesis in intravenous papillary formation induced by PGE2 and glycerol. Anat. Rec. 2017, 300, 1781–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burri, P.H. Intussusceptive microvascular growth, a new mechanism of capillary network formation. EXS 1992, 61, 32–39. [Google Scholar] [PubMed]
- Patan, S.; Munn, L.L.; Jain, R.K. Intussusceptive microvascular growth in a human colon adenocarcinoma xenograft: A novel mechanism of tumor angiogenesis. Microvasc. Res. 1996, 51, 260–272. [Google Scholar] [CrossRef] [PubMed]
- Djonov, V.G.; Kurz, H.; Burri, P.H. Optimality in the developing vascular system: Branching remodeling by means of intussusception as an efficient adaptation mechanism. Dev. Dyn. 2002, 224, 391–402. [Google Scholar] [CrossRef]
- Djonov, V.; Baum, O.; Burri, P.H. Vascular remodeling by intussusceptive angiogenesis. Cell Tissue Res. 2003, 314, 107–117. [Google Scholar] [CrossRef]
- Burri, P.H.; Hlushchuk, R.; Djonov, V. Intussusceptive angiogenesis: Its emergence, its characteristics, and its significance. Dev. Dyn. 2004, 231, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Paku, S.; Dezso, K.; Bugyik, E.; Tóvári, J.; Tímár, J.; Nagy, P.; Laszlo, V.; Klepetko, W.; Döme, B. A new mechanism for pillar formation during tumor-induced intussusceptive angiogenesis: Inverse sprouting. Am. J. Pathol. 2011, 179, 1573–1585. [Google Scholar] [CrossRef] [PubMed]
- De Spiegelaere, W.; Casteleyn, C.; Van den Broeck, W.; Plendl, J.; Bahramsoltani, M.; Simoens, P.; Djonov, V.; Cornillie, P. Intussusceptive angiogenesis: A biologically relevant form of angiogenesis. J. Vasc. Res. 2012, 49, 390–404. [Google Scholar] [CrossRef] [PubMed]
- Van Gils, J.M.; da Costa Martins, P.A.; Mol, A.; Hordijk, P.L.; Zwaginga, J.J. Transendothelial migration drives dissociation of platelet monocyte complexes. Thromb. Haemost. 2008, 100, 271–279. [Google Scholar] [PubMed]
- Kraemer, B.F.; Borst, O.; Gehring, E.M.; Schoenberger, T.; Urban, B.; Ninci, E.; Seizer, P.; Schmidt, C.; Bigalke, B.; Koch, M.; et al. PI3 kinase-dependent stimulation of platelet migration by stromal cell-derived factor 1 (SDF-1). J. Mol. Med. 2010, 88, 1277–1288. [Google Scholar] [CrossRef] [PubMed]
- Witte, A.; Rohlfing, A.K.; Dannenmann, B.; Dicenta, V.; Nasri, M.; Kolb, K.; Sudmann, J.; Castor, T.; Rath, D.; Borst, O.; et al. The chemokine CXCL14 mediates platelet function and migration via direct interaction with CXCR4. Cardiovasc. Res. 2021, 117, 903–917. [Google Scholar] [CrossRef]
- Diaz-Flores, L.; Dominguez, C. Relation between arterial intimal thickening and the vasa-vasorum. Virchows Arch. A 1985, 406, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Madrid, J.F.; Díaz-Flores, L.; Gutiérrez, R.; Varela, H.; Valladares, F.; Rancel, N.; Rodríguez, F. Participation of angiogenesis from rat femoral veins in the neovascularization of adjacent occluded arteries. Histol. Histopathol. 1998, 13, 1–11. [Google Scholar] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González-Gómez, M.; Díaz-Flores, L., Jr.; Gayoso, S.; Carrasco, J.L.; Álvarez-Argüelles, H. Ultrastructural Study of Platelet Behavior and Interrelationship in Sprouting and Intussusceptive Angiogenesis during Arterial Intimal Thickening Formation. Int. J. Mol. Sci. 2021, 22, 13001. https://doi.org/10.3390/ijms222313001
Díaz-Flores L, Gutiérrez R, García MP, González-Gómez M, Díaz-Flores L Jr., Gayoso S, Carrasco JL, Álvarez-Argüelles H. Ultrastructural Study of Platelet Behavior and Interrelationship in Sprouting and Intussusceptive Angiogenesis during Arterial Intimal Thickening Formation. International Journal of Molecular Sciences. 2021; 22(23):13001. https://doi.org/10.3390/ijms222313001
Chicago/Turabian StyleDíaz-Flores, Lucio, Ricardo Gutiérrez, Maria Pino García, Miriam González-Gómez, Lucio Díaz-Flores, Jr., Sara Gayoso, Jose Luis Carrasco, and Hugo Álvarez-Argüelles. 2021. "Ultrastructural Study of Platelet Behavior and Interrelationship in Sprouting and Intussusceptive Angiogenesis during Arterial Intimal Thickening Formation" International Journal of Molecular Sciences 22, no. 23: 13001. https://doi.org/10.3390/ijms222313001
APA StyleDíaz-Flores, L., Gutiérrez, R., García, M. P., González-Gómez, M., Díaz-Flores, L., Jr., Gayoso, S., Carrasco, J. L., & Álvarez-Argüelles, H. (2021). Ultrastructural Study of Platelet Behavior and Interrelationship in Sprouting and Intussusceptive Angiogenesis during Arterial Intimal Thickening Formation. International Journal of Molecular Sciences, 22(23), 13001. https://doi.org/10.3390/ijms222313001