
Juvenile African Clawed Frogs (Xenopus laevis) Express Growth, Metamorphosis, Mortality, Gene Expression, and Metabolic Changes When Exposed to Thiamethoxam and Clothianidin

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Methods
2.1. Animals and Husbandry
2.2. Exposures and Biological Endpoints
2.3. Dissection
2.4. RNA Extraction
2.5. Primer Development, Quantitative Real Time Polymerase Chain Reaction (qrtPCR) Assays, and Gene Expression Analysis
2.6. Sequencing
2.7. Liver Metabolism and Flow Cytometry
2.8. Statistical Analyses
3. Results
3.1. NEO Concentrations
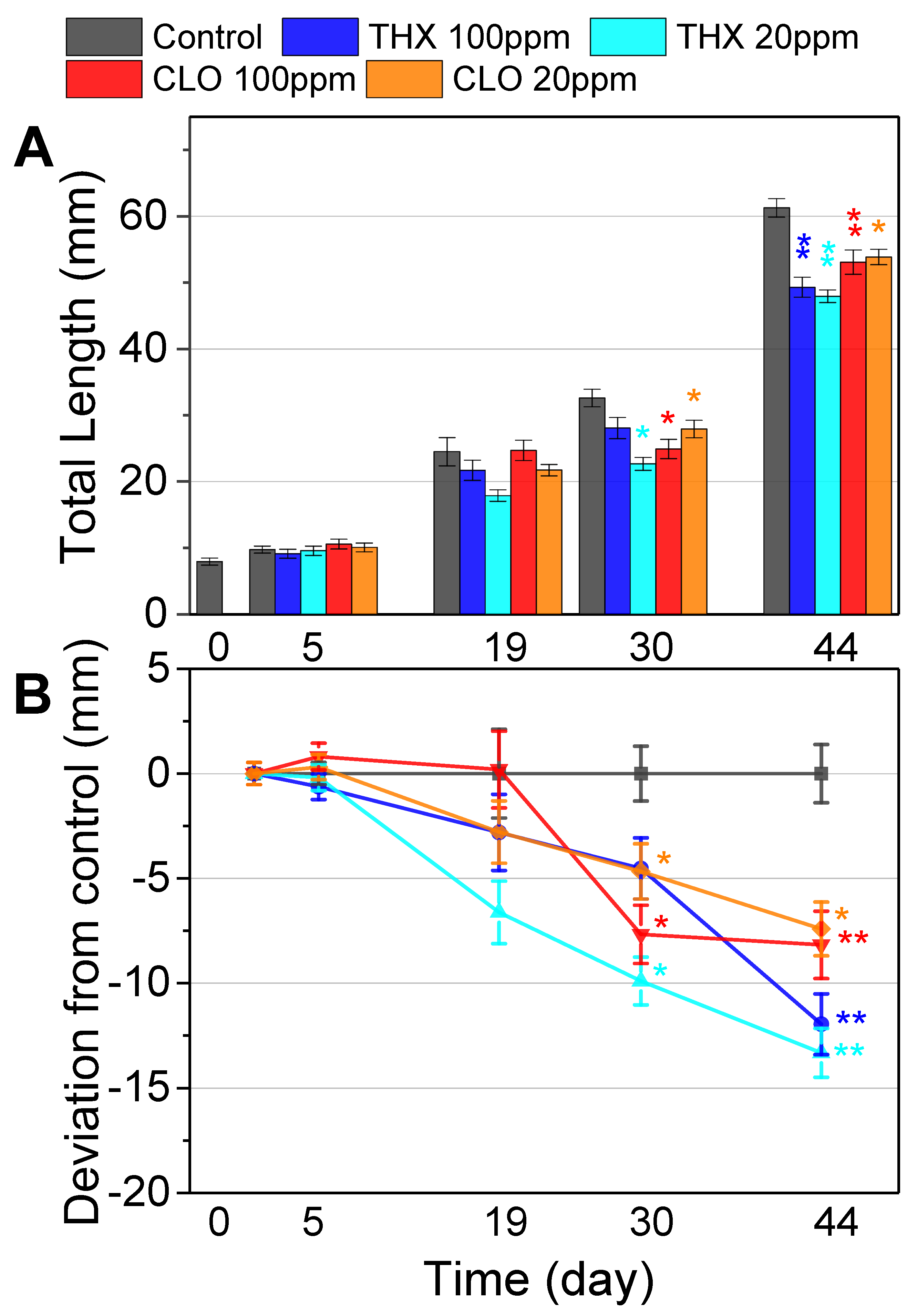
3.2. Incremental Tadpole Lengths: Over the First Exposure Period
3.3. Morphometrics: End of the First Exposure Period
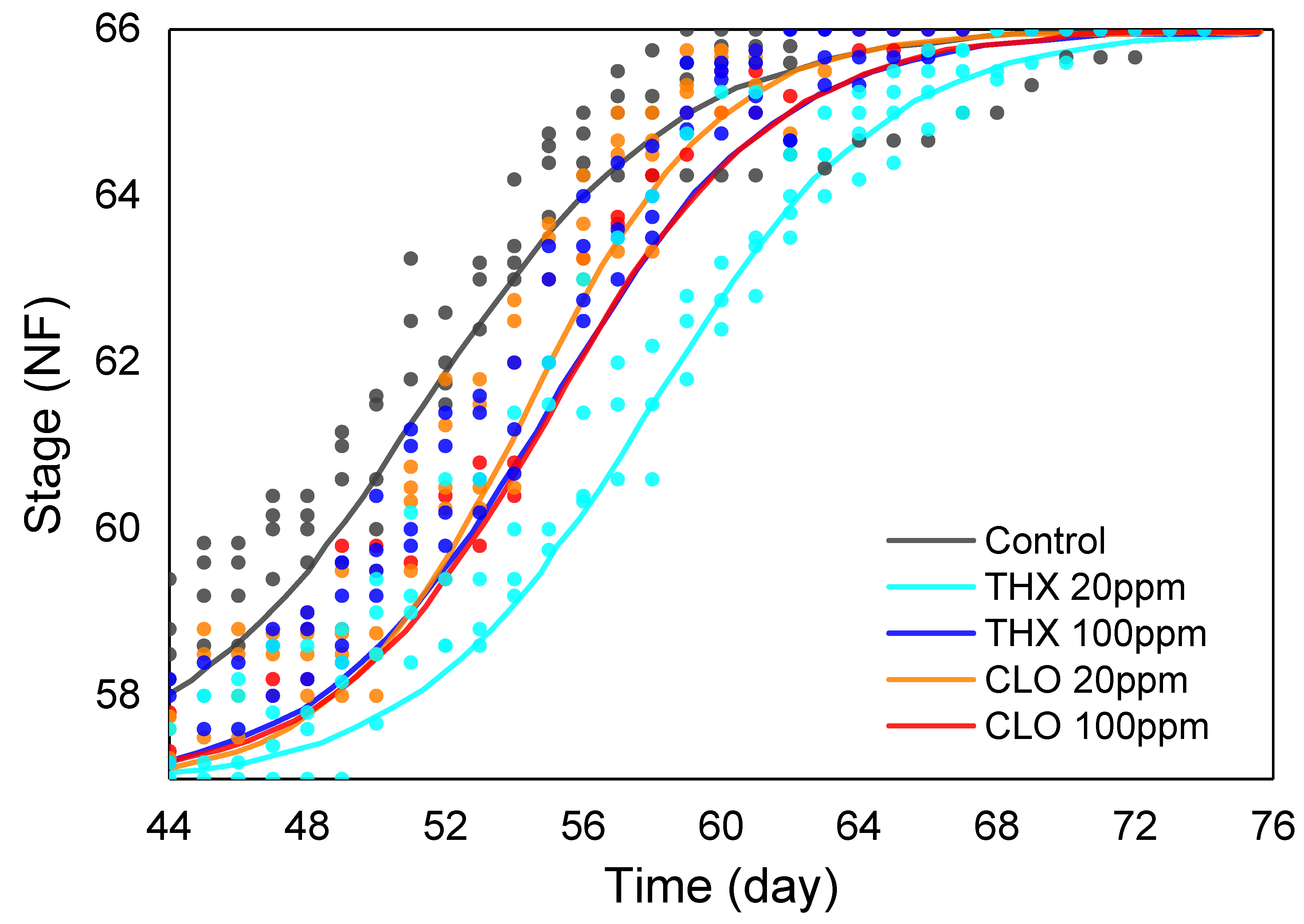
3.4. Developmental Stage: Over the Second Exposure Period
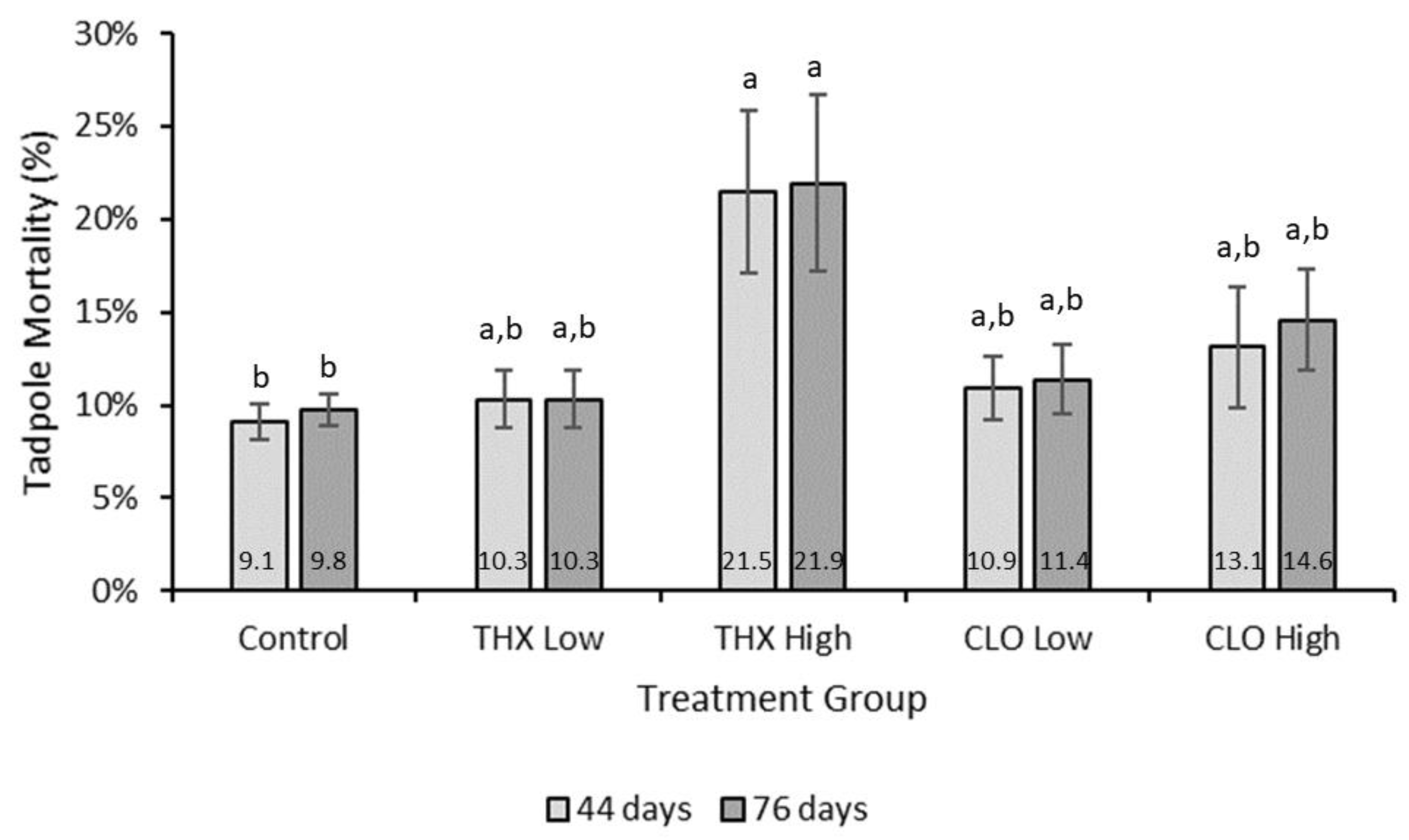
3.5. Mortality
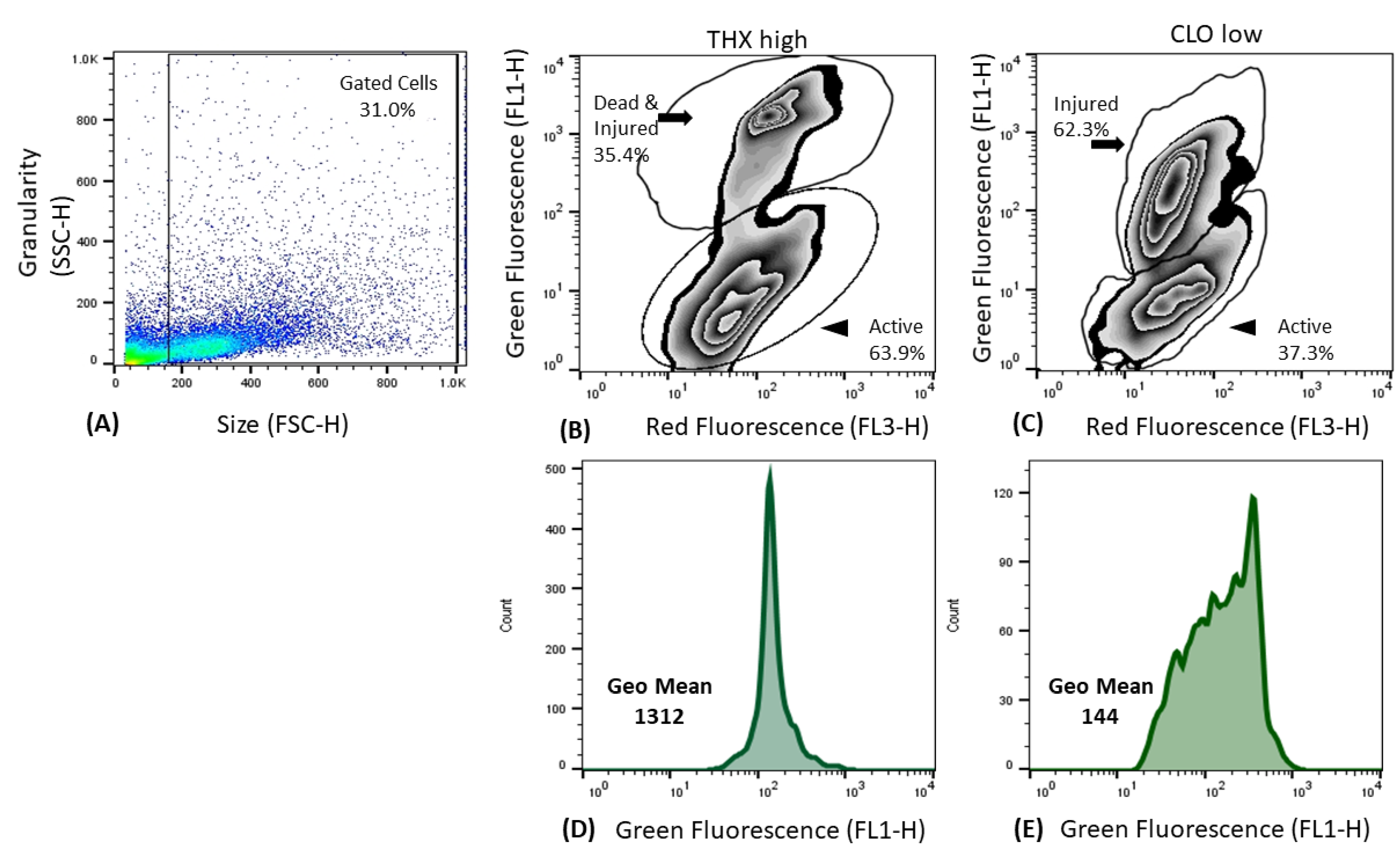
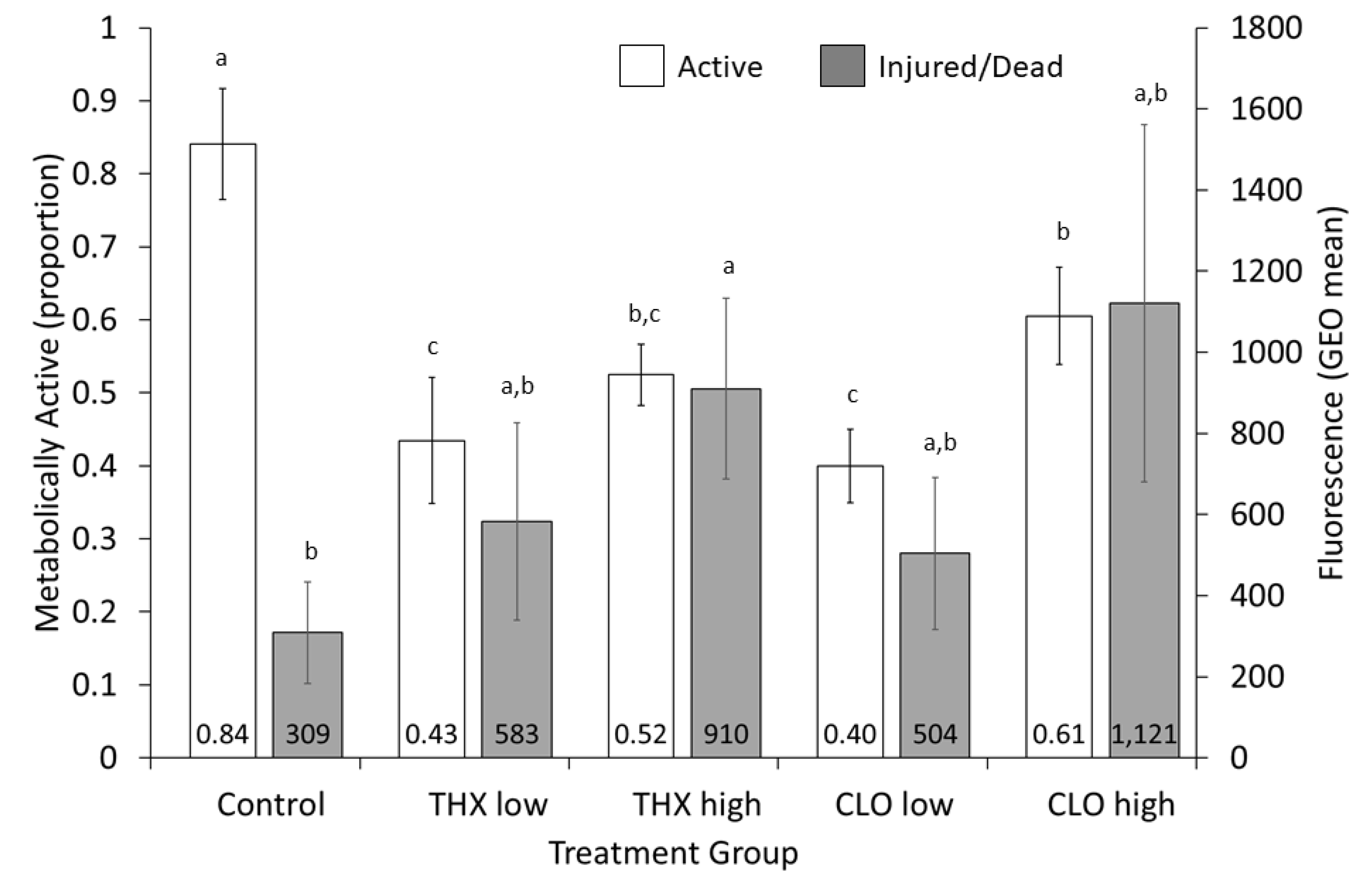
3.6. Liver Metabolism
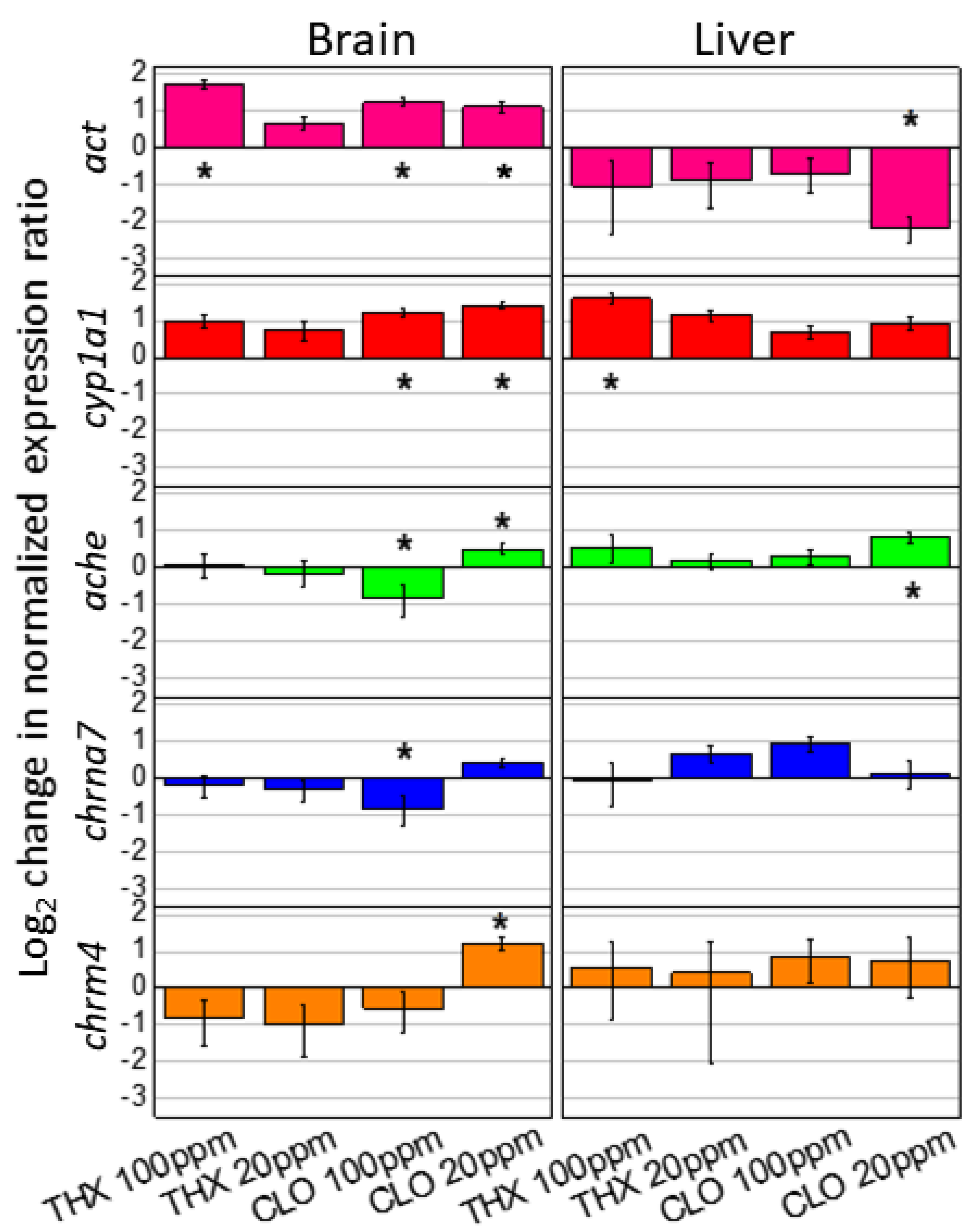
3.7. Gene Expression
4. Discussion
4.1. Survival
4.2. Growth and Development
4.3. Liver Cell Function
4.4. Gene Expression
4.4.1. Reference Genes and Cyp
4.4.2. Receptors and Acetylcholinesterase
5. Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alford, R.A.; Richards, S.J. Global amphibian declines: A problem in applied ecology. Annu. Rev. Ecol. Syst. 1999, 30, 133–165. [Google Scholar] [CrossRef] [Green Version]
- Smalling, K.L.; Reeves, R.; Muths, E.; Vandever, M.; Battaglin, W.A.; Hladik, M.L.; Pierce, C.L. Pesticide concentrations in frog tissue and wetland habitats in a landscape dominated by agriculture. Sci. Total Environ. 2015, 502, 80–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regnault, C.; Usal, M.; Veyrenc, S.; Couturier, K.; Batandier, C.; Bulteau, A.; Legjon, D.; Sapin, A.; Combourieu, B.; Chetiveaux, M.; et al. Unexpected metabolic disorders induced by endocrine disruptors in Xenopus tropicalis provide new lead for understanding amphibian decline. Proc. Natl. Acad. Sci. USA 2018, 115, E4416–E4425. [Google Scholar] [CrossRef] [Green Version]
- Gallant, A.L.; Klaver, R.W.; Casper, G.S.; Lanno, M.J. Global rates of habitat loss and implications for amphibian conservation. Copeia 2007, 4, 967–979. [Google Scholar] [CrossRef]
- Mann, R.M.; Hyne, R.V.; Choung, C.B.; Wilson, S.P. Amphibians and agricultural chemicals: Review of the risks in a complex environment. Environ. Pollut. 2009, 157, 2903–2927. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.J.; Bancroft, B.A.; Garcia, T.S. A meta-analysis of the effects of pesticides and fertilizers on survival and growth of amphibians. Sci. Total Environ. 2013, 449, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Rohr, J.R.; McCoy, K.A. A qualitative meta-analysis reveals consistent effects of atrazine on freshwater fish and amphibians. Environ. Health Perspect. 2010, 118, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Boone, M.D.; Bridges, C.M. Effects of carbaryl on green frog (Rana clamitans) tadpoles: Timing of exposure versus multiple exposures. Environ. Toxicol. Chem. 2003, 22, 2695–2702. [Google Scholar] [CrossRef]
- Mills, N.E.; Semlitsch, R.D. Competition and predation mediate the indirect effects of an insecticide on southern leopard frogs. Ecol. Appl. 2004, 14, 1041–1054. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, D.; Morrissey, C.; Mineau, P. A review of the direct and indirect effects of neonicotinoids and fipronil on vertebrate wildlife. Environ. Sci. Pollut. Res. 2015, 22, 103–118. [Google Scholar] [CrossRef] [Green Version]
- Perez-Iglesias, J.M.; Ruiz de Arcaute, C.; Nikoloff, N.; Dury, L.; Soloneski, S.; Natale, G.S.; Larramendy, M.L. The genotoxic effects of the imidacloprid-based insecticide formulation Glacoxan Imida on Montevideo tree frog Hypsiboas pulchellus tadpoles (Anura, Hylidae). Ecotoxicol. Environ. Saf. 2014, 104, 120–126. [Google Scholar] [CrossRef]
- Holtswarth, J.N.; Rowland, F.E.; Puglis, H.J.; Hladik, M.L.; Webb, E.B. Effects of the neonicotinoid insecticide clothianidin on Southern leopard frog (Rana sphenocephala) tadpole behavior. Bull. Environ. Contam. Toxicol. 2019, 103, 717–722. [Google Scholar] [CrossRef]
- Lee-Jenkins, S.S.Y.; Robinson, S.A. Effects of neonicotinoids on putative escape behavior of juvenile wood frogs (Lithobates sylvaticus) chronically exposed as tadpoles. Environ. Toxicol. Chem. 2018, 37, 3115–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ankley, G.T.; Bennett, R.S.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K.; et al. Adverse outcome pathways: A conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. 2010, 29, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Knapen, D.; Angrish, M.M.; Fortin, M.C.; Katsiadaki, I.; Leonard, M.; Margiotta-Casaluci, L.; Munn, S.; O’Brien, J.M.; Pollesch, N.; Smith, L.C.; et al. Adverse outcome pathway networks I: Development and applications. Environ. Toxicol. Chem. 2018, 37, 1723–1733. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, C.A.; Mineau, P.; Devries, J.H.; Sanchez-Bayo, F.; Liess, M.; Cavallaro, M.C.; Liber, K. Neonicotinoid contamination of global surface waters and associated risk to aquatic invertebrates: A review. Environ. Int. 2015, 74, 291–303. [Google Scholar] [CrossRef]
- Sparks, T.C. Insecticide discovery: An evaluation and analysis. Pestic. Biochem. Physiol. 2013, 107, 8–17. [Google Scholar] [CrossRef]
- Rios, F.M.; Wilcoxen, T.E.; Zimmerman, L.M. Effects of imidacloprid on Rana catesbeiana immune and nervous system. Chemosphere 2017, 188, 465–469. [Google Scholar] [CrossRef]
- Tomizawa, M.; Casida, J.E. Neonicotinoid insecticide toxicology: Mechanisms of selective action. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 247–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, W.; Tian, Y.; Shen, X. Human exposure to neonicotinoid insecticides and the evaluation of their potential toxicity: An overview. Chemosphere 2018, 192, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D. An overview of the environmental risks posed by neonicotinoid insecticides. J. Appl. Ecol. 2013, 50, 977–987. [Google Scholar] [CrossRef]
- DeCant, J.; Barrett, M. Clothianidin Registration of Prosper T400 Seed Treatment on Mustard Seed (Oilseed and Condiment) and Poncho/Votivo Seed Treatment on Cotton; U.S. Environmental Protection Agency: Washington, DC, USA, 2010; p. 101. Available online: https://archive.epa.gov/pesticides/chemicalsearch/chemical/foia/web/pdf/044309/044309-2010-11-02b.pdf (accessed on 17 November 2021).
- Bradley, P.M.; Journey, C.A.; Romanok, K.M.; Barber, L.B.; Buxton, H.T.; Foreman, W.T.; Furlong, E.T.; Glassmeyer, S.T.; Hladik, M.L.; Iwanowicz, L.R.; et al. Expanded target-chemical analysis reveals extensive mixed-organic-contaminant exposure in U.S. streams. Environ. Sci. Technol. 2017, 51, 4792–4802. [Google Scholar] [CrossRef] [PubMed]
- Miles, J.C.; Hua, J.; Sepulveda, M.S.; Krupke, C.H.; Hoverman, J.T. Effects of clothianidin on aquatic communities: Evaluating the impacts of lethal and sublethal exposure to neonicotinoids. PLoS ONE 2017, 13, e0174171. [Google Scholar] [CrossRef] [PubMed]
- Basley, K.; Goulson, D. Neonicotinoids thiamethoxam and clothianidin adversely affect the colonisation of invertebrate populations in aquatic ecosystems. Environ. Sci. Pollut. Res. 2018, 25, 9593–9599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hladik, M.L.; Kolpin, D.W.; Kuivila, K.M. Widespread occurrence of neonicotinoid insecticides in streams in a high corn and soybean producing region, USA. Environ. Pollut. 2014, 193, 189–196. [Google Scholar] [CrossRef]
- Hladik, M.L.; Kolpin, D.W. First national-scale reconnaissance of neonicotinoid insecticides in streams across the USA. Environ. Chem. 2015, 13, 12–20. [Google Scholar] [CrossRef]
- Relyea, R.A.; Diecks, N. An unforeseen chain of events: Lethal effects of pesticides on frogs at sublethal concentrations. Ecol. Appl. 2008, 18, 1728–1742. [Google Scholar] [CrossRef] [Green Version]
- Linder, G.; Lehman, C.M.; Bidwell, J.R. Ecotoxicology of amphibians and reptiles in a nutshell. In Ecotoxicology of Amphibians and Reptiles, 2nd ed.; Sparling, D.E., Linder, G., Bishop, C.A., Krest, S.K., Eds.; CRC Press: New York, NY, USA, 2010; pp. 69–103. [Google Scholar]
- Gavel, M.J.; Richardson, S.D.; Dalton, R.L.; Soos, C.; Ashby, B.; McPhee, L.; Forbes, M.R.; Robinson, S.A. Effects of 2 neonicotinoid insecticides on blood cell profiles and corticosterone concentrations of wood frogs (Lithobates sylvaticus). Environ. Toxicol. Chem. 2019, 38, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Samson-Robert, O.; Labrie, G.; Chagnon, M.; Fournier, V. Neonicotinoid-contaminated puddles of water represent a risk of intoxication for honey bees. PLoS ONE 2014, 9, e108443. [Google Scholar] [CrossRef] [Green Version]
- Nieuwkoop, P.D.; Faber, J. Normal Table of Xenopus laevis (Daudin): A Systematical and Chronological Survey of the Development from the Fertilized Egg till the End of Metamorphosis; North-Holland Publishing Company: Amsterdam, The Netherlands, 1967. [Google Scholar]
- Miko, Z.; Ujszegi, J.; Hettyey, A. Age-dependent changes in sensitivity to a pesticide in tadpoles of the common toad (Bufo bufo). Aquat. Toxicol. 2017, 187, 48–54. [Google Scholar] [CrossRef]
- Regnault, C.; Worms, I.A.M.; Oger-Desfeux, C.; MedlodeLima, C.; Veyrenc, S.; Bayle, M.; Combourieu, B.; Bonin, A.; Renaud, J.; Raveton, M.; et al. Impaired liver function in Xenopus tropicalis exposed to benzo{a}pyrene: Transcriptomic and metabolic evidence. BMC Genom. 2014, 15, 666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glennemeier, K.A.; Denver, R.J. Developmental changes in interrenal responsiveness in anuran amphibians. Integr. Comp. Biol. 2002, 42, 565–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollins-Smith, L.A. Amphibian immunity-stress, disease, and climate change. Dev. Comp. Immunol. 2017, 66, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Rollins-Smith, L.A. Metamorphosis and the amphibian immune system. Immunol. Rev. 1998, 166, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Relyea, R.A. Growth and survival of five amphibian species exposed to combinations of pesticides. Environ. Toxicol. Chem. 2004, 23, 1737–1742. [Google Scholar] [CrossRef]
- Boone, M.D.; Semlitsch, R.D.; Fairchild, J.F.; Rothermel, B.B. Effects of an insecticide on amphibians in large-scale experimental ponds. Ecol. Appl. 2004, 14, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.A.; Richardson, S.D.; Dalton, R.L.; Maissonneuve, F.; Trudeau, V.L.; Paulie, B.D.; Lee-Jenkins, S.S.Y. Sublethal effects on wood frogs chronically exposed to environmentally relevant concentrations of two neonicotinoid insecticides. Environ. Toxicol. Chem. 2017, 36, 1101–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Sluijs, J.P.; Amaral-Rogers, V.; Belzunces, L.P.; van Lexmond, M.F.I.J.B.; Bonmatin, J.-M.; Chagnon, M.; Downs, C.A.; Furlan, L.; Gibbons, D.W.; Giorio, C.; et al. Conclusions of the worldwide integrated assessment on the risks neonicotinoids and fipronil to biodiversity and ecosystem functioning. Environ. Sci. Pollut. Res. 2014, 22, 148–154. [Google Scholar] [CrossRef]
- Anderson, J.C.; Dubetz, C.; Palace, V.P. Neonicotinoids in the Canadian aquatic environment: A literature review on current use products with a focus on fate, exposure, and biological effects. Sci. Total Environ. 2015, 505, 409–422. [Google Scholar] [CrossRef]
- Swenson, T.L.; Casida, J.E. Neonicotinoid formaldehyde generators: Possible mechanism of mouse-specific hepatotoxicity/hepatocarcinogenicity of thiamethoxam. Toxicol. Lett. 2013, 216, 139–145. [Google Scholar] [CrossRef]
- Hoff, D.; Lehmann, W.; Pease, A.; Raimondo, S.; Russom, C.L.; Steeger, T. Predicting the Toxicities of Chemicals to Aquatic Animal Species; U.S. Environmental Protection Agency: Washington, DC, USA, 2010; pp. 1–127. Available online: https://www.epa.gov/sites/default/files/2015-08/documents/predicting_the_toxicities_of_chemicals_to_aquatic_animal_species.pdf (accessed on 17 November 2021).
- Bray, C.; Son, J.H.; Kumar, P.; Meizel, S. Mice deficient in CHRNA7, a subunit of the nicotinic acetylcholine receptor, produce sperm with impaired motility. J. Reprod. 2005, 73, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R.; Ebbinghaus-Kintscher, U.; Salgado, V.L.; Kaussmann, M. Thiamethoxam is a neonicotinoid precursor converted to clothianidin in insects and plants. Pestic. Biochem. Physiol. 2003, 76, 55–69. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Li, W.C.; Han, Y.; Guo, B. Study on neurotoxicity of dinotefuran, thiamethoxam and imidacloprid against Chinese lizards (Eremias argus). Chemosphere 2019, 217, 150–157. [Google Scholar] [CrossRef]
- Boily, M.; Sarrain, B.; DeBois, C.; Aras, P.; Chagnon, M. Acetylcholinesterase in honey bees (Apis mellifera) exposed to neonicotinoids, atrazine and glyphosate: Laboratory and field experiments. Environ. Sci. Pollut. Res. 2013, 20, 5603–5614. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Wang, C.; Chen, X.; Qin, Z.; Li, X.; Wang, C. Toxicity assessments with Daphnia magna of Guadipyr, a new neonicotinoid insecticide and studies of its effect on acetylcholinesterase (AChE), glutathione S-transferase (GST), catalase (CAT) and chitobiase activities. Ecotoxicol. Environ. Saf. 2013, 98, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Fulton, M.H.; Key, P.B. Acetylcholinesterase inhibition in estuarine fish and invertebrates as an indicator of organophosphorus insecticide exposure and effects. Environ. Toxicol. Chem. 2001, 20, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, M.; Casida, J.E. Imidacloprid, thiacloprid, and their imine derivatives up-regulate the α4β2 nicotinic acetylcholine receptor in M10 cells. Toxicol. Appl. Pharmacol. 2000, 169, 114–120. [Google Scholar] [CrossRef]
- Faro, L.R.F.; Tak-Kim, H.; Alfonso, M.; Duran, R. Clothianidin, a neonicotinoid insecticide, activates α4β2, α7 and muscarinic receptors to induce in vivo dopamine release from rat striatum. Toxicology 2019, 426, 152285. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A. Acetylcholine and cholinergic receptors. Brain Neurosci. Adv. 2019, 3, 1–10. [Google Scholar] [CrossRef]
- Iwamoto, D.V.; Kurylo, C.M.; Schorling, K.M.; Powell, W.H. Induction of cytochrome P450 family 1 mRNAs and activities in a cell line from the frog Xenopus laevis. Aquat. Toxicol. 2012, 114–115, 165–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparling, D.W. Ecotoxicology of organic contaminants to amphibians. In Ecotoxicology of Amphibians and Reptiles, 2nd ed.; Sparling, D.E., Linder, G., Bishop, C.A., Krest, S.K., Eds.; Society of Environmental Toxicology and Chemistry: Pensacola, FL, USA, 2010; pp. 261–288. [Google Scholar]
- Van den Brink, N.W.; Lee-de Groot, M.B.E.; de Bie, P.A.F.; Bosveld, A.T.C. Enzyme markers in frogs (Rana spp.) for monitoring risk of aquatic pollution. Aquat. Ecosyst. Health Manag. 2003, 6, 441–448. [Google Scholar] [CrossRef]
- Caron-Beaudoin, E.; Denison, M.S.; Sanderson, J.T. Effects of neonicotinoids on promoter-specific expression and activity of aromatase (CYP19) in human adrenocortical carcinoma (H295R) and primary umbilical vein endothelial (HUVEC) cells. Toxicol. Sci. 2016, 149, 134–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, H.; Tomizawa, M.; Casida, J.E. Neonicotinoid metabolic activation and inactivation established with coupled nicotinic receptor-CYP3A4 and -aldehyde oxidase systems. Toxicol. Lett. 2006, 161, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, A.; Ogilvie, B.W.; Buckley, D.B.; Kazmi, F.; Czerwinski, M.; Parkinson, O. Biotransformation of xenobiotics. In Casarett & Doull’s Essentials of Toxicology, 3rd ed.; Klaassen, C.D., Watkins, J.B., Eds.; McGraw-Hill Education: New York, NY, USA, 2015. [Google Scholar]
- Karimi, K.; Fortriede, J.D.; Lotay, V.S.; Burns, K.A.; Wang, D.Z.; Fisher, M.E.; Pells, T.J.; James-Zorn, C.; Wang, Y.; Ponferrada, V.G.; et al. Xenbase: A genomic, epigenomic and transcriptomic model organism database. Nucleic Acids Res. 2018, 46, D8610–D8868. [Google Scholar] [CrossRef]
- Use of Fishes in Research Committee (Joint Committee of the American Fisheries Society, the American Institute of Fishery Research Biologists, and the American Society of Ichthyologists and Herpetologists); Jenkins, J.A.; Bart, H., Jr.; Bowker, J.D.; Bowser, P.; MacMillan, J.; Nickum, J.; Rose, J.; Sorensen, P. Guidelines for the Use of Fishes in Research; American Fisheries Society: Bethesda, MD, USA, 2014; 90p, Available online: https://fisheries.org/policy-media/science-guidelines/guidelines-for-the-use-of-fishes-in-research/ (accessed on 15 January 2016).
- Herpetological Animal Care and Use Committee (HACC) of the American Society of Ichthyologists and Herpetologists; Beaupre, S.J.; Jacobson, E.R.; Lillywhite, H.B.; Zamudio, K. Guidelines for Use of Live Amphibians and Reptiles in Field and Laboratory Research, 2nd ed.; American Society of Ichthyologists and Herpetologists: Lawrence, KS, USA, 2004; 43p, Available online: https://asih.org/sites/default/files/2018-05/guidelines_herps_research_2004.pdf (accessed on 2 December 2021).
- Gurdon, J.B. Normal table of Xenopus laevis (Daudin): Edited by P.D. Nieuwkoop and J. Faber Garland Publishing, 1994. Trends Genet. 1995, 11, 418. [Google Scholar] [CrossRef]
- Sive, H.L.; Grainger, R.; Harland, R.M. Early Development of Xenopus laevis: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2000. [Google Scholar]
- Gurdon, J.B. Methods for nuclear transplantation in amphibia. Methods Cell Biol. 1977, 16, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Browne, R.K.; Zippel, K. Reproduction and larval rearing of amphibians. ILAR J. 2007, 48, 212–234. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Challis, J.K.; Wong, C.S. Quantum yields for direct photolysis of neonicotinoid insecticides in water: Implications for exposure to nontarget aquatic organisms. Environ. Sci. Technol. Lett. 2015, 2, 188–192. [Google Scholar] [CrossRef]
- Finnegan, M.H.; Baxter, L.R.; Maul, J.D.; Hanson, M.L.; Hoekstra, J.F. Comprehensive characterization of the acute and chronic toxicity of the neonicotinoid insecticide thiamethoxam to a suite of aquatic primary producers, invertebrates, and fish. Environ. Toxicol. Chem. 2017, 36, 2838–2848. [Google Scholar] [CrossRef] [Green Version]
- U.S. Environmental Protection Agency (USEPA). Technical Overview of Ecological Risk Assessment—Analysis Phase: Exposure Characterization. Available online: https://www.epa.gov/pesticide-science-and-assessing-pesticide-risks/technical-overview-ecological-risk-assessment-1 (accessed on 4 August 2019).
- USEPA. Pesticide Ecotoxicity Database (Formerly Environmental Effects Database; EEDB), Ecoref #344. Washington DC, 1992. 2019. Available online: https://ecotox.ipmcenters.org/ (accessed on 12 November 2021).
- De Perre, C.; Murphy, T.M.; Lydy, M.J. Fate and effects of clothianidin in fields using conservation practices. Environ. Toxicol. Chem. 2015, 34, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Padilla, S.; Corum, D.; Padnos, B.; Hunter, D.L.; Beam, A.; Houck, K.A.; Sipes, N.; Kleinstreuer, N.; Knudsen, T.B.; Dix, D.J.; et al. Zebrafish developmental screening of the ToxCastTM Phase I chemical library. Reprod. Toxicol. 2012, 33, 174–187. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency. Guidance for Using non-Definitive Endpoints in Evaluating Risks to Listed and Non-Listed Animal Species; U.S. Environmental Protection Agency: Washington, DC, USA, 2011. Available online: https://www.epa.gov/pesticide-science-and-assessing-pesticide-risks/guidance-using-non-definitive-endpoints-evaluating (accessed on 12 November 2021).
- Stehle, S.; Schulz, R. Agricultural insecticides threaten surface waters at the global scale. Proc. Natl. Acad. Sci. USA 2015, 112, 5750–5755. [Google Scholar] [CrossRef] [Green Version]
- Stehle, S.; Bub, S.; Schulz, R. Compilation and analysis of global surface water concentrations for individual insecticide compounds. Sci. Total Environ. 2018, 639, 516–525. [Google Scholar] [CrossRef]
- Hua, J.; Jones, D.K.; Mattes, B.M.; Cothran, R.D.; Relyea, R.A.; Hoverman, J.T. Evolved pesticide tolerance in amphibians: Predicting mechanisms based on pesticide novelty and mode of action. Environ. Pollut. 2015, 206, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Tennekes, H.A.; Sanchez-Bayo, F. The molecular basis of simple relationships between exposure concentration and toxic effects with time. Toxicology 2013, 309, 39–51. [Google Scholar] [CrossRef]
- USEPA. ECOTOXicology Database System. Available online: https://cfpub.epa.gov/ecotox/ (accessed on 16 September 2019).
- Hladik, M.L.; Calhoun, D.L. Analysis of the Herbicide Diuron, Three Diuron Degradates, and Six Neonicotinoid Insecticides in Water—Method Details and Application to Two Georgia Streams; U.S. Geological Survey Scientific Investigations Report 2012-5206; 2012; 10p. Available online: http://pubs.usgs.gov/sir/2012/5206/ (accessed on 15 September 2017).
- Jenkins, J.A.; Brown, B.L. Xenopus metamorphosis after neonicotinoid exposure. U.S. Geol. Surv. Data Release 2021. [Google Scholar] [CrossRef]
- Jenkins, J.A.; Brown, B.L. Gene expression and liver cell metabolism from Xenopus laevis tadpoles exposed to neonicotinoids. U.S. Geol. Surv. Data Release 2021. [Google Scholar] [CrossRef]
- Wright, K.M.; Whitaker, B.R. Amphibian Medicine and Captive Husbandry; Krieger Publishing Company: Malabar, FL, USA, 2001; 499p. [Google Scholar]
- Rio, D.C.; Ares, M., Jr.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 6, pdb.prot5439. [Google Scholar] [CrossRef]
- Benson, D.A.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2009, 38, D46–D51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, B.; Basu, C. Real-time PCR (qPCR) primer design using free online software. Biochem. Mol. Biol. Educ. 2011, 39, 145–154. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickinson, A.J.G.; Sive, H. Development of the primary mouth in Xenopus laevis. Dev. Biol. 2006, 295, 700–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Cook, P.; Fu, C.; Hickey, M.; Han, E.-S.; Miller, K.S. SAS programs for real-time RT-PCR having multiple independent samples. BioTechniques 2018, 37, 990–995. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1984; 680p. [Google Scholar]
- Rieu, I.; Power, S.J. Real-time quantitative RT-PCR: Design, calculations, and statistics. Plant Cell 2009, 21, 1031–1033. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Rugh, R. Culturing of amphibian embryos. In Experimental Embryology: Techniques and Procedures; Burgess: Minneapolis, MN, USA, 1962; pp. 50–51. [Google Scholar]
- JMP, version 13.1.0.; SAS Institute Inc.: Cary, NC, USA, 1989–2021.
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 14 April 2018).
- SAS Institute, Inc. SAS/STAT®15.1 User’s Guide; SAS Institute, Inc.: Cary, NC, USA, 2018. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice-Hall: Hoboken, NJ, USA, 2010; 960p. [Google Scholar]
- Origin(Pro), version 9.0.0.; OriginLab Corporation: Northampton, MA, USA, 2015; Available online: https://origin90.software.informer.com/9.0/ (accessed on 16 May 2016).
- Robinson, S.A.; Chlebak, R.J.; Young, S.D.; Dalton, R.L.; Gavel, M.J.; Prosser, R.S.; Bartlett, A.J.; de Solla, S.R. Clothianidin alters leukocyte profiles and elevates measures of oxidative stress in tadpoles of the amphibian, Rana pipiens. Environ. Pollut. 2021, 284, e117149. [Google Scholar] [CrossRef] [PubMed]
- Hrynyk, M.A.; Brunetti, C.; Kerr, L.; Metcalfe, C.D. Effect of imidacloprid on the survival of Xenopus tadpoles challenged with wild type frog virus 3. Aquat. Toxicol. 2018, 194, 152–158. [Google Scholar] [CrossRef]
- Sievers, M.; Hale, R.; Swearer, S.E.; Parris, K.M. Contaminant mixtures interact to impair predator-avoidance behaviours and survival in a larval amphibian. Ecotoxicol. Environ. Saf. 2018, 161, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Gavel, M.J.; Young, S.D.; Blais, N.; Forbes, M.R.; Robinson, S.A. Trematodes coupled with neonicotinoids: Effects on blood cell profiles of a model amphibian. Parasitol. Res. 2021, 120, 2135–2148. [Google Scholar] [CrossRef]
- Pochini, K.M.; Hoverman, J.T. Reciprocal effects of pesticides and pathogens on amphibian hosts: The importance of exposure order and timing. Environ. Pollut. 2017, 221, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Main, A.R.; Headley, J.V.; Peru, K.M.; Michel, N.L.; Cessna, A.J.; Morrissey, C.A. Widespread use and frequent detection of neonicotinoid insecticides in wetlands of Canada’s prairie pothole region. PLoS ONE 2014, 93, e92821. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Huber, M.; Wang, T.; Dali, W.; Lin, Z.; Chun-Sheng, Y. Progress in the studies on hormesis of low-dose pollutants. Environ. Dis. 2016, 1, 58–64. [Google Scholar] [CrossRef]
- Keller, P. Detection of Imidacloprid in Northern Leopard Frog (Rana pipiens) Brains. Bachelor’s Thesis, University of South Dakota, Vermillion, SD, USA, 2021. [Google Scholar]
- Danis, B.E.G.; Marlatt, V.L. Investigating acute and subchronic effects of neonicotinoids on northwestern salamandar larvae. Arch. Environ. Contam. Toxicol. 2021, 80, 691–707. [Google Scholar] [CrossRef] [PubMed]
- Abou-Donia, M.B. Metabolic biotransformation of xenobiotics. In Mammalian Toxicology; Abou-Donia, M.B., Ed.; John Wiley & Sons Ltd.: West Sussex, UK, 2015; pp. 101–129. [Google Scholar]
- Whyte, J.J.; Jung, R.E.; Schmitt, C.J.; Tillitt, D.E. Ethoxyresorufin-O-Deethylase (EROD) Activity in Fish as a Biomarker of Chemical Exposure; McClellan, R.O., Ed.; CRC Press: Boca Raton, FL, USA, 2000; Volume 30. [Google Scholar]
- Schwen, R.J.; Mannering, G.J. Hepatic cytochrome P-450-dependent monooxygenase systems of the trout, frog and snake—I. Components. Comp. Biochem. Physiol. Part B Comp. Biochem. 1982, 71, 431–436. [Google Scholar] [CrossRef]
- Boell, E.J.; Weber, R. Cytochrome oxidase activity in mitochondria during amphibian development. Exp. Cell Res. 1955, 9, 559–567. [Google Scholar] [CrossRef]
- Matta, J.A.; Gu, S.; Davini, W.; Bredt, D.S. Nicotinic acetylcholine receptor redux: Discovery of accessories open therapeutic vistas. Science 2021, 373, eabg6539. [Google Scholar] [CrossRef]
- Iturburu, F.G.; Zomisch, M.; Panzeri, A.M.; Crupkin, A.C.; Contardo-Jara, V.; Pflugmacher, S.; Menone, M.L. Uptake, distribution in different tissues, and genotoxicity of imidacloprid in the freshwater fish Australoheros facetus. Environ. Toxicol. Chem. 2017, 36, 699–708. [Google Scholar] [CrossRef]
- Perrin, B.J.; Ervasti, J.M. The actin gene family: Function follows isoform. Cytoskeleton 2010, 67, 630–634. [Google Scholar] [CrossRef]
- Malev, O. Toxic Effects of Selected Neonicotinoids through Different Organisational Levels: In Vitro and In Vivo Studies. Doctoral Dissertation, University of Nova Gorica, Nova Gorica, Slovenia, 2012. [Google Scholar]
- Colombo, A.; Orsi, F.; Bonfanti, P. Exposure to the organophosphorus pesticide chlorpyrifos inhibits acetylcholinesterase activity and affects muscular integrity in Xenopus laevis larvae. Chemosphere 2005, 61, 1665–1671. [Google Scholar] [CrossRef] [PubMed]
- Roger, A.J.; Sandblom, O.; Doolittle, W.F.; Philippe, H. An evaluation of elongation factor 1alpha as a phylogenetic marker for eukaryotes. Mol. Biol. Evol. 1999, 16, 218–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunderson, M.P.; Veldhoen, N.; Skirrow, R.C.; Macnab, M.K.; Ding, W.; van Aggelen, G.; Helbing, C.C. Effect of low dose exposure to the herbicide atrazine and its metabolite on cytochrome P450 aromatase and steroidogenic factor-1 mRNA levels in the brain of premetamorphic bullfrog tadpoles (Rana catesbeiana). Aquat. Toxicol. 2011, 102, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feyereisen, R. Insect P450 enzymes. Annu. Rev. Entomol. 1999, 44, 507–533. [Google Scholar] [CrossRef] [PubMed]
- Cothran, R.D.; Brown, J.M.; Relyea, R.A. Proximity to agriculture is correlated with pesticide tolerance: Evidence for the evolution of amphibian resistance to modern pesticides. Evol. Appl. 2013, 6, 832–841. [Google Scholar] [CrossRef]
- Gauthier, M. State of the art on insect nicotinic acetylcholine receptor function in learning and memory. In Insect Nicotinic Acetylcholine Receptors; Thany, S.H., Ed.; Advances in Experimental Medicine and Biology; Landes Bioscience and Springer Science+Business Media, LLC: New York, NY, USA, 2010; pp. 97–115. [Google Scholar]
- Marshall, L.M. Synaptic localization of alpha-bungarotoxin binding which blocks nicotinic transmission at frog sympathetic neurons. Proc. Natl. Acad. Sci. USA 1981, 78, 1948–1952. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Scientific Name | Biological Endpoint | Media Type | Exposure Concentration of the Active Ingredient (mg L−1) | Observation Duration (Days) | Data Source |
|---|---|---|---|---|---|---|
| Clothianidin | ||||||
| Bluegill sunfish | Lepomis macrochirus | LC50 1; mortality | freshwater | >117 | 4 | EPA #344, 1992 |
| Bluegill sunfish | Lepomis macrochirus | NOEL 2; mortality | freshwater | 117 | 4 | EPA #344, 1992 |
| Fathead minnow | Pimephales promelas | LOEC 3; growth | n.d. | 20 | 33 | EPA #344, 1992 |
| Fathead minnow | Pimephales promelas | NOEL; growth | n.d. | 9.7 | 33 | EPA #344, 1992 |
| Fathead minnow | Pimephales promelas | EC50; intoxication | freshwater | >0.5 | 4 | DePerre et al., 2015 |
| Fathead minnow | Pimephales promelas | LC50; mortality | freshwater | >0.5 | 4 | DePerre et al., 2015 |
| Sheepshead minnow | Cyprinodon variegatus | NOEL; mortality | saltwater | 93.6 | 4 | EPA #344, 1992 |
| Rainbow trout | Oncorhynchus mykiss | LC50; mortality | freshwater | >105.8 | 4 | EPA #344, 1992 |
| Rainbow trout | Oncorhynchus mykiss | NOEL; mortality | freshwater | 105.8 | 4 | EPA #344, 1992 |
| Zebrafish | Danio rerio | multiple | freshwater | 20.0 | 5 | Padilla et al. 2012 |
| Thiamethoxam | ||||||
| Bluegill sunfish | Lepomis macrochirus | LC50; mortality | freshwater | 114 | 4 | EPA #344, 1992 |
| Bluegill sunfish | Lepomis macrochirus | NOEL; mortality | freshwater | 114 | 4 | EPA #344, 1992 |
| Sheepshead minnow | Cyprinodon variegatus | LC50; mortality | salt water | >111 | 4 | EPA #344, 1992 |
| Sheepshead minnow | Cyprinodon variegatus | NOEL; mortality | salt water | 111 | 4 | EPA #344, 1992 |
| Rainbow trout | Oncorhynchus mykiss | LC50; mortality | fresh water | 100 | 4 | EPA #344, 1992 |
| Rainbow trout | Oncorhynchus mykiss | NOEL; mortality | fresh water | 100 | 4 | EPA #344, 1992 |
| Rainbow trout | Oncorhynchus mykiss | LC50; mortality | n.d. | >1005 | 4 | Syngenta SDS |
| Rainbow trout | Oncorhynchus mykiss | LC50; mortality | fresh water | >100 | 4 | EPA #344, 1992 |
| Rainbow trout | Oncorhynchus mykiss | NOEC 4 | fresh water | 20 | 45 | EPA #344, 1992 |
| Rainbow trout | Oncorhynchus mykiss | LOEC | n.d. | >20 | 45 | EPA #344, 1992 |
| Gene (Abbreviation) | Cellular Function | Sequence 5′-3′ | Base Pairs |
|---|---|---|---|
| Reference Genes | |||
| Elongation factor 1-α (ef1-a) b | Delivery of aminoacyl tRNAs to ribosomes | CTG CAC ATA TCG CCT GTA AG | 107 |
| GGC AGC ATC TCC AGA TTT C | |||
| Actin (act) b | Major protein of contractile apparatus | GGC CGT ACA ACT GGT ATT G | 93 |
| CAT GAT GGC ATG AGG TAA GG | |||
| Ornithine decarboxylase (odc) b | Polyamine biosynthesis pathway | GTA CAA GCT GTC TCA GAT GC | 92 |
| GGG AAT CCA CCA CCA ATA TC | |||
| Glyceraldehyde-3-phosphate dehydrogenase (gapdh) b | Carbohydrate metabolism | ATC AAG GCC GCC ATT AAG | 115 |
| CAA AGA TGG AGG AGT GAG TG | |||
| Target Genes | |||
| nAcetylcholine receptor subunit α7 (chrna7) c | Mediating synaptic fast signal transmission | ACC TGA AGT TTG GCT CAT GG | 159 |
| GGT ATG GTT CCT TGC AGC AT | |||
| Muscarinic acetylcholine receptor 4 (chrm4) c | Mediating synaptic fast signal transmission | ATC TTT ATC GCC ACC GTC AC | 204 |
| GAG TGG CCA GTA ACC CTT GA | |||
| Cytochrome p450 (cyp1a1) c | Monooxygenase; detoxification, resistance | AGG AGA AGA GAG TCG ATG | 220 |
| GCT CTG TCT GAT AAT CTA GG | |||
| Acetylcholinesterase (ache) c | Hydrolyses acetylcholine, ends CNS signal | ATC TGA ACT ATA ACC CAC AG | 245 |
| TGT AAT GTT GAG CAG TTT AG |
| Treatment | Tadpoles (n) a | Wet Weight (g) | p Value | Total Length (mm) | p Value | NF Stage | p Value |
|---|---|---|---|---|---|---|---|
| Control | 45.8 (4.3) | 0.73 (0.11) | - | 63.18 (2.50) | - | 56.9 (0.33) | - |
| THX 20 ppm | 69.5 (3.2) | 0.39 (0.03) | 0.022 b | 48.22 (1.22) | <0.001 | 54.2 (0.18) | 0.001 |
| THX 100 ppm | 68.3 (3.9) | 0.52 (0.07) | 0.204 | 52.12 (2.91) | 0.006 | 54.5 (0.59) | 0.002 |
| CLO 20 ppm | 52.0 (3.4) | 0.56 (0.04) | 0.245 | 54.00 (1.56) | 0.013 | 55.3 (0.43) | 0.029 |
| CLO 100 ppm | 44.8 (4.1) | 0.62 (0.10) | 0.334 | 53.79 (1.86) | 0.007 | 55.7 (0.35) | 0.054 |
| Treatment | Difference b in NF (95% Confidence Interval) | |
|---|---|---|
| NF 57; day 44 | Post-metamorphosis (NF 66; day 76) | |
| CLO 20 ppm | 1.8 (1.1, 2.4) c | 1.33 (1.16, 1.50) |
| CLO 100 ppm | 1.5 (0.7, 2.3) | 1.16 (0.97, 1.34) |
| THX 20 ppm | 2.1 (1.5, 2.7) | 1.08 (0.96, 1.20) |
| THX 100 ppm | 1.4 (0.8, 1.9) | 1.12 (0.99, 1.25) |
| Common Name | Scientific Name | Life Stage | Exposure Concentration (ug/L) | Duration (Days) | Mortality | Data Source |
|---|---|---|---|---|---|---|
| Clothianidin | ||||||
| Northern leopard frogs | Rana pipiens | tadpole | 0–100 | 56 | no | Robinson et al., 2021 |
| Northern leopard frogs | Rana pipiens | tadpole | 428 ± 66 | 14 | no | Gavel et al., 2021 |
| Northern leopard frogs | Rana pipiens | tadpole | 2.5 and 250 | on order | no | Robinson et al., 2019 |
| Wood frogs | Lithobates sylvaticus | tadpole | 2.5 and 251 | on order | no | Robinson et al., 2020 |
| Thiamethoxam | ||||||
| Northern leopard frogs | Rana pipiens | tadpole | 0–100 | 56 | no | Robinson et al., 2021 |
| Northern leopard frogs | Rana pipiens | tadpole | 304 ± 49 | 14 | no | Gavel et al., 2021 |
| Northern leopard frogs | Rana pipiens | tadpole | 2.5 and 250 | on order | no | Robinson et al., 2019 |
| Wood frogs | Lithobates sylvaticus | tadpole | 2.5 and 251 | on order | no | Robinson et al., 2020 |
| Wood frogs | Lithobates sylvaticus | tadpole | 1, 10, 100 | 42 | no | Robinson et al., 2017 |
| Wood frogs | Lithobates sylvaticus | tadpole | 200–25,200 | 14 | n.a. | Pochini and Hoverman, 2017 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jenkins, J.A.; Hartop, K.R.; Bukhari, G.; Howton, D.E.; Smalling, K.L.; Mize, S.V.; Hladik, M.L.; Johnson, D.; Draugelis-Dale, R.O.; Brown, B.L. Juvenile African Clawed Frogs (Xenopus laevis) Express Growth, Metamorphosis, Mortality, Gene Expression, and Metabolic Changes When Exposed to Thiamethoxam and Clothianidin. Int. J. Mol. Sci. 2021, 22, 13291. https://doi.org/10.3390/ijms222413291
Jenkins JA, Hartop KR, Bukhari G, Howton DE, Smalling KL, Mize SV, Hladik ML, Johnson D, Draugelis-Dale RO, Brown BL. Juvenile African Clawed Frogs (Xenopus laevis) Express Growth, Metamorphosis, Mortality, Gene Expression, and Metabolic Changes When Exposed to Thiamethoxam and Clothianidin. International Journal of Molecular Sciences. 2021; 22(24):13291. https://doi.org/10.3390/ijms222413291
Chicago/Turabian StyleJenkins, Jill A., Katherine R. Hartop, Ghadeer Bukhari, Debra E. Howton, Kelly L. Smalling, Scott V. Mize, Michelle L. Hladik, Darren Johnson, Rassa O. Draugelis-Dale, and Bonnie L. Brown. 2021. "Juvenile African Clawed Frogs (Xenopus laevis) Express Growth, Metamorphosis, Mortality, Gene Expression, and Metabolic Changes When Exposed to Thiamethoxam and Clothianidin" International Journal of Molecular Sciences 22, no. 24: 13291. https://doi.org/10.3390/ijms222413291
APA StyleJenkins, J. A., Hartop, K. R., Bukhari, G., Howton, D. E., Smalling, K. L., Mize, S. V., Hladik, M. L., Johnson, D., Draugelis-Dale, R. O., & Brown, B. L. (2021). Juvenile African Clawed Frogs (Xenopus laevis) Express Growth, Metamorphosis, Mortality, Gene Expression, and Metabolic Changes When Exposed to Thiamethoxam and Clothianidin. International Journal of Molecular Sciences, 22(24), 13291. https://doi.org/10.3390/ijms222413291