
Transcriptomic Insights on the Preventive Action of Apple (cv Granny Smith) Skin Wounding on Superficial Scald Development

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Apple Superficial Scald Incidence in Relation to Wounding
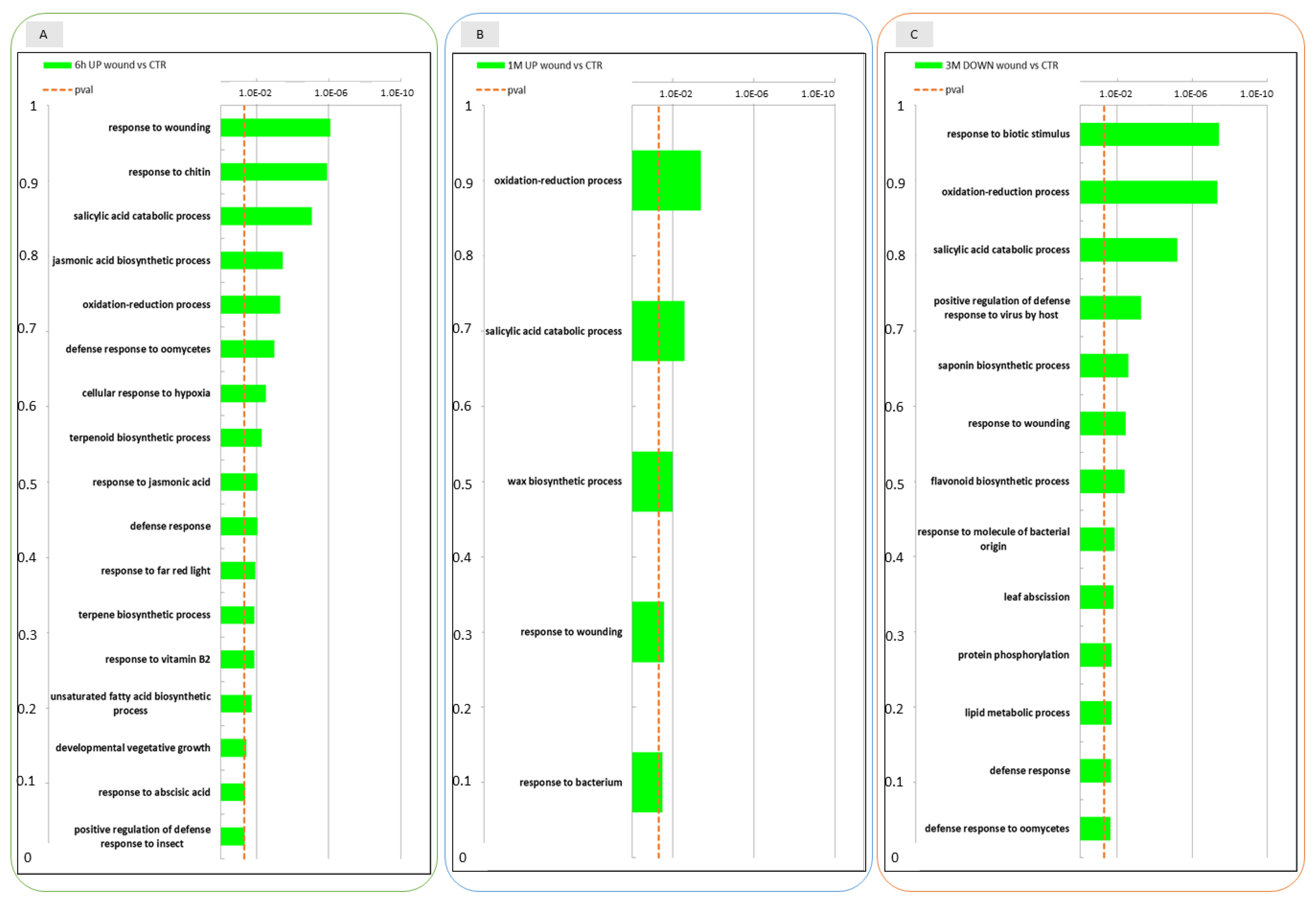
2.2. Comparison of Global Transcriptomic Profiles of Wounded versus Non-Wounded Apple Skins
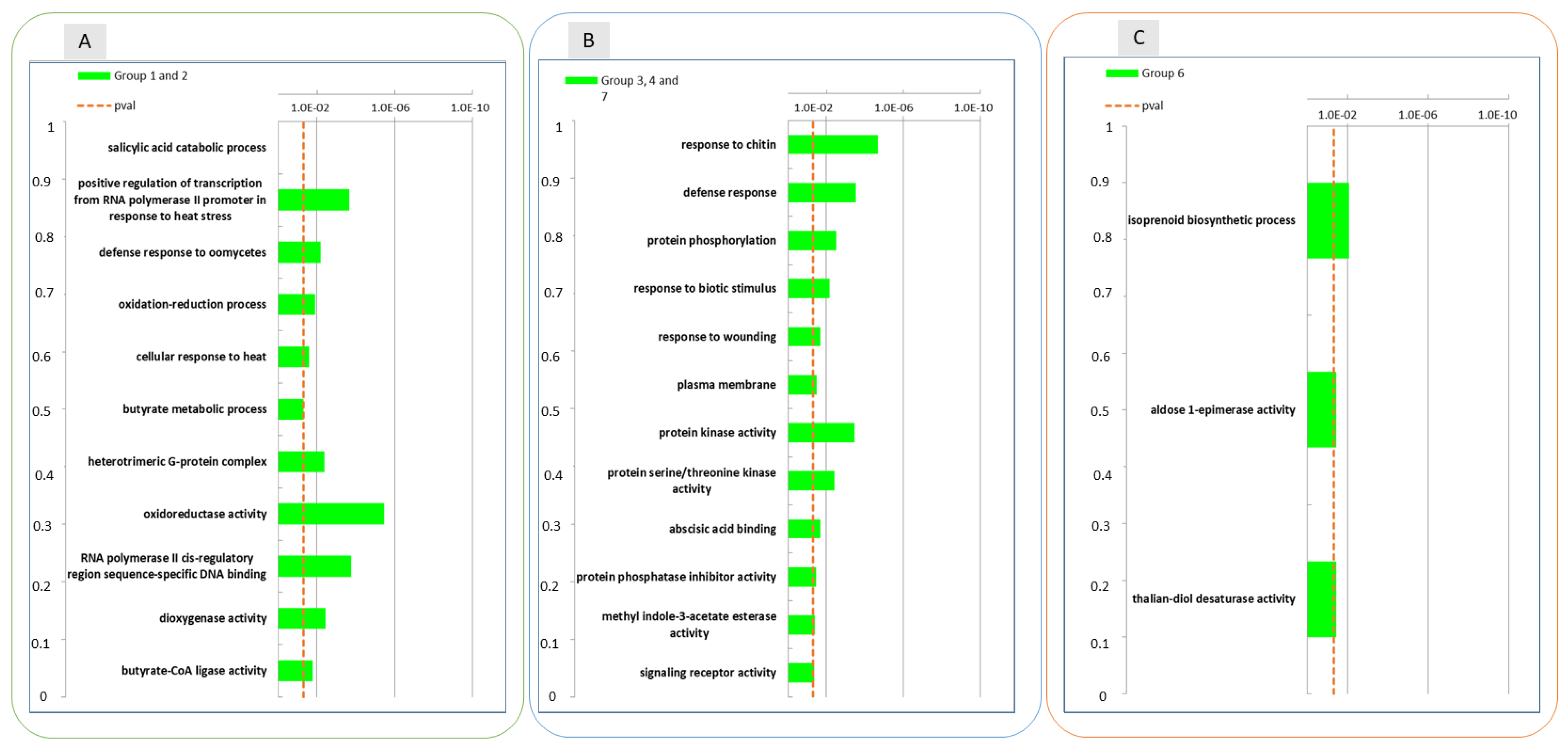
2.3. Conserved Transcriptional Regulation in Wounded and Unwounded Apple Skins during Storage
2.4. Evaluation of the Combined Effect of Wounding and Time of Storage on Transcriptional Dynamics
2.5. Pathway Analysis of Hormonal and Regulatory Genes in Apple Skin Wound Responses
3. Discussion
4. Materials and Methods
4.1. Experimental Setup, Apple Wounding and Storage Conditions
4.2. Evaluation of Scald Incidence and Sampling of Apple Skin Material
4.3. RNA Extraction and Expression Analysis
4.4. RNA-Seq Analysis and Data Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McGlasson, W.B.; Scott, K.J.; Mendoza, D.B., Jr. The refrigerated storage of tropical and subtropical products. Int. J. Refrig. 1979, 22, 199–206. [Google Scholar] [CrossRef]
- Lurie, S.; Watkins, C.B. Superficial scald, its etiology and control. Postharvest Biol. Technol. 2012, 65, 44–60. [Google Scholar] [CrossRef]
- Cainelli, N.; Ruperti, B. Biochemistry and Molecular Biology in Fruits during Cold Storage. Annu. Plant Rev. Online 2019, 2, 659–688. [Google Scholar] [CrossRef]
- Watkins, C.B.; Bramlage, W.J.; Cregoe, B.A. Superficial scald of ‘Granny Smith’ apples is expressed as a typical chilling injury. J. Am. Soc. Hortic. Sci. 1995, 120, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Sabban-Amin, R.; Feygenberga, O.; Belausovb, E.; Pesis, E. Low oxygen and 1-MCP pretreatments delay superficial scald development by reducing reactive oxygen species (ROS) accumulation in stored ‘Granny Smith’ apples. Postharvest Biol. Technol. 2011, 62, 293–304. [Google Scholar] [CrossRef]
- Arquiza, J.M.R.A.; Hay, A.G.; Nock, J.F.; Watkins, C.B. 1-Methylcyclopropene interactions with diphenylamine on diphenylamine degradation, α-farnesene and conjugated trienol concentrations, and polyphenol oxidase and peroxidase activities in apple fruit. J. Agric. Food Chem. 2005, 53, 7565–7570. [Google Scholar] [CrossRef] [PubMed]
- Rupasinghe, H.P.V.; Paliyath, G.; Murr, D.R. Sesquiterpene α-farnesene synthase: Partial purification, characterization, and activity in relation to superficial scald development in apples. J. Am. Soc. Hortic. Sci. 2000, 125, 111–119. [Google Scholar] [CrossRef]
- Busatto, N.; Farneti, B.; Tadiello, A.; Vrhovsek, U.; Cappellin, L.; Biasioli, F.; Velasco, R.; Costa, G.; Costa, F. Target metabolite and gene transcription profiling during the development of superficial scald in apple (Malus × domestica Borkh). BMC Plant Biol. 2014, 14, 193. [Google Scholar] [CrossRef]
- Zermiani, M.; Zonin, E.; Nonis, A.; Begheldo, M.; Ceccato, L.; Vezzaro, A.; Baldan, B.; Trentin, A.; Masi, A.; Pegoraro, M.; et al. Ethylene negatively regulates transcript abundance of ROP-GAP rheostat-encoding genes and affects apoplastic reactive oxygen species homeostasis in epicarps of cold stored apple fruits. J. Exp. Bot. 2015, 66, 7255–7270. [Google Scholar] [CrossRef] [Green Version]
- Karagiannis, E.; Michailidis, M.; Tanou, G.; Samiotaki, M.; Karamanoli, K.; Avramidou, E.; Ganopoulos, I.; Madesis, P.; Molassiotis, A. Ethylene-dependent and -independent superficial scald resistance mechanisms in ‘Granny Smith’ apple fruit. Sci. Rep. 2018, 8, 11436. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, A.Y.; Gil, M.I.; Biasi, W.; Mitcham, E.J. Inhibition of superficial scald in apples by wounding: Changes in lipids and phenolics. Postharvest Biol. Technol. 1997, 12, 203–212. [Google Scholar] [CrossRef]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Daccord, N.; Celton, J.M.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E.; van de Geest, H.; Bianco, L.; Micheletti, D.; Velasco, R.; et al. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 2017, 49, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-Seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeilmaker, T.; Ludwig, N.R.; Elberse, J.; Seidl, M.F.; Berke, L.; Van Doorn, A.; Schuurink, R.C.; Snel, B.; Van den Ackerveken, G. DOWNY MILDEW RESISTANT 6 and DMR6-LIKE OXYGENASE 1 are partially redundant but distinct suppressors of immunity in Arabidopsis. Plant J. 2014, 81, 210–222. [Google Scholar] [CrossRef]
- Caarls, L.; Elberse, J.; Awwanah, M.; Ludwig, N.R.; de Vries, M.; Zeilmaker, T.; Van Wees, S.C.M.; Schuurink, R.C.; Van den Ackerveken, G. Arabidopsis JASMONATE-INDUCED OXYGENASES down-regulate plant immunity by hydroxylation and inactivation of the hormone jasmonic acid. Proc. Natl. Acad. Sci. USA 2017, 114, 6388–6393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poudel, A.N.; Zhang, T.; Kwasniewski, M.; Nakabayashi, R.; Saito, K.; Koo, A.J. Mutations in jasmonoyl-L-isoleucine-12-hydroxylases suppress multiple JA-dependent wound responses in Arabidopsis thaliana. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2016, 1861, 1396–1408. [Google Scholar] [CrossRef]
- Quintero, F.J.; Blatt, M.R. A new family of K+ transporters from Arabidopsis that are conserved across phyla. FEBS Lett. 1999, 415, 206–211. [Google Scholar] [CrossRef]
- Zhao, Y.; Lin, S.; Qiu, Z.; Cao, D.; Wen, J.; Deng, X.; Wang, X.; Lin, J.; Li, X. MicroRNA857 Is Involved in the Regulation of Secondary Growth of Vascular Tissues in Arabidopsis. Plant Physiol. 2015, 169, 2539–2552. [Google Scholar] [CrossRef] [Green Version]
- Bäumler, J.; Riber, W.; Klecker, M.; Müller, L.; Dissmeyer, N.; Weig, A.R.; Mustroph, A. AtERF#111/ABR1 is a transcriptional activator involved in the wounding response. Plant J. 2019, 100, 969–990. [Google Scholar] [CrossRef] [Green Version]
- Lara, A.; Ródenas, R.; Andrés, Z.; Martínez, V.; Quintero, F.J.; Nieves-Cordones, M.; Botella, M.A.; Rubio, F. AtHAK5-mediated root high-affinity K+ uptake is regulated by the protein kinases AtCIPK1 and AtCIPK9. J. Exp. Bot. 2020, 71, 5053–5060. [Google Scholar] [CrossRef]
- Cooper, L.; Walls, R.L.; Elser, J.; Gandolfo, M.A.; Stevenson, D.W.; Smith, B.; Preece, J.; Athreya, B.; Mungall, C.J.; Rensing, S.; et al. The Plant Ontology as A Tool for Comparative Plant Anatomy and Genomic Analyses. Plant Cell Physiol. 2012, 54, e1. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Arizono, T.; Sato, R.; Trinh, M.D.L.; Hashimoto, A.; Kono, M.; Tsujii, M.; Uozumi, N.; Takaichi, S.; Masuda, S. DAY-LENGTH-DEPENDENT DELAYED-GREENING1, the Arabidopsis Homolog of the Cyanobacterial H+-Extrusion Protein, is Essential for Chloroplast pH Regulation and Optimization of Non-Photochemical Quenching. Plant Cell Physiol. 2019, 60, 2660–2671. [Google Scholar] [CrossRef] [PubMed]
- Brenner, W.G.; Leuendorf, J.E.; Cortleven, A.; Martin, L.B.B.; Schaller, H.; Schmülling, T. Analysis of CFB, a cytokinin-responsive gene of Arabidopsis thaliana encoding a novel F-box protein regulating sterol biosynthesis. J. Exp. Bot. 2017, 68, 2769–2785. [Google Scholar] [CrossRef] [Green Version]
- Mazarei, M.; Lennon, K.A.; Puthoff, D.P.; Rodermel, S.R.; Baum, T.J. Homologous soybean and Arabidopsis genes share responsiveness to cyst nematode infection. Mol. Plant Pathol. 2004, 5, 409–423. [Google Scholar] [CrossRef]
- Mansfield, T.A.; Schultes, N.P.; Mourad, G.S. AtAzg1 and AtAzg2 comprise a novel family of purine transporters in Arabidopsis. FEBS Lett. 2009, 583, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Tholl, D.; Sohrabi, R.; Huh, J.H.; Lee, S. The biochemistry of homoterpenes—Common constituents of floral and herbivore-induced plant volatile bouquets. Phytochemistry 2011, 72, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhao, F.; Chen, L.; Pan, Y.; Sun, L.; Bao, N.; Zhang, T.; Cui, C.X.; Qiu, Z.; Zhang, Y.; et al. Jasmonate-mediated wound signalling promotes plant regeneration. Nat. Plants 2019, 5, 491–497. [Google Scholar] [CrossRef]
- Coello, P.; Sassen, A.; Haywood, V.; Davis, K.R.; Walker, J.C. Biochemical characterization and expression of RLK4, a receptor-like kinase from Arabidopsis thaliana. Plant Sci. 1999, 143, 83. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, Z.; Ma, L.; Liao, C. The Ubiquitin E3 Ligase RHA2b Promotes Degradation of MYB30 in Abscisic Acid Signaling. Plant Physiol. 2018, 178, 428–440. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Wang, Y.; Zheng, H.; Lu, W.; Wu, C.; Huang, J.; Yan, K.; Yang, G.; Zheng, C. Salt-induced transcription factor MYB74 is regulated by the RNA-directed DNA methylation pathway in Arabidopsis. J. Exp. Bot. 2015, 66, 5997–6008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagor, G.H.; Zhang, S.; Kojima, S.; Simm, S.; Berberich, T.; Kusan, T. Reducing Cytoplasmic Polyamine Oxidase Activity in Arabidopsis Increases Salt and Drought Tolerance by Reducing Reactive Oxygen Species Production and Increasing Defense Gene Expression. Front. Plant Sci. 2016, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Mendy, B.; Wang’ombe, M.W.; Radakovic, Z.S.; Holbein, J.; Ilyas, M.; Chopra, D.; Holton, N.; Zipfel, C.; Grundler, F.M.; Siddique, S. Arabidopsis leucine-rich repeat receptor-like kinase NILR1 is required for induction of innate immunity to parasitic nematodes. PLoS Pathog. 2017, 13, e1006284. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, K.K.; Atamian, H.S.; Kaloshian, I.; Eulgem, T. WRKY72-type transcription factors contribute to basal immunity in tomato and Arabidopsis as well as gene-for-gene resistance mediated by the tomato R-gene Mi-1. Plant J. 2010, 63, 229–240. [Google Scholar] [CrossRef]
- González-Lamothe, R.; Tsitsigiannis, D.I.; Ludwig, A.A.; Panicot, M.; Shirasu, K.; Jones, J.D.G. The U-Box Protein CMPG1 Is Required for Efficient Activation of Defense Mechanisms Triggered by Multiple Resistance Genes in Tobacco and Tomato. Plant Cell 2006, 18, 1067–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Østergaard, L.; Lauvergeat, V.; Naested, H.; Mattsson, O.; Mundy, J. Two differentially regulated Arabidopsis genes define a new branch of the DFR superfamily. Plant Sci. 2001, 160, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Gratkowska-Zmuda, D.M.; Kubala, S.; Sarnowska, E.; Cwiek, P.; Oksinska, P.; Steciuk, J.; Rolicka, A.T.; Zaborowska, M.; Bucior, E.; Maassen, A.; et al. The SWI/SNF ATP-Dependent Chromatin Remodeling Complex in Arabidopsis Responds to Environmental Changes in Temperature-Dependent Manner. Int. J. Mol. Sci. 2020, 21, 762. [Google Scholar] [CrossRef] [Green Version]
- Kurdyukov, S.; Faust, A.; Trenkamp, S.; Bär, S.; Franke, R.; Efremova, N.; Tietjen, K.; Schreiber, L.; Saedler, H.; Yephremov, A. Genetic and biochemical evidence for involvement of HOTHEAD in the biosynthesis of long-chain alpha-,omega-dicarboxylic fatty acids and formation of extracellular matrix. Planta 2006, 224, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.N.; Li, X.T.; Yuan, J.Z.; Wang, F.Z.; Wang, S.M.; Ma, Q. Nitrate transporter NPF7.3/NRT1.5 plays an essential role in regulating phosphate deficiency responses in Arabidopsis. Biochem. Biophys. Res. Commun. 2018, 508, 314–319. [Google Scholar] [CrossRef]
- Charron, J.B.; Ouellet, F.; Houde, M.; Sarhan, F. The plant Apolipoprotein D ortholog protects Arabidopsis against oxidative stress. BMC Plant Biol. 2008, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Dhatterwal, P.; Basu, S.; Mehrotra, S.; Mehrotra, R. Genome wide analysis of W-box element in Arabidopsis thaliana reveals TGAC motif with genes down regulated by heat and salinity. Sci. Rep. 2019, 9, 1681. [Google Scholar] [CrossRef] [PubMed]
- Ascencio-Ibáñez, J.T.; Sozzani, R.; Lee, T.J.; Chu, T.M.; Wolfinger, R.D.; Cella, R.; Hanley-Bowdoin, L. Global Analysis of Arabidopsis Gene Expression Uncovers a Complex Array of Changes Impacting Pathogen Response and Cell Cycle during Geminivirus Infection. Plant Physiol. 2008, 148, 436–454. [Google Scholar] [CrossRef] [Green Version]
- Wei, P.C.; Chen, S.; Zhang, X.Q.; Zhao, P.; Xiong, Y.M.; Wang, W.L.; Chen, J.; Wang, X. An α-expansin, VfEXPA1, is involved in regulation of stomatal movement in Vicia faba L. Chin. Sci. Bull. 2011, 56, 3531–3537. [Google Scholar] [CrossRef] [Green Version]
- Keadtidumrongkul, P.; Suttangkakul, A.; Pinmanee, P.; Pattana, K.; Kittiwongwattana, C.; Apisitwanich, S.; Vuttipongchaikij, S. Growth modulation effects of CBM2a under the control of AtEXP4 and CaMV35S promoters in Arabidopsis thaliana, Nicotiana tabacum and Eucalyptus camaldulensis. Transgenic Res. 2017, 26, 447–463. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Chen, Q.; Xin, P.; Yuan, J.; Ma, Y.; Wang, X.; Xu, M.; Chu, J.; Peters, R.J.; Wang, G. CYP72A enzymes catalyse 13-hydrolyzation of gibberellins. Nat. Plants 2019, 5, 1057–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.H.; Ma, X.M.; Kojima, M.; Sakakibara, H.; Wang, Y.W.; Hou, B.K. Overexpression of glucosyltransferase UGT85A1 influences trans-zeatin homeostasis and trans-zeatin responses likely through O-glucosylation. Planta 2012, 237, 991–999. [Google Scholar] [CrossRef]
- Del Corpo, D.; Fullone, M.R.; Miele, R.; Lafond, M.; Pontiggia, D.; Grisel, S.; Kieffer-Jaquinod, S.; Giardina, T.; Bellincampi, D.; Lionetti, V. AtPME17 is a functional Arabidopsis thaliana pectin methylesterase regulated by its PRO region that triggers PME activity in the resistance to Botrytis cinerea. Mol. Plant Pathol. 2020, 21, 1620–1633. [Google Scholar] [CrossRef] [PubMed]
- Smalle, J.; Haegman, M.; Mertens, J.; Vangronsveld, J.; van Montagu, M.; van der Straeten, D. The expression pattern of the Arabidopsis ACC synthase gene 1 during rosette leaf development. J. Exp. Bot. 1999, 50, 1561. [Google Scholar] [CrossRef]
- Yamamoto, A.; Bhuiyan, M.N.; Waditee, R.; Tanaka, Y.; Esaka, M.; Oba, K.; Jagendorf, A.T.; Takabe, T. Suppressed expression of the apoplastic ascorbate oxidase gene increases salt tolerance in tobacco and Arabidopsis plants. J. Exp. Bot. 2005, 56, 1785. [Google Scholar] [CrossRef] [Green Version]
- Ji, H.; Wang, Y.; Cloix, C.; Li, K.; Jenkins, G.I.; Wang, S.; Shang, Z.; Shi, Y.; Yang, S.; Li, X. The Arabidopsis RCC1 Family Protein TCF1 Regulates Freezing Tolerance and Cold Acclimation through Modulating Lignin Biosynthesis. PLoS Genet. 2015, 11, e1005471. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Ikeda, A.; Shiojiri, K.; Ozawa, R.; Shiki, K.; Kunihiro, N.N.; Fujita, K.; Sugimoto, K.; Yamato, K.T.; Dohra, H.; et al. Identification of a hexenal reductase that modulates the composition of green leaf volatiles. Plant Physiol. 2018, 178, 552–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, R.; Kumar, D.; Sultana, A.; Bhattacharyya, D.; Chattopadhyay, S. Glutathione regulates ACC synthase transcription via WRKY33 and ACC oxidase by modulating mRNA stability to induce ethylene synthesis during stress. Plant Physiol. 2015, 169, 2963–2981. [Google Scholar] [CrossRef]
- Yang, W.Y.; Zheng, Y.; Bahn, S.C.; Pan, X.Q.; Li, M.Y.; Vu, H.S.; Roth, M.R.; Scheu, B.; Welti, R.; Hong, Y.Y.; et al. The Patatin-Containing Phospholipase A pPLAII alpha Modulates Oxylipin Formation and Water Loss in Arabidopsis thaliana. Mol. Plant 2012, 5, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Han, Y.; Zhu, X.; Shah, A.; Wang, W.; Sheng, Y.; Fan, T.; Cao, S. Negative regulation of cadmium tolerance in Arabidopsis by MMDH2. Plant Mol. Biol. 2019, 101, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Kong, X.; Li, C.; Liu, Y.; Ding, Z. Potassium Retention under Salt Stress Is Associated with Natural Variation in Salinity Tolerance among Arabidopsis Accessions. PLoS ONE 2015, 10, e0124032. [Google Scholar] [CrossRef]
- Taki, N.; Sasaki-Sekimoto, Y.; Obayashi, T.; Kikuta, A.; Kobayashi, K.; Ainai, T.; Yagi, K.; Sakurai, N.; Suzuki, H.; Masuda, T.; et al. 12-Oxo-Phytodienoic Acid Triggers Expression of a Distinct Set of Genes and Plays a Role in Wound-Induced Gene Expression in Arabidopsis. Plant Physiol. 2005, 139, 1268–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triantaphylides, C.; Krischke, M.; Hoeberichts, F.A.; Ksas, B.; Gresser, G.; Havaux, M.; Van Breusegem, F.; Mueller, M. Singlet oxygen is the major reactive oxygen species involved in photo-oxidative damage to plants. Plant Physiol. 2008, 148, 960–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, C.; Jiang, M.; Li, N.; Cao, J.; Hou, M.; Ni, D.A.; Chu, Z. Integrated bioinformatics analysis of As, Au, Cd, Pb and Cu heavy metal responsive marker genes through Arabidopsis thaliana GEO datasets. PeerJ 2019, 18, e6495. [Google Scholar] [CrossRef] [Green Version]
- Stanley Kim, H.; Yu, Y.; Snesrud, E.C.; Moy, L.P.; Linford, L.D.; Haas, B.J.; Nierman, W.C.; Quackenbush, J. Transcriptional divergence of the duplicated oxidative stress-responsive genes in the Arabidopsis genome. Plant J. 2005, 41, 212. [Google Scholar] [CrossRef]
- Hou, Y.; Guo, X.; Cyprys, P.; Zhang, Y.; Bleckmann, A.; Cai, L.; Huang, Q.; Luo, Y.; Gu, H.; Dresselhaus, T.; et al. Maternal ENODLs Are Required for Pollen Tube Reception in Arabidopsis. Curr. Biol. 2016, 26, 2343–2350. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Langlois-Meurinne, M.; Didierlaurent, L.; Chaouch, S.; Bellvert, F.; Massoud, K.; Garmier, M.; Thareau, V.; Comte, G.; Noctor, G.; et al. The secondary metabolism glycosyltransferases UGT73B3 and UGT73B5 are components of redox status in resistance of Arabidopsis to Pseudomonas syringae pv. Tomato. Plant Cell Environ. 2013, 37, 1114–1129. [Google Scholar] [CrossRef] [PubMed]
- Galva, C.; Virgin, G.K.; Helms, J.B.; Gatto, C. ATP protects against FITC labeling of Solanum lycopersicon and Arabidopsis thaliana Ca2+-ATPase ATP binding domains. Plant Physiol. Biochem. 2013, 71, 261–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Abreu-Neto, J.B.; Turchetto-Zolet, A.C.; de Oliveira, L.F.; Zanettini, M.H.; Margis-Pinheiro, M. Heavy metal-associated isoprenylated plant protein (HIPP): Characterization of a family of proteins exclusive to plants. FEBS J. 2013, 280, 1604–1616. [Google Scholar] [CrossRef] [PubMed]
- Provart, N.J.; Gil, P.; Chen, W.; Han, B.; Chang, H.S.; Wang, X.; Zhu, T. Gene expression phenotypes of Arabidopsis associated with sensitivity to low temperatures. Plant Physiol. 2003, 132, 893. [Google Scholar] [CrossRef] [Green Version]
- Zientara, K.; Wawrzyska, A.; Lukomska, J.; Moya, J.R.L.; Liszewska, F.; Assunção, A.G.L.; Aarts, M.G.M.; Sirko, A. Activity of the AtMRP3 promoter in transgenic Arabidopsis thaliana and Nicotiana tabacum plants is increased by cadmium, nickel, arsenic, cobalt and lead but not by zinc and iron. J. Biotechnol. 2008, 139, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Zhao, P.; Liang, W.; Cheng, Q.; Mu, B.; Niu, F.; Yan, J.; Chunlin, L.; Xie, H.; Kav, N.N.V.; et al. A rapeseed WRKY transcription factor phosphorylated by CPK modulates cell death and leaf senescence by regulating the expression of ROS and SA-synthesis-related genes. Agric. Food Chem. 2020, 68, 7348–7359. [Google Scholar] [CrossRef]
- Rudell, D.R.; Buchanan, D.A.; Leisso, R.S.; Whitaer, B.D.; Mattheis, J.P.; Zhu, Y.; Veranasi, V. Ripening, storage temperature, ethylene action, and oxidative stress alter apple peel phytosterol metabolism. Phytochemistry 2011, 72, 1328–1340. [Google Scholar] [CrossRef]
- Rudell, D.R.; Mattheis, J.P.; Hertog, M.L.A.T.M. Metabolomic Change Precedes Apple Superficial Scald Symptoms. J. Agric. Food Chem. 2009, 57, 8459–8466. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant Sci. 2015, 20, 219–229. [Google Scholar] [CrossRef]
- Hu, Y.; Jiang, Y.; Han, X.; Wang, H.; Pan, J.; Yu, D. Jasmonate regulates leaf senescence and tolerance to cold stress: Crosstalk with other phytohormones. J. Exp. Bot. 2017, 68, 1361–1369. [Google Scholar] [CrossRef]
- Fenn, M.A.; Giovannoni, J.J. Phytohormones in fruit development and maturation. J. Plant 2021, 105, 446–458. [Google Scholar] [CrossRef]
- Wang, S.Y.; Shi, X.C.; Liu, F.Q.; Laborda, P. Effects of exogenous methyl jasmonate on quality and preservation of postharvest fruits: A review. Food Chem. 2021, 353, 129482. [Google Scholar] [CrossRef]
- Hu, Y.; Jiang, L.; Wang, F.; Yu, D. Jasmonate Regulates the INDUCER OF CBF EXPRESSION–C-REPEAT BINDING FACTOR/DRE BINDING FACTOR1 Cascade and Freezing Tolerance in Arabidopsis. Plant Cell 2013, 25, 2907–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Song, C.; Brummell, D.A.; Qi, S.; Lin, Q.; Duan, Y. Jasmonic acid treatment alleviates chilling injury in peach fruit by promoting sugar and ethylene metabolism. Food Chem. 2021, 338, 128005. [Google Scholar] [CrossRef] [PubMed]
- Siboza, X.I.; Bertling, I.; Odindo, A.O. Salicylic acid and methyl jasmonate improve chilling tolerance in cold-stored lemon fruit (Citrus limon). J. Plant Physiol. 2014, 171, 1722–1731. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Zheng, Y.; Tang, S.; Rui, H.; Wang, C.Y. A combination of hot air and methyl jasmonate vapor treatment alleviates chilling injury of peach fruit. Postharvest Biol. Technol. 2009, 52, 24–29. [Google Scholar] [CrossRef]
- Cao, S.; Zheng, Y.; Wang, K.; Jin, P.; Rui, H. Methyl jasmonate reduces chilling injury and enhances antioxidant enzyme activity in postharvest loquat fruit. Food Chem. 2009, 115, 1458–1463. [Google Scholar] [CrossRef]
- González-Aguilar, G.A.; Tiznado-Hernández, M.E.; Zavaleta-Gatica, R.; Martínez-Téllez, M.A. Methyl jasmonate treatments reduce chilling injury and activate the defense response of guava fruits. Biochem. Biophys. Res. Commun. 2004, 313, 694–701. [Google Scholar] [CrossRef]
- Nonis, A.; Vezzaro, A.; Ruperti, B. Evaluation of RNA Extraction Methods and Identification of Putative Reference Genes for Real-Time Quantitative Polymerase Chain Reaction Expression Studies on Olive (Olea europaea L.) Fruits. J. Agric. Food Chem. 2012, 60, 6855–6865. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, D.M.; Johnson, K.; DiTommaso, T.; Tickle, T.; Couger, M.B.; Payzin-Dogru, D.; Lee, T.J.; Leigh, N.D.; Kuo, T.H.; Davis, F.G.; et al. A Tissue-Mapped Axolotl De Novo Transcriptome Enables Identification of Limb Regeneration Factors. Cell Rep. 2017, 18, 762–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DEGs log2FC > 3.5–6 h | |||||
| Description | GGDH 13 v1.1 | Arabidopsis Homologue | log2FC | Function/Regulatory Process | References |
| 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | MD14G1140400 | AT4G10490.1 | 9.1359 | defense response to oomycetes, salicylic acid catabolic process | [15] |
| 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | MD04G1154400 | AT3G11180.1 | 8.8233 | regulation of jasmonic acid mediated signaling pathway | [16] |
| 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | MD14G1141200 | AT4G10490.1 | 8.6543 | defense response to oomycetes, salicylic acid catabolic process | [15] |
| cytochrome P450%2C family 94%2C subfamily C%2C polypeptide 1 | MD03G1140700 | AT2G27690.1 | 7.0295 | response to wounding | [17] |
| 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | MD14G1141000 | AT4G10490.1 | 6.8804 | defense response to oomycetes, salicylic acid catabolic process | [15] |
| potassium transporter 1 | MD05G1223100 | AT2G30070.1 | 5.8179 | potassium ion transmembrane transport | [18] |
| laccase 7 | MD12G1157100 | AT3G09220.1 | 5.0617 | member of laccase family of genes (17 members in Arabidopsis) | [19] |
| Integrase-type DNA-binding superfamily protein | MD10G1032000 | AT5G64750.1 | 4.9703 | wounding stress response | [20] |
| UDP-glucosyl transferase 85A2 | MD08G1185700 | AT1G22360.1 | 4.7899 | glucuronosyltransferase activity | |
| Phosphorylase superfamily protein | MD15G1191300 | AT4G28940.1 | 4.7704 | nucleoside metabolic process | |
| hypothetical protein | MD06G1215300 | AT1G13360.1 | 4.7629 | cellular response to hypoxia | |
| high affinity K+ transporter 5 | MD11G1303100 | AT4G13420.1 | 4.5891 | potassium ion transport | [21] |
| terpene synthase 14 | MD10G1309900 | AT1G61680.1 | 4.5332 | terpene synthase | [22] |
| Uncharacterized protein family (UPF0114) | MD00G1038700 | AT4G19390.1 | 4.4614 | Na+ efflux activity | [23] |
| F-box family protein | MD11G1169800 | AT2G27310.1 | 4.4381 | converts 2,3-oxidosqualene to cycloartenol in the sterol biosynthesis pathway | [24] |
| RING/U-box superfamily protein | MD09G1240700 | AT2G42360.1 | 4.3325 | protein ubiquitination | |
| basic helix-loop-helix (bHLH) DNA-binding superfamily protein | MD15G1305200 | AT4G37850.1 | 4.3299 | regulation of transcription | |
| plastid movement impaired protein | MD16G1029200 | AT2G01340.1 | 4.2995 | response to nematode | [25] |
| AZA-guanine resistant1 | MD07G1231600 | AT3G10960.1 | 4.2822 | adenine transport, guanine transport | [26] |
| cytochrome P450%2C family 82%2C subfamily G%2C polypeptide 1 | MD10G1158100 | AT3G25180.1 | 4.0730 | defense response to insect wounding | [27] |
| anthranilate synthase alpha subunit 1 | MD07G1216200 | AT5G05730.1 | 4.0691 | response to wounding | [28] |
| Protein kinase superfamily protein | MD15G1076300 | AT4G00340.1 | 4.0232 | Ser/Thr receptor-like protein kinase | [29] |
| RING-H2 finger protein 2B | MD13G1051400 | AT2G01150.1 | 3.8958 | ABA response to abiotic stress/positive regulation of abscisic acid-activated signaling pathway | [30] |
| myb domain protein 74 | MD05G1327500 | AT4G05100.1 | 3.8391 | response to salt stress | [31] |
| MLP-like protein 423 | MD13G1161700 | AT1G24020.2 | 3.8376 | plant development/defence response | |
| polyamine oxidase 1 | MD02G1306200 | AT5G13700.1 | 3.7689 | reducing Reactive Oxygen Species Production and Increasing Defense Gene Expression | [32] |
| Leucine-rich repeat protein kinase family protein | MD06G1198600 | AT1G74360.1 | 3.6803 | defense response to nematode | [33] |
| Calcium-binding EF-hand family protein | MD03G1144700 | AT2G27480.1 | 3.6779 | calcium-binding EF-hand family protein | |
| WRKY DNA-binding protein 72 | MD06G1189100 | AT5G15130.1 | 3.6274 | transcriptional reprogramming associated with plant immune responses | [34] |
| ARM repeat superfamily protein | MD06G1232800 | AT5G37490.1 | 3.5579 | plant-type hypersensitive response | [35] |
| NAD(P)-binding Rossmann-fold superfamily protein | MD06G1003700 | AT2G33590.1 | 3.5458 | response to ABA | [36] |
| myb domain protein 15 | MD05G1197600 | AT3G23250.1 | 3.5358 | response to cold | [37] |
| cytochrome P450%2C family 94%2C subfamily C%2C polypeptide 1 | MD11G1171100 | AT2G27690.1 | 3.5234 | response to wound | [17] |
| cytochrome P450%2C family 71%2C subfamily B%2C polypeptide 37 | MD16G1103600 | AT3G26330.1 | 3.5031 | oxidoreductase activity | |
| DEGs log2FC > 3.5–1 month | |||||
| Description | GGDH 13 v1.1 | Arabidopsis Homologue | log2FC | Function/Regulatory Process | References |
| Glucose-methanol-choline (GMC) oxidoreductase family protein | MD03G1090900 | AT1G73050.1 | 7.2112 | drought resistance | [38] |
| 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | MD11G1074400 | AT1G52800.1 | 7.1643 | hormonal development regulation | |
| Glucose-methanol-choline (GMC) oxidoreductase family protein | MD00G1012900 | AT1G73050.1 | 5.6504 | drought resistance | [38] |
| RmlC-like cupins superfamily protein | MD10G1022900 | AT5G38940.1 | 5.0327 | response to abscisic acid (ROP10) | |
| ethylene response factor 1 | MD16G1216900 | AT3G23240.1 | 4.7101 | ethylene-activated defence response to abiotic factors | |
| 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | MD03G1140400 | AT4G10490.1 | 4.6868 | defense response to oomycetes, response to salicylic acid | [15] |
| nitrate transporter 1.5 | MD02G1228800 | AT1G32450.1 | 4.4253 | response to nitrate/nitrate transport | [39] |
| Pathogenesis-related thaumatin superfamily protein | MD17G1250000 | AT1G20030.2 | 4.4096 | oxidative stress response | [40] |
| Eukaryotic aspartyl protease family protein | MD11G1287900 | AT1G03220.1 | 4.4009 | response to abiotic stress conditions | [41] |
| Pathogenesis-related thaumatin superfamily protein | MD17G1249600 | AT1G20030.2 | 4.0734 | oxidative stress response | [40] |
| Late embryogenesis abundant (LEA) hydroxyproline-rich glycoprotein family | MD06G1094100 | AT2G46150.1 | 3.944 | pathogen response | [42] |
| expansin A1 | MD06G1195100 | AT1G69530.2 | 3.9284 | regulation of stomatal opening | [43] |
| homolog of carrot EP3-3 chitinase | MD04G1047700 | AT3G54420.1 | 3.8788 | response to environmental stresses (cold, wounding, dehydration) | |
| expansin A4 | MD01G1135600 | AT2G39700.1 | 3.8372 | growth and development modulation | [44] |
| cytochrome P450%2C family 72%2C subfamily A%2C polypeptide 9 | MD16G1056600 | AT3G14630.1 | 3.6968 | hydrolyzation of gibberellins | [45] |
| DEGs log2FC < 3.5–3 months | |||||
| Description | GGDH 13 v1.1 | Arabidopsis Homologue | log2FC | Function/Regulatory Process | References |
| UDP-Glycosyltransferase superfamily protein | MD17G1125800 | AT1G22400.1 | −5.0363 | plant growth and development | [46] |
| Plant invertase/pectin methylesterase inhibitor superfamily | MD02G1207900 | AT2G45220.1 | −4.8852 | pathogen-induced pectin methylesterases activity and resistance against B. cinerea by triggering jasmonic acid–ethylene-dependent PDF1.2 (AT5G44420) expression | [47] |
| ACC synthase 1 | MD01G1070400 | AT3G61510.1 | −4.4324 | ethylene biosynthetic process | [48] |
| Cupredoxin superfamily protein | MD15G1328900 | AT4G39830.1 | −4.2877 | salt-stress tolerance | [49] |
| NC domain-containing protein-like protein | MD01G1232800 | AT5G16360.1 | −4.1931 | unknown | |
| NAD(P)-linked oxidoreductase superfamily protein | MD12G1240300 | AT1G60690.1 | −4.1175 | unknown | |
| blue-copper-binding protein | MD02G1028300 | AT5G20230.1 | −4.1059 | response to cold/freezing | [50] |
| cinnamyl alcohol dehydrogenase 7 | MD01G1042500 | AT4G37980.1 | −4.1012 | direct plant defense at wound sites | [51] |
| 1-aminocyclopropane-1-carboxylic acid (acc) synthase 6 | MD14G1111500 | AT4G11280.1 | −4.0773 | environmental stress responses | [52] |
| phospholipase A 2A | MD15G1085800 | AT2G26560.1 | −4.0524 | response to dehydration | [53] |
| ABC-2 and Plant PDR ABC-type transporter family protein | MD09G1204300 | AT1G59870.1 | −3.9073 | ccadmium stress tolerance | [54] |
| Late embryogenesis abundant (LEA) hydroxyproline-rich glycoprotein family | MD07G1281600 | AT2G46150.1 | −3.8798 | pathogen response | [42] |
| potassium transporter 1 | MD05G1223100 | AT2G30070.1 | −3.8791 | response to salt stress | [55] |
| Late embryogenesis abundant (LEA) hydroxyproline-rich glycoprotein family | MD14G1114000 | AT2G46150.1 | −3.8079 | pathogen response | [42] |
| NAD(P)-linked oxidoreductase superfamily protein | MD08G1240600 | AT1G60750.1 | −3.8033 | response to wounding | [56] |
| S-adenosyl-L-methionine-dependent methyltransferases superfamily protein | MD08G1242800 | AT5G10830.1 | −3.7399 | ROS-induced cell death | [57] |
| cytochrome P450%2C family 81%2C subfamily D%2C polypeptide 8 | MD03G1281500 | AT4G37370.1 | −3.6903 | response to heavy metal | [58] |
| FAD-binding Berberine family protein | MD10G1244300 | AT4G20820.1 | −3.685 | oxidative stress response | [59] |
| early nodulin-like protein 14 | MD02G1028800 | AT2G25060.1 | −3.6798 | male-female communication and fertilization | [60] |
| UDP-glucosyl transferase 73B3 | MD07G1007400 | AT4G34131.1 | −3.6701 | regulation of redox status and general detoxification of ROS-reactive secondary metabolites | [61] |
| SAUR-like auxin-responsive protein family | MD07G1117400 | AT3G60690.1 | −3.6455 | Auxin-responsive protein SAUR like | |
| calcium ATPase 2 | MD02G1185000 | AT4G37640.1 | −3.6231 | Ca2+ transport against concentration gradients using ATP | [62] |
| Heavy metal transport/detoxification superfamily protein | MD10G1280500 | AT1G06330.1 | −3.5779 | response to heavy metal | [63] |
| cellulose synthase like G1 | MD02G1095700 | AT4G24010.1 | −3.5496 | response to cold | [64] |
| multidrug resistance-associated protein 3 | MD16G1109500 | AT3G13080.1 | −3.5382 | response to heavy metal | [65] |
| blue-copper-binding protein | MD02G1028600 | AT5G20230.1 | −3.5213 | ROS and SA-synthesis-related genes expression regulation | [66] |
| Mean Starch Content | Mean Fresh Weight | Sugars Brix | Firmness kg/cm2 | Acidity g/L mal.ac | |
|---|---|---|---|---|---|
| vintage 2019 | 2.1 | 212.2 | 9.7 | 8.4 | 9 |
| vintage 2020 | 2.0 | 189 | 10 | 8.1 | 7.9 |
| Gene Bank Accession Number | Name | Primer fw | Primer rv | References |
|---|---|---|---|---|
| AB030859 | MdACO | CAGTCGGATGGGACCAGAA | GCTTGGAATTTCAGGCCAGA | Dal Cin et al., 2000 |
| L29450 | MdPPO | CTGACTCGGACTGGTTGGAC | CTTCGCTACTTTGCTCAATGC | this work |
| MD02G1028600 | BCB | TCCTCCAGCTGGCTCTGTTGC | GCTACTCTGTGTTCCCCGCTCC | this work |
| MD04G1047700 | EP3 | TGGCCTCCGATCCCGTCCTT | AGGGCAGGCTGTTTACCATCACA | this work |
| MD11G1074400 | 2OG | CCTCACACGGACAAGAATTTCACA | GCAGCCTCAATCCACTCGCC | this work |
| MD10G1299400 | unknown1 | CCACTGCCCGCGATTTACG | ACCAGAGTCGGGGGCTGATG | this work |
| MD11G1072300 | FAD7 | AGCAGCAGAGCAAGCCCATCAA | GGAGGGGCACTTGGGTCGAA | this work |
| MD13G1161200 | unknown2 | GAGCACCAGCCACTATCACACCAAG | GGTGGGAGGCCTTCTCTTTGC | this work |
| MD13G1161000 | unknown3 | TGGTGGCTTCCGGCAATGGT | GGTGGGAGGCCTTCTCTTTGC | this work |
| MD13G1160900 | MLP423 | CAGTCTGCGCTCCTGCTAGGTTG | TTCCAACACCTCCATCTCCCTCA | this work |
| MD15G1411600 | LHCB6 | GGCTCGATGGCTCGCTTCC | ACCACTTGAGGAACGCCGGG | this work |
| MD04G1151300 | LHCB4.2 | CCCCGTCGCACCTTAGCACA | GTAGGCGTCGGTCGGAAGGC | this work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cainelli, N.; Forestan, C.; Angeli, D.; Villegas, T.R.; Costa, F.; Botton, A.; Rasori, A.; Bonghi, C.; Ruperti, B. Transcriptomic Insights on the Preventive Action of Apple (cv Granny Smith) Skin Wounding on Superficial Scald Development. Int. J. Mol. Sci. 2021, 22, 13425. https://doi.org/10.3390/ijms222413425
Cainelli N, Forestan C, Angeli D, Villegas TR, Costa F, Botton A, Rasori A, Bonghi C, Ruperti B. Transcriptomic Insights on the Preventive Action of Apple (cv Granny Smith) Skin Wounding on Superficial Scald Development. International Journal of Molecular Sciences. 2021; 22(24):13425. https://doi.org/10.3390/ijms222413425
Chicago/Turabian StyleCainelli, Nadia, Cristian Forestan, Dario Angeli, Tomas Roman Villegas, Fabrizio Costa, Alessandro Botton, Angela Rasori, Claudio Bonghi, and Benedetto Ruperti. 2021. "Transcriptomic Insights on the Preventive Action of Apple (cv Granny Smith) Skin Wounding on Superficial Scald Development" International Journal of Molecular Sciences 22, no. 24: 13425. https://doi.org/10.3390/ijms222413425
APA StyleCainelli, N., Forestan, C., Angeli, D., Villegas, T. R., Costa, F., Botton, A., Rasori, A., Bonghi, C., & Ruperti, B. (2021). Transcriptomic Insights on the Preventive Action of Apple (cv Granny Smith) Skin Wounding on Superficial Scald Development. International Journal of Molecular Sciences, 22(24), 13425. https://doi.org/10.3390/ijms222413425