
GDF15 Supports the Inflammatory Response of PdL Fibroblasts Stimulated by P. gingivalis LPS and Concurrent Compression

 ,
,
Abstract
:1. Introduction
2. Results
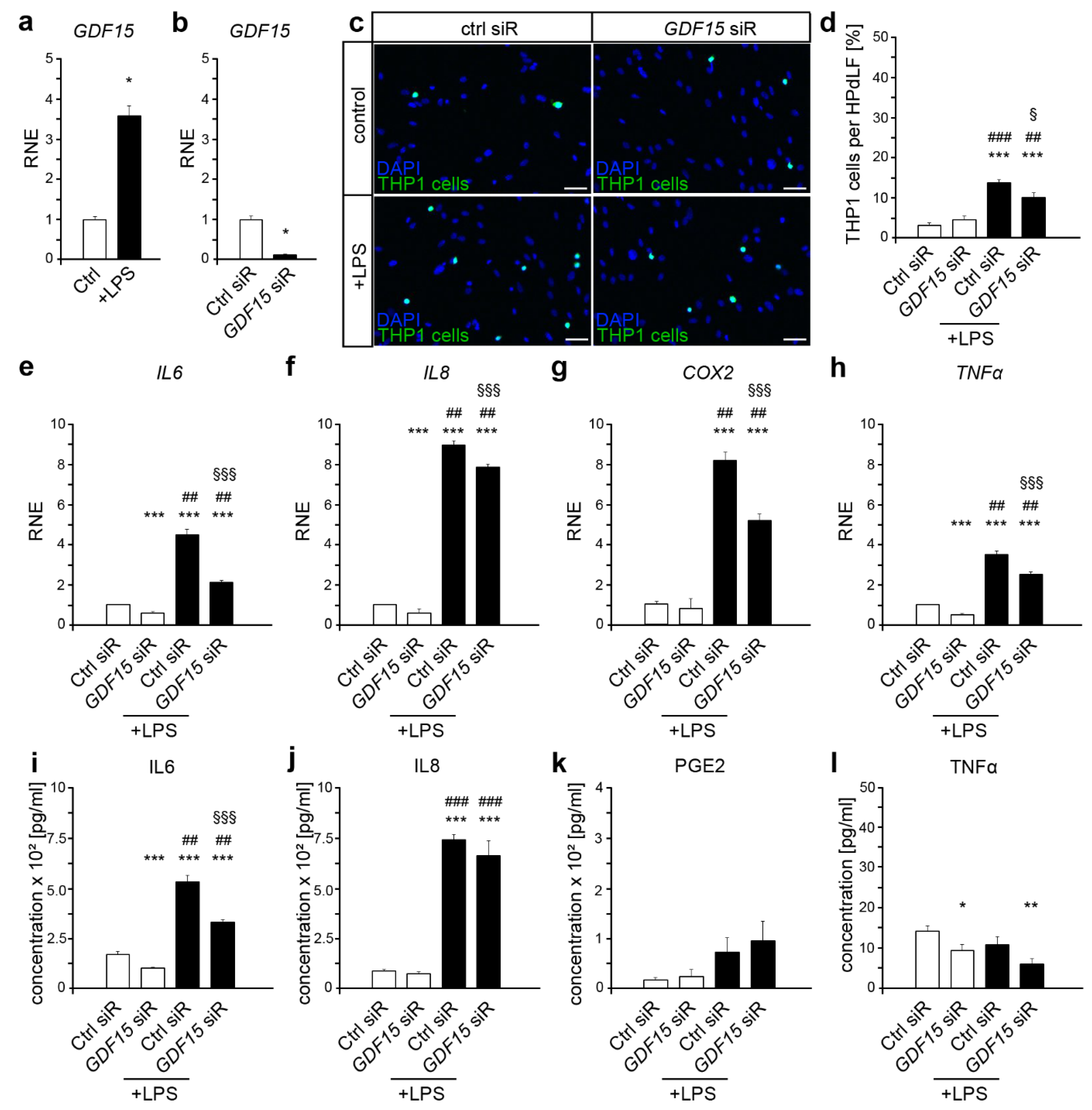
2.1. GDF15 Is Involved in the Inflammatory Response to P. gingivalis LPS
2.2. GDF15 Promotes the Inflammatory Response of HPdLF to Compressive Force
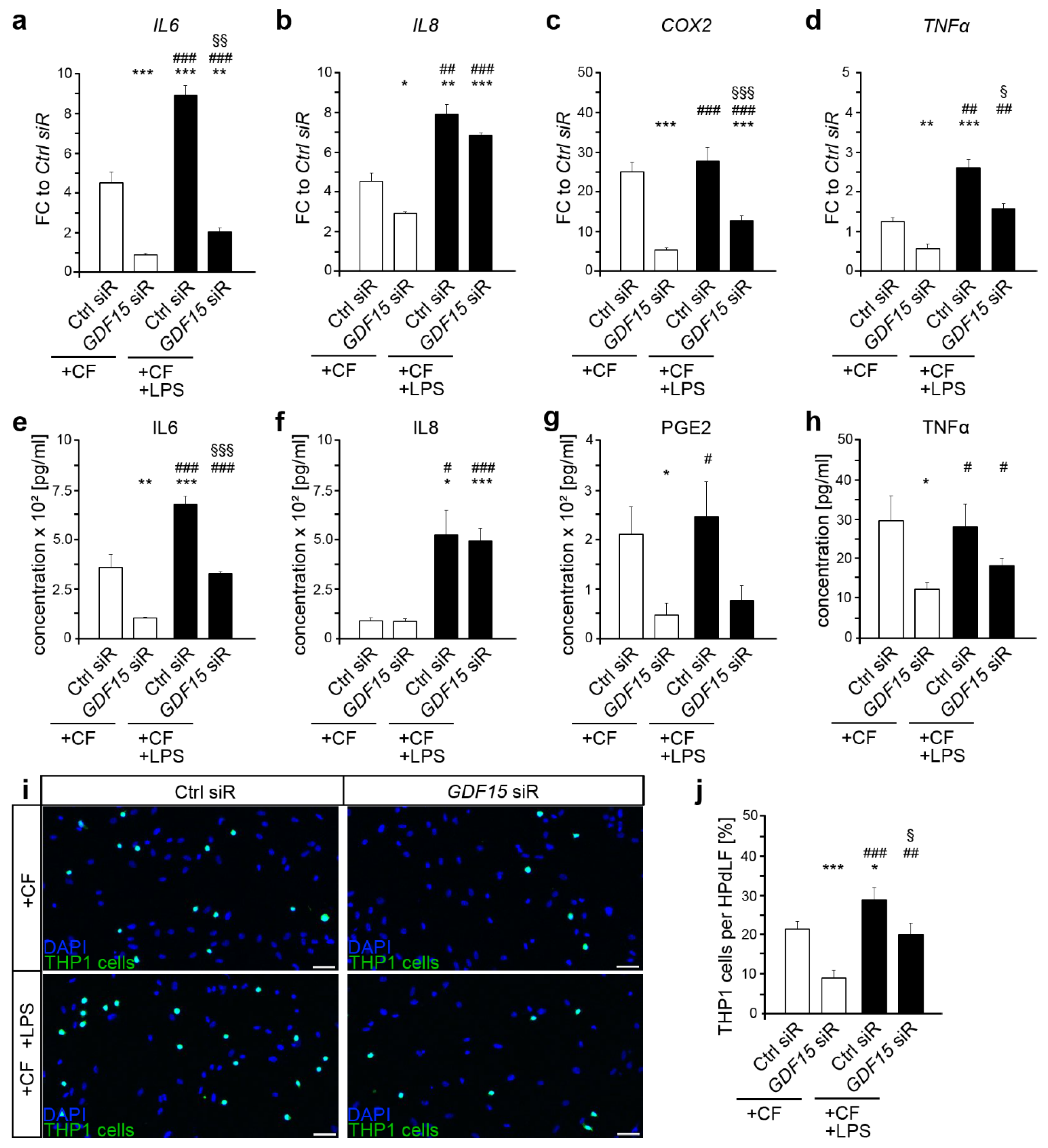
2.3. Additional Exposure to Bacterial Stimulants Enhanced the Inflammatory Response in Mechanically Stressed HPdLF, Even in the Presence of GDF15 Deficiency
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. siRNA-Mediated Knockdown
4.3. P. gingivalis LPS Stimulation
4.4. Mechanical Compression
4.5. THP1 Cell Adherence Assay
4.6. RNA Extraction and Quantitative PCR
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Microscopy, Image Analysis and Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bootcov, M.R.; Bauskin, A.R.; Valenzuela, S.M.; Moore, A.G.; Bansal, M.; He, X.Y.; Zhang, H.P.; Donnellan, M.; Mahler, S.; Pryor, K.; et al. Mic-1, a novel macrophage inhibitory cytokine, is a divergent member of the tgf-beta superfamily. Proc. Natl. Acad. Sci. USA 1997, 94, 11514–11519. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Legere, S.; Haidl, I.; Marshall, J.; MacLeod, J.B.; Aguiar, C.; Lutchmedial, S.; Hassan, A.; Brunt, K.R.; Kienesberger, P.; et al. Serum gdf15, a promising biomarker in obese patients undergoing heart surgery. Front. Cardiovasc. Med. 2020, 7, 103. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.B.; Sapinoso, L.M.; Kern, S.G.; Brown, D.A.; Liu, T.; Bauskin, A.R.; Ward, R.L.; Hawkins, N.J.; Quinn, D.I.; Russell, P.J.; et al. Large-scale delineation of secreted protein biomarkers overexpressed in cancer tissue and serum. Proc. Natl. Acad. Sci. USA 2003, 100, 3410–3415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Huang, Y.; Lyu, Y.; Dai, W.; Tong, Y.; Li, Y. Gdf15 as a biomarker of ageing. Exp. Gerontol. 2021, 146, 111228. [Google Scholar] [CrossRef]
- Bottner, M.; Suter-Crazzolara, C.; Schober, A.; Unsicker, K. Expression of a novel member of the tgf-beta superfamily, growth/differentiation factor-15/macrophage-inhibiting cytokine-1 (gdf-15/mic-1) in adult rat tissues. Cell Tissue Res. 1999, 297, 103–110. [Google Scholar]
- Hsiao, E.C.; Koniaris, L.G.; Zimmers-Koniaris, T.; Sebald, S.M.; Huynh, T.V.; Lee, S.J. Characterization of growth-differentiation factor 15, a transforming growth factor beta superfamily member induced following liver injury. Mol. Cell. Biol. 2000, 20, 3742–3751. [Google Scholar] [CrossRef]
- Bauskin, A.R.; Brown, D.A.; Kuffner, T.; Johnen, H.; Luo, X.W.; Hunter, M.; Breit, S.N. Role of macrophage inhibitory cytokine-1 in tumorigenesis and diagnosis of cancer. Cancer Res. 2006, 66, 4983–4986. [Google Scholar] [CrossRef] [Green Version]
- Breit, S.N.; Johnen, H.; Cook, A.D.; Tsai, V.W.; Mohammad, M.G.; Kuffner, T.; Zhang, H.P.; Marquis, C.P.; Jiang, L.; Lockwood, G.; et al. The tgf-beta superfamily cytokine, mic-1/gdf15: A pleotrophic cytokine with roles in inflammation, cancer and metabolism. Growth Factors 2011, 29, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Deng, C.; Li, Y.P. Tgf-beta and bmp signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [Green Version]
- Vanhara, P.; Lincova, E.; Kozubik, A.; Jurdic, P.; Soucek, K.; Smarda, J. Growth/differentiation factor-15 inhibits differentiation into osteoclasts—A novel factor involved in control of osteoclast differentiation. Differ. Res. Biol. Divers. 2009, 78, 213–222. [Google Scholar] [CrossRef]
- Hinoi, E.; Ochi, H.; Takarada, T.; Nakatani, E.; Iezaki, T.; Nakajima, H.; Fujita, H.; Takahata, Y.; Hidano, S.; Kobayashi, T.; et al. Positive regulation of osteoclastic differentiation by growth differentiation factor 15 upregulated in osteocytic cells under hypoxia. J. Bone Miner. Res. 2012, 27, 938–949. [Google Scholar] [CrossRef]
- Westhrin, M.; Moen, S.H.; Holien, T.; Mylin, A.K.; Heickendorff, L.; Olsen, O.E.; Sundan, A.; Turesson, I.; Gimsing, P.; Waage, A.; et al. Growth differentiation factor 15 (gdf15) promotes osteoclast differentiation and inhibits osteoblast differentiation and high serum gdf15 levels are associated with multiple myeloma bone disease. Haematologica 2015, 100, e511–e514. [Google Scholar] [CrossRef] [Green Version]
- Symmank, J.; Zimmermann, S.; Goldschmitt, J.; Schiegnitz, E.; Wolf, M.; Wehrbein, H.; Jacobs, C. Mechanically-induced gdf15 secretion by periodontal ligament fibroblasts regulates osteogenic transcription. Sci. Rep. 2019, 9, 11516. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Li, Q.; Zhu, Y.; Hu, W. Gdf15 induced by compressive force contributes to osteoclast differentiation in human periodontal ligament cells. Exp. Cell Res. 2020, 387, 111745. [Google Scholar] [CrossRef]
- Nanci, A.; Bosshardt, D.D. Structure of periodontal tissues in health and disease. Periodontology 2000 2006, 40, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Trubiani, O.; Pizzicannella, J.; Caputi, S.; Marchisio, M.; Mazzon, E.; Paganelli, R.; Paganelli, A.; Diomede, F. Periodontal ligament stem cells: Current knowledge and future perspectives. Stem Cells Dev. 2019, 28, 995–1003. [Google Scholar] [CrossRef]
- Caputi, S.; Trubiani, O.; Sinjari, B.; Trofimova, S.; Diomede, F.; Linkova, N.; Diatlova, A.; Khavinson, V. Effect of short peptides on neuronal differentiation of stem cells. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738419828613. [Google Scholar] [CrossRef] [Green Version]
- Basdra, E.K.; Komposch, G. Osteoblast-like properties of human periodontal ligament cells: An in vitro analysis. Eur. J. Orthod. 1997, 19, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jacox, L.A.; Little, S.H.; Ko, C.C. Orthodontic tooth movement: The biology and clinical implications. Kaohsiung J. Med. Sci. 2018, 34, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, D.; Nebel, D.; Bratthall, G.; Nilsson, B.O. The human periodontal ligament cell: A fibroblast-like cell acting as an immune cell. J. Periodontal. Res. 2011, 46, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Proff, P.; Reicheneder, C.; Faltermeier, A.; Kubein-Meesenburg, D.; Romer, P. Effects of mechanical and bacterial stressors on cytokine and growth-factor expression in periodontal ligament cells. J. Orofac. Orthop. Fortschr. Kieferorthopadie 2014, 75, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Fukasawa, S. Is inflammation a friend or foe for orthodontic treatment?: Inflammation in orthodontically induced inflammatory root resorption and accelerating tooth movement. Int. J. Mol. Sci. 2021, 22, 2388. [Google Scholar] [CrossRef]
- Weltman, B.; Vig, K.W.; Fields, H.W.; Shanker, S.; Kaizar, E.E. Root resorption associated with orthodontic tooth movement: A systematic review. Am. J. Orthod. Dentofac. Orthop. 2010, 137, 462–476, discussion 412A. [Google Scholar] [CrossRef] [Green Version]
- Wishney, M. Potential risks of orthodontic therapy: A critical review and conceptual framework. Aust. Dent. J. 2017, 62 (Suppl. S1), 86–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsulaiman, A.A.; Kaye, E.; Jones, J.; Cabral, H.; Leone, C.; Will, L.; Garcia, R. Incisor malalignment and the risk of periodontal disease progression. Am. J. Orthod. Dentofac. Orthop. 2018, 153, 512–522. [Google Scholar] [CrossRef]
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal diseases. Nat. Rev. Dis. Primers 2017, 3, 17038. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, H.F.M.; Patil, S.H.; Pangam, T.S.; Rathod, K.V. Polymicrobial synergy and dysbiosis: An overview. J. Indian Soc. Periodontol. 2018, 22, 101–106. [Google Scholar] [CrossRef]
- Nakayama, M.; Ohara, N. Molecular mechanisms of porphyromonas gingivalis-host cell interaction on periodontal diseases. Jpn. Dent. Sci. Rev. 2017, 53, 134–140. [Google Scholar] [CrossRef]
- D’Ercole, S.; D’Addazio, G.; Di Lodovico, S.; Traini, T.; Di Giulio, M.; Sinjari, B. Porphyromonas gingivalis load is balanced by 0.20% chlorhexidine gel. A randomized, double-blind, controlled, microbiological and immunohistochemical human study. J. Clin. Med. 2020, 9, 284. [Google Scholar]
- Gao, L.; Kang, M.; Zhang, M.J.; Reza Sailani, M.; Kuraji, R.; Martinez, A.; Ye, C.; Kamarajan, P.; Le, C.; Zhan, L.; et al. Polymicrobial periodontal disease triggers a wide radius of effect and unique virome. npj Biofilms Microbiomes 2020, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Shi, J.; Zhao, Y.; Yan, F.; Lei, L.; Li, H. Porphyromonas gingivalis triggers inflammatory responses in periodontal ligament cells by succinate-succinate dehydrogenase-hif-1alpha axis. Biochem. Biophys. Res. Commun. 2020, 522, 184–190. [Google Scholar] [CrossRef]
- Symmank, J.; Appel, S.; Bastian, J.A.; Knaup, I.; Marciniak, J.; Hennig, C.L.; Doding, A.; Schulze-Spate, U.; Jacobs, C.; Wolf, M. Hyperlipidemic conditions impact force-induced inflammatory response of human periodontal ligament fibroblasts concomitantly challenged with p. Gingivalis-lps. Int. J. Mol. Sci. 2021, 22, 6069. [Google Scholar] [CrossRef]
- Nativel, B.; Couret, D.; Giraud, P.; Meilhac, O.; d’Hellencourt, C.L.; Viranaicken, W.; Da Silva, C.R. Porphyromonas gingivalis lipopolysaccharides act exclusively through tlr4 with a resilience between mouse and human. Sci. Rep. 2017, 7, 15789. [Google Scholar] [CrossRef] [PubMed]
- Darveau, R.P.; Pham, T.T.; Lemley, K.; Reife, R.A.; Bainbridge, B.W.; Coats, S.R.; Howald, W.N.; Way, S.S.; Hajjar, A.M. Porphyromonas gingivalis lipopolysaccharide contains multiple lipid a species that functionally interact with both toll-like receptors 2 and 4. Infect. Immun. 2004, 72, 5041–5051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, I.; Colaco, H.G.; Neves-Costa, A.; Seixas, E.; Velho, T.R.; Pedroso, D.; Barros, A.; Martins, R.; Carvalho, N.; Payen, D.; et al. Cxcl5-mediated recruitment of neutrophils into the peritoneal cavity of gdf15-deficient mice protects against abdominal sepsis. Proc. Natl. Acad. Sci. USA 2020, 117, 12281–12287. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Zhao, F.; Zhao, Y.; Liu, H.; Wang, Z. Atf4-mediated gdf15 suppresses lps-induced inflammation and muc5ac in human nasal epithelial cells through the pi3k/akt pathway. Life Sci. 2021, 275, 119356. [Google Scholar] [CrossRef]
- Wu, Q.; Jiang, D.; Schaefer, N.R.; Harmacek, L.; O’Connor, B.P.; Eling, T.E.; Eickelberg, O.; Chu, H.W. Overproduction of growth differentiation factor 15 promotes human rhinovirus infection and virus-induced inflammation in the lung. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 314, L514–L527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ago, T.; Sadoshima, J. Gdf15, a cardioprotective tgf-beta superfamily protein. Circ. Res. 2006, 98, 294–297. [Google Scholar] [CrossRef] [Green Version]
- Emmerson, P.J.; Duffin, K.L.; Chintharlapalli, S.; Wu, X. Gdf15 and growth control. Front. Physiol. 2018, 9, 1712. [Google Scholar] [CrossRef] [Green Version]
- Luan, H.H.; Wang, A.; Hilliard, B.K.; Carvalho, F.; Rosen, C.E.; Ahasic, A.M.; Herzog, E.L.; Kang, I.; Pisani, M.A.; Yu, S.; et al. Gdf15 is an inflammation-induced central mediator of tissue tolerance. Cell 2019, 178, 1231–1244.e11. [Google Scholar] [CrossRef]
- Assadi, A.; Zahabi, A.; Hart, R.A. Gdf15, an update of the physiological and pathological roles it plays: A review. Pflug. Arch. 2020, 472, 1535–1546. [Google Scholar] [CrossRef]
- Abulizi, P.; Loganathan, N.; Zhao, D.; Mele, T.; Zhang, Y.; Zwiep, T.; Liu, K.; Zheng, X. Growth differentiation factor-15 deficiency augments inflammatory response and exacerbates septic heart and renal injury induced by lipopolysaccharide. Sci. Rep. 2017, 7, 1037. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Kosak, J.P.; Kim, J.K.; Kissling, G.; Germolec, D.R.; Zeldin, D.C.; Bradbury, J.A.; Baek, S.J.; Eling, T.E. Nag-1/gdf15 transgenic mouse has less white adipose tissue and a reduced inflammatory response. Mediat. Inflamm. 2013, 2013, 641851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalli, M.; Minia, A.; Pliaka, V.; Fotis, C.; Alexopoulos, L.G.; Stylianopoulos, T. Solid stress-induced migration is mediated by gdf15 through akt pathway activation in pancreatic cancer cells. Sci. Rep. 2019, 9, 978. [Google Scholar] [CrossRef] [PubMed]
- Kalli, M.; Voutouri, C.; Minia, A.; Pliaka, V.; Fotis, C.; Alexopoulos, L.G.; Stylianopoulos, T. Mechanical compression regulates brain cancer cell migration through mek1/erk1 pathway activation and gdf15 expression. Front. Oncol. 2019, 9, 992. [Google Scholar] [CrossRef] [Green Version]
- Ratnam, N.M.; Peterson, J.M.; Talbert, E.E.; Ladner, K.J.; Rajasekera, P.V.; Schmidt, C.R.; Dillhoff, M.E.; Swanson, B.J.; Haverick, E.; Kladney, R.D.; et al. Nf-kappab regulates gdf-15 to suppress macrophage surveillance during early tumor development. J. Clin. Investig. 2017, 127, 3796–3809. [Google Scholar] [CrossRef] [Green Version]
- Moelants, E.A.; Mortier, A.; Van Damme, J.; Proost, P. Regulation of tnf-alpha with a focus on rheumatoid arthritis. Immunol. Cell Biol. 2013, 91, 393–401. [Google Scholar] [CrossRef]
- Szalecki, M.; Malinowska, A.; Prokop-Piotrkowska, M.; Janas, R. Interactions between the growth hormone and cytokines—A review. Adv. Med. Sci. 2018, 63, 285–289. [Google Scholar] [CrossRef]
- Akdis, M.; Aab, A.; Altunbulakli, C.; Azkur, K.; Costa, R.A.; Crameri, R.; Duan, S.; Eiwegger, T.; Eljaszewicz, A.; Ferstl, R.; et al. Interleukins (from il-1 to il-38), interferons, transforming growth factor beta, and tnf-alpha: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2016, 138, 984–1010. [Google Scholar] [CrossRef] [Green Version]
- Marchant, A.; Bruyns, C.; Vandenabeele, P.; Ducarme, M.; Gerard, C.; Delvaux, A.; De Groote, D.; Abramowicz, D.; Velu, T.; Goldman, M. Interleukin-10 controls interferon-gamma and tumor necrosis factor production during experimental endotoxemia. Eur. J. Immunol. 1994, 24, 1167–1171. [Google Scholar] [CrossRef]
- Muchamuel, T.; Menon, S.; Pisacane, P.; Howard, M.C.; Cockayne, D.A. Il-13 protects mice from lipopolysaccharide-induced lethal endotoxemia: Correlation with down-modulation of tnf-alpha, ifn-gamma, and il-12 production. J. Immunol. 1997, 158, 2898–2903. [Google Scholar]
- Marciniak, J.; Lossdorfer, S.; Knaup, I.; Bastian, A.; Craveiro, R.B.; Jager, A.; Wolf, M. Orthodontic cell stress modifies proinflammatory cytokine expression in human pdl cells and induces immunomodulatory effects via tlr-4 signaling in vitro. Clin. Oral Investig. 2020, 24, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Klein, Y.; Fleissig, O.; Polak, D.; Barenholz, Y.; Mandelboim, O.; Chaushu, S. Immunorthodontics: In vivo gene expression of orthodontic tooth movement. Sci. Rep. 2020, 10, 8172. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kita, M.; Yamamoto, K.; Akamatsu, Y.; Oseko, F.; Kanamura, N. Mechanical stress enhances production of cytokines in human periodontal ligament cells induced by porphyromonas gingivalis. Arch. Oral Biol. 2011, 56, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Suriben, R.; Chen, M.; Higbee, J.; Oeffinger, J.; Ventura, R.; Li, B.; Mondal, K.; Gao, Z.; Ayupova, D.; Taskar, P.; et al. Antibody-mediated inhibition of gdf15-gfral activity reverses cancer cachexia in mice. Nat. Med. 2020, 26, 1264–1270. [Google Scholar] [CrossRef]
- Lerner, L.; Tao, J.; Liu, Q.; Nicoletti, R.; Feng, B.; Krieger, B.; Mazsa, E.; Siddiquee, Z.; Wang, R.; Huang, L.; et al. Map3k11/gdf15 axis is a critical driver of cancer cachexia. J. Cachexia Sarcopenia Muscle 2016, 7, 467–482. [Google Scholar] [CrossRef]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Insights into mechanisms of gdf15 and receptor gfral: Therapeutic targets. Trends Endocrinol. Metab. TEM 2020, 31, 939–951. [Google Scholar] [CrossRef]
- Wen, W.; Chau, E.; Jackson-Boeters, L.; Elliott, C.; Daley, T.D.; Hamilton, D.W. Tgf-ss1 and fak regulate periostin expression in pdl fibroblasts. J. Dent. Res. 2010, 89, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Artz, A.; Butz, S.; Vestweber, D. Gdf-15 inhibits integrin activation and mouse neutrophil recruitment through the alk-5/tgf-betarii heterodimer. Blood 2016, 128, 529–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschneck, C.; Batschkus, S.; Proff, P.; Kostler, J.; Spanier, G.; Schroder, A. Valid gene expression normalization by rt-qpcr in studies on hpdl fibroblasts with focus on orthodontic tooth movement and periodontitis. Sci. Rep. 2017, 7, 14751. [Google Scholar] [CrossRef] [Green Version]
- Symmank, J.; Chorus, M.; Appel, S.; Marciniak, J.; Knaup, I.; Bastian, A.; Hennig, C.L.; Doding, A.; Schulze-Spate, U.; Jacobs, C.; et al. Distinguish fatty acids impact survival, differentiation and cellular function of periodontal ligament fibroblasts. Sci. Rep. 2020, 10, 15706. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Gene Symbol | NCBI Gene ID | Primer Sequence | Length |
|---|---|---|---|---|
| growth differentiation factor 15 | GDF15 | 9518 | fw CCGAAGACTCCAGATTCCGA rew CCCGAGAGATACGCAGGTG | 180 bp |
| C-X-C motif chemokine ligand 8 | IL8 | 3576 | fw TTGGCAGCCTTCCTGATTTCT rew GGTCCACTCTCAATCACTCTCA | 149 bp |
| Interleukin 6 | IL6 | 3569 | fw CATCCTCGACGGCATCTCAG rew TCACCAGGCAAGTCTCCTCA | 164 bp |
| Prostaglandin-endoperoxide synthase 2 | PTGS2(COX2) | 5743 | fw GATGATTGCCCGACTCCCTT rew GGCCCTCGCTTATGATCTGT | 185 bp |
| Ribosomal protein L22 | RPL22 | 6146 | fw TGATTGCACCCACCCTGTAG rev GGTTCCCAGCTTTTCCGTTC | 98 bp |
| TATA-box binding protein | TBP | 6908 | fw CGGCTGTTTAACTTCGCTTCC rev TGGGTTATCTTCACACGCCAAG | 86 bp |
| Tumor necrosis factor | TNFα | 7124 | fw CACGCTCTTCTGCCTGCTG rev AGGCTTGTCACTCGGGGTT | 130 bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stemmler, A.; Symmank, J.; Steinmetz, J.; von Brandenstein, K.; Hennig, C.-L.; Jacobs, C. GDF15 Supports the Inflammatory Response of PdL Fibroblasts Stimulated by P. gingivalis LPS and Concurrent Compression. Int. J. Mol. Sci. 2021, 22, 13608. https://doi.org/10.3390/ijms222413608
Stemmler A, Symmank J, Steinmetz J, von Brandenstein K, Hennig C-L, Jacobs C. GDF15 Supports the Inflammatory Response of PdL Fibroblasts Stimulated by P. gingivalis LPS and Concurrent Compression. International Journal of Molecular Sciences. 2021; 22(24):13608. https://doi.org/10.3390/ijms222413608
Chicago/Turabian StyleStemmler, Albert, Judit Symmank, Julia Steinmetz, Katrin von Brandenstein, Christoph-Ludwig Hennig, and Collin Jacobs. 2021. "GDF15 Supports the Inflammatory Response of PdL Fibroblasts Stimulated by P. gingivalis LPS and Concurrent Compression" International Journal of Molecular Sciences 22, no. 24: 13608. https://doi.org/10.3390/ijms222413608
APA StyleStemmler, A., Symmank, J., Steinmetz, J., von Brandenstein, K., Hennig, C. -L., & Jacobs, C. (2021). GDF15 Supports the Inflammatory Response of PdL Fibroblasts Stimulated by P. gingivalis LPS and Concurrent Compression. International Journal of Molecular Sciences, 22(24), 13608. https://doi.org/10.3390/ijms222413608