
Resveratrol Supplementation Attenuates Cognitive and Molecular Alterations under Maternal High-Fat Diet Intake: Epigenetic Inheritance over Generations




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
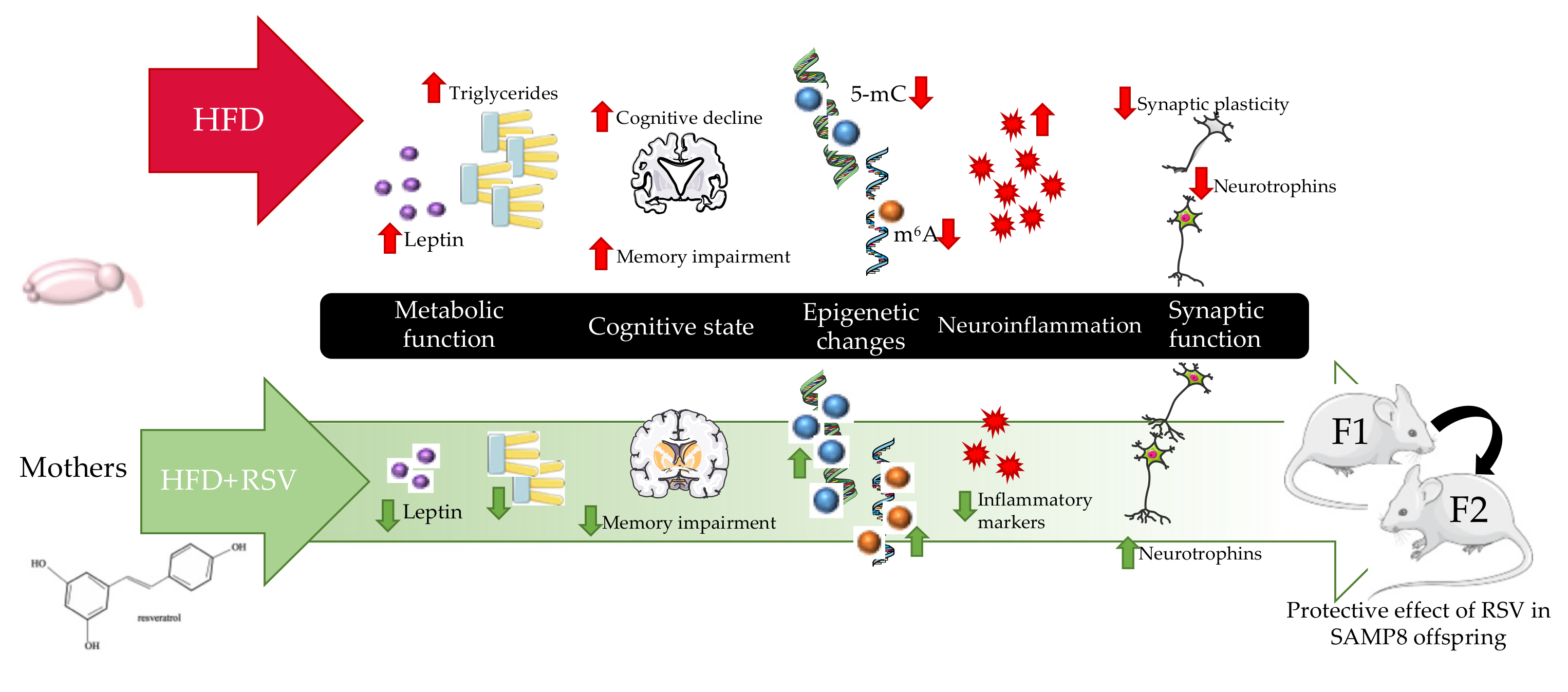
Abstract
:1. Introduction
2. Results
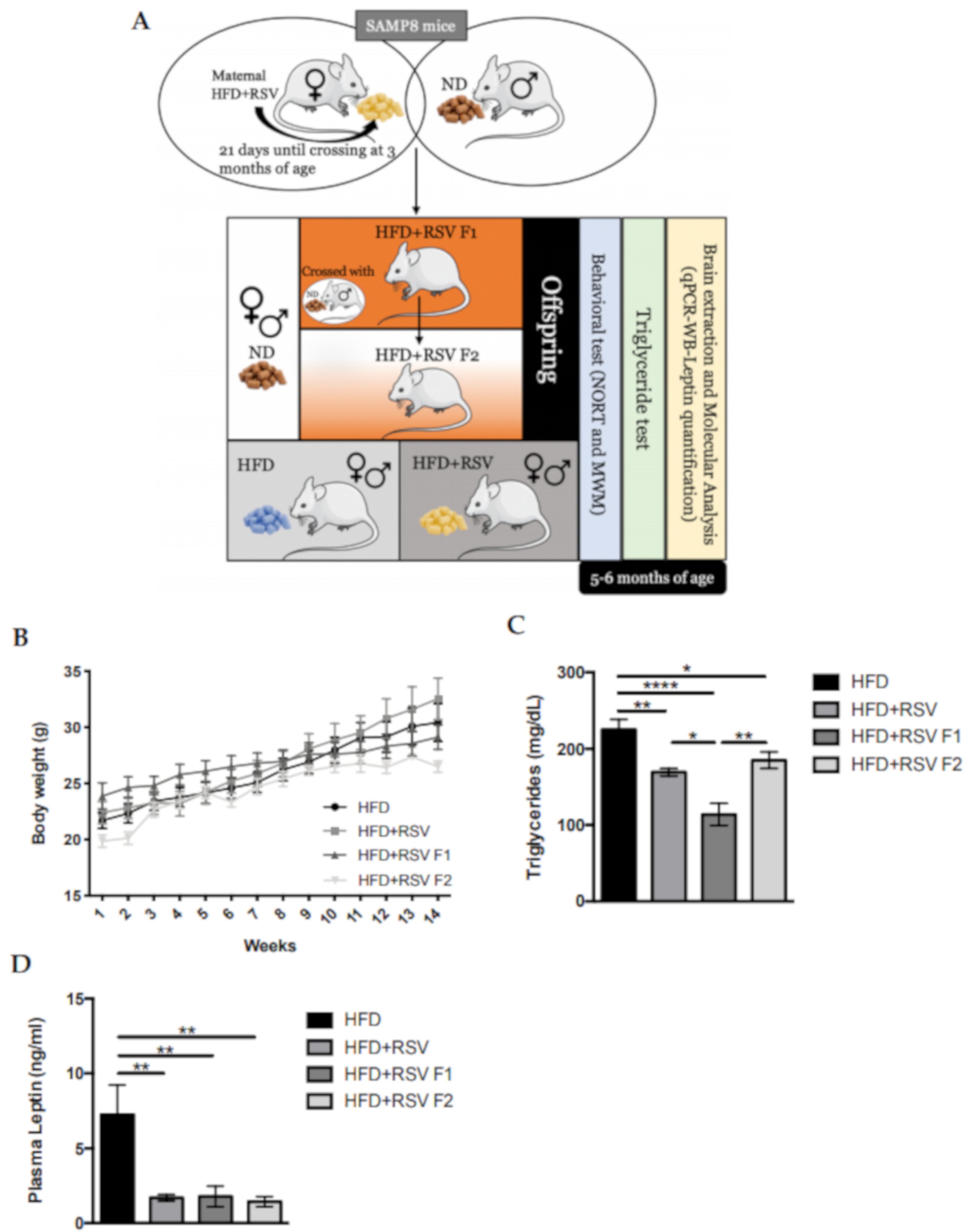
2.1. Body Weight Progression in SAMP8 HFD + RSV and Its Offspring
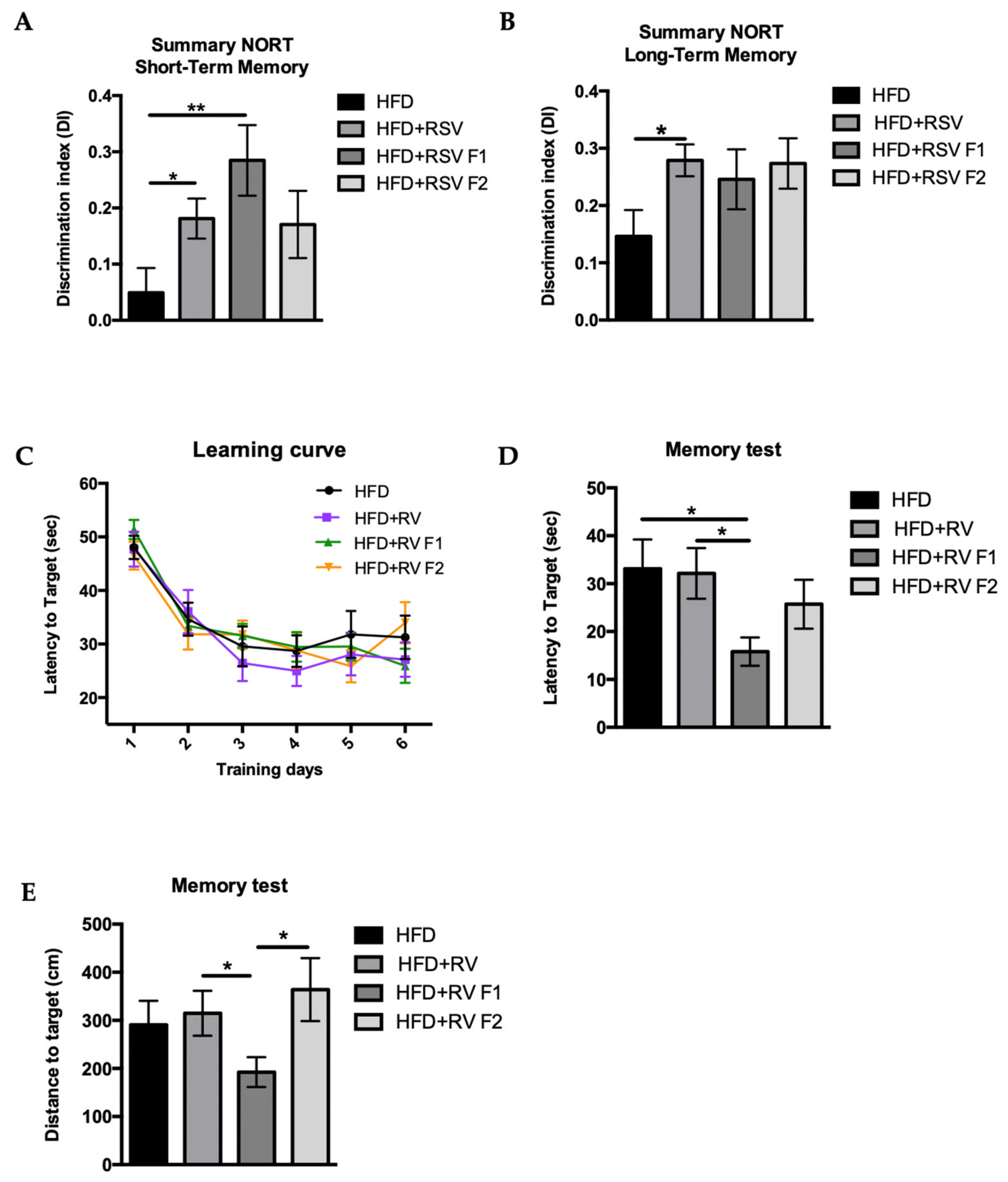
2.2. RSV Prevented Cognitive Impairment Induced by HFD in SAMP8 Mice across Generations
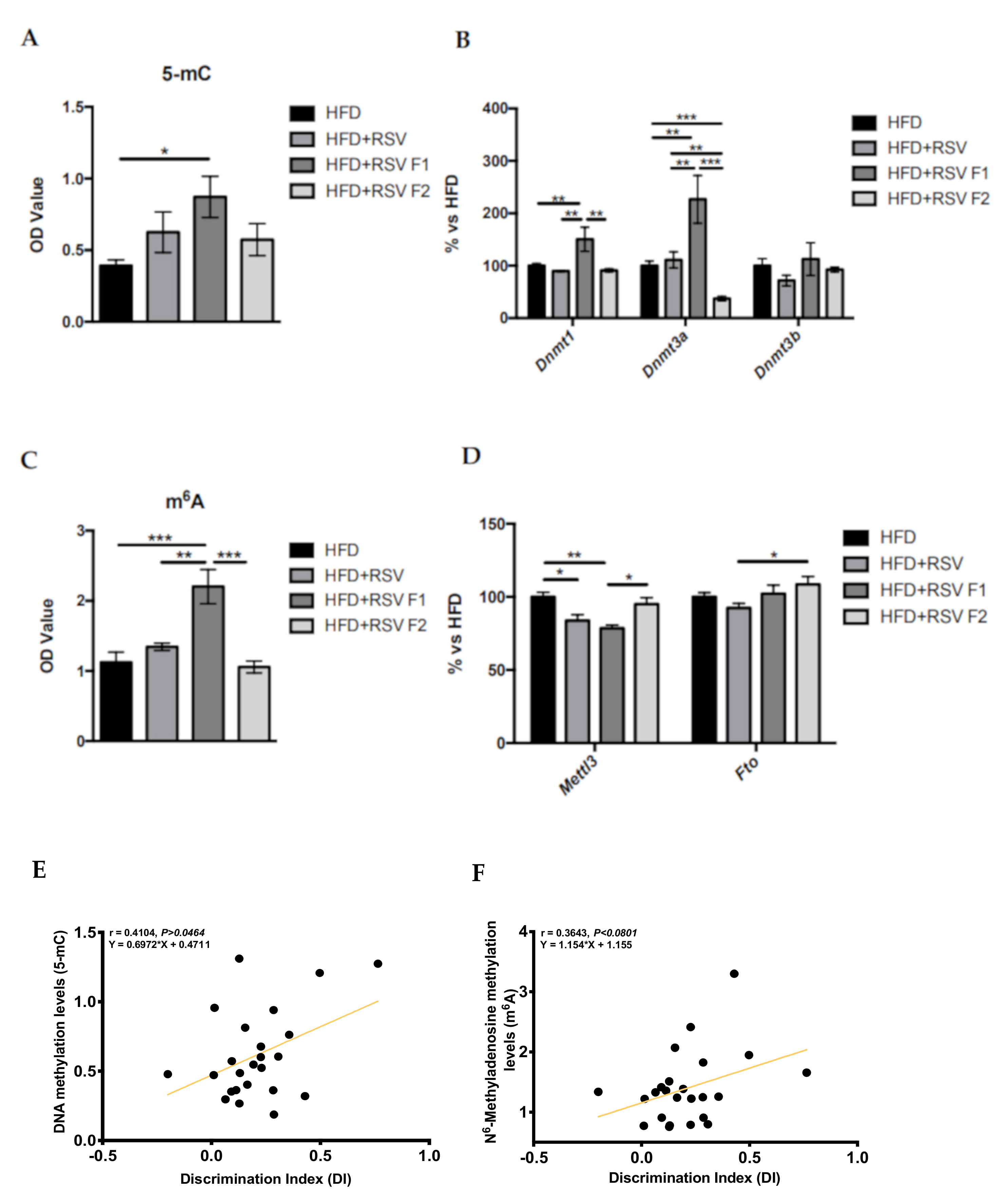
2.3. Adult Offspring of SAMP8 Mice Showed Global Changes in Methylation Patterns and Its Machinery after Maternal HFD + RSV Diet, as Well as a Correlation with Cognitive Improvement
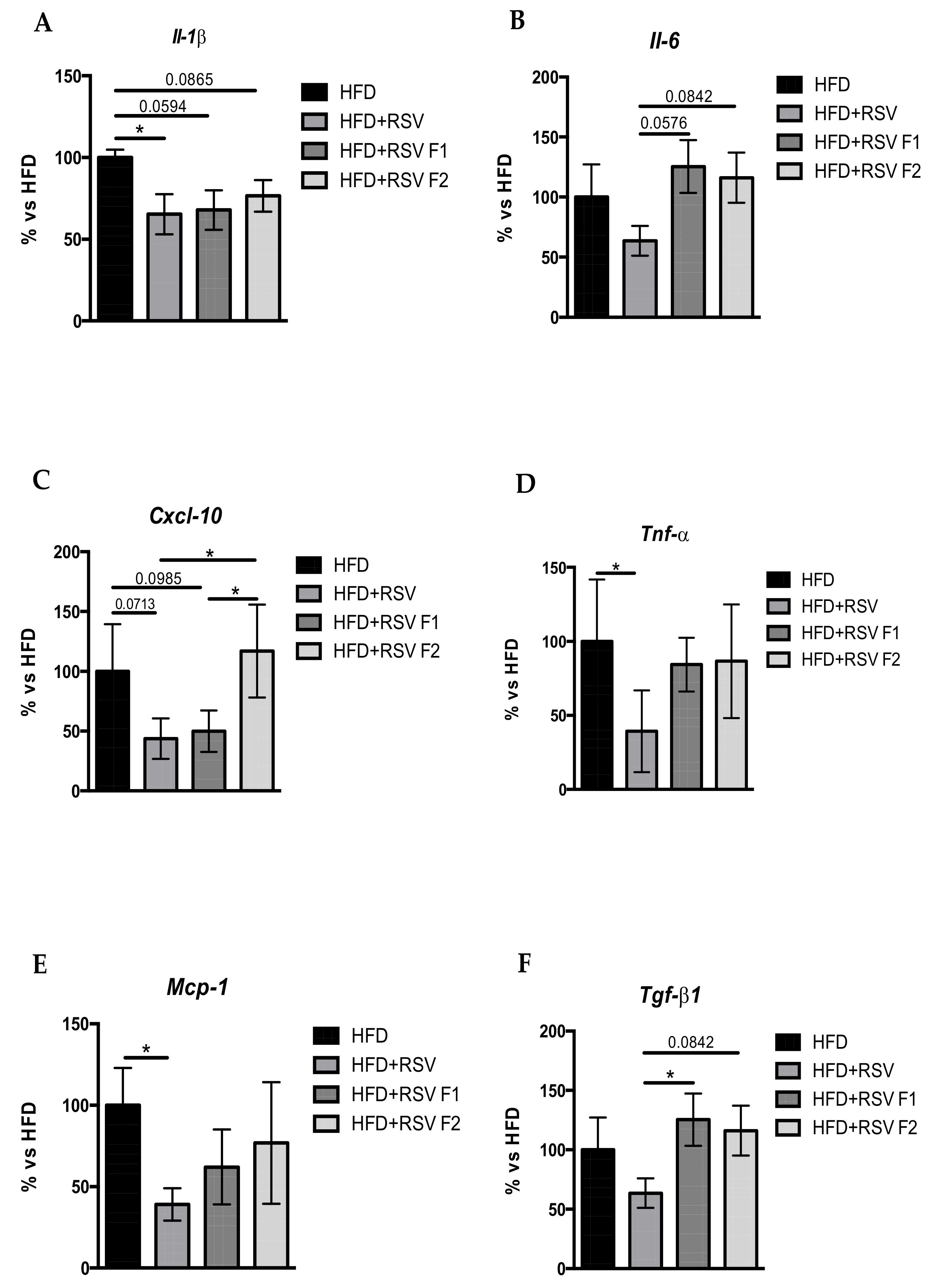
2.4. RSV Diet Modified the Inflammatory Markers in the Hippocampus of SAMP8 and Its Offspring under HFD
2.5. RSV Diet Rescued Synaptic Dysfunction, Reverting the Effects of HFD in the Hippocampus of SAMP8 Mice and Its Offspring
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Behavioral Test
4.2.1. Novel Object Recognition Test (NORT)
4.2.2. Morris Water Maze (MWM)
4.3. Immunodetection Experiments
4.3.1. Brain Processing
4.3.2. Plasma Isolated, Leptin and Triglycerides Quantification
4.3.3. Western Blotting (WB)
4.4. RNA Extraction and Gene Expression Determination by q-PCR
4.5. Global DNA Methylation Determination
4.6. m6A RNA Methylation Quantification
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| Akt | Protein kinase B |
| ANOVA | One-way analysis of variance |
| APP/PS1 mice | Double transgenic mice of amyloid precursor protein and presenilin 1 |
| BDNF | Bran-derived neurotrophic factor |
| cDNA | Complementary DNA |
| CNS | Central nervous system |
| Ct | Cycle threshold |
| Cxcl-10 | C-X-C motif chemokine ligand 10 |
| DI | Discrimination index |
| DNA | Desoxyribonucleic acid |
| 5-hmC | Hydroxymethylation |
| 5-mC | DNA methylation |
| Dnmt1 | DNA methyltransferase 1 |
| Dnmt3a | DNA methyltransferase 3 alpha |
| Dnmt3b | DNA methyltransferase 3 beta |
| Fto | Alpha-ketoglutarate dependent dioxygenase |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GDNF | Glial cell-derived neurotrophic factor |
| HFD | High fat diet |
| HFD+RSV | High fat diet with resveratrol parental group |
| HFD+RSV F1 | High fat diet with resveratrol first generation |
| HFD+RSV F2 | High fat diet with resveratrol second generation |
| Il-6 | Interleukin 6 |
| Il-1β | Interleukin 1 beta |
| LTP | Long term potentiation |
| m6A | N-6-methyladenosine |
| Mcp-1 | Monocyte chemoattractant protein 1 |
| MeS | Metabolic syndrome |
| Mettl3 | Methyltransferase like 3 |
| mRNA | Mature messenger RNA |
| MWM | Morris water maze |
| ND | Normal diet |
| NF-κβ | Nuclear factor NF-kappa-B |
| NGF | Nerve growth factor |
| NORT | Novel object recognition test |
| NT3 | Neurotrophin-3 |
| OD | Optical density |
| OS | Oxidative stress |
| p-Akt | Phosphorylated RAC-alpha serine/threonine-protein kinase |
| PCR | Polymerase chain reaction |
| PVDF | Polyvinylidene difluoride |
| qPCR | Real-time quantitative polymerase chain reaction |
| RNA | Ribonucleic acid |
| RT-PCR | Reverse transcriptional-polymerase chain reaction |
| RSV | Resveratrol |
| SAMP8 | Senescence-accelerated mouse prone 8 |
| SAMR1 | Senescence-accelerated mouse resistant 1 |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrolysis |
| SEM | Standard error of the mean |
| SIRT1 | NAD-dependent protein deacetylase sirtuin-1 |
| TBS | Tris-buffer saline |
| TBS-T | Tris-buffer saline tween 20 |
| TG | Triglycerides |
| TGF-β1 | Transforming growth factor beta 1 |
| TN | Time with new object |
| TNF-α | Tumor necrosis factor-alpha |
| TO | Time with old object |
| TrkA | Neurotrophic receptor tyrosine kinase 1 |
| TrkB | Neurotrophic receptor tyrosine kinase 2 |
| WB | Western blot |
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzheimer’s Association. 2018 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2018, 14, 367–429. [Google Scholar] [CrossRef]
- Sugiura, M. Functional neuroimaging of normal aging: Declining brain, adapting brain. Ageing Res. Rev. 2016, 30, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Chernoff, R. The symbiotic relationship between oral health, nutrition, and aging. Generations 2016, 40, 32–38. [Google Scholar]
- Bhatti, G.K.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Lifestyle Modifications and Nutritional Interventions in Aging-Associated Cognitive Decline and Alzheimer’s Disease. Front. Aging Neurosci. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Spencer, S.J.; D’Angelo, H.; Soch, A.; Watkins, L.R.; Maier, S.F.; Barrientos, R.M. High-fat diet and aging interact to produce neuroinflammation and impair hippocampal-and amygdalar-dependent memory. Neurobiol. Aging 2017, 58, 88–101. [Google Scholar] [CrossRef]
- Wang, Z.; Ge, Q.; Wu, Y.; Zhang, J.; Gu, Q.; Han, J. Impairment of Long-term Memory by a Short-term High-fat Diet via Hippocampal Oxidative Stress and Alterations in Synaptic Plasticity. Neuroscience 2020, 424, 24–33. [Google Scholar] [CrossRef]
- Griñan-Ferré, C.; Palomera-Ávalos, V.; Puigoriol-Illamola, D.; Camins, A.; Porquet, D.; Plá, V.; Aguado, F.; Pallàs, M. Behaviour and cognitive changes correlated with hippocampal neuroinflammaging and neuronal markers in female SAMP8, a model of accelerated senescence. Exp. Gerontol. 2016, 80, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Mottahedin, A.; Ardalan, M.; Chumak, T.; Riebe, I.; Ek, J.; Mallard, C. Effect of neuroinflammation on synaptic organization and function in the developing brain: Implications for neurodevelopmental and neurodegenerative disorders. Front. Cell. Neurosci. 2017, 11, 190. [Google Scholar] [CrossRef] [Green Version]
- Di Benedetto, S.; Müller, L.; Wenger, E.; Düzel, S.; Pawelec, G. Contribution of neuroinflammation and immunity to brain aging and the mitigating effects of physical and cognitive interventions. Neurosci. Biobehav. Rev. 2017, 75, 114–128. [Google Scholar] [CrossRef] [Green Version]
- Gibon, J.; Barker, P.A. Neurotrophins and proneurotrophins: Focus on synaptic activity and plasticity in the brain. Neuroscientist 2017, 23, 587–604. [Google Scholar] [CrossRef] [PubMed]
- Leal, G.; Bramham, C.R.; Duarte, C.B. BDNF and hippocampal synaptic plasticity. In Vitamins and Hormones; Litwack, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 104, pp. 153–195. ISBN 0083-6729. [Google Scholar]
- Ren, H.L.; Lv, C.N.; Xing, Y.; Geng, Y.; Zhang, F.; Bu, W.; Wang, M.W. Downregulated nuclear factor E2-related factor 2 (Nrf2) aggravates cognitive impairments via neuroinflammation and synaptic plasticity in the senescence-accelerated mouse prone 8 (SAMP8) mouse: A model of accelerated senescence. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berson, A.; Nativio, R.; Berger, S.L.; Bonini, N.M. Epigenetic regulation in neurodegenerative diseases. Trends. Neurosci. 2018, 41, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wei, Q.; Liu, F.-F.; Hu, F.; Xie, A.; Zhu, L.-Q.; Liu, D. Synaptic dysfunction in Alzheimer’s disease: Aβ, tau, and epigenetic alterations. Mol. Neurobiol. 2018, 55, 3021–3032. [Google Scholar] [CrossRef] [PubMed]
- Gangisetty, O.; Cabrera, M.A.; Murugan, S. Impact of epigenetics in aging and age related neurodegenerative diseases. Front. Biosci. 2018, 23, 1445–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griñán-Ferré, C.; Corpas, R.; Puigoriol-Illamola, D.; Palomera-Ávalos, V.; Sanfeliu, C.; Pallàs, M. Understanding epigenetics in the neurodegeneration of Alzheimer’s disease: SAMP8 mouse model. J. Alzheimer’s Dis. 2018, 62, 943–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Song, M.; Qu, J.; Liu, G.-H. Epigenetic modifications in cardiovascular aging and diseases. Circ. Res. 2018, 123, 773–786. [Google Scholar] [CrossRef]
- Rosen, E.D.; Kaestner, K.H.; Natarajan, R.; Patti, M.-E.; Sallari, R.; Sander, M.; Susztak, K. Epigenetics and epigenomics: Implications for diabetes and obesity. Diabetes 2018, 67, 1923–1931. [Google Scholar] [CrossRef] [Green Version]
- Seki, Y.; Suzuki, M.; Guo, X.; Glenn, A.S.; Vuguin, P.M.; Fiallo, A.; Du, Q.; Ko, Y.-A.; Yu, Y.; Susztak, K. In utero exposure to a high-fat diet programs hepatic hypermethylation and gene dysregulation and development of metabolic syndrome in male mice. Endocrinology 2017, 158, 2860–2872. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Yan, J.; Teng, M.; Yan, S.; Zhou, Z.; Zhu, W. In utero and lactational exposure to BDE-47 promotes obesity development in mouse offspring fed a high-fat diet: Impaired lipid metabolism and intestinal dysbiosis. Arch. Toxicol. 2018, 92, 1847–1860. [Google Scholar] [CrossRef]
- Yang, J.L.; Jiang, H.; Pan, F.; Ho, C.S.H.; Ho, R.C.M. The effects of high-fat-diet combined with chronic unpredictable mild stress on depression-like behavior and leptin/leprb in male rats. Sci. Rep. 2016, 6, 35239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, M.M.; Dias-Rocha, C.P.; Reis-Gomes, C.F.; Wang, H.; Atella, G.C.; Cordeiro, A.; Pazos-Moura, C.C.; Joss-Moore, L.; Trevenzoli, I.H. Maternal high-fat diet impairs leptin signaling and up-regulates type-1 cannabinoid receptor with sex-specific epigenetic changes in the hypothalamus of newborn rats. Psychoneuroendocrinology 2019, 103, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Masuyama, H.; Mitsui, T.; Eguchi, T.; Tamada, S.; Hiramatsu, Y. The effects of paternal high-fat diet exposure on offspring metabolism with epigenetic changes in the mouse adiponectin and leptin gene promoters. Am. J. Physiol. Metab. 2016, 311, E236–E245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, R.M.; Bueno, F.G.; Schamber, C.R.; de Mello, J.C.P.; de Oliveira, J.C.; Francisco, F.A.; Moreira, V.M.; Junior, M.D.F.; Pedrino, G.R.; de Freitas Mathias, P.C. Maternal diet-induced obesity during suckling period programs offspring obese phenotype and hypothalamic leptin/insulin resistance. J. Nutr. Biochem. 2018, 61, 24–32. [Google Scholar] [CrossRef]
- Zhang, Q.; Xiao, X.; Zheng, J.; Li, M.; Yu, M.; Ping, F.; Wang, T.; Wang, X. A Maternal High-Fat Diet Induces DNA Methylation Changes That Contribute to Glucose Intolerance in Offspring. Front. Endocrinol. (Lausanne) 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhang, C.-X.; Liu, Y.-M.; Chen, K.-L.; Chen, G. A comparative study of anti-aging properties and mechanism: Resveratrol and caloric restriction. Oncotarget 2017, 8, 65717. [Google Scholar] [CrossRef] [Green Version]
- Costa, D.; Scognamiglio, M.; Fiorito, C.; Benincasa, G.; Napoli, C. Genetic background, epigenetic factors and dietary interventions which influence human longevity. Biogerontology 2019, 20, 605–626. [Google Scholar] [CrossRef]
- Lange, K.W. Red wine, resveratrol, and Alzheimer’s disease. Mov. Nutr. Health Dis. 2018, 2. [Google Scholar] [CrossRef]
- Szkudelski, T.; Szkudelska, K. Potential of resveratrol in mitigating metabolic disturbances induced by ethanol. Biomed. Pharmacother. 2018, 101, 579–584. [Google Scholar] [CrossRef]
- Pineda-Ramírez, N.; Gutiérrez Aguilar, G.F.; Espinoza-Rojo, M.; Aguilera, P. Current evidence for AMPK activation involvement on resveratrol-induced neuroprotection in cerebral ischemia. Nutr. Neurosci. 2018, 21, 229–247. [Google Scholar] [CrossRef]
- Izquierdo, V.; Palomera-Ávalos, V.; López-Ruiz, S.; Canudas, A.-M.; Pallàs, M.; Griñán-Ferré, C. Maternal resveratrol supplementation prevents cognitive decline in senescent mice offspring. Int. J. Mol. Sci. 2019, 20, 1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.; Feng, Q.; Cheng, J.; Zheng, J. Maternal resveratrol consumption and its programming effects on metabolic health in offspring mechanisms and potential implications. Biosci. Rep. 2018, 38, BSR20171741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, M.-H.; Sheen, J.-M.; Lin, I.; Yu, H.-R.; Tiao, M.-M.; Tain, Y.-L.; Huang, L.-T. Effects of Maternal Resveratrol on Maternal High-Fat Diet/Obesity with or without Postnatal High-Fat Diet. Int. J. Mol. Sci. 2020, 21, 3428. [Google Scholar] [CrossRef] [PubMed]
- Akiguchi, I.; Pallàs, M.; Budka, H.; Akiyama, H.; Ueno, M.; Han, J.; Yagi, H.; Nishikawa, T.; Chiba, Y.; Sugiyama, H. SAMP8 mice as a neuropathological model of accelerated brain aging and dementia: Toshio Takeda’s legacy and future directions. Neuropathology 2017, 37, 293–305. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Liu, J.; Shi, J.-S. SAMP8 mice as a model of age-related cognition decline with underlying mechanisms in Alzheimer’s disease. J. Alzheimer’s Dis. 2020, 75, 385–395. [Google Scholar] [CrossRef]
- Pan, W.; Han, S.; Kang, L.; Li, S.; Du, J.; Cui, H. Effects of dihydrotestosterone on synaptic plasticity of the hippocampus in mild cognitive impairment male SAMP8 mice. Exp. Ther. Med. 2016, 12, 1455–1463. [Google Scholar] [CrossRef] [Green Version]
- Cosín-Tomás, M.; Álvarez-López, M.J.; Companys-Alemany, J.; Kaliman, P.; González-Castillo, C.; Ortuño-Sahagún, D.; Pallàs, M.; Griñán-Ferré, C. Temporal integrative analysis of mRNA and microRNAs expression profiles and epigenetic alterations in female SAMP8, a model of age-related cognitive decline. Front. Genet. 2018, 9, 596. [Google Scholar] [CrossRef]
- Griñan-Ferré, C.; Pérez-Cáceres, D.; Gutiérrez-Zetina, S.M.; Camins, A.; Palomera-Avalos, V.; Ortuño-Sahagún, D.; Rodrigo, M.T.; Pallàs, M. Environmental enrichment improves behavior, cognition, and brain functional markers in young senescence-accelerated prone mice (SAMP8). Mol. Neurobiol. 2016, 53, 2435–2450. [Google Scholar] [CrossRef]
- Griñan-Ferré, C.; Puigoriol-Illamola, D.; Palomera-Ávalos, V.; Pérez-Cáceres, D.; Companys-Alemany, J.; Camins, A.; Ortuño-Sahagún, D.; Rodrigo, M.T.; Pallàs, M. Environmental enrichment modified epigenetic mechanisms in SAMP8 mouse hippocampus by reducing oxidative stress and inflammaging and achieving neuroprotection. Front. Aging Neurosci. 2016, 8, 241. [Google Scholar] [CrossRef]
- Zhao, C.; Noble, J.M.; Marder, K.; Hartman, J.S.; Gu, Y.; Scarmeas, N. Dietary patterns, physical activity, sleep, and risk for dementia and cognitive decline. Curr. Nutr. Rep. 2018, 7, 335–345. [Google Scholar] [CrossRef]
- Gardener, S.L.; Rainey-Smith, S.R. The role of nutrition in cognitive function and brain ageing in the elderly. Curr. Nutr. Rep. 2018, 7, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.P.; Hoppe, J.B.; Saccomori, A.B.; Dos Santos, B.G.; Sagini, J.P.; Crestani, M.S.; August, P.M.; Hözer, R.M.; Grings, M.; Parmeggiani, B. Physical exercise during pregnancy prevents cognitive impairment induced by amyloid-β in adult offspring rats. Mol. Neurobiol. 2019, 56, 2022–2038. [Google Scholar] [CrossRef] [PubMed]
- Kusuyama, J.; Alves-Wagner, A.B.; Makarewicz, N.S.; Goodyear, L.J. Effects of maternal and paternal exercise on offspring metabolism. Nat. Metab. 2020, 2, 858–872. [Google Scholar] [CrossRef] [PubMed]
- Jha, S.K.; Jha, N.K.; Kumar, D.; Ambasta, R.K.; Kumar, P. Linking mitochondrial dysfunction, metabolic syndrome and stress signaling in Neurodegeneration. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 1132–1146. [Google Scholar] [CrossRef]
- De Boleti, A.P.A.; Almeida, J.A.; Migliolo, L. Impact of the metabolic syndrome on the evolution of neurodegenerative diseases. Neural Regen. Res. 2020, 16, 688. [Google Scholar]
- Rui, Y.; Cheng, J.; Qin, L.; Shan, C.; Chang, J.; Wang, G.; Wan, Z. Effects of vitamin D and resveratrol on metabolic associated markers in liver and adipose tissue from SAMP8 mice. Exp. Gerontol. 2017, 93, 16–28. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Huang, L.-T.; Sheen, J.-M.; Hou, C.-Y.; Yeh, Y.-T.; Chiang, C.-P.; Lin, I.-C.; Tiao, M.-M.; Tsai, C.-C.; Lin, Y.-J. Resveratrol treatment improves the altered metabolism and related dysbiosis of gut programed by prenatal high-fat diet and postnatal high-fat diet exposure. J. Nutr. Biochem. 2020, 75, 108260. [Google Scholar] [CrossRef]
- Palomera-Ávalos, V.; Griñán-Ferré, C.; Izquierdo, V.; Camins, A.; Sanfeliu, C.; Canudas, A.M.; Pallàs, M. Resveratrol modulates response against acute inflammatory stimuli in aged mouse brain. Exp. Gerontol. 2018, 102, 3–11. [Google Scholar] [CrossRef]
- Palomera-Avalos, V.; Griñán-Ferré, C.; Izquierdo, V.; Camins, A.; Sanfeliu, C.; Pallàs, M. Metabolic stress induces cognitive disturbances and inflammation in aged mice: Protective role of resveratrol. Rejuvenation Res. 2017, 20, 202–217. [Google Scholar] [CrossRef]
- Yu, H.; Sheen, J.; Tiao, M.; Tain, Y.; Chen, C.; Lin, I.; Lai, Y.; Tsai, C.; Lin, Y.; Tsai, C. Resveratrol Treatment Ameliorates Leptin Resistance and Adiposity Programed by the Combined Effect of Maternal and Post-Weaning High-Fat Diet. Mol. Nutr. Food Res. 2019, 63, 1801385. [Google Scholar] [CrossRef]
- Ford, D.; Ions, L.J.; Alatawi, F.; Wakeling, L.A. The potential role of epigenetic responses to diet in ageing. Proc. Nutr. Soc. 2011, 70, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Vanden Berghe, W. Epigenetic impact of dietary polyphenols in cancer chemoprevention: Lifelong remodeling of our epigenomes. Pharmacol. Res. 2012, 65, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Komorowska, J.; Wątroba, M.; Szukiewicz, D. Review of beneficial effects of resveratrol in neurodegenerative diseases such as Alzheimer’s disease. Adv. Med. Sci. 2020, 65, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Huang, N.; Xu, S.; Luo, Y.; Li, Y.; Jin, H.; Yu, C.; Shi, J.; Jin, F. Signaling mechanisms underlying inhibition of neuroinflammation by resveratrol in neurodegenerative diseases. J. Nutr. Biochem. 2020, 88, 108552. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lopez, O.; Milagro, F.I.; Riezu-Boj, J.I.; Martinez, J.A. Epigenetic signatures underlying inflammation: An interplay of nutrition, physical activity, metabolic diseases, and environmental factors for personalized nutrition. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2020, 1–21. [Google Scholar] [CrossRef]
- Zhang, N.; Li, Z.; Xu, K.; Wang, Y.; Wang, Z. Resveratrol protects against high-fat diet induced renal pathological damage and cell senescence by activating SIRT1. Biol. Pharm. Bull. 2016, 39, 1448–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, S.; Jiang, J.; Zhang, G.; Bu, Y.; Zhang, G.; Zhao, X. Resveratrol and caloric restriction prevent hepatic steatosis by regulating SIRT1-autophagy pathway and alleviating endoplasmic reticulum stress in high-fat diet-fed rats. PLoS ONE 2017, 12, e0183541. [Google Scholar]
- Zou, T.; Chen, D.; Yang, Q.; Wang, B.; Zhu, M.; Nathanielsz, P.W.; Du, M. Resveratrol supplementation of high-fat diet-fed pregnant mice promotes brown and beige adipocyte development and prevents obesity in male offspring. J. Physiol. 2017, 595, 1547–1562. [Google Scholar] [CrossRef]
- Tsai, T.-A.; Tsai, C.-K.; Huang, L.-T.; Sheen, J.-M.; Tiao, M.-M.; Tain, Y.-L.; Chen, C.-C.; Lin, I.; Lai, Y.-J.; Tsai, C.-C. Maternal Resveratrol Treatment Re-Programs and Maternal High-Fat Diet-Induced Retroperitoneal Adiposity in Male Offspring. Int. J. Environ. Res. Public Health 2020, 17, 2780. [Google Scholar] [CrossRef] [Green Version]
- Karpova, N.N.; Sales, A.J.; Joca, S.R. Epigenetic basis of neuronal and synaptic plasticity. Curr. Top. Med. Chem. 2017, 17, 771–793. [Google Scholar] [CrossRef]
- Pauwels, S.; Ghosh, M.; Duca, R.C.; Bekaert, B.; Freson, K.; Huybrechts, I.; Langie, S.A.S.; Koppen, G.; Devlieger, R.; Godderis, L. Maternal intake of methyl-group donors affects DNA methylation of metabolic genes in infants. Clin. Epigenetics 2017, 9, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velazquez, M.R.; Batistel, F.; Rodriguez, J.M.P.; Relling, A.E. Effects of maternal dietary omega-3 polyunsaturated fatty acids and methionine during late gestation on fetal growth, DNA methylation, and mRNA relative expression of genes associated with the inflammatory response, lipid metabolism and DNA methylation in placenta and offspring’s liver in sheep. J. Anim. Sci. Biotechnol. 2020, 11, 1–15. [Google Scholar]
- Kwan, S.T.C.; King, J.H.; Grenier, J.K.; Yan, J.; Jiang, X.; Roberson, M.S.; Caudill, M.A. Maternal choline supplementation during normal murine pregnancy alters the placental epigenome: Results of an exploratory study. Nutrients 2018, 10, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowacka-Woszuk, J.; Madeja, Z.E.; Chmurzynska, A. Prenatal caloric restriction alters lipid metabolism but not hepatic Fasn gene expression and methylation profiles in rats. BMC Genet. 2017, 18, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.; Liu, Z.; Xu, Y.; Liu, X.; Wang, D.; Li, F.; Wang, Y.; Bi, J. Abnormality of m6A mRNA methylation is involved in Alzheimer’s disease. Front. Neurosci. 2020, 14, 98. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Camats-Perna, J.; Medeiros, R.; Anggono, V.; Widagdo, J. Altered Expression of the m6A Methyltransferase METTL3 in Alzheimer’s Disease. Eneuro 2020. [Google Scholar] [CrossRef]
- Miyanohara, J.; Kakae, M.; Nagayasu, K.; Nakagawa, T.; Mori, Y.; Arai, K.; Shirakawa, H.; Kaneko, S. TRPM2 channel aggravates CNS inflammation and cognitive impairment via activation of microglia in chronic cerebral hypoperfusion. J. Neurosci. 2018, 38, 3520–3533. [Google Scholar] [CrossRef]
- Sharma, V.; Bryant, C.; Montero, M.; Creegan, M.; Slike, B.; Krebs, S.J.; Ratto-Kim, S.; Valcour, V.; Sithinamsuwan, P.; Chalermchai, T. Monocyte and CD4+ T-cell antiviral and innate responses associated with HIV-1 inflammation and cognitive impairment. Aids 2020, 34, 1289–1301. [Google Scholar] [CrossRef]
- Pan, W.; Yu, H.; Huang, S.; Zhu, P. Resveratrol protects against TNF-α-induced injury in human umbilical endothelial cells through promoting sirtuin-1-induced repression of NF-KB and p38 MAPK. PLoS ONE 2016, 11, e0147034. [Google Scholar] [CrossRef] [Green Version]
- Devi, S.A.; Chamoli, A. Polyphenols as an Effective Therapeutic Intervention Against Cognitive Decline During Normal and Pathological Brain Aging. In Reviews on New Drug Targets in Age-Related Disorders; Springer: Cham, Switzerland, 2020; pp. 159–174. ISBN 978-3-030-42667-5. [Google Scholar]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef]
- Lu, H.; Wang, B.; Cui, N.; Zhang, Y. Artesunate suppresses oxidative and inflammatory processes by activating Nrf2 and ROS-dependent p38 MAPK and protects against cerebral ischemia-reperfusion injury. Mol. Med. Rep. 2018, 17, 6639–6646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, Y.; Han, Q.; Xu, Y.; Hu, G.; Xing, H. The inflammatory injury of heart caused by ammonia is realized by oxidative stress and abnormal energy metabolism activating inflammatory pathway. Sci. Total Environ. 2020, 742, 140532. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Liu, Y.; Ren, G.; Yin, L.; Liang, X.; Geng, T.; Dang, H.; An, R. Resveratrol limits diabetes-associated cognitive decline in rats by preventing oxidative stress and inflammation and modulating hippocampal structural synaptic plasticity. Brain Res. 2016, 1650, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Oliveros, A.; Jang, M.-H. Dysfunctional mitochondrial bioenergetics and synaptic degeneration in Alzheimer disease. Int. Neurourol. J. 2019, 23, S5. [Google Scholar] [CrossRef] [Green Version]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-derived neurotrophic factor: A key molecule for memory in the healthy and the pathological brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef]
- Heisz, J.J.; Clark, I.B.; Bonin, K.; Paolucci, E.M.; Michalski, B.; Becker, S.; Fahnestock, M. The effects of physical exercise and cognitive training on memory and neurotrophic factors. J. Cogn. Neurosci. 2017, 29, 1895–1907. [Google Scholar] [CrossRef]
- Canu, N.; Amadoro, G.; Triaca, V.; Latina, V.; Sposato, V.; Corsetti, V.; Severini, C.; Ciotti, M.T.; Calissano, P. The intersection of NGF/TrkA signaling and amyloid precursor protein processing in Alzheimer’s disease neuropathology. Int. J. Mol. Sci. 2017, 18, 1319. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xie, Y.; Zhang, T.; Bo, S.; Bai, X.; Liu, H.; Li, T.; Liu, S.; Zhou, Y.; Cong, X. Resveratrol reverses chronic restraint stress-induced depression-like behaviour: Involvement of BDNF level, ERK phosphorylation and expression of Bcl-2 and Bax in rats. Brain Res. Bull. 2016, 125, 134–143. [Google Scholar] [CrossRef]
- Rosa, P.M.; Martins, L.A.M.; Souza, D.O.; Quincozes-Santos, A. Glioprotective effect of resveratrol: An emerging therapeutic role for oligodendroglial cells. Mol. Neurobiol. 2018, 55, 2967–2978. [Google Scholar] [CrossRef]
- Wang, X.; Ma, S.; Yang, B.; Huang, T.; Meng, N.; Xu, L.; Xing, Q.; Zhang, Y.; Zhang, K.; Li, Q. Resveratrol promotes hUC-MSCs engraftment and neural repair in a mouse model of Alzheimer’s disease. Behav. Brain Res. 2018, 339, 297–304. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Izquierdo, V.; Palomera-Ávalos, V.; Pallàs, M.; Griñán-Ferré, C. Resveratrol Supplementation Attenuates Cognitive and Molecular Alterations under Maternal High-Fat Diet Intake: Epigenetic Inheritance over Generations. Int. J. Mol. Sci. 2021, 22, 1453. https://doi.org/10.3390/ijms22031453
Izquierdo V, Palomera-Ávalos V, Pallàs M, Griñán-Ferré C. Resveratrol Supplementation Attenuates Cognitive and Molecular Alterations under Maternal High-Fat Diet Intake: Epigenetic Inheritance over Generations. International Journal of Molecular Sciences. 2021; 22(3):1453. https://doi.org/10.3390/ijms22031453
Chicago/Turabian StyleIzquierdo, Vanesa, Verónica Palomera-Ávalos, Mercè Pallàs, and Christian Griñán-Ferré. 2021. "Resveratrol Supplementation Attenuates Cognitive and Molecular Alterations under Maternal High-Fat Diet Intake: Epigenetic Inheritance over Generations" International Journal of Molecular Sciences 22, no. 3: 1453. https://doi.org/10.3390/ijms22031453
APA StyleIzquierdo, V., Palomera-Ávalos, V., Pallàs, M., & Griñán-Ferré, C. (2021). Resveratrol Supplementation Attenuates Cognitive and Molecular Alterations under Maternal High-Fat Diet Intake: Epigenetic Inheritance over Generations. International Journal of Molecular Sciences, 22(3), 1453. https://doi.org/10.3390/ijms22031453